
Identification of Tumor Suppressive Genes Regulated by miR-31-5p and miR-31-3p in Head and Neck Squamous Cell Carcinoma

 , ,
, ,
Abstract
:1. Introduction
2. Results
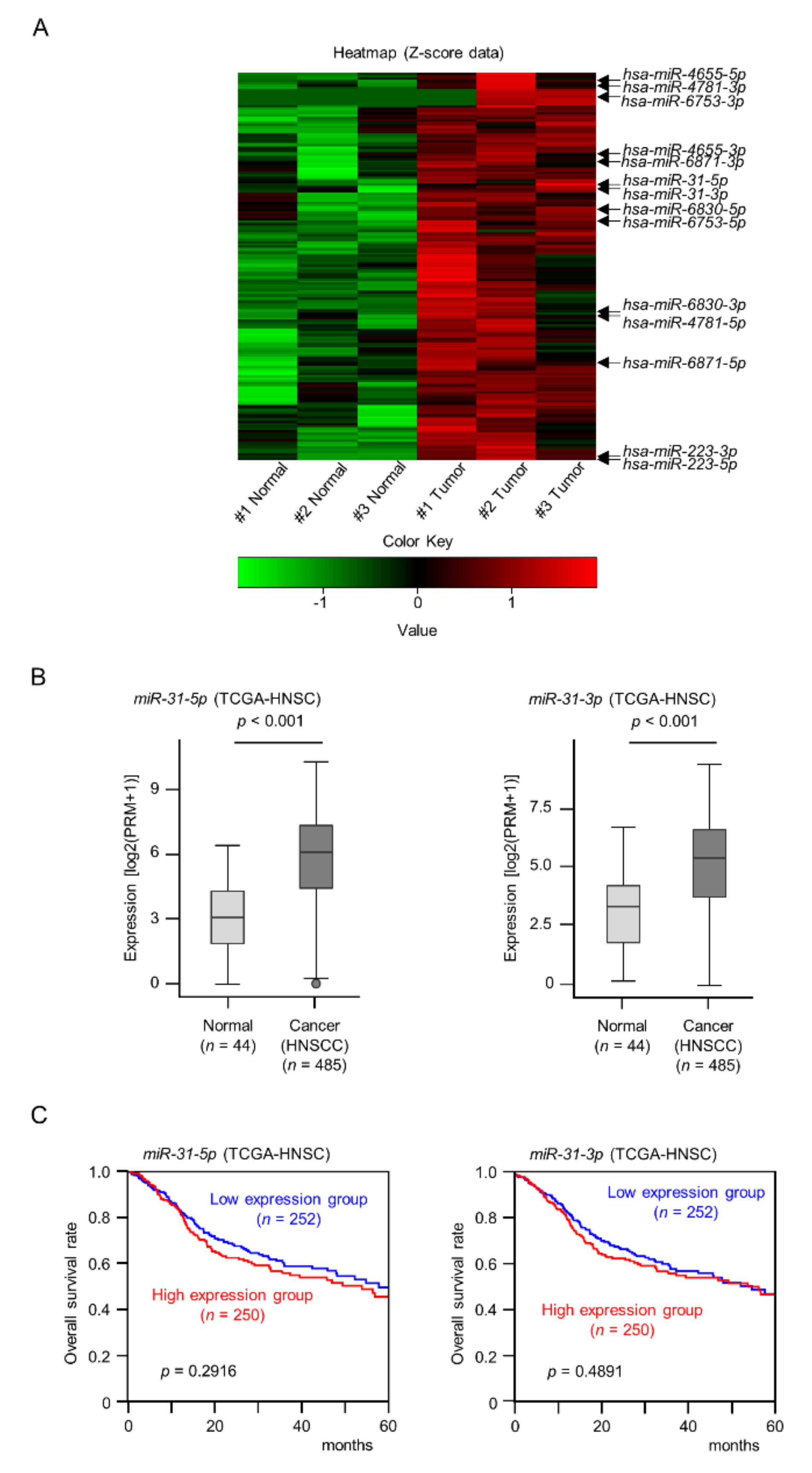
2.1. Identification of the miRNA Expression Signature of HNSCC by RNA Sequencing
2.2. Expression Levels and Clinical Significance of miR-31-5p and miR-31-3p in HNSCC
2.3. Effects of Inhibition of miR-31-5p and miR-31-3p Expression on the Proliferation, Migration, and Invasion of HNSCC Cells
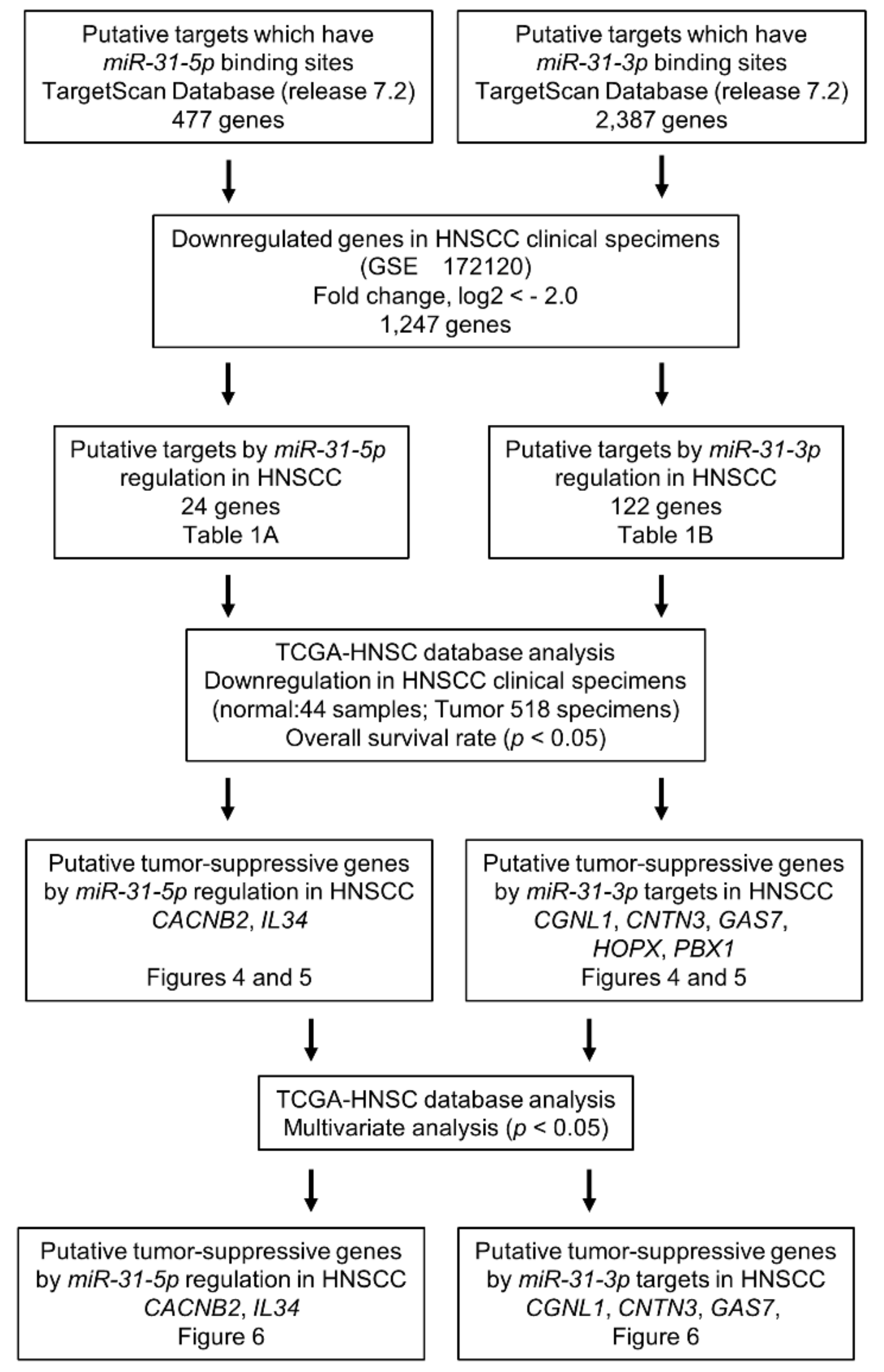
2.4. Screening of miR-31-5p and miR-31-3p Targets in HNSCC Cells
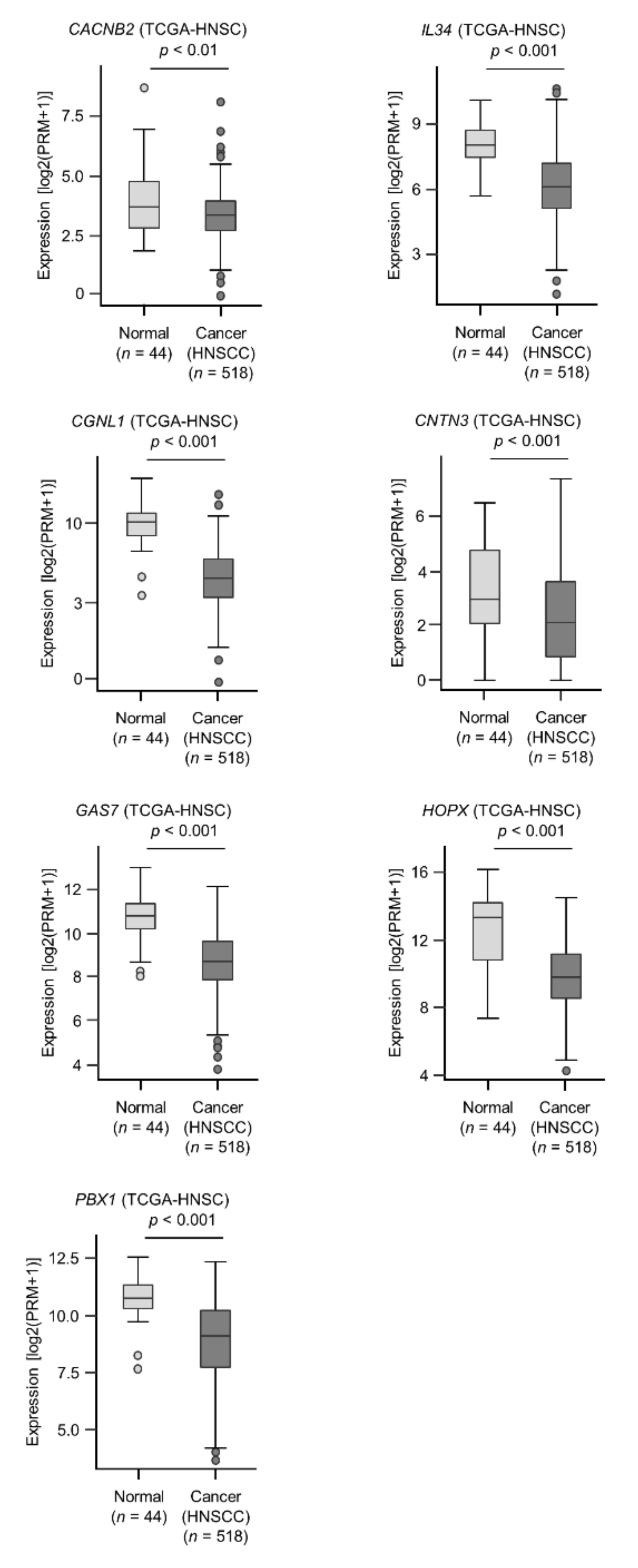
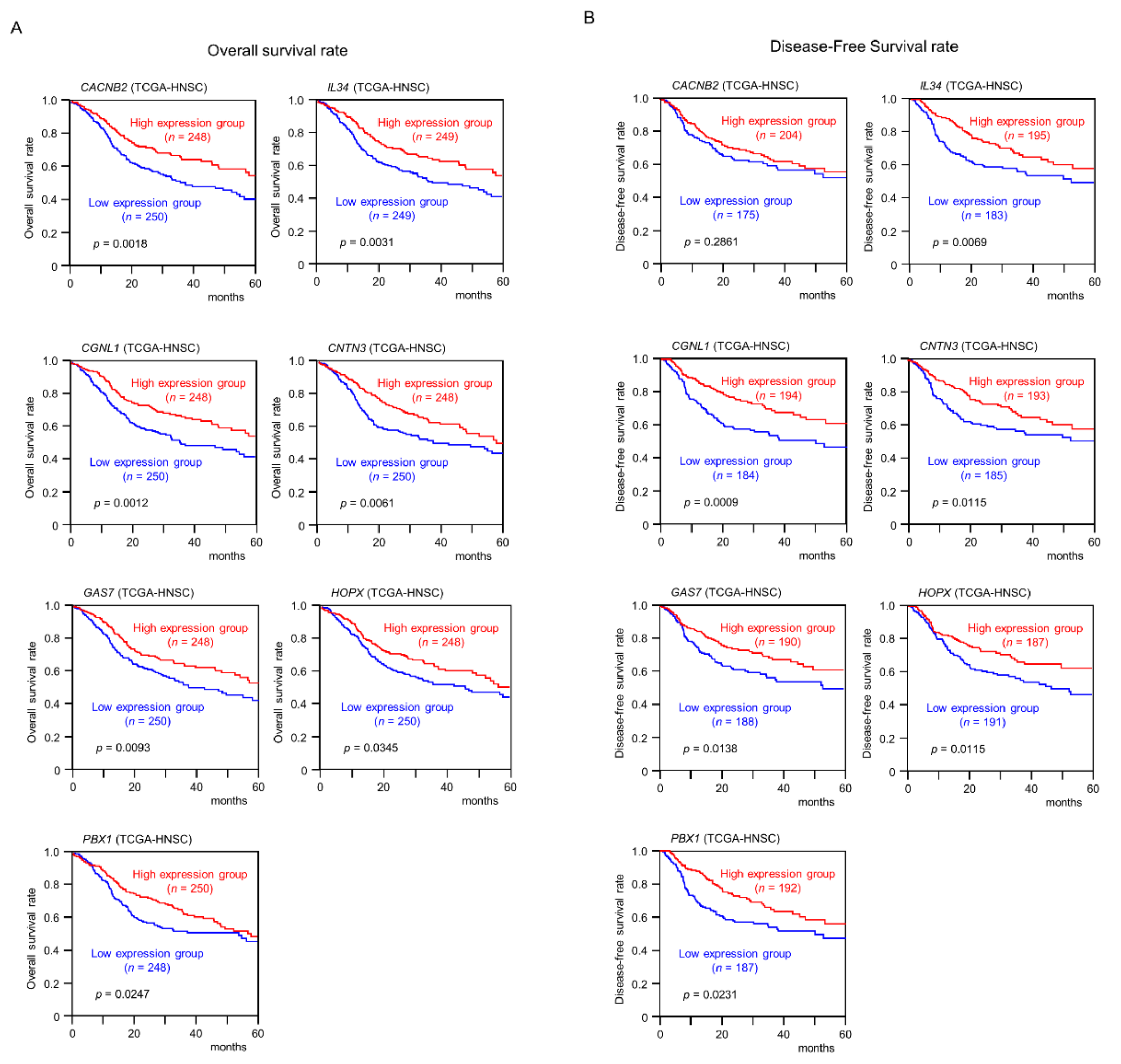
2.5. Clinical Significance of miR-31-5p and miR-31-3p Targets in HNSCC Cells
2.6. Direct Regulation of CGNL1 by miR-31-3p in HNSCC Cells
3. Discussion
4. Materials and Methods
4.1. Clinical HNSCC and Normal Epithelial Tissue Specimens and HNSCC Cell Lines
4.2. Determination of the miRNA Expression Signature in HNSCC by RNA Sequencing
4.3. RNA Extraction and Quantitative Reverse-Transcription Polymerase Chain Reaction (qRT-PCR)
4.4. Transfection of Mirnas Precursors and Inhibitors into HNSCC Cells
4.5. Functional Assays (Cell Proliferation, Migration, and Invasion) in HNSCC Cells
4.6. Analysis of the Clinical Significance of HNSCC Patients Using TCGA-HNSC Data
4.7. Western Blotting
4.8. Plasmid Construction and Dual-Luciferase Reporter Assays
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leemans, C.R.; Braakhuis, B.J.; Brakenhoff, R.H. The molecular biology of head and neck cancer. Nat. Rev. Cancer 2011, 11, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Chow, L.Q.M. Head and Neck Cancer. N. Engl. J. Med. 2020, 382, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Bonner, J.A.; Harari, P.M.; Giralt, J.; Cohen, R.B.; Jones, C.U.; Sur, R.K.; Raben, D.; Baselga, J.; Spencer, S.A.; Zhu, J.; et al. Radiotherapy plus cetuximab for locoregionally advanced head and neck cancer: 5-year survival data from a phase 3 randomised trial, and relation between cetuximab-induced rash and survival. Lancet Oncol. 2010, 11, 21–28. [Google Scholar] [CrossRef]
- Ferris, R.L.; Blumenschein, G., Jr.; Fayette, J.; Guigay, J.; Colevas, A.D.; Licitra, L.; Harrington, K.; Kasper, S.; Vokes, E.E.; Even, C.C.; et al. Nivolumab for recurrent squamous-cell carcinoma of the head and neck. Br. Dent. J. 2016, 221, 632. [Google Scholar] [CrossRef]
- Hsieh, J.C.; Wang, H.M.; Wu, M.H.; Chang, K.P.; Chang, P.H.; Liao, C.T.; Liau, C.T. Review of emerging biomarkers in head and neck squamous cell carcinoma in the era of immunotherapy and targeted therapy. Head Neck 2019, 41 (Suppl. S1), 19–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betel, D.; Wilson, M.; Gabow, A.; Marks, D.S.; Sander, C. The microRNA.org resource: Targets and expression. Nucleic Acids Res. 2008, 36, D149–D153. [Google Scholar] [CrossRef] [Green Version]
- Anfossi, S.; Babayan, A.; Pantel, K.; Calin, G.A. Clinical utility of circulating non-coding RNAs—An update. Nat. Rev. Clin. Oncol. 2018, 15, 541–563. [Google Scholar] [CrossRef]
- Krek, A.; Grün, D.; Poy, M.N.; Wolf, R.; Rosenberg, L.; Epstein, E.J.; MacMenamin, P.; da Piedade, I.; Gunsalus, K.C.; Stoffel, M.; et al. Combinatorial microRNA target predictions. Nat. Genet. 2005, 37, 495–500. [Google Scholar] [CrossRef]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Croce, C.M. MicroRNA dysregulation in cancer: Diagnostics, monitoring and therapeutics. A comprehensive review. EMBO Mol. Med. 2012, 4, 43–59. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Okada, R.; Koshizuka, K.; Yamada, Y.; Moriya, S.; Kikkawa, N.; Kinoshita, T.; Hanazawa, T.; Seki, N. Regulation of Oncogenic Targets by miR-99a-3p (Passenger Strand of miR-99a-Duplex) in Head and Neck Squamous Cell Carcinoma. Cells 2019, 8, 1535. [Google Scholar] [CrossRef] [Green Version]
- Yamada, Y.; Koshizuka, K.; Hanazawa, T.; Kikkawa, N.; Okato, A.; Idichi, T.; Arai, T.; Sugawara, S.; Katada, K.; Okamoto, Y.; et al. Passenger strand of miR-145-3p acts as a tumor-suppressor by targeting MYO1B in head and neck squamous cell carcinoma. Int. J. Oncol. 2018, 52, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Koshizuka, K.; Hanazawa, T.; Kikkawa, N.; Katada, K.; Okato, A.; Arai, T.; Idichi, T.; Osako, Y.; Okamoto, Y.; Seki, N. Antitumor miR-150-5p and miR-150-3p inhibit cancer cell aggressiveness by targeting SPOCK1 in head and neck squamous cell carcinoma. Auris Nasus Larynx 2018, 45, 854–865. [Google Scholar] [CrossRef] [PubMed]
- Koshizuka, K.; Hanazawa, T.; Kikkawa, N.; Arai, T.; Okato, A.; Kurozumi, A.; Kato, M.; Katada, K.; Okamoto, Y.; Seki, N. Regulation of ITGA3 by the anti-tumor miR-199 family inhibits cancer cell migration and invasion in head and neck cancer. Cancer Sci. 2017, 108, 1681–1692. [Google Scholar] [CrossRef] [Green Version]
- Mitra, R.; Adams, C.M.; Jiang, W.; Greenawalt, E.; Eischen, C.M. Pan-cancer analysis reveals cooperativity of both strands of microRNA that regulate tumorigenesis and patient survival. Nat. Commun. 2020, 11, 968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, T.; Ma, P.; Wu, D.; Shu, Y.; Gao, W. Functions and mechanisms of microRNA-31 in human cancers. Biomed. Pharmacother. 2018, 108, 1162–1169. [Google Scholar] [CrossRef]
- Gao, Y.; Yi, J.; Zhang, K.; Bai, F.; Feng, B.; Wang, R.; Chu, X.; Chen, L.; Song, H. Downregulation of MiR-31 stimulates expression of LATS2 via the hippo pathway and promotes epithelial-mesenchymal transition in esophageal squamous cell carcinoma. J. Exp. Clin. Cancer Res. 2017, 36, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.J.; Tsai, M.M.; Hung, P.S.; Kao, S.Y.; Liu, T.Y.; Wu, K.J.; Chiou, S.H.; Lin, S.C.; Chang, K.W. miR-31 ablates expression of the HIF regulatory factor FIH to activate the HIF pathway in head and neck carcinoma. Cancer Res. 2010, 70, 1635–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, B.; Cao, X.; Zhang, W.; Pan, G.; Yi, Q.; Zhong, W.; Yan, D. MicroRNA-31-5p enhances the Warburg effect via targeting FIH. FASEB J. 2019, 33, 545–556. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.C.; Kao, S.Y.; Yang, C.C.; Tu, H.F.; Wu, C.H.; Chang, K.W.; Lin, S.C. EGF up-regulates miR-31 through the C/EBPβ signal cascade in oral carcinoma. PLoS ONE 2014, 9, e108049. [Google Scholar] [CrossRef]
- Hung, P.S.; Tu, H.F.; Kao, S.Y.; Yang, C.C.; Liu, C.J.; Huang, T.Y.; Chang, K.W.; Lin, S.C. miR-31 is upregulated in oral premalignant epithelium and contributes to the immortalization of normal oral keratinocytes. Carcinogenesis 2014, 35, 1162–1171. [Google Scholar] [CrossRef] [Green Version]
- She, B.R.; Liou, G.G.; Lin-Chao, S. Association of the growth-arrest-specific protein Gas7 with F-actin induces reorganization of microfilaments and promotes membrane outgrowth. Exp. Cell Res. 2002, 273, 34–44. [Google Scholar] [CrossRef]
- Gotoh, A.; Hidaka, M.; Hirose, K.; Uchida, T. Gas7b (growth arrest specific protein 7b) regulates neuronal cell morphology by enhancing microtubule and actin filament assembly. J. Biol. Chem. 2013, 288, 34699–34706. [Google Scholar] [CrossRef] [Green Version]
- Tseng, R.C.; Chang, J.W.; Mao, J.S.; Tsai, C.D.; Wu, P.C.; Lin, C.J.; Lu, Y.L.; Liao, S.Y.; Cheng, H.C.; Hsu, H.S.; et al. Growth-arrest-specific 7C protein inhibits tumor metastasis via the N-WASP/FAK/F-actin and hnRNP U/β-TrCP/β-catenin pathways in lung cancer. Oncotarget 2015, 6, 44207–44221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Yeo, K.S.; Lopez, G.; Zhang, C.; Dankert Eggum, E.N.; Rokita, J.L.; Ung, C.Y.; Levee, T.M.; Her, Z.P.; Howe, C.J.; et al. GAS7 Deficiency Promotes Metastasis in MYCN-driven Neuroblastoma. Cancer Res. 2021, 81, 2995–3007. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Kono, T.; Monte, M.M.; Kuse, H.; Costa, M.M.; Korenaga, H.; Maehr, T.; Husain, M.; Sakai, M.; Secombes, C.J. Identification of IL-34 in teleost fish: Differential expression of rainbow trout IL-34, MCSF1 and MCSF2, ligands of the MCSF receptor. Mol. Immunol. 2013, 53, 398–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ushach, I.; Zlotnik, A. Biological role of granulocyte macrophage colony-stimulating factor (GM-CSF) and macrophage colony-stimulating factor (M-CSF) on cells of the myeloid lineage. J. Leukoc. Biol. 2016, 100, 481–489. [Google Scholar] [CrossRef]
- Lelios, I.; Cansever, D.; Utz, S.G.; Mildenberger, W.; Stifter, S.A.; Greter, M. Emerging roles of IL-34 in health and disease. J. Exp. Med. 2020, 217, e20190290. [Google Scholar] [CrossRef] [PubMed]
- Van Itallie, C.M.; Anderson, J.M. Architecture of tight junctions and principles of molecular composition. Semin. Cell Dev. Biol. 2014, 36, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Chrifi, I.; Hermkens, D.; Brandt, M.M.; van Dijk, C.G.M.; Bürgisser, P.E.; Haasdijk, R.; Pei, J.; van de Kamp, E.H.M.; Zhu, C.; Blonden, L.; et al. Cgnl1, an endothelial junction complex protein, regulates GTPase mediated angiogenesis. Cardiovasc. Res. 2017, 113, 1776–1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paschoud, S.; Guillemot, L.; Citi, S. Distinct domains of paracingulin are involved in its targeting to the actin cytoskeleton and regulation of apical junction assembly. J. Biol. Chem. 2012, 287, 13159–13169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Citi, S.; Guerrera, D.; Spadaro, D.; Shah, J. Epithelial junctions and Rho family GTPases: The zonular signalosome. Small GTPases 2014, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Guillemot, L.; Paschoud, S.; Jond, L.; Foglia, A.; Citi, S. Paracingulin regulates the activity of Rac1 and RhoA GTPases by recruiting Tiam1 and GEF-H1 to epithelial junctions. Mol. Biol. Cell 2008, 19, 4442–4453. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshizuka, K.; Nohata, N.; Hanazawa, T.; Kikkawa, N.; Arai, T.; Okato, A.; Fukumoto, I.; Katada, K.; Okamoto, Y.; Seki, N. Deep sequencing-based microRNA expression signatures in head and neck squamous cell carcinoma: Dual strands of pre-miR-150 as antitumor miRNAs. Oncotarget 2017, 8, 30288–30304. [Google Scholar] [CrossRef] [Green Version]
- Toda, H.; Kurozumi, S.; Kijima, Y.; Idichi, T.; Shinden, Y.; Yamada, Y.; Arai, T.; Maemura, K.; Fujii, T.; Horiguchi, J.; et al. Molecular pathogenesis of triple-negative breast cancer based on microRNA expression signatures: Antitumor miR-204-5p targets AP1S3. J. Hum. Genet. 2018, 63, 1197–1210. [Google Scholar] [CrossRef]
- Toda, H.; Seki, N.; Kurozumi, S.; Shinden, Y.; Yamada, Y.; Nohata, N.; Moriya, S.; Idichi, T.; Maemura, K.; Fujii, T.; et al. RNA-sequence-based microRNA expression signature in breast cancer: Tumor-suppressive miR-101-5p regulates molecular pathogenesis. Mol. Oncol. 2020, 14, 426–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, M.; Goto, Y.; Tanaka, T.; Okada, R.; Moriya, S.; Idichi, T.; Noda, M.; Sasaki, K.; Kita, Y.; Kurahara, H.; et al. RNA sequencing-based microRNA expression signature in esophageal squamous cell carcinoma: Oncogenic targets by antitumor miR-143-5p and miR-143-3p regulation. J. Hum. Genet. 2020, 65, 1019–1034. [Google Scholar] [CrossRef] [PubMed]
- Anaya, J. OncoLnc: Linking TCGA survival data to mRNAs, miRNAs, and lncRNAs. PeerJ Comput. Sci. 2016, 2, e67. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | ||||
|---|---|---|---|---|
| Entrez Gene ID | Gene Symbol | Gene Name | Fold Change (log2 < −2.0) | Total Sites |
| 5563 | PRKAA2 | protein kinase, AMP-activated, alpha 2 catalytic subunit | −4.56 | 1 |
| 83699 | SH3BGRL2 | SH3 domain binding glutamate-rich protein like 2 | −4.45 | 1 |
| 6517 | SLC2A4 | solute carrier family 2 (facilitated glucose transporter), member 4 | −4.29 | 1 |
| 2252 | FGF7 | fibroblast growth factor 7 | −3.81 | 1 |
| 55607 | PPP1R9A | protein phosphatase 1, regulatory subunit 9A | −3.73 | 1 |
| 5549 | PRELP | proline/arginine-rich end leucine-rich repeat protein | −3.66 | 2 |
| 5083 | PAX9 | paired box 9 | −3.62 | 1 |
| 26084 | ARHGEF26 | Rho guanine nucleotide exchange factor (GEF) 26 | −3.58 | 1 |
| 252995 | FNDC5 | fibronectin type III domain containing 5 | −3.50 | 1 |
| 51209 | RAB9B | RAB9B, member RAS oncogene family | −3.23 | 1 |
| 2899 | GRIK3 | glutamate receptor, ionotropic, kainate 3 | −2.88 | 1 |
| 401474 | SAMD12 | sterile alpha motif domain containing 12 | −2.84 | 1 |
| 60529 | ALX4 | ALX homeobox 4 | −2.63 | 1 |
| 64399 | HHIP | hedgehog interacting protein | −2.53 | 1 |
| 146433 | IL34 | interleukin 34 | −2.45 | 1 |
| 84144 | SYDE2 | synapse defective 1, Rho GTPase, homolog 2 (C. elegans) | −2.44 | 2 |
| 619279 | ZNF704 | zinc finger protein 704 | −2.40 | 1 |
| 783 | CACNB2 | calcium channel, voltage-dependent, beta 2 subunit | −2.34 | 1 |
| 5493 | PPL | periplakin | −2.27 | 1 |
| 3670 | ISL1 | ISL LIM homeobox 1 | −2.24 | 2 |
| 389208 | TMPRSS11F | transmembrane protease, serine 11F | −2.17 | 1 |
| 168667 | BMPER | BMP binding endothelial regulator | −2.16 | 1 |
| 1983 | EIF5 | eukaryotic translation initiation factor 5 | −2.14 | 1 |
| 5100 | PCDH8 | protocadherin 8 | −2.06 | 2 |
| B | ||||
| Entrez Gene ID | Gene Symbol | Gene Name | Fold Change (log2 < −2.0) | Total Sites |
| 420 | ART4 | ADP-ribosyltransferase 4 (Dombrock blood group) | −6.92 | 1 |
| 5075 | PAX1 | paired box 1 | −6.47 | 1 |
| 1805 | DPT | dermatopontin | −5.69 | 1 |
| 10218 | ANGPTL7 | angiopoietin-like 7 | −5.09 | 1 |
| 2315 | MLANA | melan-A | −4.81 | 1 |
| 55286 | C4orf19 | chromosome 4 open reading frame 19 | −4.76 | 1 |
| 8839 | WISP2 | WNT1 inducible signaling pathway protein 2 | −4.72 | 1 |
| 440854 | CAPN14 | calpain 14 | −4.70 | 1 |
| 6422 | SFRP1 | secreted frizzled-related protein 1 | −4.70 | 1 |
| 114905 | C1QTNF7 | C1q and tumor necrosis factor related protein 7 | −4.66 | 1 |
| 9068 | ANGPTL1 | angiopoietin-like 1 | −4.56 | 1 |
| 5563 | PRKAA2 | protein kinase, AMP-activated, alpha 2 catalytic subunit | −4.56 | 2 |
| 5104 | SERPINA5 | serpin peptidase inhibitor, clade A (alpha-1 antiproteinase, antitrypsin), member 5 | −4.45 | 1 |
| 148213 | ZNF681 | zinc finger protein 681 | −4.27 | 1 |
| 127435 | PODN | podocan | −4.20 | 1 |
| 53405 | CLIC5 | chloride intracellular channel 5 | −4.16 | 1 |
| 85477 | SCIN | scinderin | −4.09 | 1 |
| 255798 | SMCO1/C3orf43 | single-pass membrane protein with coiled-coil domains 1 | −4.01 | 1 |
| 53353 | LRP1B | low density lipoprotein receptor-related protein 1B | −4.00 | 1 |
| 23242 | COBL | cordon-bleu WH2 repeat protein | −3.89 | 1 |
| 5570 | PKIB | protein kinase (cAMP-dependent, catalytic) inhibitor beta | −3.84 | 1 |
| 440730 | TRIM67 | tripartite motif containing 67 | −3.83 | 1 |
| 2252 | FGF7 | fibroblast growth factor 7 | −3.81 | 1 |
| 84525 | HOPX | HOP homeobox | −3.81 | 1 |
| 389432 | SAMD5 | sterile alpha motif domain containing 5 | −3.79 | 1 |
| 8736 | MYOM1 | myomesin 1 | −3.68 | 1 |
| 5549 | PRELP | proline/arginine-rich end leucine-rich repeat protein | −3.66 | 2 |
| 137735 | ABRA | actin-binding Rho activating protein | −3.58 | 1 |
| 785 | CACNB4 | calcium channel, voltage-dependent, beta 4 subunit | −3.57 | 3 |
| 79442 | LRRC2 | leucine rich repeat containing 2 | −3.55 | 2 |
| 339512 | C1orf110 | chromosome 1 open reading frame 110 | −3.50 | 1 |
| 10894 | LYVE1 | lymphatic vessel endothelial hyaluronan receptor 1 | −3.43 | 1 |
| 3768 | KCNJ12 | potassium channel, inwardly rectifying subfamily J, member 12 | −3.36 | 1 |
| 171024 | SYNPO2 | synaptopodin 2 | −3.35 | 1 |
| 114786 | XKR4 | XK, Kell blood group complex subunit-related family, member 4 | −3.33 | 1 |
| 84952 | CGNL1 | cingulin-like 1 | −3.30 | 2 |
| 55335 | NIPSNAP3B | nipsnap homolog 3B (C. elegans) | −3.27 | 1 |
| 3479 | IGF1 | insulin-like growth factor 1 (somatomedin C) | −3.26 | 2 |
| 2690 | GHR | growth hormone receptor | −3.20 | 1 |
| 8522 | GAS7 | growth arrest-specific 7 | −3.18 | 1 |
| 2066 | ERBB4 | erb-b2 receptor tyrosine kinase 4 | −3.14 | 2 |
| 202333 | CMYA5 | cardiomyopathy associated 5 | −3.13 | 1 |
| 22865 | SLITRK3 | SLIT and NTRK-like family, member 3 | −3.13 | 1 |
| 51666 | ASB4 | ankyrin repeat and SOCS box containing 4 | −3.08 | 1 |
| 22871 | NLGN1 | neuroligin 1 | −3.08 | 1 |
| 4958 | OMD | osteomodulin | −3.08 | 1 |
| 5178 | PEG3 | paternally expressed 3 | −3.06 | 1 |
| 29119 | CTNNA3 | catenin (cadherin-associated protein), alpha 3 | −3.04 | 2 |
| 8529 | CYP4F2 | cytochrome P450, family 4, subfamily F, polypeptide 2 | −3.01 | 1 |
| 343450 | KCNT2 | potassium channel, sodium activated subfamily T, member 2 | −3.00 | 1 |
| 5087 | PBX1 | pre-B-cell leukemia homeobox 1 | −2.98 | 1 |
| 387758 | FIBIN | fin bud initiation factor homolog (zebrafish) | −2.96 | 1 |
| 57689 | LRRC4C | leucine rich repeat containing 4C | −2.96 | 1 |
| 79071 | ELOVL6 | ELOVL fatty acid elongase 6 | −2.95 | 1 |
| 6542 | SLC7A2 | solute carrier family 7 (cationic amino acid transporter, y+ system), member 2 | −2.94 | 1 |
| 6450 | SH3BGR | SH3 domain binding glutamate-rich protein | −2.93 | 1 |
| 7276 | TTR | transthyretin | −2.92 | 2 |
| 23732 | FRRS1L/C9orf4 | ferric-chelate reductase 1-like | −2.89 | 1 |
| 220963 | SLC16A9 | solute carrier family 16, member 9 | −2.88 | 1 |
| 55 | ACPP | acid phosphatase, prostate | −2.84 | 1 |
| 401474 | SAMD12 | sterile alpha motif domain containing 12 | −2.84 | 1 |
| 8153 | RND2 | Rho family GTPase 2 | −2.83 | 1 |
| 7135 | TNNI1 | troponin I type 1 (skeletal, slow) | −2.82 | 1 |
| 340596 | LHFPL1 | lipoma HMGIC fusion partner-like 1 | −2.77 | 1 |
| 26974 | ZNF285 | zinc finger protein 285 | −2.74 | 1 |
| 2053 | EPHX2 | epoxide hydrolase 2, cytoplasmic | −2.73 | 1 |
| 386618 | KCTD4 | potassium channel tetramerization domain containing 4 | −2.73 | 1 |
| 1183 | CLCN4 | chloride channel, voltage-sensitive 4 | −2.69 | 1 |
| 291 | SLC25A4 | solute carrier family 25 (mitochondrial carrier; adenine nucleotide translocator), member 4 | −2.68 | 1 |
| 4023 | LPL | lipoprotein lipase | −2.65 | 1 |
| 32 | ACACB | acetyl-CoA carboxylase beta | −2.64 | 1 |
| 55244 | SLC47A1 | solute carrier family 47 (multidrug and toxin extrusion), member 1 | −2.64 | 1 |
| 84620 | ST6GAL2 | ST6 beta-galactosamide alpha-2,6-sialyltranferase 2 | −2.62 | 1 |
| 26032 | SUSD5 | sushi domain containing 5 | −2.61 | 1 |
| 6857 | SYT1 | synaptotagmin I | −2.61 | 2 |
| 6391 | SDHC | succinate dehydrogenase complex, subunit C, integral membrane protein, 15kDa | −2.60 | 1 |
| 5506 | PPP1R3A | protein phosphatase 1, regulatory subunit 3A | −2.58 | 2 |
| 367 | AR | androgen receptor | −2.57 | 2 |
| 64399 | HHIP | hedgehog interacting protein | −2.53 | 1 |
| 56898 | BDH2 | 3-hydroxybutyrate dehydrogenase, type 2 | −2.52 | 2 |
| 9077 | DIRAS3 | DIRAS family, GTP-binding RAS-like 3 | −2.52 | 1 |
| 154661 | RUNDC3B | RUN domain containing 3B | −2.52 | 1 |
| 8796 | SCEL | sciellin | −2.52 | 1 |
| 50937 | CDON | cell adhesion associated, oncogene regulated | −2.49 | 1 |
| 6660 | SOX5 | SRY (sex determining region Y)-box 5 | −2.48 | 1 |
| 56172 | ANKH | ANKH inorganic pyrophosphate transport regulator | −2.46 | 1 |
| 6092 | ROBO2 | roundabout, axon guidance receptor, homolog 2 (Drosophila) | −2.46 | 1 |
| 158326 | FREM1 | FRAS1 related extracellular matrix 1 | −2.45 | 1 |
| 10345 | TRDN | triadin | −2.45 | 1 |
| 158866 | ZDHHC15 | zinc finger, DHHC-type containing 15 | −2.44 | 1 |
| 55283 | MCOLN3 | mucolipin 3 | −2.42 | 1 |
| 653316 | FAM153C | family with sequence similarity 153, member C, pseudogene | −2.41 | 1 |
| 348158 | ACSM2B | acyl-CoA synthetase medium-chain family member 2B | −2.39 | 1 |
| 11227 | GALNT5 | polypeptide N-acetylgalactosaminyltransferase 5 | −2.39 | 1 |
| 3169 | FOXA1 | forkhead box A1 | −2.37 | 1 |
| 284716 | RIMKLA | ribosomal modification protein rimK-like family member A | −2.37 | 2 |
| 253559 | CADM2 | cell adhesion molecule 2 | −2.36 | 1 |
| 144453 | BEST3 | bestrophin 3 | −2.35 | 1 |
| 2258 | FGF13 | fibroblast growth factor 13 | −2.35 | 1 |
| 57863 | CADM3 | cell adhesion molecule 3 | −2.34 | 1 |
| 140456 | ASB11 | ankyrin repeat and SOCS box containing 11, E3 ubiquitin protein ligase | −2.32 | 2 |
| 346389 | MACC1 | metastasis associated in colon cancer 1 | −2.30 | 2 |
| 9378 | NRXN1 | neurexin 1 | −2.30 | 1 |
| 151887 | CCDC80 | coiled-coil domain containing 80 | −2.29 | 2 |
| 266977 | GPR110 | G protein-coupled receptor 110 | −2.28 | 1 |
| 3481 | IGF2 | insulin-like growth factor 2 | −2.27 | 1 |
| 57554 | LRRC7 | leucine rich repeat containing 7 | −2.27 | 1 |
| 80310 | PDGFD | platelet derived growth factor D | −2.25 | 1 |
| 342926 | ZNF677 | zinc finger protein 677 | −2.25 | 1 |
| 341640 | FREM2 | FRAS1 related extracellular matrix protein 2 | −2.24 | 1 |
| 5067 | CNTN3 | contactin 3 (plasmacytoma associated) | −2.22 | 1 |
| 4919 | ROR1 | receptor tyrosine kinase-like orphan receptor 1 | −2.20 | 1 |
| 948 | CD36 | CD36 molecule (thrombospondin receptor) | −2.19 | 1 |
| 23171 | GPD1L | glycerol-3-phosphate dehydrogenase 1-like | −2.18 | 1 |
| 64102 | TNMD | tenomodulin | −2.18 | 2 |
| 55638 | SYBU | syntabulin (syntaxin-interacting) | −2.17 | 1 |
| 6586 | SLIT3 | slit homolog 3 (Drosophila) | −2.13 | 2 |
| 2247 | FGF2 | fibroblast growth factor 2 (basic) | −2.11 | 1 |
| 115827 | RAB3C | RAB3C, member RAS oncogene family | −2.11 | 2 |
| 203859 | ANO5 | anoctamin 5 | −2.10 | 1 |
| 80110 | ZNF614 | zinc finger protein 614 | −2.10 | 1 |
| 115265 | DDIT4L | DNA-damage-inducible transcript 4-like | −2.03 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oshima, S.; Asai, S.; Seki, N.; Minemura, C.; Kinoshita, T.; Goto, Y.; Kikkawa, N.; Moriya, S.; Kasamatsu, A.; Hanazawa, T.; et al. Identification of Tumor Suppressive Genes Regulated by miR-31-5p and miR-31-3p in Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2021, 22, 6199. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126199
Oshima S, Asai S, Seki N, Minemura C, Kinoshita T, Goto Y, Kikkawa N, Moriya S, Kasamatsu A, Hanazawa T, et al. Identification of Tumor Suppressive Genes Regulated by miR-31-5p and miR-31-3p in Head and Neck Squamous Cell Carcinoma. International Journal of Molecular Sciences. 2021; 22(12):6199. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126199
Chicago/Turabian StyleOshima, Sachi, Shunichi Asai, Naohiko Seki, Chikashi Minemura, Takashi Kinoshita, Yusuke Goto, Naoko Kikkawa, Shogo Moriya, Atsushi Kasamatsu, Toyoyuki Hanazawa, and et al. 2021. "Identification of Tumor Suppressive Genes Regulated by miR-31-5p and miR-31-3p in Head and Neck Squamous Cell Carcinoma" International Journal of Molecular Sciences 22, no. 12: 6199. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126199