
Superior Alignment of Human iPSC-Osteoblasts Associated with Focal Adhesion Formation Stimulated by Oriented Collagen Scaffold

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
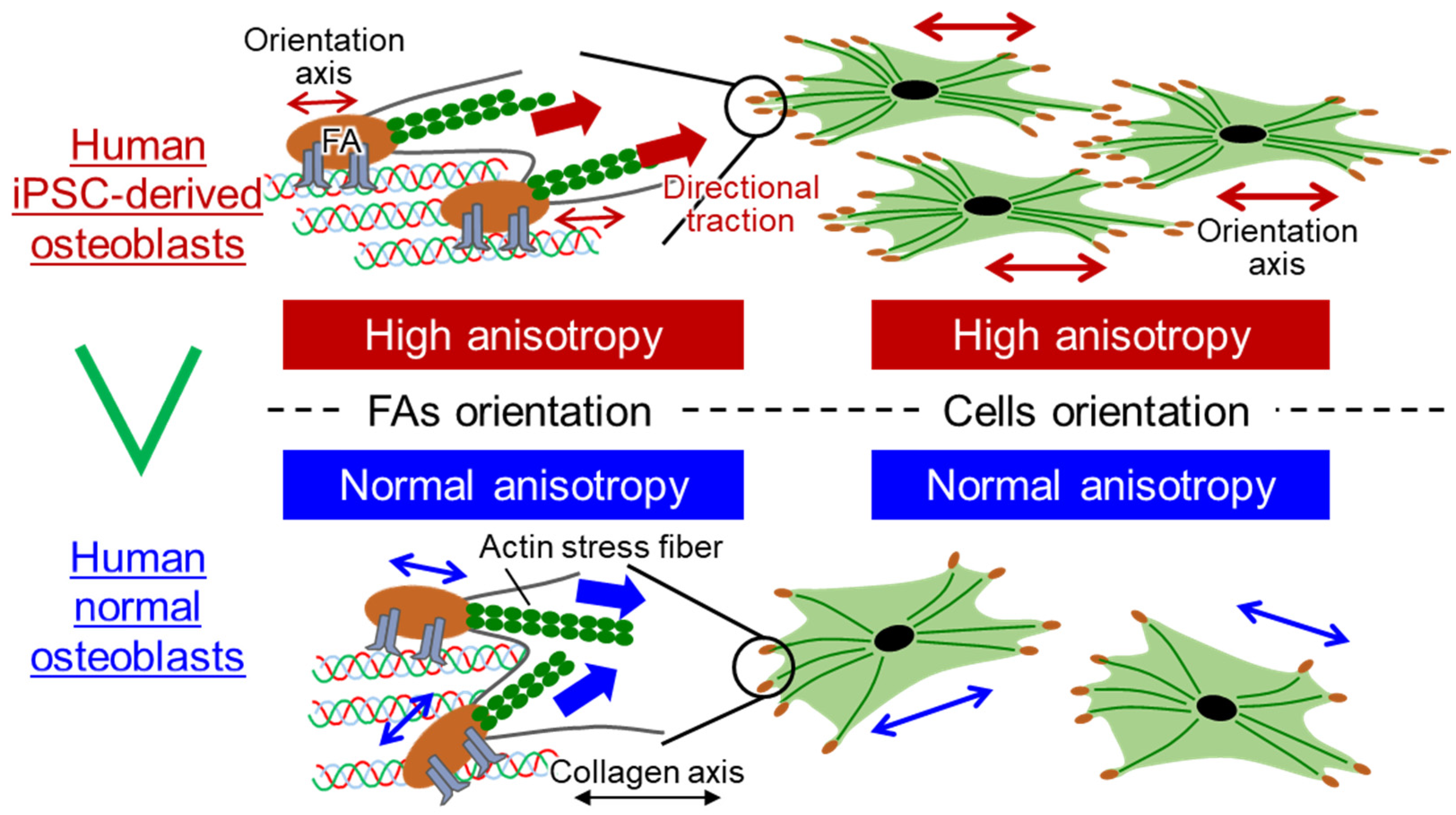{kind=link}
Abstract
:1. Introduction
2. Results
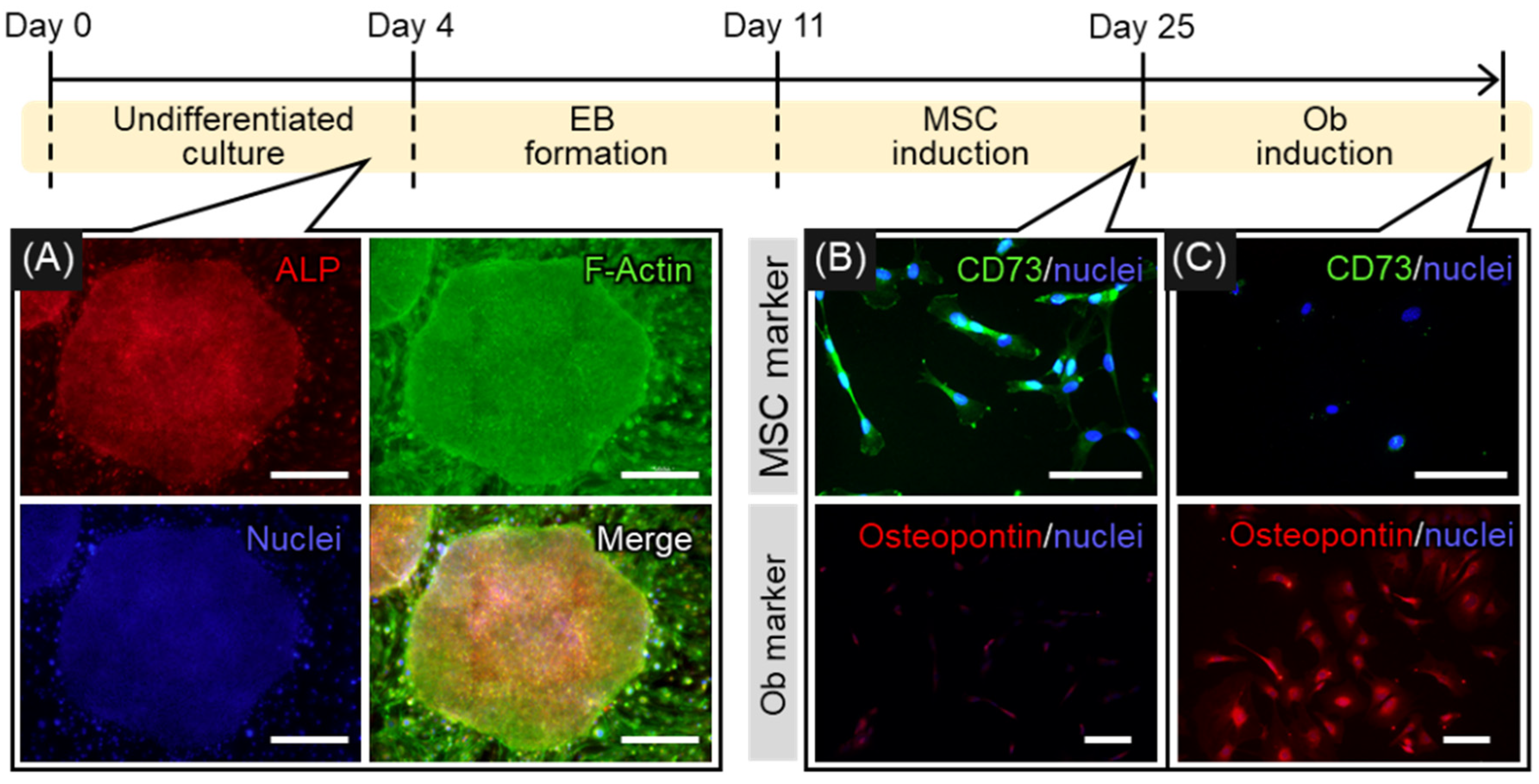
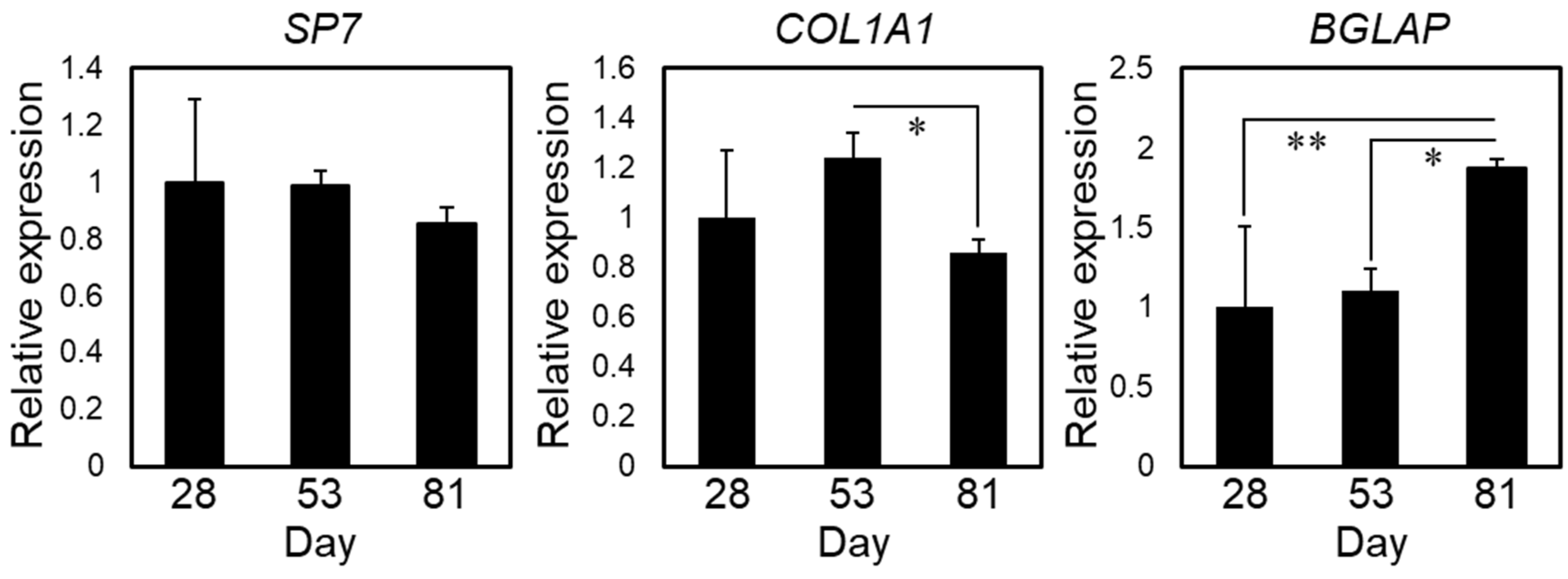
2.1. Induction of Osteogenic Differentiation of hiPSCs
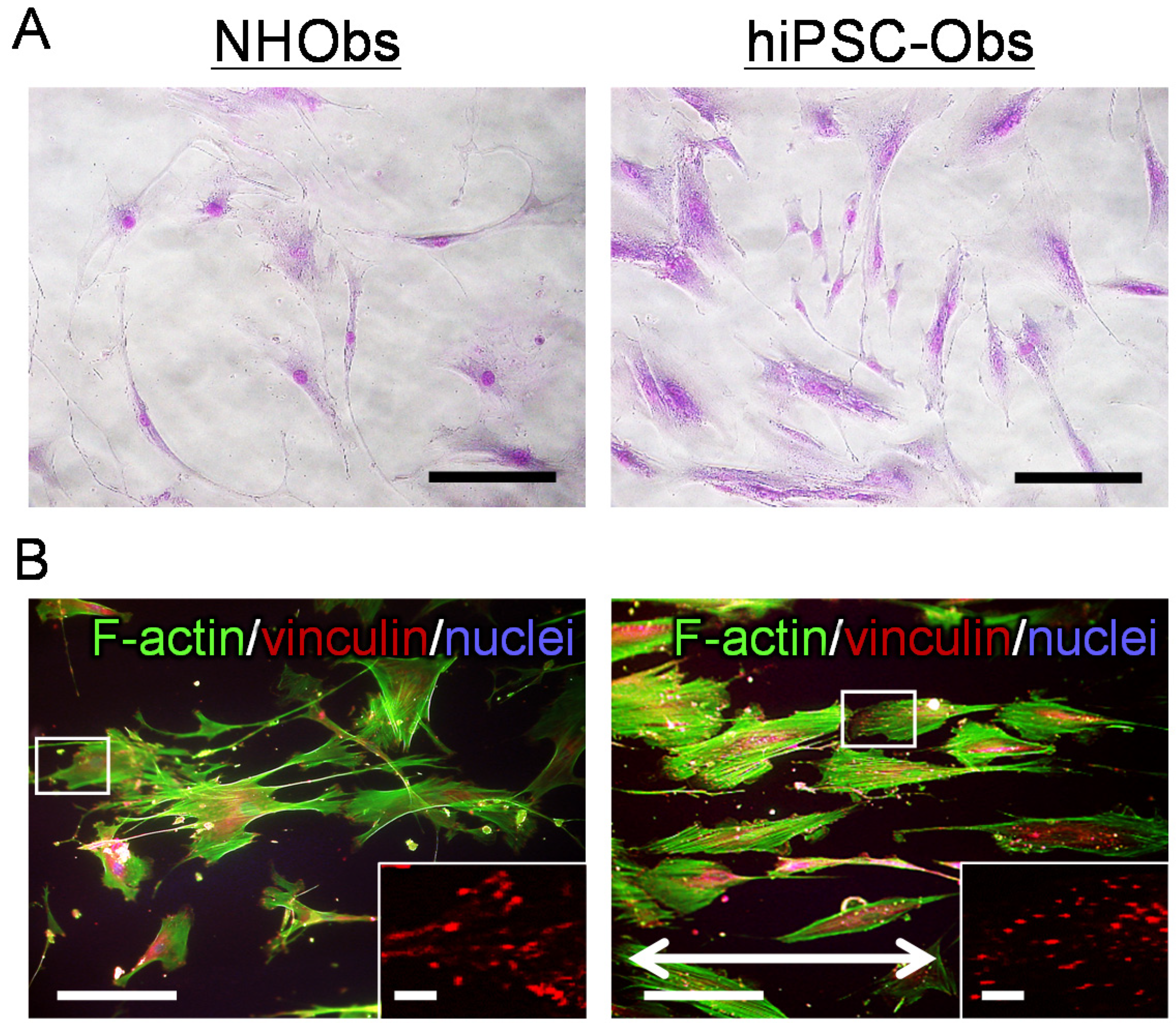
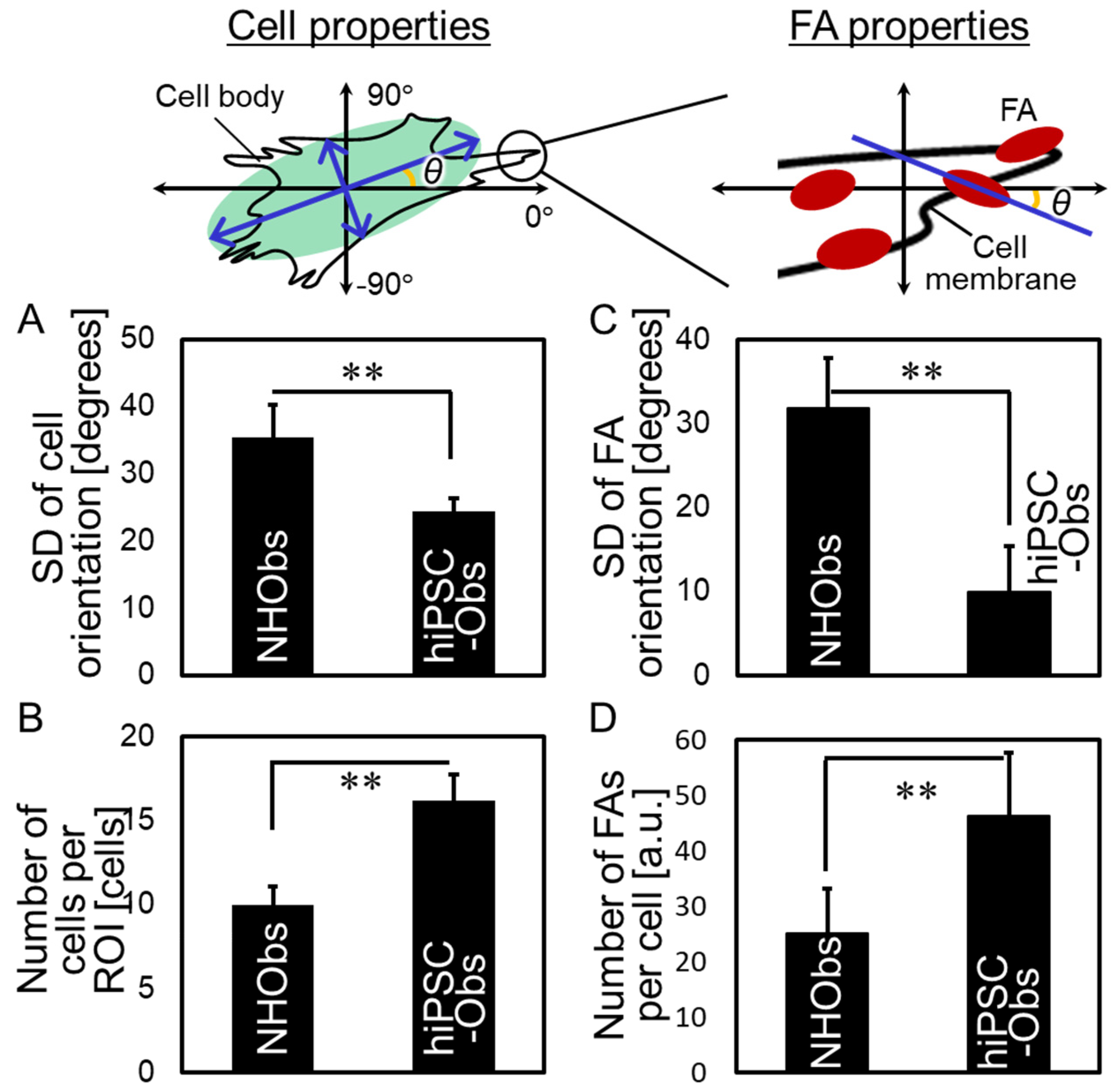
2.2. Cellular Behavior on a Bone-Mimetic Collagen Scaffold
3. Discussion
4. Materials and Methods
4.1. Culture of SNL Feeder Cells
4.2. Culture of hiPSCs
4.3. Osteogenic Differentiation of hiPSCs
4.4. Culture of NHObs
4.5. Gene Expression Analysis
4.6. Immunostaining
4.7. Quantitative Analysis of the Properties of Cells and Focal Adhesions (FAs)
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Bock, C.; Kiskinis, E.; Verstappen, G.; Gu, H.; Boulting, G.; Smith, Z.D.; Ziller, M.; Croft, G.F.; Amoroso, M.W.; Oakley, D.H.; et al. Reference Maps of Human Es and Ips Cell Variation Enable High-Throughput Characterization of Pluripotent Cell Lines. Cell 2011, 144, 439–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Im, G.I. Embryonic Stem Cells and Induced Pluripotent Stem Cells for Skeletal Regeneration. Tissue Eng. B 2014, 20, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Yang, B.; Hu, K.; Cao, C.; Man, Y.; Wang, P. Deriving Osteogenic Cells from Induced Pluripotent Stem Cells for Bone Tissue Engineering. Tissue Eng. B 2017, 23, 1–8. [Google Scholar] [CrossRef]
- Chamberlain, G.; Fox, J.; Ashton, B.; Middleton, J. Concise Review: Mesenchymal Stem Cells: Their Phenotype, Differentiation Capacity, Immunological Features, and Potential for Homing. Stem Cells 2007, 25, 2739–2749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumaran, S.T.; Arun, K.V.; Sudarsan, S.; Talwar, A.; Srinivasan, N. Osteoblast Response to Commercially Available Demineralized Bone Matrices—An In-Vitro Study. Indian J. Dent. Res. 2010, 21, 3–9. [Google Scholar] [PubMed]
- Lou, X. Induced Pluripotent Stem Cells as a New Strategy for Osteogenesis and Bone Regeneration. Stem Cell Rev. Rep. 2015, 11, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Geng, X.; Chen, Z.; Zhao, L. A Novel and Expeditious Method to Fabricate Superhydrophobic Metal Carboxylate Surface. Appl. Phys. A 2012, 106, 35–40. [Google Scholar] [CrossRef]
- Bilousova, G.; Jun, D.H.; King, K.B.; De Langhe, S.; Chick, W.S.; Torchia, E.C.; Chow, K.S.; Klemm, D.J.; Roop, D.R.; Majka, S.M. Osteoblasts Derived from Induced Pluripotent Stem Cells Form Calcified Structures in Scaffolds Both In Vitro and In Vivo. Stem Cells 2011, 29, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Tashiro, K.; Inamura, M.; Kawabata, K.; Sakurai, F.; Yamanishi, K.; Hayakawa, T.; Mizuguchi, H. Efficient Adipocyte and Osteoblast Differentiation from Mouse Induced Pluripotent Stem Cells by Adenoviral Transduction. Stem Cells 2009, 27, 1802–1811. [Google Scholar] [CrossRef]
- Illich, D.J.; Demir, N.; Stojković, M.; Scheer, M.; Rothamel, D.; Neugebauer, J.; Hescheler, J.; Zöller, J.E. Concise Review: Induced Pluripotent Stem Cells and Lineage Reprogramming: Prospects for Bone Regeneration. Stem Cells 2011, 29, 555–563. [Google Scholar] [CrossRef]
- Ishimoto, T.; Nakano, T.; Umakoshi, Y.; Yamamoto, M.; Tabata, Y. Degree of Biological Apatite c-Axis Orientation Rather Than Bone Mineral Density Controls Mechanical Function in Bone Regenerated Using Recombinant Bone Morphogenetic protein-2. J. Bone Miner. Res. 2013, 28, 1170–1179. [Google Scholar] [CrossRef] [PubMed]
- Matsugaki, A.; Isobe, Y.; Saku, T.; Nakano, T. Quantitative Regulation of Bone-Mimetic, Oriented Collagen/Apatite Matrix Structure Depends on the Degree of Osteoblast Alignment on Oriented Collagen Substrates. J. Biomed. Mater. Res. A 2015, 103, 489–499. [Google Scholar] [CrossRef]
- Ozasa, R.; Matsugaki, A.; Isobe, Y.; Saku, T.; Yun, H.S.; Nakano, T. Construction of Human Induced Pluripotent Stem Cell-Derived Oriented Bone Matrix Microstructure by Using In Vitro Engineered Anisotropic Culture Model. J. Biomed. Mater. Res. A 2018, 106, 360–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiger, B.; Spatz, J.P.; Bershadsky, A.D. Environmental Sensing Through Focal Adhesions. Nat. Rev. Mol. Cell Biol. 2009, 10, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, N.J.; Jones, P.L. Extracellular Matrix and Integrin Signalling: The Shape of Things to Come. Biochem. J. 1999, 339, 481–488. [Google Scholar] [CrossRef]
- Matsugaki, A.; Aramoto, G.; Ninomiya, T.; Sawada, H.; Hata, S.; Nakano, T. Abnormal Arrangement of a Collagen/Apatite Extracellular Matrix Orthogonal to Osteoblast Alignment Is Constructed by a Nanoscale Periodic Surface Structure. Biomaterials 2015, 37, 134–143. [Google Scholar] [CrossRef]
- Nakanishi, Y.; Matsugaki, A.; Kawahara, K.; Ninomiya, T.; Sawada, H.; Nakano, T. Unique Arrangement of Bone Matrix Orthogonal to Osteoblast Alignment Controlled by Tspan11-Mediated Focal Adhesion Assembly. Biomaterials 2019, 209, 103–110. [Google Scholar] [CrossRef]
- Matsugaki, A.; Ozasa, R.; Isobe, Y.; Saku, T.; Nakano, T. Oriented Collagen Scaffolds for Anisotropic Bone Tissue Construction In Vitro. In Materials Science Forum; Trans Tech Publications Ltd.: Bäch SZ, Switzerland, 2014; pp. 1303–1306. [Google Scholar]
- Liu, G.; David, B.T.; Trawczynski, M.; Fessler, R.G. Advances in Pluripotent Stem Cells: History, Mechanisms, Technologies, and Applications. Stem Cell Rev. Rep. 2020, 16, 3–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballini, A.; Boccaccio, A.; Saini, R.; Pham, P.V.; Tatullo, M. Dental-Derived Stem Cells and Their Secretome and Interactions with Bioscaffolds/Biomaterials in Regenerative Medicine: From the In Vitro Research to Translational Applications. Stem Cell Int. 2017, 6975251. [Google Scholar] [CrossRef] [PubMed]
- Boccaccio, A.; Uva, A.E.; Fiorentino, M.; Monno, G.; Ballini, A.; Desiate, A. Optimal Load for Bone Tissue Scaffolds with An Assigned Geometry. Int. J. Med. Sci. 2018, 15, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Cantore, S.; Crincoli, V.; Boccaccio, A.; Uva, A.E.; Fiorentino, M.; Monno, G.; Bollero, P.; Derla, C.; Fabiano, F.; Ballini, A.; et al. Recent Advances in Endocrine, Metabolic and Immune Disorders: Mesenchymal Stem Cells (MSCs) and Engineered Scaffolds. Endocr. Metab. Immune. Disord. Drug Targets 2018, 18, 466–469. [Google Scholar] [CrossRef]
- Dang, Q.T.; Huynh, T.D.; Inchingolo, F.; Dipalma, G.; Inchingolo, A.D.; Cantore, S.; Paduanelli, G.; Nguyen, K.C.D.; Ballini, A.; Isacco, C.G.; et al. Human Chondrocytes from Human Adipose Tissue-Derived Mesenchymal Stem Cells Seeded on A Dermal-Derived Collagen Matrix Sheet: Our Preliminary Results for A Ready To Go Biotechnological Cartilage Graft in Clinical Practice. Stem Cell Int. 2021, 2021, 6664697. [Google Scholar]
- Matsugaki, A.; Aramoto, G.; Nakano, T. The Alignment of MC3T3-E1 Osteoblasts on Steps of Slip Traces Introduced by Dislocation Motion. Biomaterials 2012, 33, 7327–7335. [Google Scholar] [CrossRef]
- Matsugaki, A.; Yamazaki, D.; Nakano, T. Selective Patterning of netrin-1 as a Novel Guiding Cue for Anisotropic Dendrogenesis in Osteocytes. Mater. Sci. Eng. C 2020, 108, 110391. [Google Scholar] [CrossRef] [PubMed]
- Sugino, A.; Ohtsuki, C.; Tsuru, K.; Hayakawa, S.; Nakano, T.; Okazaki, Y.; Osaka, A. Effect of Spatial Design and Thermal Oxidation on Apatite Formation on Ti-15Zr-4Ta-4Nb Alloy. Acta Biomater. 2009, 5, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Ozasa, R.; Matsugaki, A.; Isobe, Y.; Saku, T.; Nakano, T. Directional Control of Mature Osteoblast Derived from Juvenile Mouse Calvariae. Mater. Trans. 2017, 58, 958–962. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Matsugaki, A.; Sekita, A.; Nakano, T. Alteration of Osteoblast Arrangement via Direct Attack by Cancer Cells: New Insights into Bone Metastasis. Sci. Rep. 2017, 7, 44824. [Google Scholar] [CrossRef] [Green Version]
- Matsugaki, A.; Harada, T.; Kimura, Y.; Sekita, A.; Nakano, T. Dynamic Collision Behavior Between Osteoblasts and Tumor Cells Regulates the Disordered Arrangement of Collagen Fiber/Apatite Crystals in Metastasized Bone. Int. J. Mol. Sci. 2018, 19, 3474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsugaki, A.; Fujiwara, N.; Nakano, T. Continuous Cyclic Stretch Induces Osteoblast Alignment and Formation of Anisotropic Collagen Fiber Matrix. Acta Biomater. 2013, 9, 7227–7235. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.J.; Boyde, A.; Pawley, J.B. Osteoblasts and Collagen Orientation. Cell Tissue Res. 1975, 159, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Baslé, M.F.; Grizon, F.; Pascaretti, C.; Lesourd, M.; Chappard, D. Shape and Orientation of Osteoblast-Like Cells (Saos-2) Are Influenced by Collagen Fibers in Xenogenic Bone Biomaterial. J. Biomed. Mater. Res. 1998, 40, 350–357. [Google Scholar] [CrossRef]
- Ishimoto, T.; Ozasa, R.; Nakano, K.; Weinmann, M.; Schnitter, C.; Stenzel, M.; Matsugaki, A.; Nagase, T.; Matsuzaka, T.; Todai, M.; et al. Development of TiNbTaZrMo Bio-High Entropy Alloy (BioHEA) Super-Solid Solution by Selective Laser Melting, and Its Improved Mechanical Property and Biocompatibility. Scr. Mater. 2021, 194, 113658. [Google Scholar] [CrossRef]
- Taubenberger, A.V.; Woodruff, M.A.; Bai, H.; Muller, D.J.; Hutmacher, D.W. The Effect of Unlocking RGD-Motifs in Collagen I on Pre-Osteoblast Adhesion and Differentiation. Biomaterials 2010, 31, 2827–2835. [Google Scholar] [CrossRef]
- Yamazaki, C.M.; Kadoya, Y.; Hozumi, K.; Okano-Kosugi, H.; Asada, S.; Kitagawa, K.; Nomizu, M.; Koide, T. A Collagen-Mimetic Triple Helical Supramolecule That Evokes Integrin-Dependent Cell Responses. Biomaterials 2010, 31, 1925–1934. [Google Scholar] [CrossRef]
- Tan, X.; Xu, A.; Zhao, T.; Zhao, Q.; Zhang, J.; Fan, C.; Deng, Y.; Freywald, A.; Genth, H.; Xiang, J. Simulated Microgravity Inhibits Cell Focal Adhesions Leading to Reduced Melanoma Cell Proliferation and Metastasis via FAK/RhoA-Regulated mTORC1 and AMPK Pathways. Sci. Rep. 2018, 8, 3769. [Google Scholar] [CrossRef] [Green Version]
- Changede, R.; Sheetz, M. Integrin and Cadherin Clusters: A Robust Way to Organize Adhesions for Cell Mechanics. BioEssays 2017, 39, 1–12. [Google Scholar] [CrossRef]
- Arnaout, M.A.; Goodman, S.L.; Xiong, J.P. Structure and Mechanics of Integrin-Based Cell Adhesion. Curr. Opin. Cell Biol. 2007, 19, 495–507. [Google Scholar] [CrossRef] [Green Version]
- Chiang, H.S.; Yang, R.S.; Huang, T.F. The Arg–Gly–Asp-Containing Peptide, Rhodostomin, Inhibits In Vitro Cell Adhesion to Extracellular Matrices and Platelet Aggregation Caused by Saos-2 Human Osteosarcoma Cells. Br. J. Cancer 1995, 71, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Fiore, V.F.; Wong, S.S.; Tran, C.; Tan, C.; Xu, W.; Sulchek, T.; White, E.S.; Hagood, J.S.; Barker, T.H. αvβ3 Integrin Drives Fibroblast Contraction and Strain Stiffening of Soft Provisional Matrix During Progressive Fibrosis. J.C.I. Insight 2018, 3, e97597. [Google Scholar] [CrossRef] [Green Version]
- Lobert, V.H.; Brech, A.; Pedersen, N.M.; Wesche, J.; Oppelt, A.; Malerød, L.; Stenmark, H. Ubiquitination of α5β1 Integrin Controls Fibroblast Migration Through Lysosomal Degradation of Fibronectin-Integrin Complexes. Dev. Cell 2010, 19, 148–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hozumi, K.; Teranishi, Y.; Enomoto, S.; Katagiri, F.; Kikkawa, Y.; Nomizu, M. Identification of Specific Integrin Cross-Talk for Dermal Fibroblast Cell Adhesion Using a Mixed Peptide-Chitosan Matrix. J. Biomater. Appl. 2019, 33, 893–902. [Google Scholar] [CrossRef]
- Ishimoto, T.; Sato, B.; Lee, J.W.; Nakano, T. Co-Deteriorations of Anisotropic Extracellular Matrix Arrangement and Intrinsic Mechanical Property in c-src Deficient Osteopetrotic Mouse Femur. Bone 2017, 103, 216–223. [Google Scholar] [CrossRef]
- Bishop, N. Bone Material Properties in Osteogenesis Imperfecta. J. Bone Miner. Res. 2016, 31, 699–708. [Google Scholar] [CrossRef] [Green Version]
- Ozasa, R.; Ishimoto, T.; Miyabe, S.; Hashimoto, J.; Hirao, M.; Yoshikawa, H.; Nakano, T. Osteoporosis Changes Collagen/Apatite Orientation and Young’s Modulus in Vertebral Cortical Bone of Rat. Calcif. Tissue Int. 2019, 104, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Matsugaki, A.; Ishimoto, T.; Nakano, T. Evaluation of Crystallographic Orientation of Biological Apatite in Vertebral Cortical Bone in Ovariectomized Cynomolgus Monkeys Treated with Minodronic Acid and Alendronate. J. Bone Miner. Metab. 2016, 34, 234–241. [Google Scholar] [CrossRef]
- Sekita, A.; Matsugaki, A.; Ishimoto, T.; Nakano, T. Synchronous Disruption of Anisotropic Arrangement of the Osteocyte Network and Collagen/Apatite in Melanoma Bone Metastasis. J. Struct. Biol. 2017, 197, 260–270. [Google Scholar] [CrossRef]
- Sekita, A.; Matsugaki, A.; Nakano, T. Disruption of Collagen/Apatite Alignment Impairs Bone Mechanical Function in Osteoblastic Metastasis Induced by Prostate Cancer. Bone 2017, 97, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Wakamatsu, T.; Iwasaki, Y.; Yamamoto, S.; Matsuo, K.; Goto, S.; Narita, I.; Kazama, J.J.; Tanaka, K.; Ito, A.; Ozasa, R.; et al. Type I Angiotensin II Receptor Blockade Reduces Uremia-Induced Deterioration of Bone Material Properties. J. Bone Miner. Res. 2021, 36, 67–79. [Google Scholar] [CrossRef]
- Ozasa, R.; Nakatsu, M.; Moriguchi, A.; Sasaki, K.; Ishimoto, T.; Okada, M.; Matsumoto, T.; Nakano, T. Analysis of Bone Regeneration Based on the Relationship Between the Orientations of Collagen and Apatite in Mouse Femur. Mater. Trans. 2020, 61, 381–386. [Google Scholar] [CrossRef] [Green Version]
- Kashii, M.; Hashimoto, J.; Nakano, T.; Umakoshi, Y.; Yoshikawa, H. Alendronate Treatment Promotes Bone Formation with a Less Anisotropic Microstructure During Intramembranous Ossification in Rats. J. Bone Miner. Metab. 2008, 26, 24–33. [Google Scholar] [CrossRef]
- Shiraishi, A.; Miyabe, S.; Nakano, T.; Umakoshi, Y.; Ito, M.; Mihara, M. The Combination Therapy with Alfacalcidol and Risedronate Improves the Mechanical Property in Lumbar Spine by Affecting the Material Properties in an Ovariectomized Rat Model of Osteoporosis. BMC Musculoskelet. Disord. 2009, 10, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsugaki, A.; Matsumoto, S.; Nakano, T. A Novel Role of Interleukin-6 as a Regulatory Factor of Inflammation-Associated Deterioration in Osteoblast Arrangement. Int. J. Mol. Sci. 2020, 21, 6659. [Google Scholar] [CrossRef] [PubMed]
- Aubin, J.E. Regulation of Osteoblast Formation and Function. Rev. Endocr. Metab. Disord. 2001, 2, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Price, P.A.; Poser, J.W.; Raman, N. Primary Structure of the γ Carboxyglutamic Acid Containing Protein from Bovine Bone. Proc. Natl. Acad. Sci. USA 1976, 73, 3374–3375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauschka, P.V.; Carr, S.A. Calcium-Dependent α-Helical Structure in Osteocalcin. Biochemistry 1982, 21, 2538–2547. [Google Scholar] [CrossRef] [PubMed]
- Hauschka, P.V.; Wians, F.H., Jr. Osteocalcin-Hydroxyapatite Interaction in the Extracellular Organic Matrix of Bone. Anat. Rec. 1989, 224, 180–188. [Google Scholar] [CrossRef]
- Chen, L.; Jacquet, R.; Lowder, E.; Landis, W.J. Refinement of Collagen-Mineral Interaction: A Possible Role for Osteocalcin in Apatite Crystal Nucleation, Growth and Development. Bone 2015, 71, 7–16. [Google Scholar] [CrossRef]
- Price, P.A.; Williamson, M.K.; Lothringer, J.W. Origin of the Vitamin K-Dependent Bone Protein Found in Plasma and Its Clearance by Kidney and Bone. J. Biol. Chem. 1981, 256, 12760–12766. [Google Scholar] [CrossRef]
- Moriishi, T.; Ozasa, R.; Ishimoto, T.; Nakano, T.; Hasegawa, T.; Miyazaki, T.; Liu, W.; Fukuyama, R.; Wang, Y.; Komori, H.; et al. Osteocalcin Is Necessary for the Alignment of Apatite Crystallites, but Not Glucose Metabolism, Testosterone Synthesis, or Muscle Mass. PLoS Genet. 2020, 16, 1008586. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.; Quintanilla, R.H.; Grecian, S.; Gee, K.R.; Rao, M.S.; Lakshmipathy, U. Novel live alkaline phosphatase substrate for identification of pluripotent stem cells. Stem Cell Rev. 2012, 8, 1021–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shuzui, E.; Kim, M.-H.; Azuma, K.; Fujinaga, Y.; Kino-oka, M. Maintenance of an undifferentiated state of human-induced pluripotent stem cells through botulinum hemagglutinin-mediated regulation of cell behavior. J. Biosci. Bioeng. 2019, 127, 744–751. [Google Scholar] [CrossRef]
- Villa-Diaz, L.G.; Brown, S.E.; Liu, Y.; Ross, A.M.; Lahann, J.; Parent, J.M.; Krebsbach, P.H. Derivation of Mesenchymal Stem Cells from Human Induced Pluripotent Stem Cells Cultured on Synthetic Substrates. Stem Cells 2012, 30, 1174–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, S.E.; Tong, W.; Krebsbach, P.H. The Derivation of Mesenchymal Stem Cells from Human Embryonic Stem Cells. Cells Tissues Organs 2009, 189, 256–260. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozasa, R.; Matsugaki, A.; Matsuzaka, T.; Ishimoto, T.; Yun, H.-S.; Nakano, T. Superior Alignment of Human iPSC-Osteoblasts Associated with Focal Adhesion Formation Stimulated by Oriented Collagen Scaffold. Int. J. Mol. Sci. 2021, 22, 6232. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126232
Ozasa R, Matsugaki A, Matsuzaka T, Ishimoto T, Yun H-S, Nakano T. Superior Alignment of Human iPSC-Osteoblasts Associated with Focal Adhesion Formation Stimulated by Oriented Collagen Scaffold. International Journal of Molecular Sciences. 2021; 22(12):6232. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126232
Chicago/Turabian StyleOzasa, Ryosuke, Aira Matsugaki, Tadaaki Matsuzaka, Takuya Ishimoto, Hui-Suk Yun, and Takayoshi Nakano. 2021. "Superior Alignment of Human iPSC-Osteoblasts Associated with Focal Adhesion Formation Stimulated by Oriented Collagen Scaffold" International Journal of Molecular Sciences 22, no. 12: 6232. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126232