
Screen Key Genes Associated with Distraction-Induced Osteogenesis of Stem Cells Using Bioinformatics Methods



Abstract
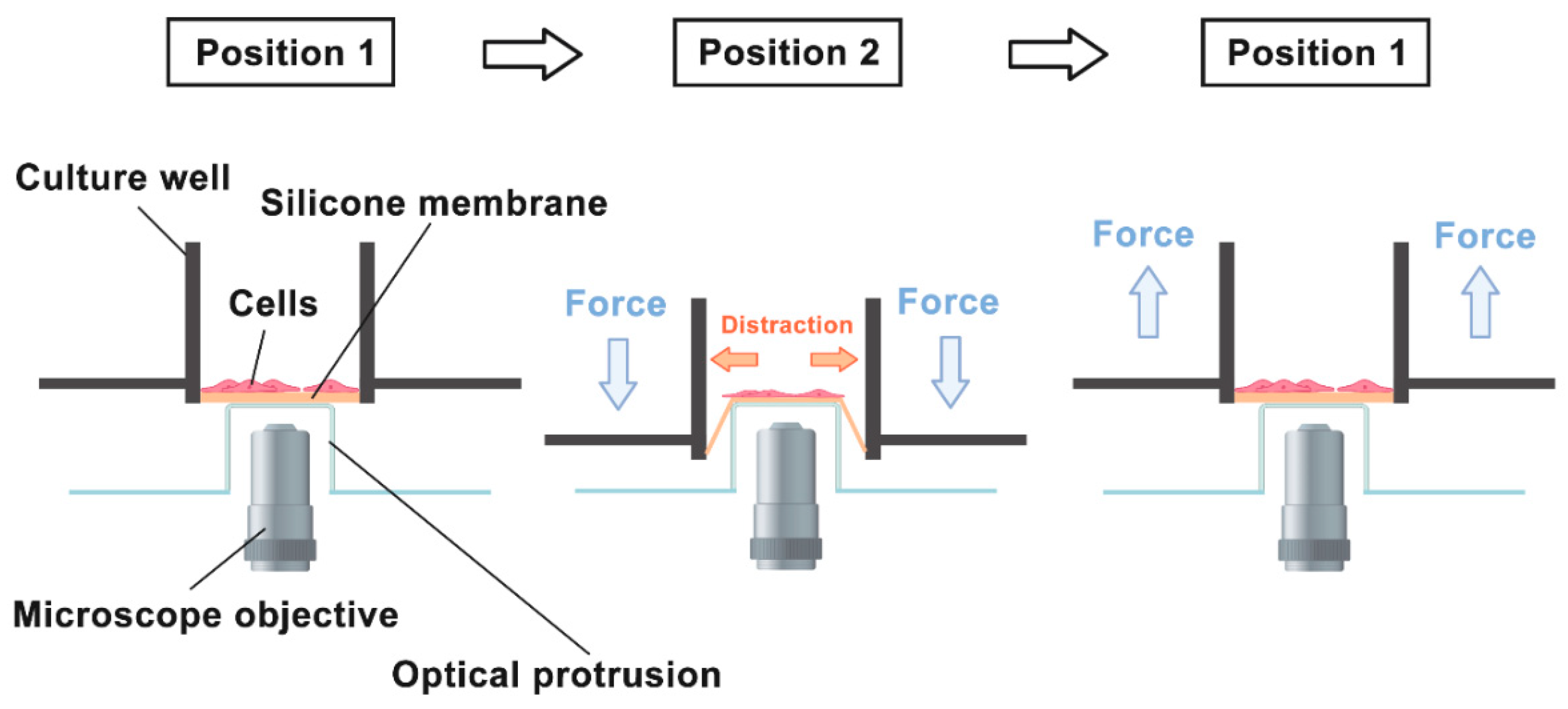
:1. Introduction
2. Results
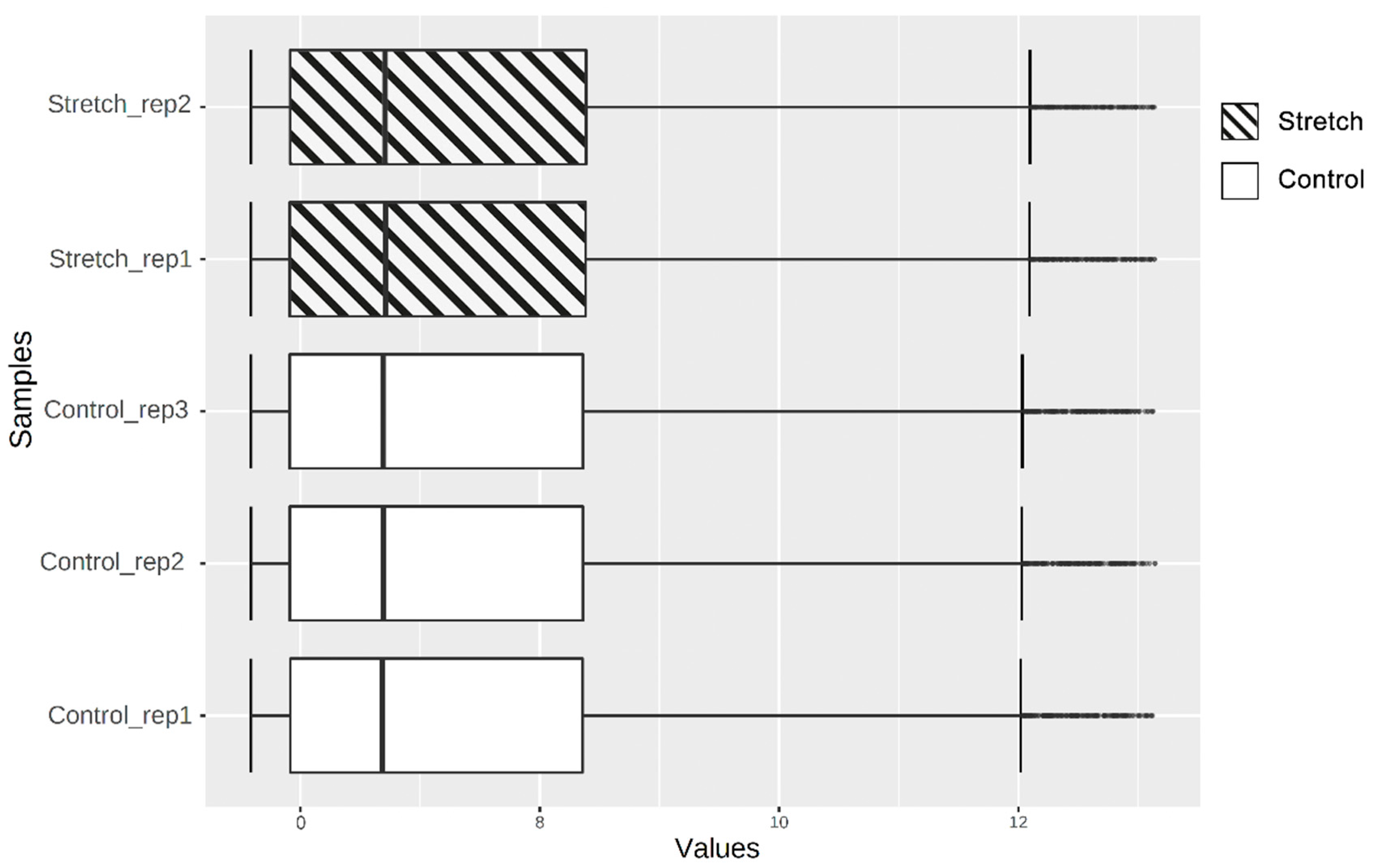
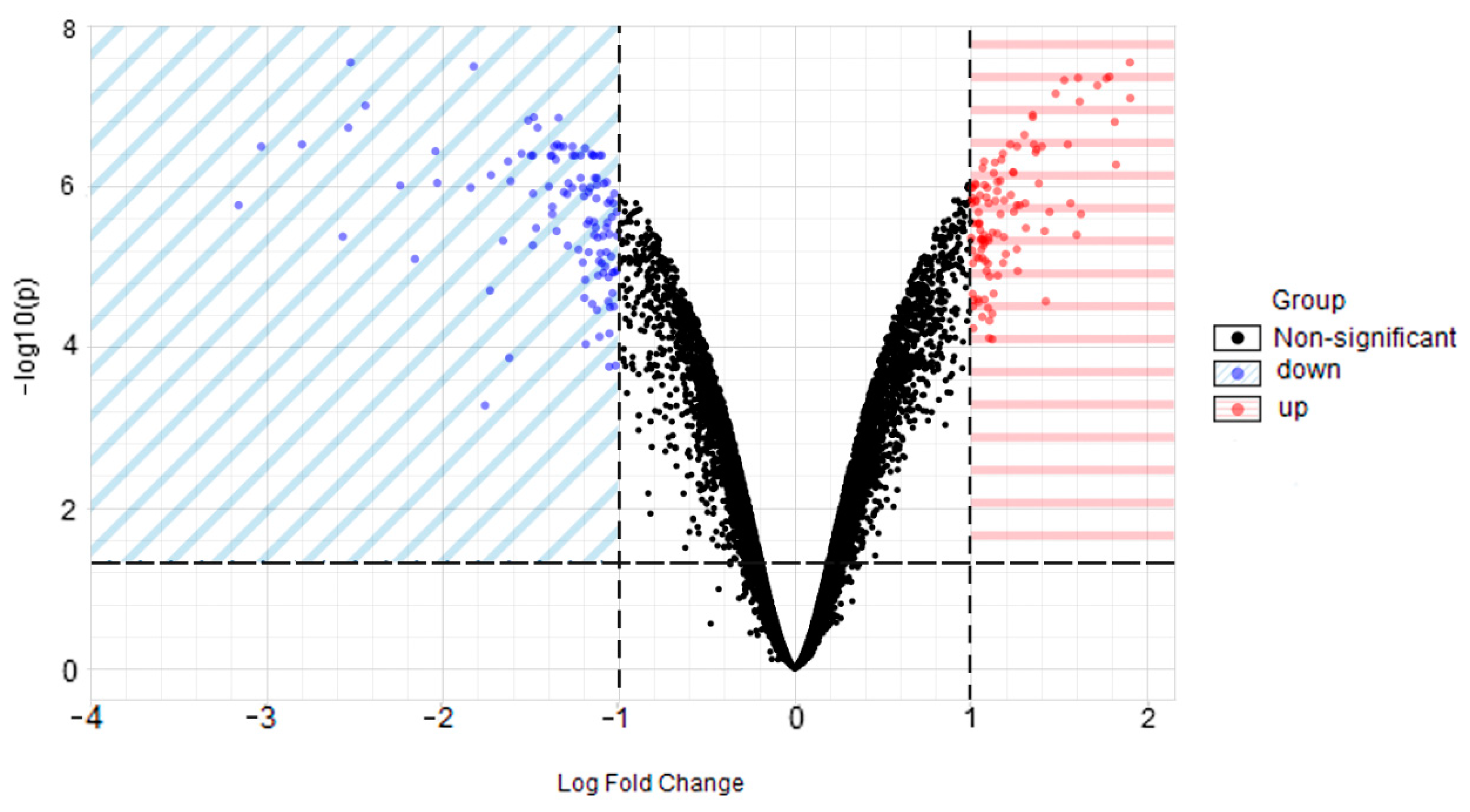
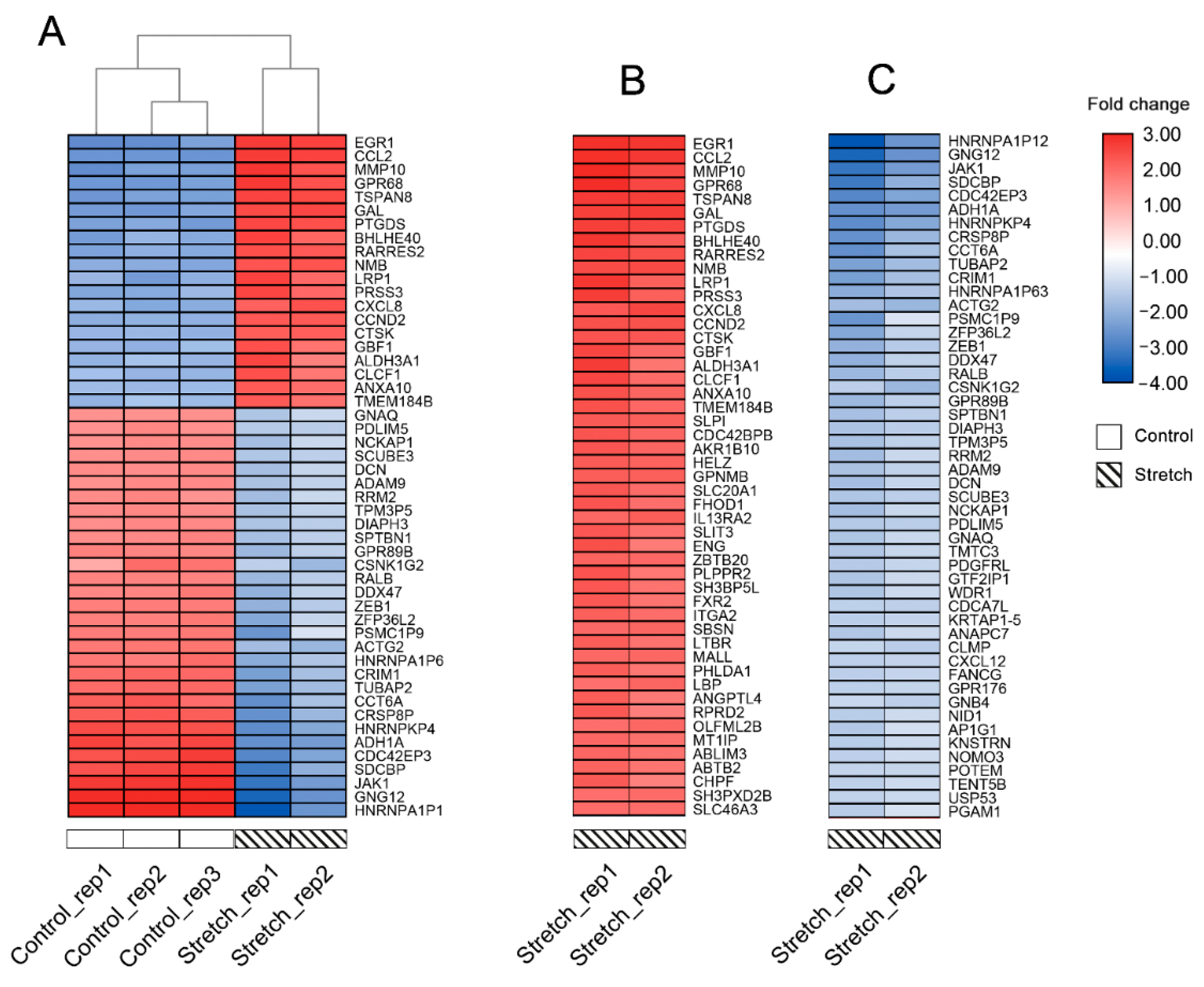
2.1. Identification of DEGs
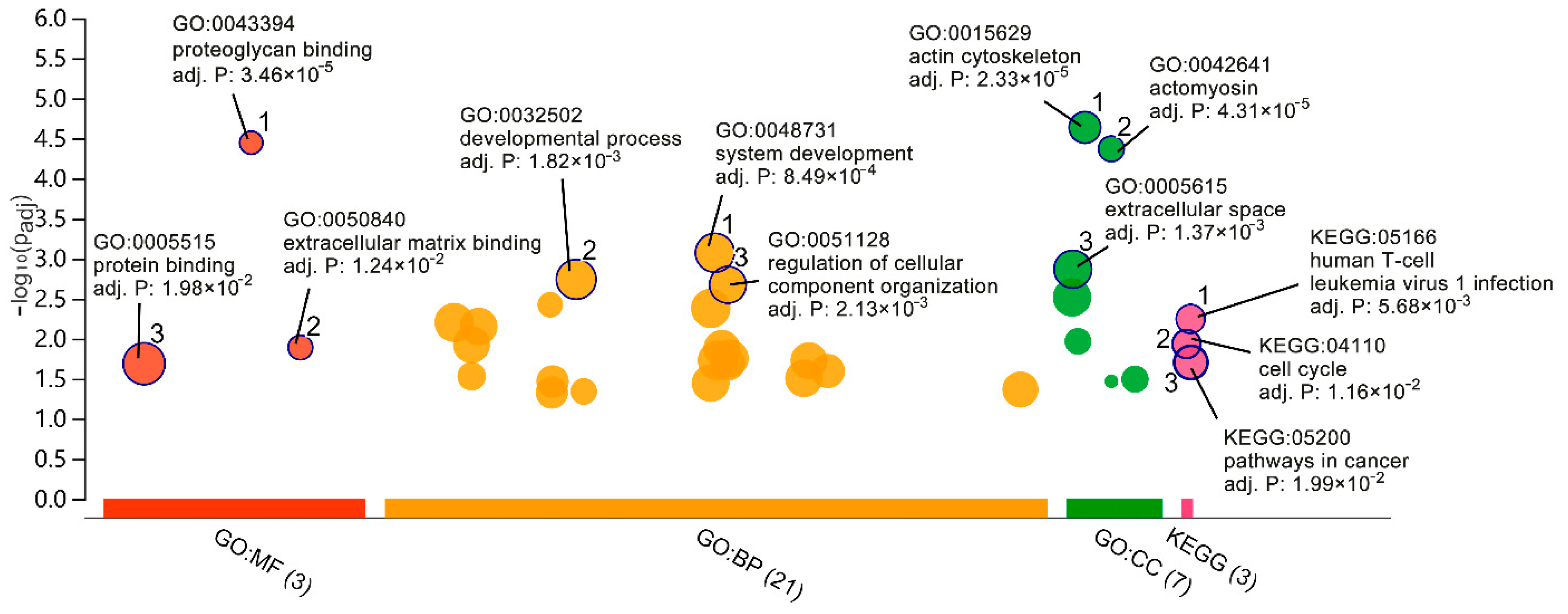
2.2. GO and Pathway Enrichment Analyses
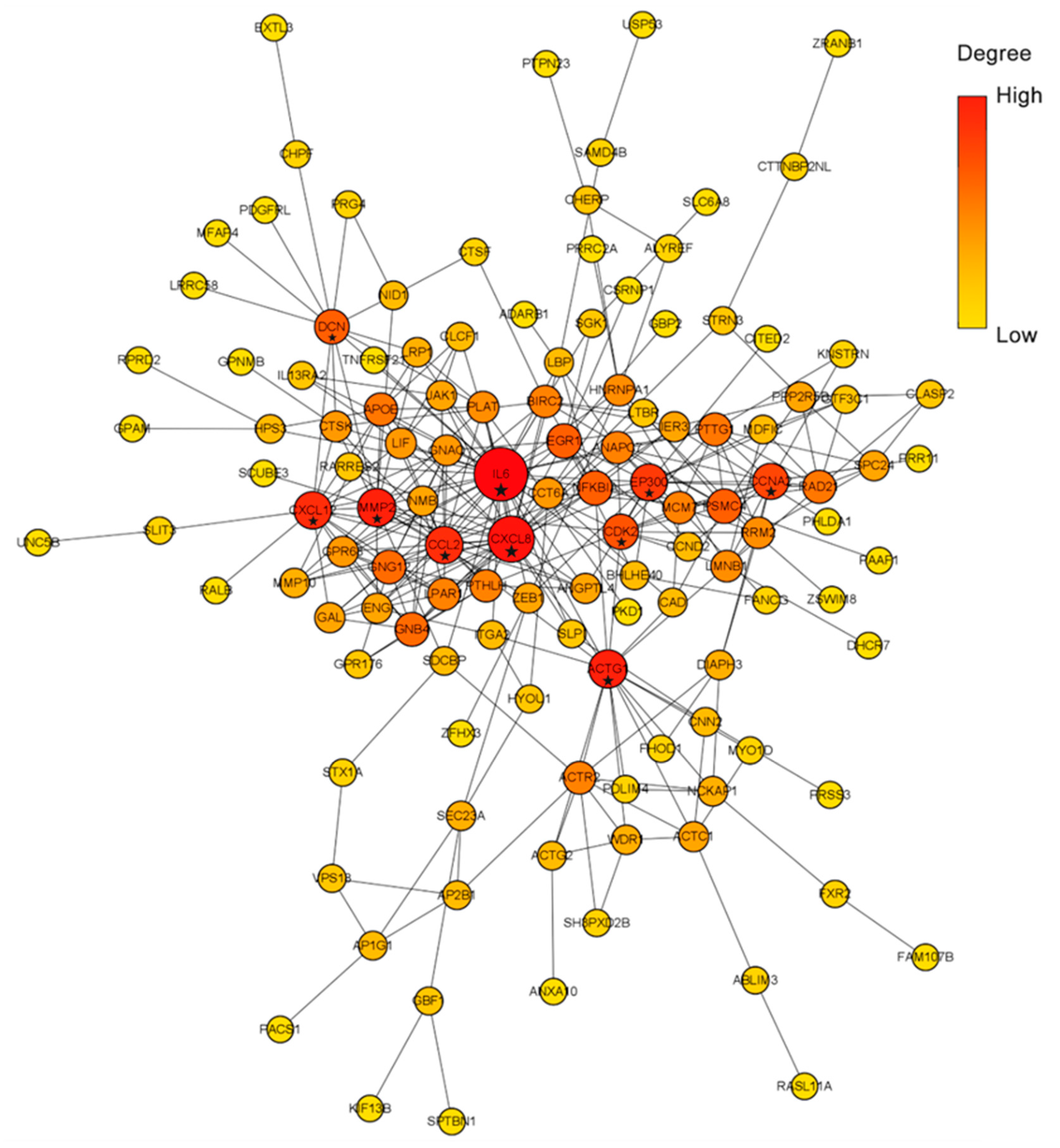
2.3. PPI Network Construction
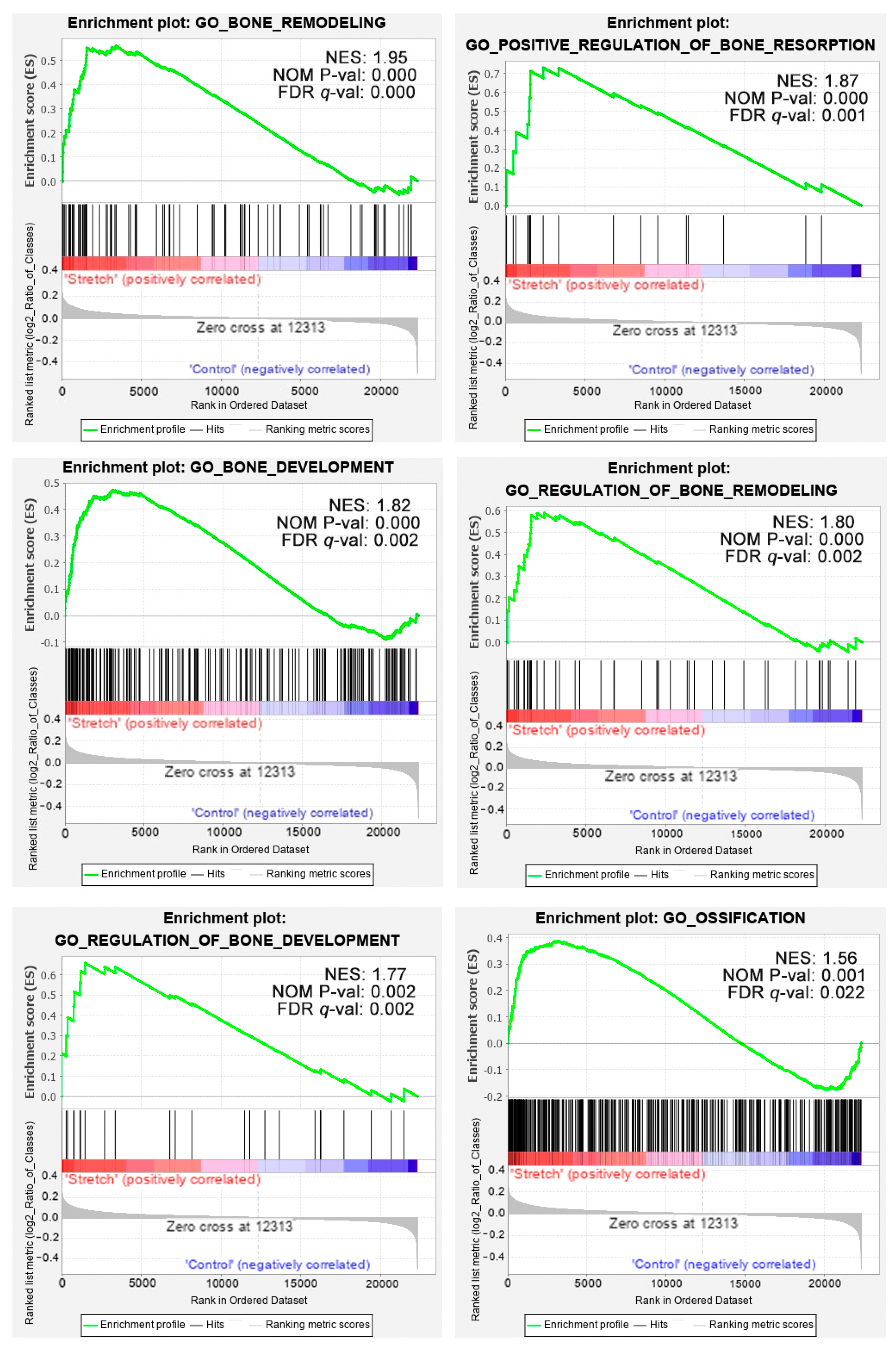
2.4. Gene Set Enrichment Analysis
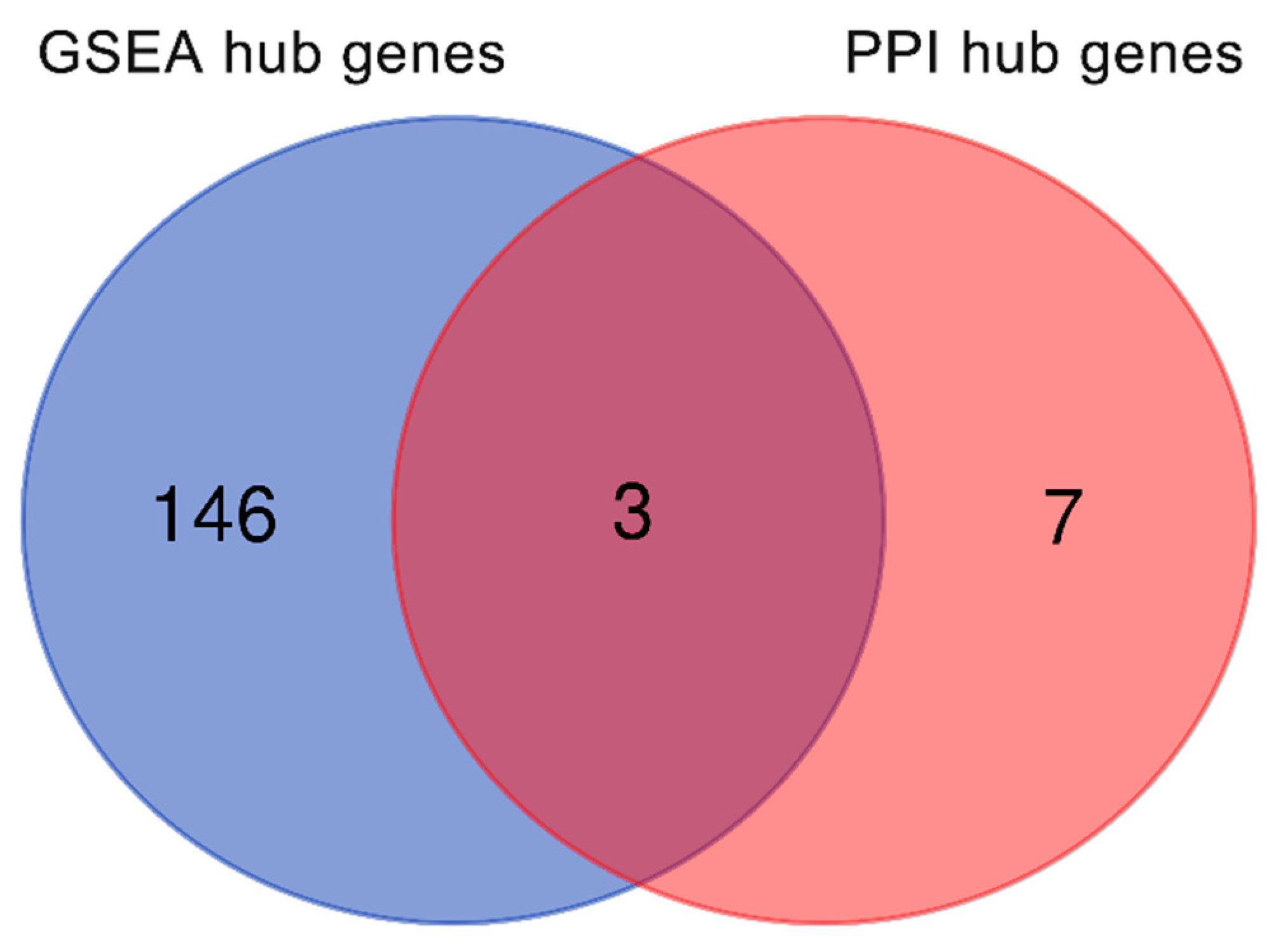
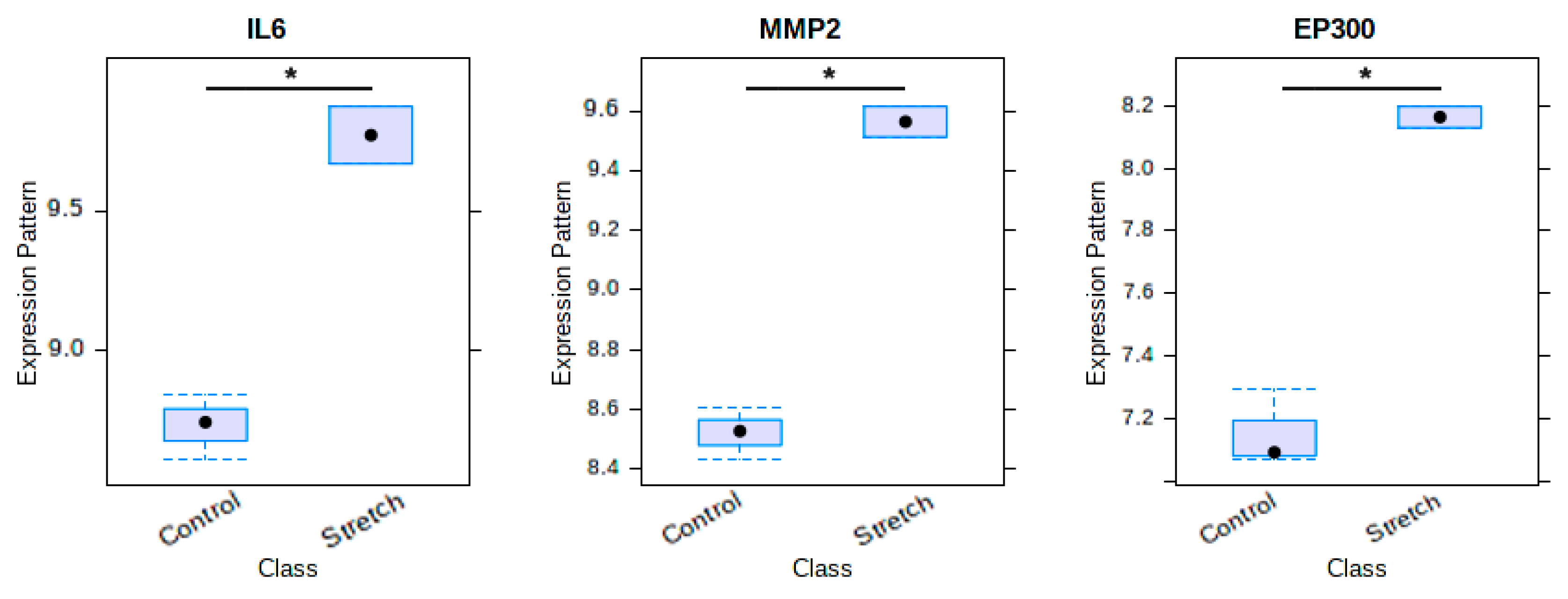
2.5. Venn Diagram of Osteogenic Hub Genes
3. Discussion
4. Materials and Methods
4.1. Microarray Data Information
4.2. Identification of DEGs
4.3. GO and Pathway Enrichment Analyses
4.4. PPI Network Construction
4.5. Gene Set Enrichment Analysis
4.6. Venn Diagram of Osteogenic Hub Genes
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Informations | Data |
|---|---|
| Donor Information | |
| Age | 21 yo |
| Race | Black |
| Gender | Male |
| Cell Information | |
| Cell type | Human Bone Marrow Mesenchymal Stem Cell |
| Frozen Date | 11 April 2006 |
| Cell Passage | 2 |
| Number of cells | ≥750,000 cells/0.5 mL–1,530,000 cells in total |
| Vitality-Trypan Blue | ≥75% |
| Virus Tests | |
| HIV | Negative |
| HBV | Negative |
| HCV | Negative |
| Adipogenic Analysis | |
| Oil Red O Method | Positive |
| Chondrogenic Analysis | |
| Proteoglycan–Saffron Staining | Positive |
| Type II Collagen (on 14th and 21st days) | Positive |
| Osteogenic Analysis | |
| Calcium Deposition | Positive |
| Markers | |
| CD105, CD166, CD29, CD44 | 90% Positive |
| CD14, CD34, CD45 | <5% Positive |
Appendix B
| Collection | Gene Set | Size |
|---|---|---|
| C5: GO BP | GO_DIRECT_OSSIFICATION | 6 |
| C5: GO BP | GO_BONE_REMODELING | 91 |
| C5: GO BP | GO_BONE_MINERALIZATION | 112 |
| C5: GO BP | GO_BONE_MATURATION | 22 |
| C5: GO BP | GO_BONE_GROWTH | 47 |
| C5: GO BP | GO_BONE_DEVELOPMENT | 219 |
| C5: GO BP | GO_OSSIFICATION | 396 |
| C5: GO BP | GO_OSSIFICATION_INVOLVED_IN_BONE_REMODELING | 5 |
| C5: GO BP | GO_POSITIVE_REGULATION_OF_BONE_DEVELOPMENT | 9 |
| C5: GO BP | GO_POSITIVE_REGULATION_OF_BONE_MINERALIZATION | 39 |
| C5: GO BP | GO_POSITIVE_REGULATION_OF_BONE_RESORPTION | 19 |
| C5: GO BP | GO_POSITIVE_REGULATION_OF_CELL_PROLIFERATION_IN_BONE_MARROW | 8 |
| C5: GO BP | GO_POSITIVE_REGULATION_OF_OSSIFICATION | 88 |
| C5: GO BP | GO_REGULATION_OF_BONE_DEVELOPMENT | 24 |
| C5: GO BP | GO_REGULATION_OF_BONE_MINERALIZATION | 74 |
| C5: GO BP | GO_REGULATION_OF_BONE_MINERALIZATION_INVOLVED_IN_BONE_MATURATION | 5 |
| C5: GO BP | GO_REGULATION_OF_BONE_REMODELING | 48 |
| C5: GO BP | GO_REGULATION_OF_OSSIFICATION | 200 |
| C5: GO BP | GO_REPLACEMENT_OSSIFICATION | 28 |
Appendix C
| Software/Website | Website Address |
|---|---|
| ArrayExpress database | https://www.ebi.ac.uk/arrayexpress/ (accessed on 27 June 2020) |
| NetworkAnalyst 3.0 | https://www.networkanalyst.ca (accessed on 28 June 2020) |
| g:Profiler | http://biit.cs.ut.ee/gprofiler/ (accessed on 1 July 2020) |
| STRING database | https://string-db.org (accessed on 5 July 2020) |
| Cytoscape software (version 3.8.0) | https://cytoscape.org (accessed on 5 July 2020) |
| Gene set enrichment analysis software (version 4.0.3) | https://www.gsea-msigdb.org/gsea/downloads.jsp (accessed on 12 July 2020) |
| Molecular Signatures Database | https://www.gsea-msigdb.org/gsea/msigdb/index.jsp (accessed on 12 July 2020) |
| Venn diagram | http://bioinformatics.psb.ugent.be/webtools/Venn/ (accessed on 17 July 2020) |
| SPSS Statistics 22.0 | https://www.ibm.com/support/pages/downloading-ibm-spss-statistics-22 (accessed on 23 July 2020) |
References
- Guerado, E.C.E.; Caso, E. Challenges of bone tissue engineering in orthopaedic patients. World J. Orthop. 2017, 8, 87–98. [Google Scholar] [CrossRef]
- Borzunov, D.Y.; Kolchin, S.N.; Malkova, T.A. Role of the Ilizarov non-free bone plasty in the management of long bone defects and nonunion: Problems solved and unsolved. World J. Orthop. 2020, 11, 304–318. [Google Scholar] [CrossRef]
- Gubin, A.V.; Borzunov, D.Y.; Marchenkova, L.O.; Malkova, T.A.; ISmirnova, L. Contribution of G.A. Ilizarov to bone reconstruction: Historical achievements and state of the art. Strateg. Trauma Limb Reconstr. 2016, 11, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Xu, J.; Xu, L.; Zhang, J.; Chan, K.; Pan, X.; Li, G. MiR-503 Promotes Bone Formation in Distraction Osteogenesis through Suppressing Smurf1 Expression. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Morillo, C.M.R.; Sloniak, M.C.; Gonçalves, F.; Villar, C. Efficacy of stem cells on bone consolidation of distraction osteogenesis in animal models: A systematic review. Braz. Oral Res. 2018, 32, e83. [Google Scholar] [CrossRef]
- Grayson, W.L.; Bunnell, B.A.; Martin, E.; Frazier, T.; Hung, B.P.; Gimble, J.M. Stromal cells and stem cells in clinical bone regeneration. Nat. Rev. Endocrinol. 2015, 11, 140–150. [Google Scholar] [CrossRef]
- Jones, E.A.; Giannoudis, P.V.; Kouroupis, D. Bone repair with skeletal stem cells: Rationale, progress to date and clinical application. Ther. Adv. Musculoskelet. Dis. 2016, 8, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Robey, P.G. “Mesenchymal stem cells”: Fact or fiction, and implications in their therapeutic use. F1000Research 2017, 6, 524. [Google Scholar] [CrossRef] [Green Version]
- Montes-Medina, L.; Hernández-Fernández, A.; Gutiérrez-Rivera, A.; Ripalda-Cemboráin, P.; Bitarte, N.; Pérez-López, V.; Granero-Moltó, F.; Prosper, F.; Izeta, A. Effect of bone marrow stromal cells in combination with biomaterials in early phases of distraction osteogenesis: An experimental study in a rabbit femur model. Injury 2018, 49, 1979–1986. [Google Scholar] [CrossRef]
- Yang, Y.; Lin, S.; Wang, B.; Gu, W.; Li, G. Stem cell therapy for enhancement of bone consolidation in distraction osteogenesis: A contemporary review of experimental studies. Bone Joint Res. 2017, 66, 385–390. [Google Scholar] [CrossRef] [Green Version]
- Gessmann, J.; Köller, M.; Godry, H.; Schildhauer, T.A.; Seybold, D. Regenerate augmentation with bone marrow concentrate after traumatic bone loss. Orthop. Rev. 2011, 3, e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoey, D.A.; Tormey, S.; Ramcharan, S.; O’Brien, F.J.; Jacobs, C.R. Primary cilia-mediated mechanotransduction in human mesenchymal stem cells. Stem Cells 2012, 30, 2561–2570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saidova, A.A.; Vorobjev, I.A. Lineage Commitment, Signaling Pathways, and the Cytoskeleton Systems in Mesenchymal Stem Cells. Tissue Eng. Part B Rev. 2020, 26, 13–25. [Google Scholar] [CrossRef]
- Ai-Aql, Z.S.; Alagl, A.S.; Graves, D.T.; Gerstenfeld, L.C.; Einhorn, T.A. Molecular mechanisms controlling bone formation during fracture healing and distraction osteogenesis. J. Dent. Res. 2008, 87, 107–118. [Google Scholar] [CrossRef]
- Athar, A.; Füllgrabe, A.; George, N.; Iqbal, H.; Huerta, L.; Ali, A.; Snow, C.; Fonseca, N.A.; Petryszak, R.; Papatheodorou, I.; et al. ArrayExpress update–from bulk to single-cell expression data. Nucleic Acids Res. 2019, 47, D711–D715. [Google Scholar] [CrossRef] [PubMed]
- Fang, B.; Liu, Y.; Zheng, D.; Shan, S.; Wang, C.; Gao, Y.; Wang, J.; Xie, Y.; Zhang, Y.; Li, Q. The effects of mechanical stretch on the biological characteristics of human adipose-derived stem cells. J. Cell. Mol. Med. 2019, 23, 4244–4255. [Google Scholar] [CrossRef] [Green Version]
- Bao, G.; Suresh, S. Cell and molecular mechanics of biological materials. Nat. Mater. 2003, 2, 715–725. [Google Scholar] [CrossRef]
- Zhu, C.; Bao, G.; Wang, N. Cell mechanics: Mechanical response, cell adhesion, and molecular deformation. Annu. Rev. Biomed. Eng. 2000, 2, 189–226. [Google Scholar] [CrossRef] [PubMed]
- Burridge, K.; Fath, K.; Kelly, T.; Nuckolls, G.; Turner, C. Focal Adhesions: Transmembrane Junctions between the Extracellular Matrix and the Cytoskeleton. Annu. Rev. Cell Biol. 1988, 4, 487–525. [Google Scholar] [CrossRef]
- Ingber, D. Integrins as mechanochemical transducers. Curr. Opin. Cell Biol. 1991, 3, 841–848. [Google Scholar] [CrossRef]
- Morgan, E.F.; Longaker, M.T.; Carter, D.R. Relationships between tissue dilatation and differentiation in distraction osteogenesis. Matrix Biol. 2006, 25, 94–103. [Google Scholar] [CrossRef] [Green Version]
- Ransom, R.C.; Carter, A.C.; Salhotra, A.; Leavitt, T.; Marecic, O.; Murphy, M.P.; Lopez, M.L.; Wei, Y.; Marshall, C.D.; Shen, E.Z.; et al. Mechanoresponsive stem cells acquire neural crest fate in jaw regeneration. Nat. Cell Biol. 2018, 563, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Kon, T.; Cho, T.J.; Aizawa, T.; Yamazaki, M.; Nooh, N.; Graves, D.; Gerstenfeld, L.C.; Einhorn, T.A. Expression of osteoprotegerin, receptor activator of NF-kappaB ligand (osteoprotegerin ligand) and related proinflammatory cytokines during fracture healing. J. Bone Miner. Res. 2001, 16, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Cho, T.-J.; Kim, J.A.; Chung, C.Y.; Yoo, W.J.; Gerstenfeld, L.C.; Einhorn, T.A.; Choi, I.H. Expression and Role of Interleukin-6 in Distraction Osteogenesis. Calcif. Tissue Int. 2007, 80, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Cillo, J.E.; Gassner, R.; Koepsel, R.R.; Buckley, M.J. Growth factor and cytokine gene expression in mechanically strained human osteoblast-like cells: Implications for distraction osteogenesis. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontol. 2000, 90, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.-E.; Lee, S.Y. IL-6 is produced by adipose-derived stromal cells and promotes osteogenesis. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2013, 1833, 2608–2616. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; Wang, L.; Zhao, Y.; Cao, J.; Wang, T.; Liu, P.; Zhang, Y.; Yang, X.; Cheng, X.; Liu, B.; et al. Sympathetic denervation-induced MSC mobilization in distraction osteogenesis associates with inhibition of MSC migration and osteogenesis by norepinephrine/adrb3. PLoS ONE 2014, 9, e105976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mott, J.D.; Werb, Z. Regulation of matrix biology by matrix metalloproteinases. Curr. Opin. Cell Biol. 2004, 16, 558–564. [Google Scholar] [CrossRef] [Green Version]
- Karadag, A.; Fisher, L.W. Bone sialoprotein enhances migration of bone marrow stromal cells through matrices by bridging MMP-2 to alpha(v)beta3-integrin. J. Bone Miner. Res. 2006, 21, 1627–1636. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wu, B.; Jia, S.; Zhao, Y.; Hou, R.; Liu, X.; Wang, X.; Chen, L.; Yang, X.; Lei, D.; et al. The mechanically activated p38/MMP-2 signaling pathway promotes bone marrow mesenchymal stem cell migration in rats. Arch. Oral Biol. 2017, 76, 55–60. [Google Scholar] [CrossRef]
- Almalki, S.G.; Agrawal, D.K. Effects of matrix metalloproteinases on the fate of mesenchymal stem cells. Stem Cell Res. Ther. 2016, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kasper, G.; Glaeser, J.D.; Geißler, S.; Ode, A.; Tuischer, J.; Matziolis, G.; Perka, C.; Duda, G.N. Matrix Metalloprotease Activity Is an Essential Link Between Mechanical Stimulus and Mesenchymal Stem Cell Behavior. Stem Cells 2007, 25, 1985–1994. [Google Scholar] [CrossRef]
- Hu, Z.; Wang, Y.; Sun, Z.; Wang, H.; Zhou, H.; Zhang, L.; Zhang, S.; Cao, X. miRNA-132-3p inhibits osteoblast differentiation by targeting Ep300 in simulated microgravity. Sci. Rep. 2015, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Hu, Z.; Shi, F.; Dong, J.; Dang, L.; Wang, Y.; Sun, Z.; Zhou, H.; Zhang, S.; Cao, X.; et al. Osteoblast-targeted delivery of miR-33-5p attenuates osteopenia development induced by mechanical unloading in mice. Cell Death Dis. 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Zhang, L.; Wang, H.; Wang, Y.; Tan, Y.; Dang, L.; Wang, K.; Sun, Z.; Li, G.; Cao, X.; et al. Targeted silencing of miRNA-132-3p expression rescues disuse osteopenia by promoting mesenchymal stem cell osteogenic differentiation and osteogenesis in mice. Stem Cell Res. Ther. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Zhou, G.; Soufan, O.; Ewald, J.; Hancock, R.E.W.; Basu, N.; Xia, J. NetworkAnalyst 3.0: A visual analytics platform for comprehensive gene expression profiling and meta-analysis. Nucleic Acids Res. 2019, 47, W234–W241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, L.J.; Kuhn, M.; Stark, M.; Chaffron, S.; Creevey, C.; Muller, J.; Doerks, T.; Julien, P.; Roth, A.; Simonovic, M.; et al. STRING 8—A global view on proteins and their functional interactions in 630 organisms. Nucleic Acids Res. 2009, 37 (Suppl. S1), D412–D416. [Google Scholar] [CrossRef] [PubMed]









| Category | GO ID | Description | Gene Count | Adjusted p-Value |
|---|---|---|---|---|
| BP | GO:0048731 | system development | 83 | 8.49 × 10−4 |
| BP | GO:0032502 | developmental process | 100 | 1.82 × 10−3 |
| BP | GO:0051128 | regulation of cellular component organisation | 51 | 2.13 × 10−3 |
| BP | GO:0022617 | extracellular matrix disassembly | 8 | 3.78 × 10−3 |
| BP | GO:0048518 | positive regulation of biological process | 96 | 4.18 × 10−3 |
| CC | GO:0015629 | actin cytoskeleton | 21 | 2.33 × 10−5 |
| CC | GO:0042641 | actomyosin mesenchymal transition | 9 | 4.31 × 10−5 |
| CC | GO:0005615 | extracellular space | 62 | 1.37 × 10−3 |
| CC | GO:0005576 | extracellular region | 73 | 3.08 × 10−3 |
| CC | GO:0005884 | actin filament | 8 | 1.08 × 10−2 |
| MF | GO:0043394 | proteoglycan binding | 7 | 3.46 × 10−5 |
| MF | GO:0050840 | extracellular matrix binding | 6 | 1.24 × 10−2 |
| MF | GO:0005515 | protein binding | 156 | 1.98 × 10−2 |
| KEGG | KEGG:05166 | human T-cell leukaemia virus 1 infection | 11 | 5.68 × 10−3 |
| KEGG | KEGG:04110 | cell cycle | 8 | 1.16 × 10−2 |
| KEGG | KEGG:05200 | pathways in cancer | 17 | 1.99 × 10−2 |
| Rank | Gene Symbol | Degree |
|---|---|---|
| 1 | IL6 | 38 |
| 2 | CXCL8 | 28 |
| 3 | MMP2 | 17 |
| 3 | ACTG1 | 17 |
| 5 | CCL2 | 16 |
| 5 | CXCL12 | 16 |
| 7 | EP300 | 15 |
| 8 | CCNA2 | 14 |
| 9 | CDK2 | 13 |
| 10 | DCN | 12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Hua, J.; Song, W. Screen Key Genes Associated with Distraction-Induced Osteogenesis of Stem Cells Using Bioinformatics Methods. Int. J. Mol. Sci. 2021, 22, 6505. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126505
Chen J, Hua J, Song W. Screen Key Genes Associated with Distraction-Induced Osteogenesis of Stem Cells Using Bioinformatics Methods. International Journal of Molecular Sciences. 2021; 22(12):6505. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126505
Chicago/Turabian StyleChen, Jishizhan, Jia Hua, and Wenhui Song. 2021. "Screen Key Genes Associated with Distraction-Induced Osteogenesis of Stem Cells Using Bioinformatics Methods" International Journal of Molecular Sciences 22, no. 12: 6505. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126505