
Genome-Wide Identification, Diversification, and Expression Analysis of Lectin Receptor-Like Kinase (LecRLK) Gene Family in Cucumber under Biotic Stress

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Identification and Physicochemical Properties of CsaLecRLK Genes
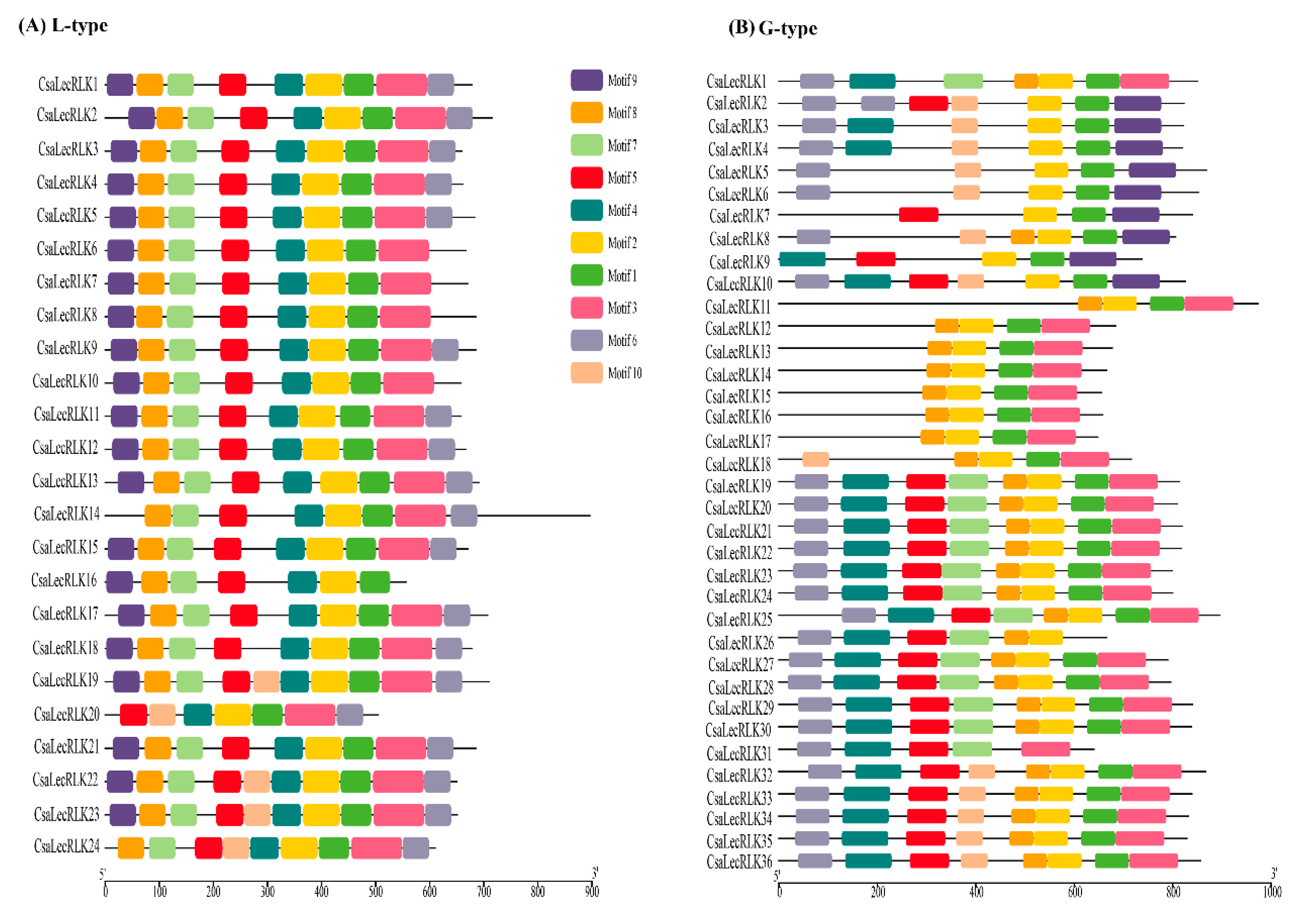
2.2. Phylogenetic Relationship, Motif Composition and Gene Structure Analysis of CsaLecRLKs Genes
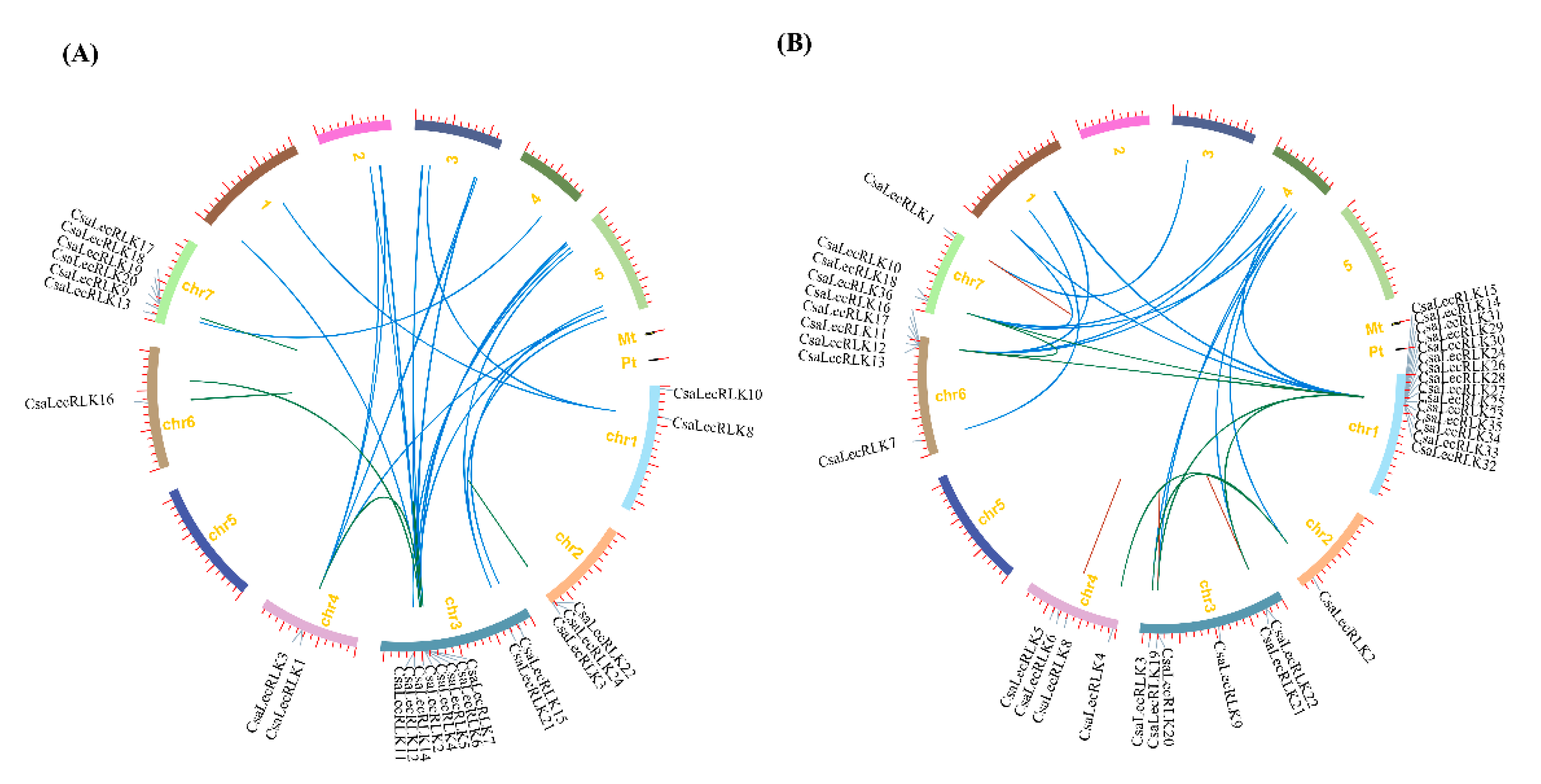
2.3. Chromosomal Location and Gene Duplication Analysis of CsaLecRLK Genes
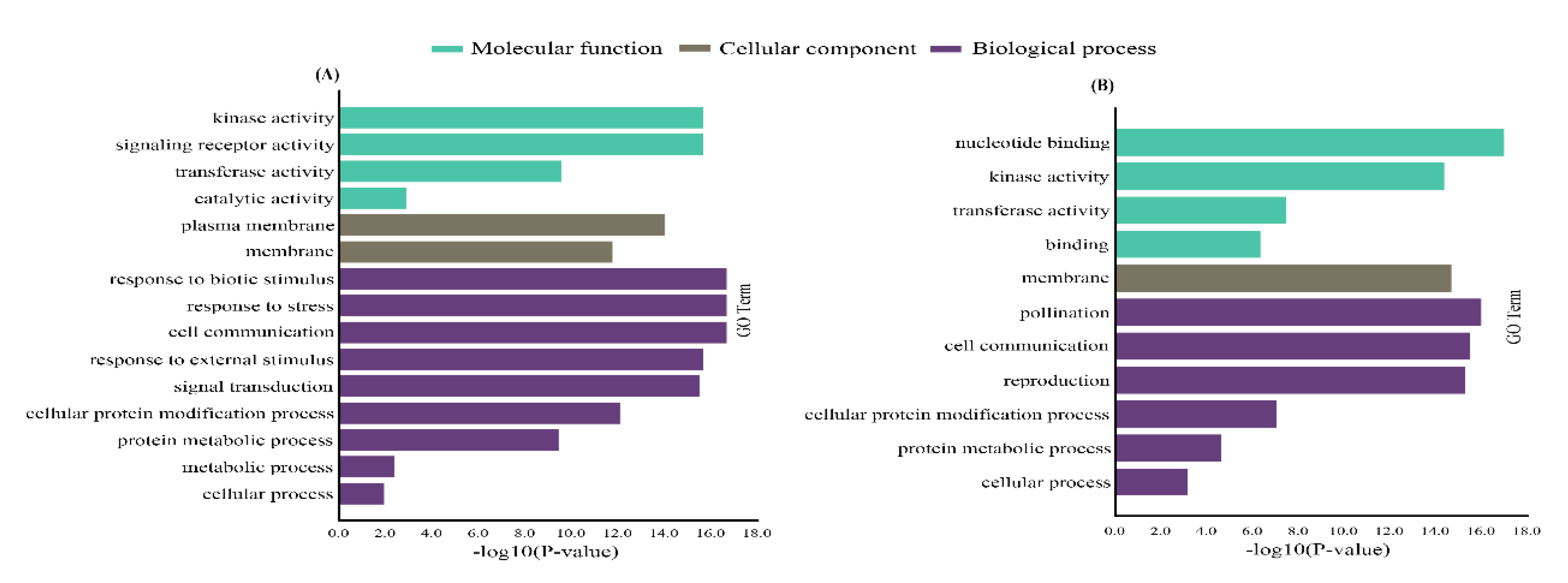
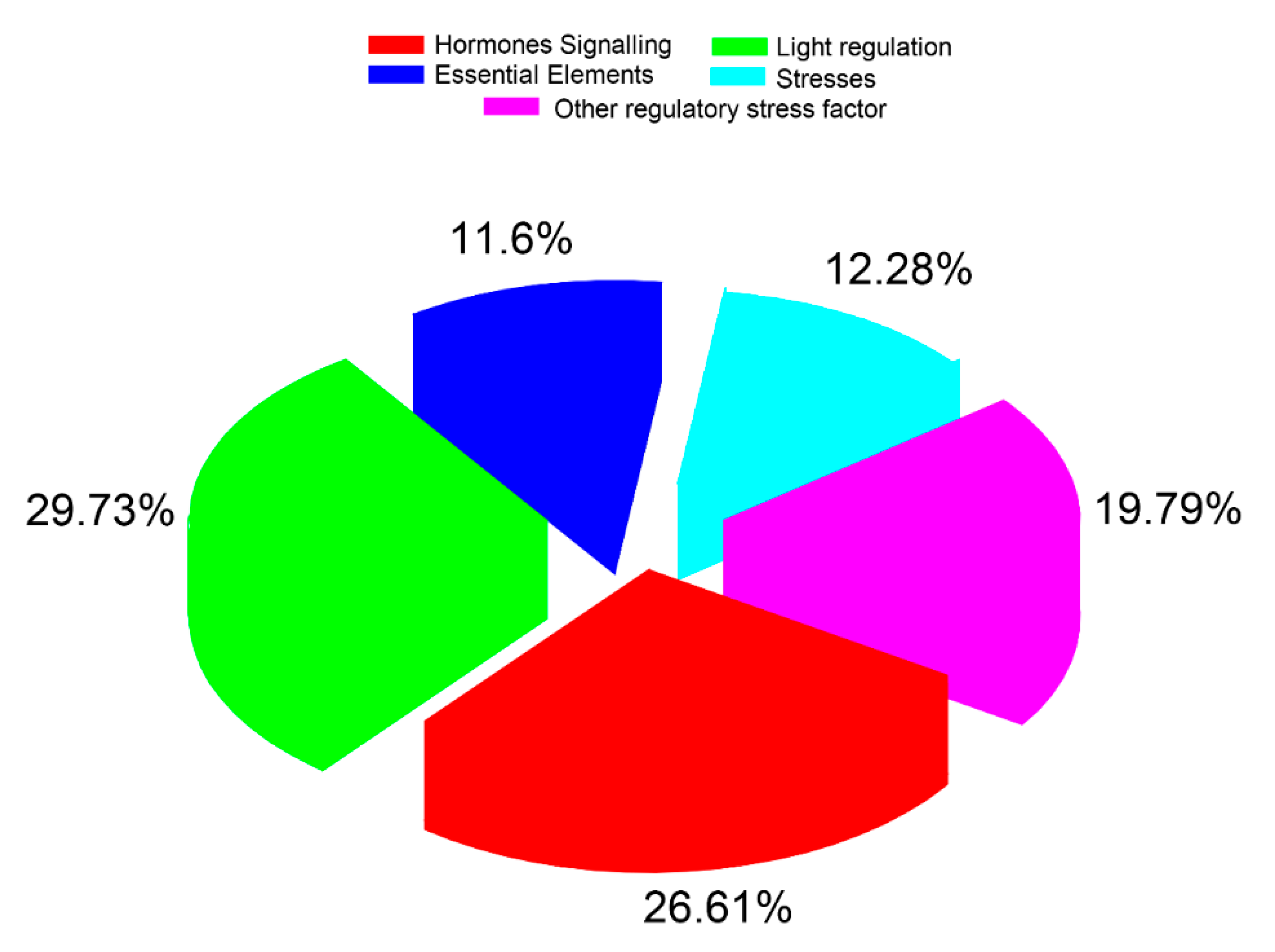
2.4. Gene Ontology (GO) and Promoter Analysis of CsaLecRLK Genes
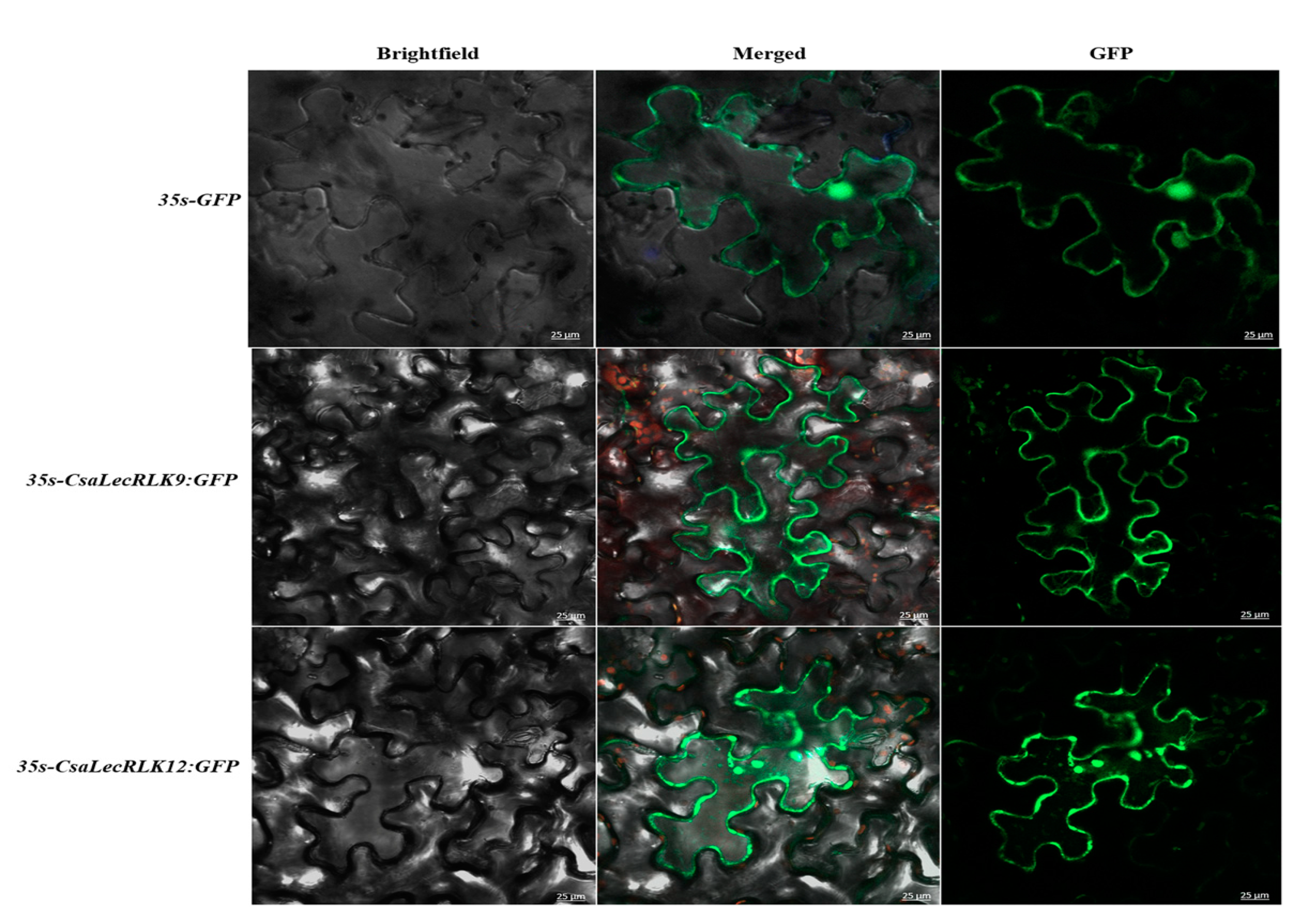
2.5. Subcellular Localization of CsaLecRLKs
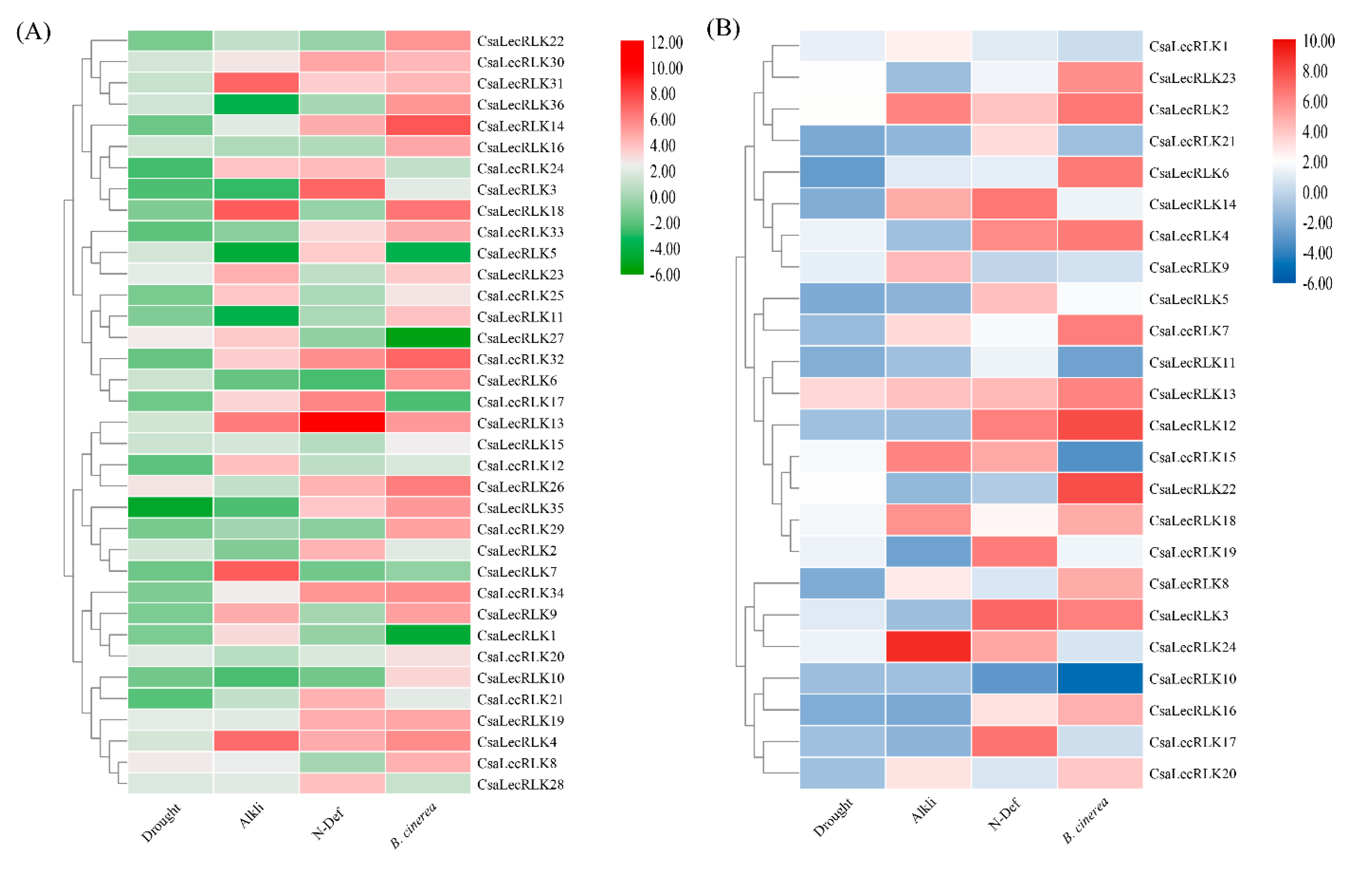
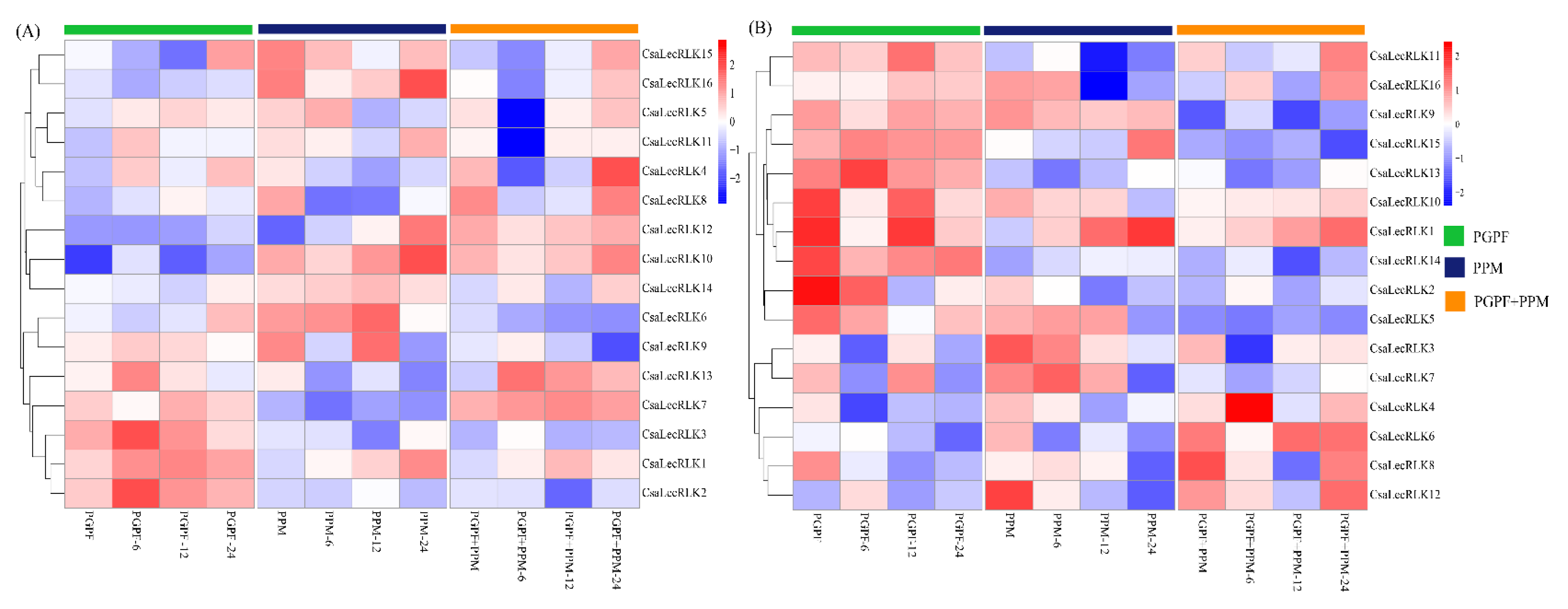
2.6. Expression Pattern and qRT-PCR Validation of CsaLecRLK Genes
3. Discussion
4. Materials and Methods
4.1. Identification and Sequence Retrieval for CsaLecRLK
4.2. Phylogenetic Analysis of CsaLecRLK
4.3. Ka/Ks for Duplicated CsaLecRLK Genes and Their Rate of Divergence
4.4. Conserved Motifs, Exon-Intron Structure Analysis, and Physicochemical Parameters of CsaLecRLK Proteins
4.5. Cis-Elements Predictions of CsaLecRLK
4.6. Chromosomal Location and Syntenic Relationship, and Gene Onotology Enrichment Analysis
4.7. Subcellular Localization of CsaLecRLKs
4.8. Plant Materials and Treatment
4.9. Seed Priming with PGPF-Trichoderma Harzianum Isolate (TriH_JSB27)
4.10. RNA Isolation and Expression Profiling of C. sativus under Biotic Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jogaiah, S.; Praveen, S.; De Britto, S.; Konappa, N.; Udayashankar, A.C. Exogenous priming of chitosan induces upregulation of phytohormones and resistance against cucumber powdery mildew disease is correlated with localized biosynthesis of defense enzymes. Int. J. Biol. Macromol. 2020, 162, 1825–1838. [Google Scholar] [CrossRef]
- Jogaiah, S.; Abdelrahman, M.; Tran, L.-S.P.; Ito, S.-I. Different mechanisms of Trichoderma virens-mediatedresistance in tomato against Fusarium wilt involve the jasmonic and salicylic acid pathways. Mol. Plantpathol. 2018, 19, 870–882. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Xu, N.; Huang, J.; Gao, F.; Zou, H.; Boudsocq, M.; Coaker, G.; Liu, J. A Lectin Receptor-Like Kinase Mediates Pattern-Triggered Salicylic Acid Signaling. Plant Physiol. 2017, 174, 2501–2514. [Google Scholar] [CrossRef] [Green Version]
- Vaid, N.; Macovei, A.; Tuteja, N. Knights in action: Lectin receptor-like kinases in plant development and stress responses. Mol. Plant 2013, 6, 1405–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiu, S.H.; Bleecker, A.B. Receptor-like kinases from Arabidopsis form a monophyletic gene family related to animal receptor kinases. Proc. Natl. Acad. Sci. USA 2001, 98, 10763–10768. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Labbé, J.; Muchero, W.; Yang, X.; Jawdy, S.S.; Kennedy, M.; Johnson, J.; Sreedasyam, A.; Schmutz, J.; Tuskan, G.A.; et al. Genome-wide analysis of lectin receptor-like kinases in Populus. BMC Genom. 2016, 17, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiu, S.H.; Karlowski, W.M.; Pan, R.; Tzeng, Y.H.; Mayer, K.F.; Li, W.H. Comparative analysis of the receptor-like kinase family in Arabidopsis and rice. Plant Cell 2004, 16, 1220–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, T.; Wang, J.; Zhang, B.; Hou, X. Genome-wide analysis of Lectin Receptor-Like Kinases in Tomato (Solanum lycopersicum) and its association with the infection of tomato yellow leaf curl virus. Plant Mol. Biol. Rep. 2018, 36, 429–438. [Google Scholar] [CrossRef]
- Elola, M.T.; Blidner, A.G.; Ferragut, F.; Bracalente, C.; Rabinovich, G.A. Assembly, organization and regulation of cell-surface receptors by lectin–glycan complexes. Biochem. J. 2015, 469, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Jianying, L.; Xiaojing, P.; Tao, L.; Zhijie, N.; Gong, C. The roles of direct recognition by animal Lectins in antiviral immunity and viral pathogenesis. Molecules 2015, 20, 2272–2295. [Google Scholar] [CrossRef] [Green Version]
- Aker, J.; de Vries, S.C. Plasma membrane receptor complexes. Plant Physiol. 2008, 147, 1560–1564. [Google Scholar] [CrossRef] [Green Version]
- Rieu, I.; Laux, T. Signaling pathways maintaining stem cells at the plant shoot apex. Semin. Cell Dev. Biol. 2009, 20, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Jiangqi, W.; Kevin, A.L.; Jason, T.D.; Frans, E.T.; John, W.C. BAK1, an Arabidopsis LRR Receptor-like Protein Kinase, interacts with BRI1 and modulates brassinosteroid signaling. Cell 2002, 110, 213–222. [Google Scholar] [CrossRef]
- Wei, Z.; Wang, J.; Yang, S.; Song, Y. Identification and expression analysis of the LRR-RLK gene family in tomato (Solanum lycopersicum) Heinz 1706. Genome 2015, 58, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Tabata, S.; Hirakawa, H.; Asamizu, E.; Shirasawa, K.; Isobe, S.; Kaneko, T.; Nakamura, Y.; Shibata, D.; Aoki, K. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Nsibo, D.L.; Juhar, H.M.; Govers, F.; Bouwmeester, K. Ectopic expression of Arabidopsis L-type lectin receptor kinase genes LecRK-I.9 and LecRK-IX.1 in Nicotiana benthamiana confers Phytophthora resistance. Plant Cell. Rep. 2016, 35, 845–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labbe, J.; Muchero, W.; Czarnecki, O.; Wang, J.; Wang, X.; Bryan, A.C.; Zheng, K.; Yang, Y.; Xie, M.; Zhang, J.; et al. Mediation of plant-mycorrhizal interaction by a lectin receptor-like kinase. Nat. Plants 2019, 5, 676–680. [Google Scholar] [CrossRef]
- Lv, D.; Wang, G.; Xiong, L.-R.; Sun, J.-X.; Chen, Y.; Guo, C.-L.; Yu, Y.; He, H.-L.; Cai, R.; Pan, J.-S. Genome-Wide Identification and Characterization of Lectin Receptor-Like Kinase Gene Family in Cucumber and Expression Profiling Analysis under Different Treatments. Genes 2020, 11, 1032. [Google Scholar] [CrossRef]
- Liu, P.L.; Yuan, H.; Shi, P.H.; Meng, Y.; Xie, J.B.; Lulu, X. Duplication and diversification of lectin receptor-like kinases (LecRLK) genes in soybean. Sci. Rep. 2018, 8, 5861. [Google Scholar] [CrossRef] [Green Version]
- Haider, M.S.; Kurjogi, M.; Khalil, U.R.; Fiaz, M.; Tariq, P.; Jiu, S.; Jia, H.; Fang, J. Grapevine immune signaling network in response to drought stress as revealed by transcriptomic analysis. Plant Physiol. Biochem. 2017, 121, 187–195. [Google Scholar] [CrossRef]
- Shumayla, S.; Sharma, S.; Pandey, A.K.; Singh, K.; Upadhyay, S.K. Molecular characterization and global expression analysis of Lectin Receptor Kinases in Bread Wheat (Triticum aestivum). PLoS ONE 2016, 11, e0153925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiu, S.-H.; Bleecker, A.B. Expansion of the Receptor-like Kinase/Pelle Gene Family and Receptor-like proteins in Arabidopsis. Plant Physiol. 2003, 132, 530–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, H.; Heibl, C.; Renner, S.S. Gourds afloat: A dated phylogeny reveals an Asian origin of the gourd family (Cucurbitaceae) and numerous oversea dispersal events. Proc. R. Soc. B 2009, 276, 843–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.; Liu, Y.W.; Zhou, J.M.; Zhao, S.P.; Zhang, X.H.; Min, D.H. Genome-wide analysis of the lectin receptor-like kinase family in foxtail millet (Setaria italica L.). Plant Cell Tissue Organ Cult. (PCTOC) 2016, 127, 335–346. [Google Scholar] [CrossRef]
- Khan, N.; Han, K.; Hu, C.M.; Naseri, E.; Haider, M.S.; Ayaz, A.; Khan, W.A.; Wang, J.; Hou, X. Genome-wide identification, evolution, and transcriptional profiling of PP2C Gene family in Brassica rapa. BioMed Res. Int. 2019, 2019, 2965035. [Google Scholar] [CrossRef] [Green Version]
- Koschmann, J.; Machens, F.; Becker, M.; Niemeyer, J.; Schulze, J.; Bülow, L.; Stahl, D.J.; Hehl, R. Integration of bioinformatics and synthetic promoters leads to the discovery of novel elicitor-responsive cis-regulatory sequences in Arabidopsis. Plant Physiol. 2012, 160, 178–191. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.G.; Ellis, C.; Devoto, A. The Jasmonate Signal Pathway. Plant Cell 2002, 14, S153–S164. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wang, F.; Yu, G.; Zhang, X.; Jia, C.; Qin, J.; Pan, H. Functional Analysis of the Maize C-Repeat/DRE Motif-Binding Transcription Factor CBF3 Promoter in Response to Abiotic Stress. Int. J. Mol. Sci. 2015, 16, 12131–12146. [Google Scholar] [CrossRef] [Green Version]
- Yura, T.; Nakahigashi, K. Regulation of the Heat-Shock Response. Curr. Opin. Microbiol. 1998, 117, 1135–1141. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Organization of cis-acting regulatory elements in osmotic- and cold-stress-responsive promoters. Trends Plant Sci. 2005, 10, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Haider, M.S.; Khan, N.; Tariq, P.; Liu, Z.; Maazullah, N.; Jogaiah, S.; Mushtaq, N.; Jiu, S.; Fang, J. Genome-wide identification, evolution, and molecular characterization of the PP2C gene family in woodland strawberry. Gene 2019, 702, 27–35. [Google Scholar] [CrossRef]
- Sun, X.L.; Yu, Q.Y.; Tang, L.L.; Ji, W.; Bai, X.; Cai, H.; Liu, X.F.; Ding, X.D.; Zhu, Y.M. GsSRK, a G-type lectin S-receptor-like serine/threonineprotein kinase, is a positive regulator of plant tolerance to salt stress. J. Plant Physiol. 2013, 170, 505–515. [Google Scholar] [CrossRef]
- Chen, T.; Lv, Y.; Zhao, T.; Li, N.; Yang, Y.; Yu, W.; He, X.; Liu, T.; Zhang, B. Comparative transcriptome profiling of a resistant vs. susceptible tomato (Solanum lycopersicum) cultivar in response to infection by tomato yellow leaf curl virus. PLoS ONE 2013, 8, e80816. [Google Scholar] [CrossRef]
- Ma, N.; Liu, C.; Li, H.; Wang, J.; Zhang, B.; Lin, J.; Chang, Y. Genome-wide identification of Lectin Receptor Kinases in pear: Functional characterization of the L-Type LecRLK Gene PbLRK138. Gene 2018, 66111–66121. [Google Scholar] [CrossRef] [PubMed]
- Sasabe, M.; Naito, K.; Suenaga, H.; Ikeda, T.; Toyoda, K.; Inagaki, Y.; Shiraishi, T.; Ichinose, Y. Elicitin-responsive lectin-like receptor kinase genes in BY-2 cells. DNA Seq. 2007, 18152–18159. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Bouwmeester, K. L-type lectin receptor kinases: New forces in plant immunity. PLoS Pathog. 2017, 13, e1006433. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.M.; Dubery, I.A.; van Heerden, H. Identification and molecular characterisation of a Lectin Receptor-like Kinase (GhLecRK-2) from cotton. Plant Mol. Biol. Rep. 2013, 319–320. [Google Scholar] [CrossRef]
- Wang, Z.; Cheng, J.; Fan, A.; Zhao, J.; Yu, Z.; Li, Y.; Zhang, H.; Xiao, J.; Muhammad, F.; Wang, H.; et al. LecRK-V, an L-type lectin receptor kinase in haynaldia villosa, plays positive role in resistance to wheat powdery mildew. Plant Biotechnol. J. 2018, 16, 50–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.B.; Sun, Y.D.; Liu, H.; Wang, Y.Y.; Jia, Y.L.; Xu, M.H. Natural variation of rice blast resistance gene Pi-d2. Genet. Mol. Res. 2015, 14, 1235–1249. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Doerks, T.; Bork, P. SMART 7: Recent updates to the protein domain annotation resource. Nucleic Acids Res. 2012, 40, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, M.A.; Haubold, B.; Mitchell-Olds, T. Comparative evolutionary analysis of chalcone synthase and alcohol dehydrogenase loci in Arabidopsis, Arabis, and Related Genera (Brassicaceae). Mol. Biol. Evol. 2000, 17, 1483–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, 202–208. [Google Scholar] [CrossRef]
- Hu, B.; Jinpu, J.; Anyuan, G.; He, Z.; Jingchu, L.; Ge, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Khan, N.; Fatima, F.; Haider, M.S.; Shazadee, H.; Liu, Z.; Zheng, T.; Fang, J. Genome-wide identification and expression profiling of the polygalacturonase (PG) and pectin methylesterase (PME) genes in Grapevine (Vitis vinifera L.). Int. J. Mol. Sci. 2019, 20, 3180. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Xia, R.; Chen, H.; He, Y. Tbtools, a toolkit for biologists integrating various hts-data handling tools with a user-friendly interface. bioRxiv 2018. [Google Scholar] [CrossRef]
- Jogaiah, S.; Mostafa, A.; Tran, L.S.P.; Shin-ichi, I. Characterization of rhizosphere fungi that mediate resistance in tomato against bacterial wilt disease. J. Exp. Bot. 2013, 64, 3829–3842. [Google Scholar] [CrossRef] [Green Version]
- Haider, M.S.; Jogaiah, S.; Tariq, P.; Zhao, Y.; Nadeem, K.; Fang, J. Physiological and transcriptional variations inducing complex adaptive mechanisms in grapevine by salt stress. Environ. Exp. Bot. 2019, 162, 455–467. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene 1 | Gene 2 | Ks | Ka | Ka/Ks | Divergence Time | Selection Pressure | Gene Duplications |
|---|---|---|---|---|---|---|---|
| Between L-type genes | |||||||
| CsaLecRLK1 | CsaLecRLK5 | 1.19 | 0.40 | 0.33 | 39.67 | Purifying Selection | WGD or Segmental |
| CsaLecRLK14 | CsaLecRLK16 | 0.91 | 0.53 | 0.58 | 30.33 | Purifying Selection | WGD or Segmental |
| CsaLecRLK2 | CsaLecRLK4 | 0.51 | 0.55 | 1.09 | 17.00 | Positive Selection | Tandem |
| CsaLecRLK6 | CsaLecRLK7 | 0.34 | 0.38 | 1.10 | 11.33 | Positive Selection | Tandem |
| CsaLecRLK18 | CsaLecRLK19 | 0.18 | 0.28 | 1.55 | 6.00 | Positive Selection | Tandem |
| CsaLecRLK22 | CsaLecRLK23 | 0.52 | 0.14 | 0.26 | 17.33 | Purifying Selection | Tandem |
| CsaLecRLK8 | CsaLecRLK9 | 0.79 | 0.38 | 0.48 | 26.33 | Purifying Selection | Dispersed |
| CsaLecRLK10 | CsaLecRLK13 | 0.84 | 0.55 | 0.65 | 28.00 | Purifying Selection | Dispersed |
| CsaLecRLK20 | CsaLecRLK21 | 1.73 | 0.30 | 0.17 | 57.67 | Purifying Selection | Dispersed |
| CsaLecRLK3 | CsaLecRLK11 | 0.89 | 0.59 | 0.67 | 29.67 | Purifying Selection | Proximal |
| CsaLecRLK12 | CsaLecRLK17 | 0.85 | 0.61 | 0.72 | 28.33 | Purifying Selection | Proximal |
| Between G-type genes | |||||||
| CsaLecRLK2 | CsaLecRLK3 | 0.22 | 0.41 | 1.85 | 7.33 | Positive Selection | WGD or Segmental |
| CsaLecRLK4 | CsaLecRLK10 | 0.77 | 0.65 | 0.84 | 25.67 | Purifying Selection | WGD or Segmental |
| CsaLecRLK12 | CsaLecRLK15 | 0.97 | 0.45 | 0.46 | 32.33 | Purifying Selection | WGD or Segmental |
| CsaLecRLK17 | CsaLecRLK18 | 1.36 | 0.43 | 0.31 | 45.33 | Purifying Selection | WGD or Segmental |
| CsaLecRLK19 | CsaLecRLK20 | 0.98 | 0.16 | 0.16 | 32.67 | Purifying Selection | WGD or Segmental |
| CsaLecRLK22 | CsaLecRLK25 | 0.65 | 0.49 | 0.76 | 21.67 | Purifying Selection | WGD or Segmental |
| CsaLecRLK27 | CsaLecRLK33 | 0.81 | 0.49 | 0.60 | 27.00 | Purifying Selection | WGD or Segmental |
| CsaLecRLK35 | CsaLecRLK36 | 1.03 | 0.47 | 0.46 | 34.33 | Purifying Selection | WGD or Segmental |
| CsaLecRLK8 | CsaLecRLK11 | 0.81 | 0.58 | 0.72 | 27.00 | Purifying Selection | Tandem |
| CsaLecRLK13 | CsaLecRLK16 | 0.67 | 0.53 | 0.79 | 22.33 | Purifying Selection | Tandem |
| CsaLecRLK21 | CsaLecRLK23 | 1.07 | 0.48 | 0.44 | 35.67 | Purifying Selection | Tandem |
| CsaLecRLK24 | CsaLecRLK26 | 0.41 | 0.38 | 0.93 | 13.67 | Purifying Selection | Tandem |
| CsaLecRLK28 | CsaLecRLK29 | 0.53 | 0.60 | 1.12 | 17.67 | Positive Selection | Tandem |
| CsaLecRLK30 | CsaLecRLK31 | 0.48 | 0.14 | 0.29 | 16.00 | Purifying Selection | Tandem |
| CsaLecRLK6 | CsaLecRLK7 | 0.96 | 0.57 | 0.59 | 32.00 | Purifying Selection | Dispersed |
| CsaLecRLK5 | CsaLecRLK14 | 0.54 | 0.61 | 1.13 | 18.00 | Positive Selection | Proximal |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haider, M.S.; De Britto, S.; Nagaraj, G.; Gurulingaiah, B.; Shekhar, R.; Ito, S.-i.; Jogaiah, S. Genome-Wide Identification, Diversification, and Expression Analysis of Lectin Receptor-Like Kinase (LecRLK) Gene Family in Cucumber under Biotic Stress. Int. J. Mol. Sci. 2021, 22, 6585. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126585
Haider MS, De Britto S, Nagaraj G, Gurulingaiah B, Shekhar R, Ito S-i, Jogaiah S. Genome-Wide Identification, Diversification, and Expression Analysis of Lectin Receptor-Like Kinase (LecRLK) Gene Family in Cucumber under Biotic Stress. International Journal of Molecular Sciences. 2021; 22(12):6585. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126585
Chicago/Turabian StyleHaider, Muhammad Salman, Savitha De Britto, Geetha Nagaraj, Bhavya Gurulingaiah, Ravikant Shekhar, Shin-ichi Ito, and Sudisha Jogaiah. 2021. "Genome-Wide Identification, Diversification, and Expression Analysis of Lectin Receptor-Like Kinase (LecRLK) Gene Family in Cucumber under Biotic Stress" International Journal of Molecular Sciences 22, no. 12: 6585. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126585