
Role of Long Noncoding RNAs ZlMSTRG.11348 and UeMSTRG.02678 in Temperature-Dependent Culm Swelling in Zizania latifolia

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Plant Culm Morphology
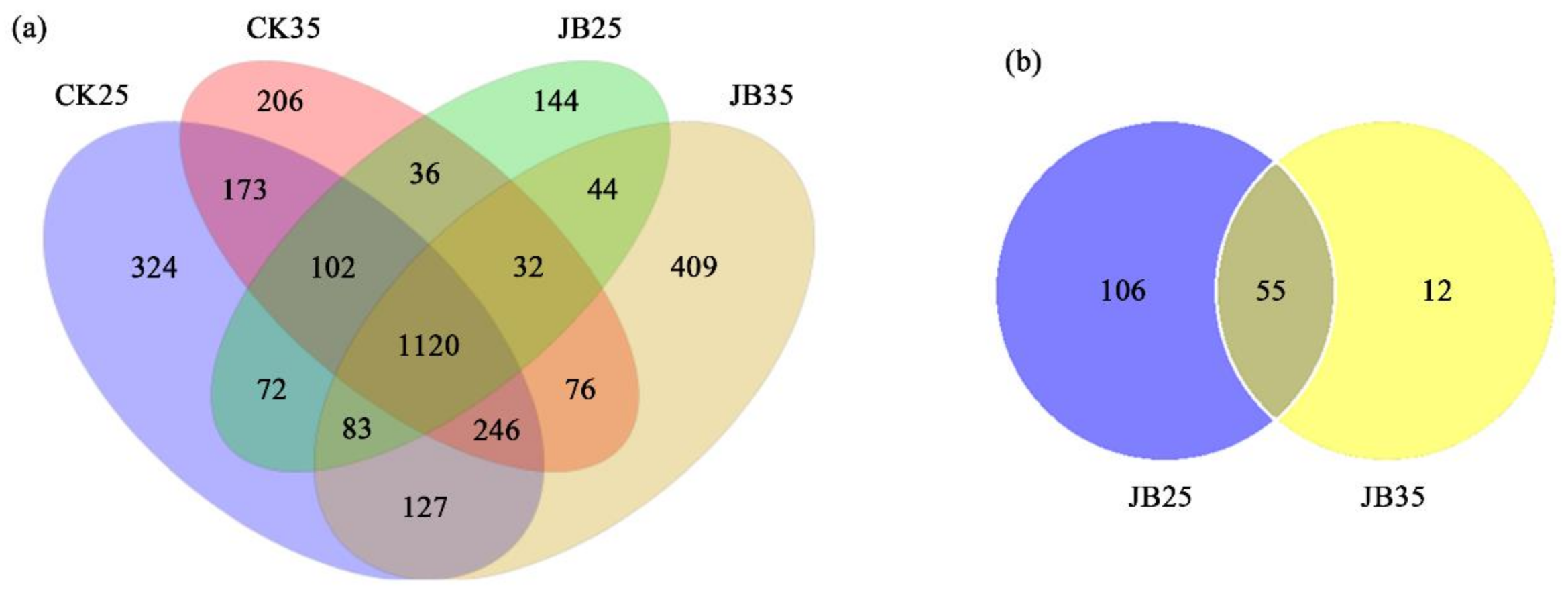
2.2. Identification and Characterisation of lncRNAs in Z. latifolia and U. esculenta
2.3. lncRNA and mRNA Expression in Z. latifolia and U. esculenta
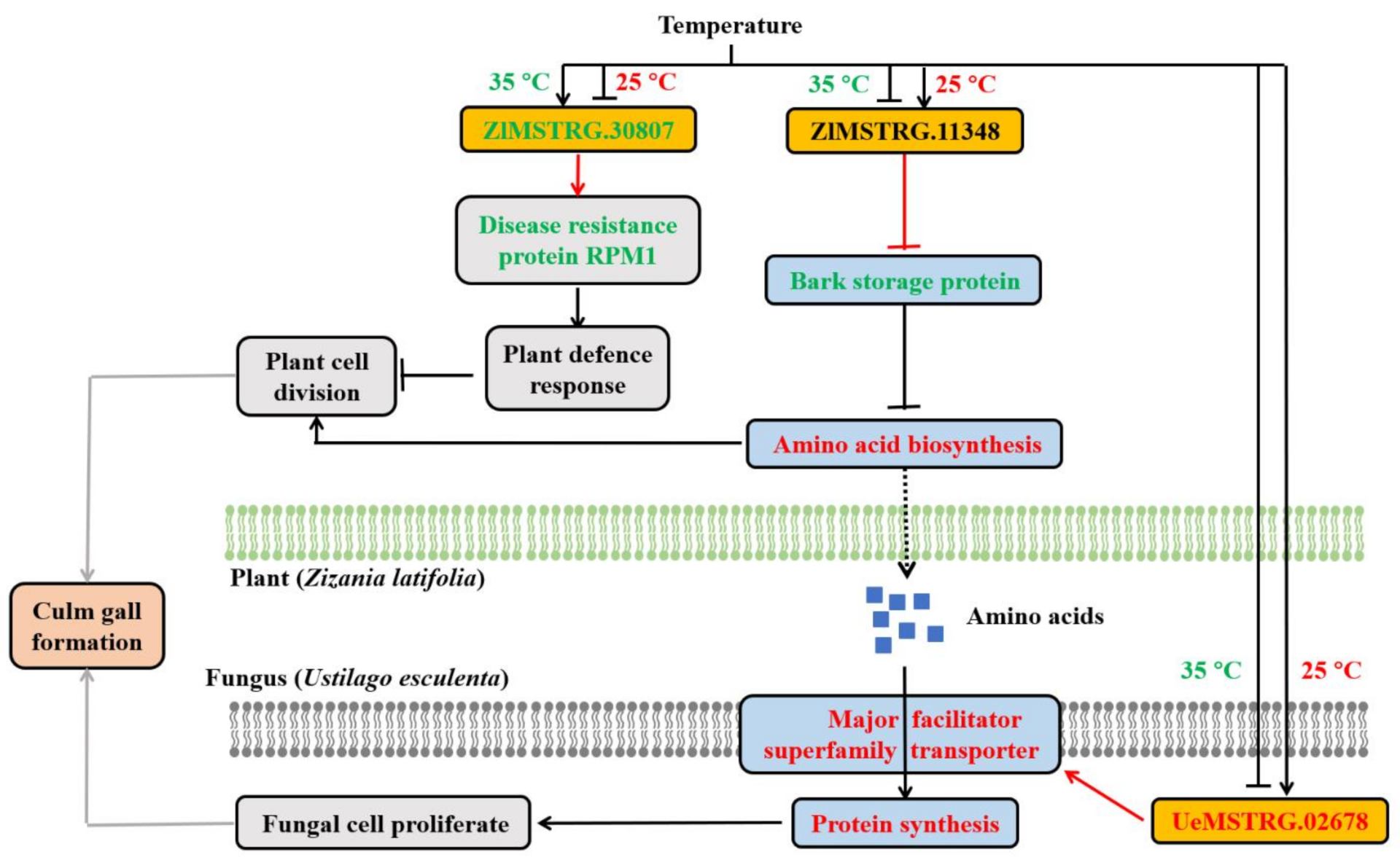
2.4. Functional Analysis of lncRNAs
2.5. Plant Defence Response
2.6. Plant Hormones
2.7. Amino Acid Metabolism
3. Discussion
3.1. Culm Gall Formation Is Regulated by Temperature
3.2. Temperature Alters lncRNA-Mediated Genes Related to Plant Defence Response and Hormone
3.3. Expression of Amino Acid Metabolism and Sugar Metabolism Genes Influences Gall Formation
4. Materials and Methods
4.1. Plant Growth Conditions and Sampling
4.2. Fungal Strains and Culture Conditions
4.3. RNA Extraction, Library Construction, and Sequencing
4.4. Transcriptome Assembly and lncRNA Identification
4.5. Differential Expression of lncRNAs and mRNAs
4.6. Prediction and Functional Analysis of lncRNA Targets
4.7. Quantitative Reverse-Transcription PCR
4.8. Bioinformatics Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chan, Y.S.; Thrower, L.B. The host-parasite relationship between Zizania-caduciflora Turcz and Ustilago-esculenta P Henn.1. Structure and development of the host-parasite combination. New Phytol. 1980, 85, 201–207. [Google Scholar] [CrossRef]
- Yan, N.; Wang, X.Q.; Xu, X.F.; Guo, D.P.; Wang, Z.D.; Zhang, J.Z.; Hyde, K.D.; Liu, H.L. Plant growth and photosynthetic performance of Zizania latifolia are altered by endophytic Ustilago esculenta infection. Physiol. Mol. Plant Pathol. 2013, 83, 75–83. [Google Scholar] [CrossRef]
- Wang, Z.D.; Yan, N.; Wang, Z.H.; Zhang, X.H.; Zhang, J.Z.; Xue, H.M.; Wang, L.X.; Zhan, Q.; Xu, Y.P.; Guo, D.P. RNA-seq analysis provides insight into reprogramming of culm development in Zizania latifolia induced by Ustilago esculenta. Plant Mol. Biol. 2017, 95, 533–547. [Google Scholar] [CrossRef] [PubMed]
- Lanver, D.; Muller, A.N.; Happel, P.; Schweizer, G.; Haas, F.B.; Franitza, M.; Pellegrin, C.; Reissmann, S.; Altmuller, J.; Rensing, S.A.; et al. The biotrophic development of Ustilago maydis studied by RNA-Seq analysis. Plant Cell 2018, 30, 300–323. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.P.; Shi, M.; Huang, J.Z.; Guo, D.P.; Hu, Z.L. Effects of different altitudes on growth and culm-swelling of Zizania caduciflora (Turca.). China Veg. 2013, 12, 88–93. [Google Scholar]
- Yan, N.; Xu, X.F.; Wang, Z.D.; Huang, J.Z.; Guo, D.P. Interactive effects of temperature and light intensity on photosynthesis and antioxidant enzyme activity in Zizania latifolia Turcz. plants. Photosynthetica 2013, 51, 127–138. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Chu, F.Q.; Guo, D.P.; Hyde, K.D.; Xie, G.L. Cytology and ultrastructure of interactions between Ustilago esculenta and Zizania latifolia. Mycol. Prog. 2012, 11, 499–508. [Google Scholar] [CrossRef]
- Velasquez, A.C.; Castroverde, C.D.M.; He, S.Y. Plant-pathogen warfare under changing climate conditions. Curr. Biol. 2018, 28, R619–R634. [Google Scholar] [CrossRef] [Green Version]
- Menna, A.; Nguyen, D.; Guttman, D.S.; Desveaux, D. Elevated temperature differentially influences effector-triggered immunity outputs in Arabidopsis. Front. Plant Sci. 2015, 6, 995:1–995:7. [Google Scholar] [CrossRef] [Green Version]
- Huot, B.; Castroverde, C.D.M.; Velasquez, A.C.; Hubbard, E.; Pulman, J.A.; Yao, J.; Childs, K.L.; Tsuda, K.; Montgomery, B.L.; He, S.Y. Dual impact of elevated temperature on plant defence and bacterial virulence in Arabidopsis. Nat. Commun. 2017, 8, 1808:1–1808:12. [Google Scholar] [CrossRef]
- Kim, Y.S.; An, C.; Park, S.; Gilmour, S.J.; Wang, L.; Renna, L.; Brandizzi, F.; Grumet, R.; Thomashow, M.F. CAMTA-mediated regulation of salicylic acid immunity pathway genes in Arabidopsis exposed to low temperature and pathogen infection. Plant Cell 2017, 29, 2465–2477. [Google Scholar] [CrossRef] [Green Version]
- Nadal, M.; Garcia-Pedrajas, M.D.; Gold, S.E. Dimorphism in fungal plant pathogens. FEMS Microbiol. Lett. 2008, 284, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.H.; Chen, Y.; Ma, C.J.; Liu, J.; Liguo, C.; Ma, A.M. Effects of environmental factors on dimorphic transition of the jelly mushroom Tremella fuciformis. Cryptogam. Mycol. 2011, 32, 421–428. [Google Scholar] [CrossRef]
- Zhu, L.B.; Wang, Y.; Zhang, Z.B.; Yang, H.L.; Yan, R.M.; Zhu, D. Influence of environmental and nutritional conditions on yeast–mycelial dimorphic transition in Trichosporon cutaneum. Biotechnol. Biotechol. Equip. 2017, 31, 516–526. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.; Song, Y.N.; Deng, W.Y.; Gordon, M.P.; Nester, E.W. The regulatory VirA protein of Agrobacterium tumefaciens does not function at elevated temperatures. J. Bacteriol. 1993, 21, 6830–6835. [Google Scholar] [CrossRef] [Green Version]
- Baron, C.; Domke, N.; Beinhofer, M.; Hapfelmeier, S. Elevated temperature differentially affects virulence, VirB protein accumulation, and T-pilus formation in different Agrobacterium tumefaciens and Agrobacterium vitis strains. J. Bacteriol. 2001, 183, 6852–6861. [Google Scholar] [CrossRef] [Green Version]
- Pratelli, R.; Pilot, G. Regulation of amino acid metabolic enzymes and transporters in plants. J. Exp. Bot. 2014, 65, 5535–5556. [Google Scholar] [CrossRef]
- Muthusamy, T.; Cordes, T.; Handzlik, M.K.; You, L.; Lim, E.W.; Gengatharan, J.; Pinto, A.F.M.; Badur, M.G.; Kolar, M.J.; Wallace, M.; et al. Serine restriction alters sphingolipid diversity to constrain tumour growth. Nature 2020, 586, 790–795. [Google Scholar] [CrossRef]
- Possemato, R.; Marks, K.M.; Shaul, Y.D.; Pacold, M.E.; Kim, D.; Birsoy, K.; Sethumadhavan, S.; Woo, H.K.; Jang, H.G.; Jha, A.K.; et al. Functional genomics reveal that the serine synthesis pathway is essential in breast cancer. Nature 2011, 476, 346–350. [Google Scholar] [CrossRef] [Green Version]
- Knott, S.R.V.; Wagenblast, E.; Khan, S.; Kim, S.Y.; Soto, M.; Wagner, M.; Turgeon, M.O.; Fish, L.; Erard, N.; Gable, A.L.; et al. Asparagine bioavailability governs metastasis in a model of breast cancer. Nature 2018, 554, 378–381. [Google Scholar] [CrossRef]
- Leone, R.D.; Zhao, L.; Englert, J.M.; Sun, I.M.; Oh, M.H.; Sun, I.H.; Arwood, M.L.; Bettencourt, I.A.; Patel, C.H.; Wen, J.; et al. Glutamine blockade induces divergent metabolic programs to overcome tumor immune evasion. Science 2019, 366, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- McCann, M.P.; Snetselaar, K.M. A genome-based analysis of amino acid metabolism in the biotrophic plant pathogen Ustilago maydis. Fungal Genet. Biol. 2008, 45, S77–S87. [Google Scholar] [CrossRef] [PubMed]
- Saint-Macary, M.E.; Barbisan, C.; Gagey, M.J.; Frelin, O.; Beffa, R.; Lebrun, M.H.; Droux, M. Methionine biosynthesis is essential for infection in the rice blast fungus Magnaporthe oryzae. PLoS ONE 2015, 10, e0111108:1–e0111108:22. [Google Scholar] [CrossRef]
- Kerppola, T.K.; Kahn, M.L. Symbiotic phenotypes of auxotrophic mutants of Rhizobium meliloti 104A14. J. Gen. Microbiol. 1988, 134, 913–919. [Google Scholar]
- Mulley, G.; White, J.P.; Karunakaran, R.; Prell, J.; Bourdes, A.; Bunnewell, S.; Hill, L.; Poole, P.S. Mutation of GOGAT prevents pea bacteroid formation and N2 fixation by globally downregulating transport of organic nitrogen sources. Mol. Microbiol. 2011, 80, 149–167. [Google Scholar] [CrossRef]
- Resendis-Antonio, O.; Hernandez, M.; Salazar, E.; Contreras, S.; Batallar, G.M.; Mora, Y.; Encarnacion, S. Systems biology of bacterial nitrogen fixation: High-throughput technology and its integrative description with constraint-based modeling. BMC Syst. Biol. 2011, 5, 120:1–120:15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, M.F. Key roles of microsymbiont amino acid metabolism in rhizobia-legume interactions. Crit. Rev. Microbiol. 2014, 41, 411–451. [Google Scholar] [CrossRef]
- Salmeron-Santiago, K.G.; Pardo, J.P.; Flores-Herrera, O.; Mendoza-Hernandez, G.; Miranda-Arango, M.; Guerra-Sanchez, G. Response to osmotic stress and temperature of the fungus Ustilago maydis. Arch. Microbiol. 2011, 193, 701–709. [Google Scholar] [CrossRef]
- Chekanova, J.A. Long non-coding RNAs and their functions in plants. Curr. Opin. Plant Biol. 2015, 27, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Jin, Z.X.; Gao, S.W.; Ma, W.Y.; Lyu, X.N.; Cao, X.L.; Yao, Y.X. Identification and functional prediction of salt stress-related long noncoding RNAs in grapevine roots. Environ. Exp. Bot. 2020, 179, 104215:1–104215:11. [Google Scholar] [CrossRef]
- Zhu, Q.H.; Stephen, S.; Taylor, J.; Helliwell, C.A.; Wang, M.B. Long noncoding RNAs responsive to Fusarium oxysporum infection in Arabidopsis thaliana. New Phytol. 2014, 201, 574–584. [Google Scholar] [CrossRef]
- Seo, J.S.; Sun, H.X.; Park, B.S.; Huang, C.H.; Yeh, S.D.; Jung, C.; Chua, N.H. ELF18-INDUCED LONG-NONCODING RNA associates with mediator to enhance expression of innate immune response genes in Arabidopsis. Plant Cell 2017, 29, 1024–1038. [Google Scholar] [CrossRef] [Green Version]
- Peter, K.; Ryan, A.; Maxim, I.; Sebastian, M. Transcriptional read-through of the long noncoding RNA SVALKA governs plant cold acclimation. Nat. Commun. 2018, 9, 4561:1–4561:11. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, G.; Singh, A.; Verma, D.; Sharma, S.; Singh, K. Genome-wide investigation of regulatory roles of lncRNAs in response to heat and drought stress in Brassica juncea (Indian mustard). Environ. Exp. Bot. 2020, 171, 103922. [Google Scholar] [CrossRef]
- Wu, J.; Zhao, W.; Wang, Z.; Xiang, X.; Zhang, S.; Liu, L. Long non-coding RNA SNHG20 promotes the tumorigenesis of oral squamous cell carcinoma via targeting miR-197/LIN28 axis. J. Cell. Mol. Med. 2019, 23, 680–688. [Google Scholar] [CrossRef]
- Chen, D.L.; Ju, H.Q.; Lu, Y.X.; Chen, L.Z.; Zeng, Z.L.; Zhang, D.S.; Luo, H.Y.; Wang, F.; Qiu, M.Z.; Wang, D.S.; et al. Long non-coding RNA XIST regulates gastric cancer progression by acting as a molecular sponge of miR-101 to modulate EZH2 expression. J. Exp. Clin. Canc. Res. 2016, 35, 142:1–142:13. [Google Scholar] [CrossRef] [Green Version]
- Yan, N.; Wang, X.Q.; Wang, Z.D.; Zhang, Y.L.; Xue, H.M.; Guo, D.P. Antioxidative system and chlorophyll fluorescence of Zizania latifolia Turcz. plants are affected by Ustilago esculenta infection. Acta Ecol. Sin. 2013, 33, 1584–1593. [Google Scholar]
- Wang, Z.H.; Yan, N.; Luo, X.; Guo, S.S.; Xue, S.Q.; Liu, J.Q.; Zhang, J.Z.; Guo, D.P. Gene expression in the smut fungus Ustilago esculenta governs swollen gall metamorphosis in Zizania latifolia. Microb. Pathogenesis. 2020, 143, 104107:1–104107:11. [Google Scholar] [CrossRef]
- Morrison, E.N.; Emery, R.J.; Saville, B.J. Phytohormone involvement in the Ustilago maydis-Zea mays pathosystem: Relationships between abscisic acid and cytokinin levels and strain virulence in infected cob tissue. PLoS ONE 2015, 10, e0130945:1–e0130945:23. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Gao, X.; Feng, B.; Sheen, J.; Shan, L.; He, P. Plant immune response to pathogens differs with changing temperatures. Nat. Commun. 2013, 4, 2530:1–2530:9. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Zhou, D.; Du, Q.; Chen, J.; Wang, Q.; Zhang, D. Identification and allelic dissection uncover roles of lncRNAs in secondary growth of Populus tomentosa. DNA Res. 2017, 24, 473–486. [Google Scholar] [CrossRef] [Green Version]
- Lozano-Duran, R.; Zipfel, C. Trade-off between growth and immunity: Role of brassinosteroids. Trends Plant Sci. 2015, 20, 12–19. [Google Scholar] [CrossRef]
- Gao, Z.; Chung, E.H.; Eitas, T.K.; Dangl, J.L. Plant intracellular innate immune receptor resistance to Pseudomonas syringae pv. maculicola 1 (RPM1) is activated at, and functions on, the plasma membrane. Proc. Natl. Acad. Sci. USA 2011, 108, 7619–7624. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Feng, B.M.; He, P.; Shan, L.B. From chaos to harmony: Responses and signaling upon microbial pattern recognition. Annu. Rev. Phytopathol. 2017, 55, 109–137. [Google Scholar] [CrossRef]
- Guo, D.P.; Jiang, Y.T.; Zeng, G.W.; Shah, G.A. Stem swelling of stem mustard, as affected by temperature and growth regulators. Sci. Hortic. 1994, 60, 153–160. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Ren, J.S.; Clifton, I.J.; Schofield, C.J. Crystal structure and mechanistic implications of 1-aminocyclopropane-1-carboxylic acid oxidase-the ethylene-forming enzyme. Chem. Biol. 2004, 11, 1383–1394. [Google Scholar] [CrossRef] [Green Version]
- Pre, M.; Atallah, M.; Champion, A.; De Vos, M.; Pieterse, C.M.; Memelink, J. The AP2/ERF domain transcription factor ORA59 integrates jasmonic acid and ethylene signals in plant defense. Plant Physiol. 2008, 147, 1347–1357. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.J. Signaling crosstalk between salicylic acid and ethylene/jasmonate in plant defense: Do we understand what they are whispering? Int. J. Mol. Sci. 2019, 20, 671. [Google Scholar] [CrossRef] [Green Version]
- Palmer, D.A.; Bender, C.L. Effects of environmental and nutritional factors on production of the polyketide phytotoxin coronatine by Pseudomonas syringae pv. glycinea. Appl. Environ. Microbiol. 1993, 59, 1619–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Dijk, K.; Fouts, D.E.; Rehm, A.H.; Hill, A.R.; Collmer, A.; Alfano, J.R. The Avr (effector) proteins HrmA (HopPsyA) and AvrPto are secreted in culture from Pseudomonas syringae pathovars via the Hrp (type III) protein secretion system in a temperature- and pH-sensitive manner. J. Bacteriol. 1999, 181, 4790–4797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weingart, H.; Stubner, S.; Schenk, A.; Ullrich, M.S. Impact of temperature on in planta expression of genes involved in synthesis of the Pseudomonas syringae phytotoxin coronatine. Mol. Plant Microbe Interact. 2004, 17, 1095–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemetsberger, C.; Herrberger, C.; Zechmann, B.; Hillmer, M.; Doehlemann, G. The Ustilago maydis effector Pep1 suppresses plant immunity by inhibition of host peroxidase activity. PLoS Pathog. 2012, 8, e1002684:1–e1002684:14. [Google Scholar] [CrossRef] [Green Version]
- Froldi, F.; Pachnis, P.; Szuperak, M.; Costas, O.; Fernando, T.; Gould, A.P.; Cheng, L.Y. Histidine is selectively required for the growth of Myc-dependent dedifferentiation tumours in the Drosophila CNS. EMBO J. 2019, 38, e99895:1–e99895:12. [Google Scholar] [CrossRef]
- Sanders, H.K.; Becker, G.E.; Nason, A. Glycine-cytochrome c reductase from Nitrobacter agilis. J. Biol. Chem. 1972, 247, 2015–2025. [Google Scholar] [CrossRef]
- Lancien, M.; Martin, M.; Hsieh, M.H.; Leustek, T.; Goodman, H.; Coruzzi, G.M. Arabidopsis glt1-T mutant defines a role for NADH-GOGAT in the non-photorespiratory ammonium assimilatory pathway. Plant J. 2002, 29, 347–358. [Google Scholar] [CrossRef]
- Nunes, J.E.; Ducati, R.G.; Breda, A.; Rosado, L.A.; de Souza, B.M.; Palma, M.S.; Santos, D.S.; Basso, L.A. Molecular, kinetic, thermodynamic, and structural analyses of Mycobacterium tuberculosis hisD-encoded metal-dependent dimeric histidinol dehydrogenase (EC 1.1.1.23). Arch. Biochem. Biophys. 2011, 512, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Wildhagen, H.; Durr, J.; Ehlting, B.; Rennenberg, H. Seasonal nitrogen cycling in the bark of field-grown grey poplar is correlated with meteorological factors and gene expression of bark storage proteins. Tree Physiol. 2010, 30, 1096–1110. [Google Scholar] [CrossRef] [Green Version]
- Lizarraga-Guerra, R.; Lopez, M.G. Content of free amino acids in huitlacoche (Ustilago maydis). J. Agric. Food Chem. 1996, 44, 2556–2559. [Google Scholar] [CrossRef]
- Ye, Z.H.; Pan, Y.; Zhang, Y.F.; Cui, H.F.; Jin, G.L.; McHardy, A.C.; Fan, L.J.; Yu, X.P. Comparative whole-genome analysis reveals artificial selection effects on Ustilago esculenta genome. DNA Res. 2017, 24, 635–648. [Google Scholar] [CrossRef] [Green Version]
- Sekito, T.; Fujiki, Y.; Ohsumi, Y.; Kakinuma, Y. Novel families of vacuolar amino acid transporters. IUBMB Life 2008, 60, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Walerowski, P.; Gundel, A.; Yahaya, N.; Truman, W.; Sobczak, M.; Olszak, M.; Rolfe, S.; Borisjuk, L.; Malinowski, R. Clubroot disease stimulates early steps of phloem differentiation and recruits SWEET sucrose transporters within developing galls. Plant Cell 2018, 30, 3058–3073. [Google Scholar] [CrossRef] [Green Version]
- Koyama, Y.; Yao, I.; Akimoto, S.I. Aphid galls accumulate high concentrations of amino acids a support for the nutrition hypothesis for gall formation. Entomol. Exp. Appl. 2004, 113, 35–44. [Google Scholar] [CrossRef]
- Li, R.; Yu, C.; Li, Y.; Lam, T.W.; Yiu, S.M.; Kristiansen, K.; Wang, J. SOAP2: An improved ultrafast tool for short read alignment. Bioinformatics 2009, 25, 1966–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, L.; Zhang, Y.; Ye, Z.Q.; Liu, X.Q.; Zhao, S.Q.; Wei, L.; Gao, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 2007, 35, W345–W349. [Google Scholar] [CrossRef]
- Sun, L.; Luo, H.; Bu, D.; Zhao, G.; Yu, K.; Zhang, C.; Liu, Y.; Chen, R.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166:1–e166:8. [Google Scholar] [CrossRef]
- Mistry, J.; Bateman, A.; Finn, R.D. Predicting active site residue annotations in the Pfam database. BMC Bioinform. 2007, 8, 298:1–298:14. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550:1–550:21. [Google Scholar] [CrossRef] [Green Version]
- Tafer, H.; Hofacker, I.L. RNAplex: A fast tool for RNA-RNA interaction search. Bioinformatics 2008, 24, 2657–2663. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Arezi, B.; Xing, W.; Sorge, J.A.; Hogrefe, H.H. Amplification efficiency of thermostable DNA polymerases. Anal. Biochem. 2003, 321, 226–235. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| lncRNA ID | Log2FC (JB25/JB35) | Cis-Target Gene ID | Log2FC (JB25/JB35) | Annotation |
|---|---|---|---|---|
| UeMSTRG.02678 | 17.44935825 | Ue.05653 | 11.40205275 | Major facilitator superfamily transporter |
| UeMSTRG.00837 | 4.255446947 | Ue.00835 | 2.530165481 | Uncharacterised protein SPSC_00409 |
| UeMSTRG.03069 | 2.581856123 | Ue.06235 | 1.608946133 | Hypothetical protein PSEUBRA_SCAF8g02247 |
| UeMSTRG.00868 | 1.521002017 | Ue.00907 | 1.211732405 | Probable glutathione-dependent formaldehyde dehydrogenase |
| Gene ID | Log2FC (JB25/JB35) | Log2FC (CK25/CK35) | Annotation | |
|---|---|---|---|---|
| Ethylene biosynthesis | Zl.12708 | NA | 3.080401507 | 1-aminocyclopropane-1-carboxylate oxidase 1 isoform X1 |
| Zl.09313 | NA | −1.126827996 | 1-aminocyclopropane-1-carboxylate oxidase | |
| Zl.07467 | −6.974635202 | −4.772315741 | Putative 1-aminocyclopropane-1-carboxylate oxidase | |
| Zl.10315 | −4.669026766 | −1.083063024 | 1-aminocyclopropane-1-carboxylate synthase | |
| Zl.09320 | −2.994981925 | −1.715749108 | 1-aminocyclopropane-1-carboxylate oxidase (ACC oxidase) | |
| Zl.04853 | −1.801681538 | NA | S-adenosylmethionine synthetase | |
| Cytokinin metabolism | Zl.15021 | 7.055282436 | NA | Adenylate isopentenyltransferase |
| Zl.03496 | 4.120206262 | NA | PREDICTED: cis-zeatin O-glucosyltransferase 1-like | |
| Zl.19719 | 4.101538026 | NA | Cytokinin dehydrogenase 8-like | |
| Zl.20687 | 2.633461018 | NA | Isopentenyltransferase 8 | |
| Zl.22357 | 1.429568818 | 2.365983275 | Eukaryotic tRNA isopentenyltransferase | |
| Zl.09144 | NA | 3.54689446 | Cytokinin dehydrogenase 9 | |
| Zl.19009 | NA | 2.130198723 | PREDICTED: protein G1-like4 | |
| Zl.04106 | NA | 1.230663683 | Adenylate isopentenyltransferase | |
| Zl.05044 | −4.667587558 | 1.315840765 | Putative cytokinin dehydrogenase | |
| Zl.12768 | −4.113062664 | 1.16901907 | PREDICTED: cytokinin dehydrogenase 4-like | |
| Zl.02021 | −3.741466986 | NA | Putative cis-zeatin O-glucosyltransferase | |
| Zl.14070 | −1.29936594 | NA | PREDICTED: adenylate isopentenyltransferase-like |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.-H.; Yan, N.; Luo, X.; Guo, S.-S.; Xue, S.-Q.; Liu, J.-Q.; Zhang, S.-S.; Zheng, L.-W.; Zhang, J.-Z.; Guo, D.-P. Role of Long Noncoding RNAs ZlMSTRG.11348 and UeMSTRG.02678 in Temperature-Dependent Culm Swelling in Zizania latifolia. Int. J. Mol. Sci. 2021, 22, 6020. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22116020
Wang Z-H, Yan N, Luo X, Guo S-S, Xue S-Q, Liu J-Q, Zhang S-S, Zheng L-W, Zhang J-Z, Guo D-P. Role of Long Noncoding RNAs ZlMSTRG.11348 and UeMSTRG.02678 in Temperature-Dependent Culm Swelling in Zizania latifolia. International Journal of Molecular Sciences. 2021; 22(11):6020. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22116020
Chicago/Turabian StyleWang, Zheng-Hong, Ning Yan, Xi Luo, Sai-Sai Guo, Shu-Qin Xue, Jiang-Qiong Liu, Shen-Shen Zhang, Li-Wen Zheng, Jing-Ze Zhang, and De-Ping Guo. 2021. "Role of Long Noncoding RNAs ZlMSTRG.11348 and UeMSTRG.02678 in Temperature-Dependent Culm Swelling in Zizania latifolia" International Journal of Molecular Sciences 22, no. 11: 6020. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22116020