
Molecular and Electrophysiological Role of Diabetes-Associated Circulating Inflammatory Factors in Cardiac Arrhythmia Remodeling in a Metabolic-Induced Model of Type 2 Diabetic Rat

 , ,
, ,
Abstract
:1. Introduction
2. Results
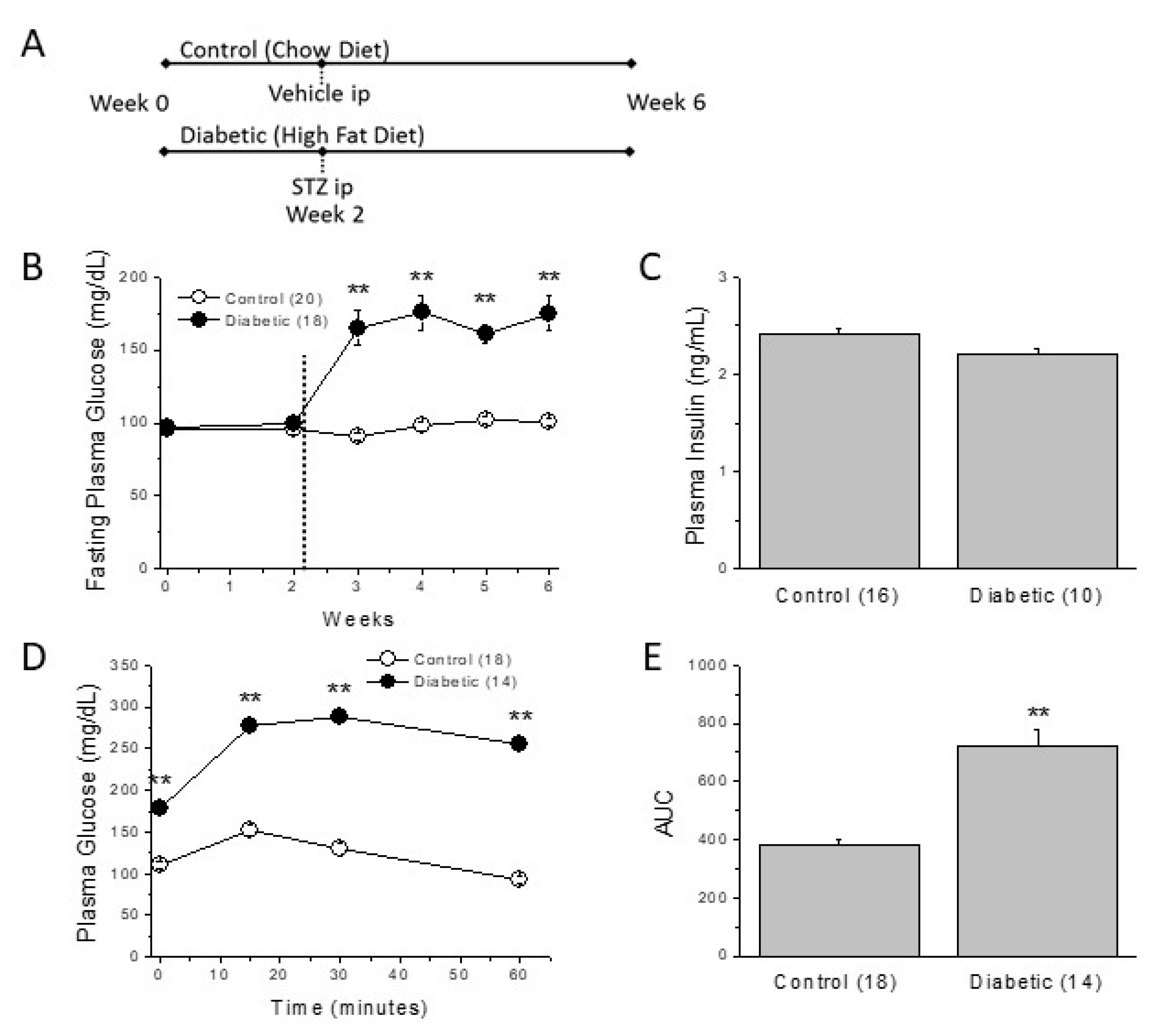
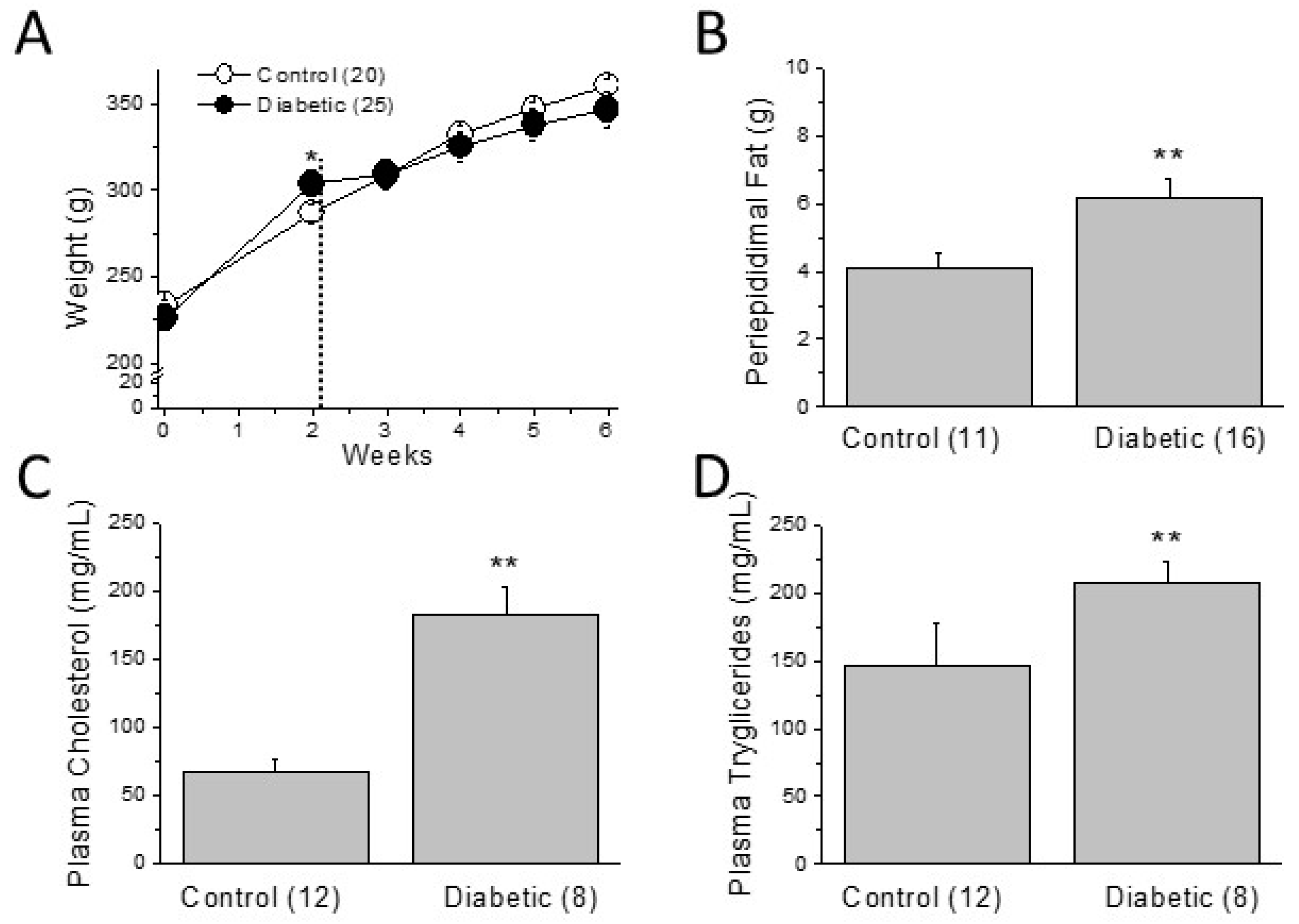
2.1. Metabolic Characterization of the Type 2 Diabetic Model
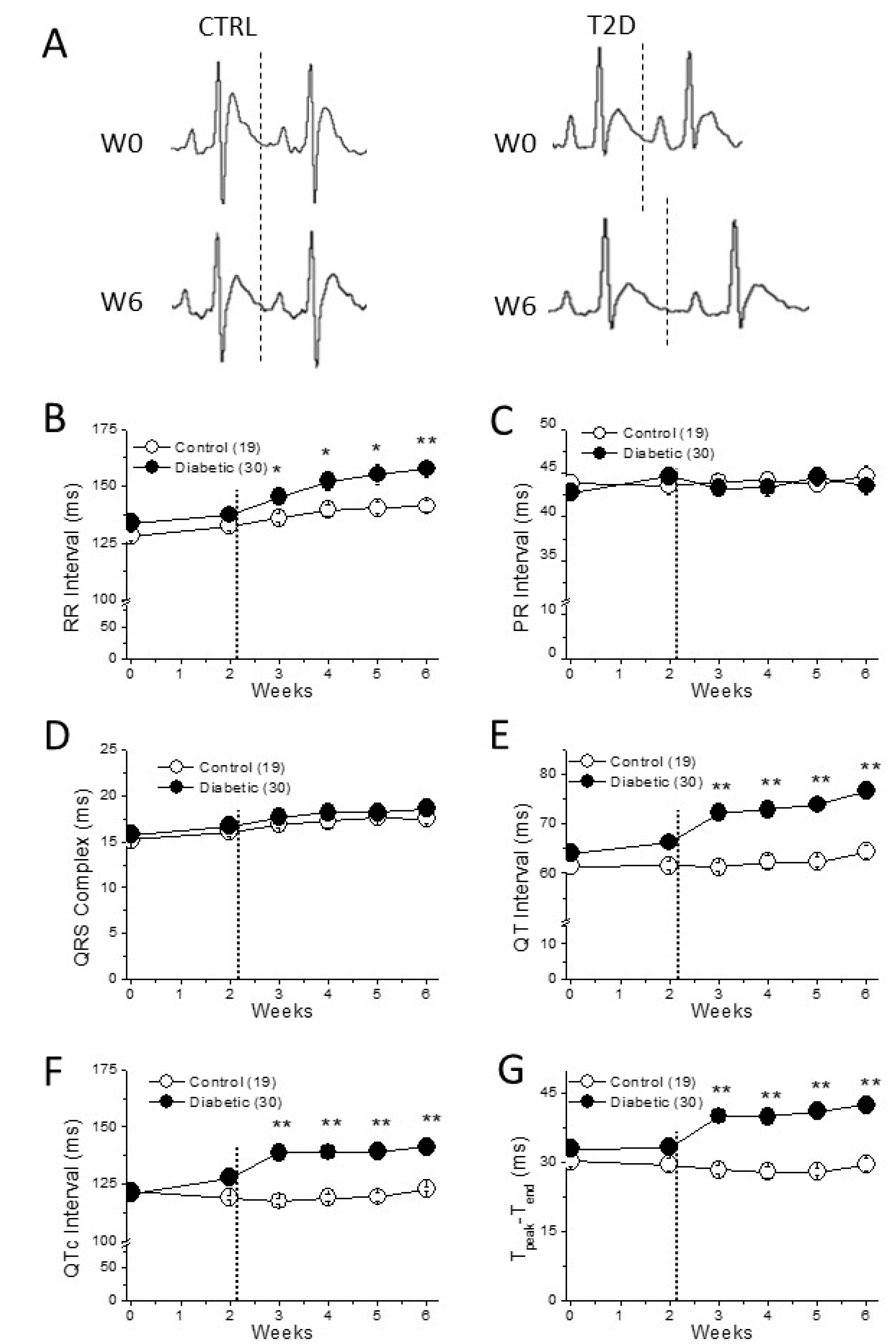
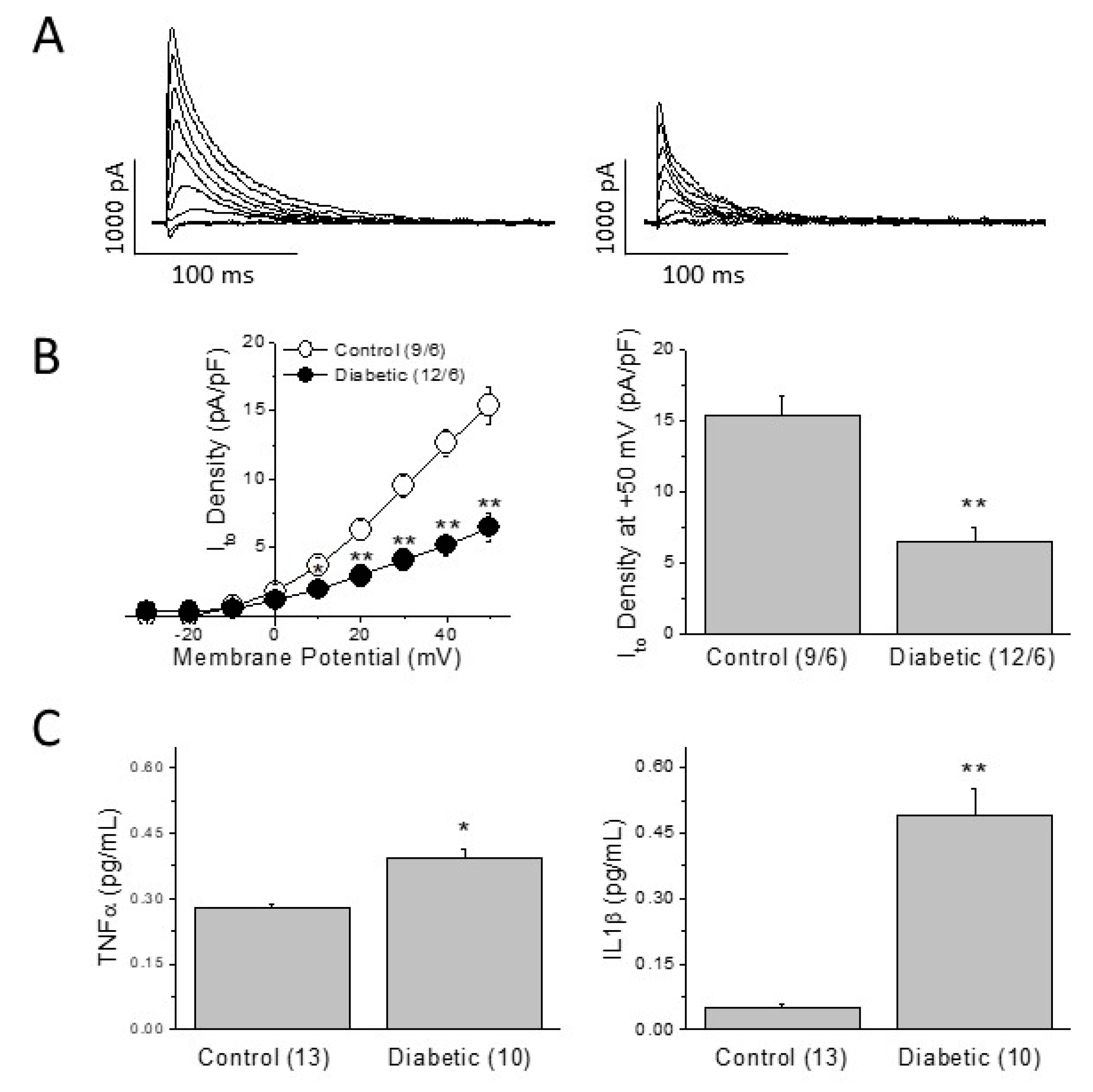
2.2. Cardiac Electrical Remodeling in the Type 2 Diabetic Model
2.3. Mechanisms of the Diabetes-Induced Electrical Remodeling
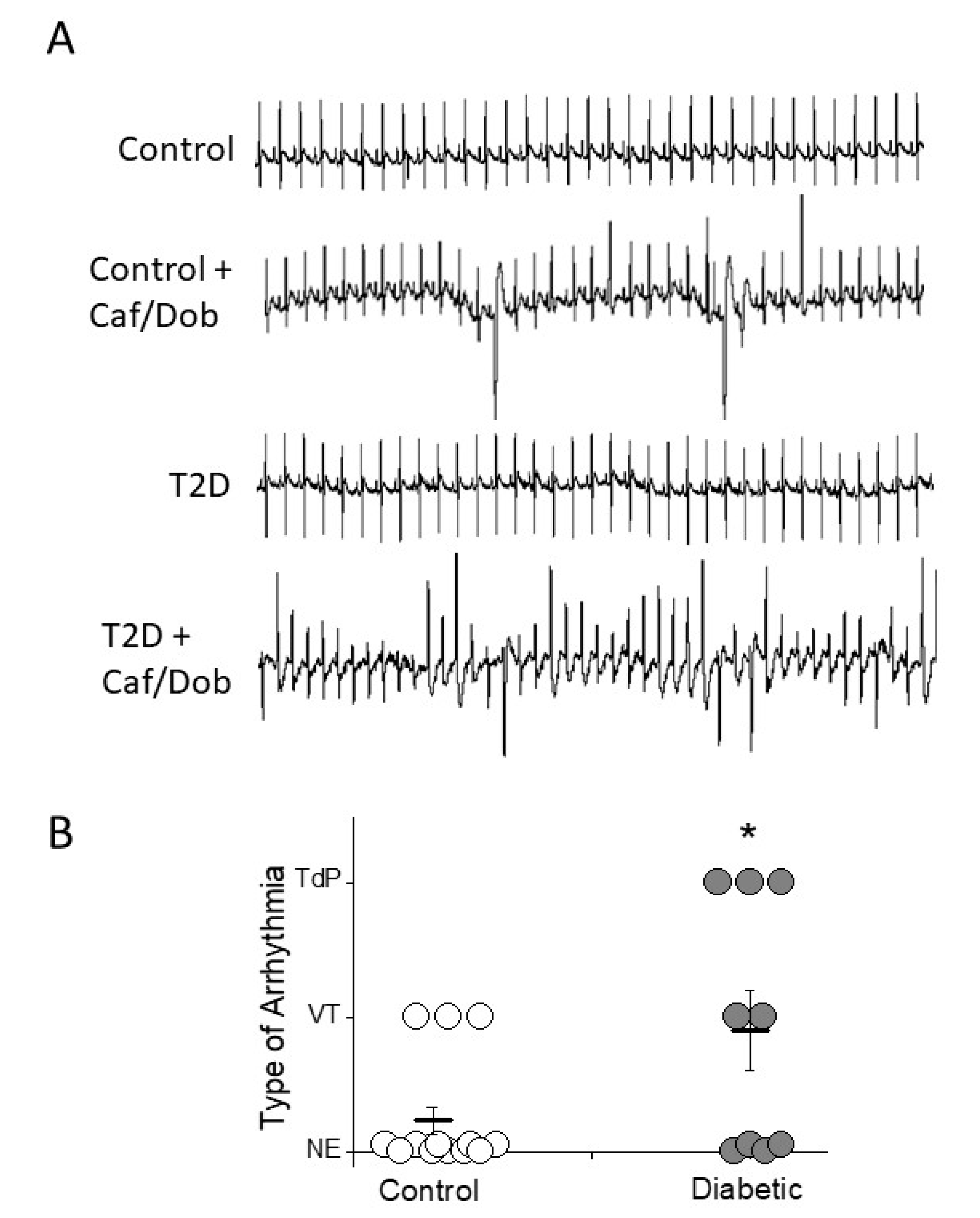
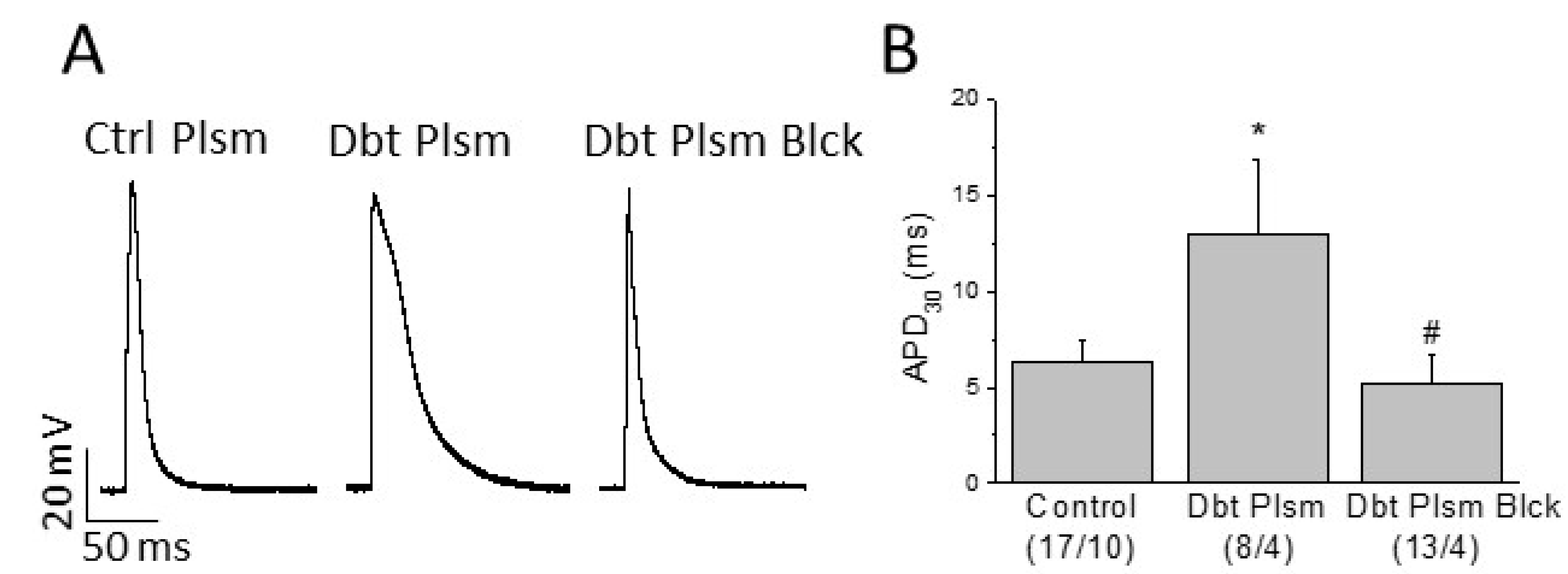
2.4. Role of Circulating Factors on Cardiac Electrical Remodeling
3. Discussion
3.1. Limitations
3.2. Summary
4. Materials and Methods
4.1. Induction of Diabetes and In Vivo Treatments
4.2. Blood Measurements
4.3. Combined Intraperitoneal Insulin and Glucose Tolerance Test, IPIGTT
4.4. In Vivo ECG Recordings
4.5. Cardiomyocyte Isolation
4.6. Patch-Clamp Experiments
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- El-Atat, F.A.; McFarlane, S.I.; Sowers, J.R.; Bigger, J.T. Sudden cardiac death in patients with diabetes. Curr. Diabetes Rep. 2004, 4, 187–193. [Google Scholar] [CrossRef]
- Tse, G.; Lai, E.T.H.; Tse, V.; Yeo, J.M. Molecular and electrophysiological mechanisms underlying cardiac arrhythmogenesis in diabetes mellitus. J. Diabetes Res. 2016, 2016, 2848759. [Google Scholar] [CrossRef] [Green Version]
- Sanguinetti, M.C.; Tristani-Firouzi, M. hERG Potassium Channels and Cardiac Arrhythmia. Nature 2006, 440, 463–469. [Google Scholar] [CrossRef]
- Kahn, J.K.; Sisson, J.C.; Vinik, A.I. QT interval prolongation and sudden cardiac death in diabetic autonomic neuropathy. J. Clin. Endocrinol. Metab. 1987, 64, 751–754. [Google Scholar] [CrossRef]
- Bellavere, F.; Ferri, M.; Guarini, L.; Bax, G.; Piccoli, A.; Cardone, C.; Fedele, D. Prolonged QT period in diabetic autonomic neuropathy—a possible role in sudden cardiac death. Br. Heart J. 1988, 59, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Naas, A.; Davidson, N.C.; Thompson, C.; Cummings, F.; Ogston, S.; Jung, R.T.; Newton, W.; Struthers, A.D. QT and QTc dispersion are accurate predictors of cardiac death in newly diagnosed non-insulin dependent diabetes: Cohort study. Br. Med. J. 1998, 316, 745–746. [Google Scholar] [CrossRef] [Green Version]
- Lengyel, C.; Virag, L.; Biro, T.; Jost, N.; Magyar, J.; Biliczki, P.; Kocsis, E.; Skoumal, P.; Nánási, P.P.; Tóth, M.; et al. Diabetes mellitus attenuates the repolarization reserve in mammalian heart. Cardiovasc. Res. 2007, 73, 512–520. [Google Scholar] [CrossRef] [Green Version]
- Torres-Jacome, J.; Gallego, M.; Rodriguez-Robledo, J.M.; Sanchez-Chapula, J.A.; Casis, O. Improvement of the metabolic status recovers cardiac potassium channel synthesis in experimental diabetes. Acta Physiol. 2013, 207, 447–459. [Google Scholar] [CrossRef]
- Niwa, N.; Nerbonne, J.M. Molecular determinants of cardiac transient outward potassium current (Ito) expression and regulation. J. Mol. Cell. Cardiol. 2010, 48, 12–25. [Google Scholar] [CrossRef] [Green Version]
- Dutka, M.; Bobiński, R.; Ulman-Włodarz, I.; Hajduga, M.; Bujok, J.; Pająk, C.; Ćwiertnia, M. Various aspects of inflammation in heart failure. Heart Fail. Rev. 2020, 25, 537–548. [Google Scholar] [CrossRef] [Green Version]
- Van Linthout, S.; Tschöpe, C. Inflammation—Cause or Consequence of Heart Failure or Both? Curr. Heart Fail. Rep. 2017, 14, 251–265. [Google Scholar] [CrossRef] [Green Version]
- Monnerat-Cahli, G.; Alarcon, M.L.; Vasconcellos, L.R.; Hochman-Mendez, C.; Brasil, G.; Bassani, R.A.; Casis, O.; Malan, D.; Travassos, L.H.; Sepúlveda, M.; et al. Macrophage-dependent IL-1 beta production induces cardiac arrhythmias in diabetic mice. Nat. Commun. 2016, 7, 13344. [Google Scholar] [CrossRef] [Green Version]
- Monnerat-Cahli, G.; Alonso, H.; Gallego, M.; Alarcón, M.L.; Bassani, R.A.; Casis, O.; Medei, E. Toll-like receptor 4 activation promotes cardiac arrhythmias by decreasing the transient outward potassium current (Ito) through an IRF3-dependent and MyD88-independent pathway. J. Mol. Cell. Cardiol. 2014, 76, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Velasco, M.; Ruiz-Hurtado, G.; Hurtado, O.; Moro, M.A.; Delgado, C. TNF-alpha downregulates transient outward potassium current in rat ventricular myocytes through iNOS overexpression and oxidant species generation. Am. J. Physiol.-Heart Circ. Physiol. 2007, 293, H238–H245. [Google Scholar] [CrossRef] [Green Version]
- Devaraj, S.; Dasu, M.R.; Rockwood, J.; Winter, W.; Griffen, S.C.; Jialal, I. Increased toll-like receptor (TLR) 2 and TLR4 expression in monocytes from patients with type 1 diabetes: Further evidence of a pro-inflammatory state. J. Clin. Endocrinol. Metab. 2008, 93, 578–583. [Google Scholar] [CrossRef] [Green Version]
- Dasu, M.R.; Devaraj, S.; Park, S.; Jialal, I. Increased toll-like receptor (TLR) activation and TLR ligands in recently diagnosed type 2 diabetic subjects. Diabetes Care 2010, 33, 861–868. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.M.; Kim, J.J.; Kim, H.J.; Shong, M.; Ku, B.J.; Jo, E.K. Upregulated NLRP3 inflammasome activation in patients with type 2 diabetes. Diabetes 2013, 62, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Magyar, J.; Rusznak, Z.; Szentesi, P.; Szucs, G.; Kovacs, L. Action-potentials and potassium currents in rat ventricular muscle during experimental diabetes. J. Mol. Cell. Cardiol. 1992, 24, 841–853. [Google Scholar] [CrossRef]
- Casis, O.; Gallego, M.; Iriarte, M.; Sanchez-Chapula, J.A. Effects of diabetic cardiomyopathy on regional electrophysiologic characteristics of rat ventricle. Diabetologia 2000, 43, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Lengyel, C.; Virag, L.; Kovacs, P.P.; Kristof, A.; Pacher, P.; Kocsis, E.; Koltay, Z.M.; Nánási, P.P.; Tóth, M.; Kecskeméti, V.; et al. Role of slow delayed rectifier K+-current in QT prolongation in the alloxan-induced diabetic rabbit heart. Acta Physiol. 2008, 192, 359–368. [Google Scholar] [CrossRef]
- Reed, M.J.; Meszaros, K.; Entes, L.J.; Claypool, M.D.; Pinkett, J.G.; Gadbois, T.M.; Reaven, G.M. New Rat Model of Type 2 Diabetes: The Fat-Fed, Streptozotocin-Treated Rat. Metabolism 2000, 49, 1390–1394. [Google Scholar] [CrossRef]
- Ti, Y.; Xie, G.; Wang, Z.H.; Bi, X.L.; Ding, W.Y.; Wang, J.; Jiang, G.J.; Bu, P.L.; Zhang, Y.; Zhong, M.; et al. TRB3 Gene Silencing Alleviates Diabetic Cardiomyopathy in a Type 2 Diabetic Rat Model. Diabetes 2011, 60, 2963–2974. [Google Scholar] [CrossRef] [Green Version]
- Ionut, V.; Liu, H.; Mooradian, V.; Castro, A.V.B.; Kabir, M.; Stefanovski, D.; Zheng, D.; Kirkman, E.L.; Bergman, R.N. Novel canine models of obese prediabetes and mild type 2 diabetes. Am. J. Physiol.-Endocrinol. Metab. 2010, 298, E38–E48. [Google Scholar] [CrossRef]
- Podell, B.K.; Ackart, D.F.; Richardson, M.A.; DiLisio, J.E.; Pulford, B.; Basaraba, R.J. A model of type 2 diabetes in the guinea pig using sequential diet-induced glucose intolerance and streptozotocin treatment. Dis. Mod. Mech. 2017, 10, 151–162. [Google Scholar] [CrossRef] [Green Version]
- Rout, D.; Chandra, D.U.; Kanhar, S.; Swain, S.K.; Sahoo, A.K. The modulatory role of prime identified compounds in the bioactive fraction of Homalium zeylanicum in high-fat diet fed-streptozotocin-induced type 2 diabetic rats. J. Ethnopharmacol. 2020, 260, 113099. [Google Scholar] [CrossRef] [PubMed]
- Shawky, L.M.; Morsi, A.A.; El Bana, E.; Hanafy, S.M. The Biological Impacts of Sitagliptin on the Pancreas of a Rat Model of Type 2 Diabetes Mellitus: Drug Interactions with Metformin. Biology 2019, 9, 6. [Google Scholar] [CrossRef] [Green Version]
- Navabi, R.; Negahdari, B.; Hajizadeh-Saffar, E.; Hajinasrollah, M.; Jenab, Y.; Rabbani, S.; Pakzad, M.; Hassani, S.N.; Hezavehei, M.; Jafari-Atrabi, M.; et al. Combined therapy of mesenchymal stem cells with a GLP-1 receptor agonist, liraglutide, on an inflammatory-mediated diabetic non-human primate model. Life Sci. 2021, 276, 119374. [Google Scholar] [CrossRef] [PubMed]
- Jialal, I.; Singh, G. Management of diabetic dyslipidemia: An update. World J. Diabetes 2019, 10, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Pfister, R.; Cairns, R.; Erdmann, E.; Schneider, C.A.; PROactive investigators. Prognostic impact of electrocardiographic signs in patients with Type 2 diabetes and cardiovascular disease: Results from the PROactive study. Diabetes Med. 2011, 28, 1206–1212. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.X.; Antzelevitch, C. Cellular basis for the normal T wave and the electrocardiographic manifestations of the long QT syndrome. Circulation 1998, 98, 1928–1936. [Google Scholar] [CrossRef]
- Tse, G.; Gong, M.; Wong, W.T.; Georgopoulos, S.; Letsas, K.P.; Vassiliou, V.S.; Chan, Y.S.; Yan, B.P.; Wong, S.H.; Wu, W.K.K.; et al. The Tpeak-Tend interval as an electrocardiographic risk marker of arrhythmic and mortality outcomes: A systematic review and meta-analysis. Heart Rhythm 2017, 14, 1131–1137. [Google Scholar] [CrossRef] [Green Version]
- Bloom, L.J.; Groeneveld, S.A.; Wulterkens, B.M.; van Rees, B.; Nguyen, U.C.; Roudijk, R.W.; Cluitmans, M.; Volders, P.G.A.; Hassink, R.J. Novel use of repolarization parameters in electrocardiographic imaging to uncover arrhythmogenic substrate. J. Electrocardiol. 2020, 59, 116–121. [Google Scholar] [CrossRef]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic Adipose Tissue Inflammation Linking Obesity to Insulin Resistance and Type 2 Diabetes. Front. Physiol. 2020, 10, 1607. [Google Scholar] [CrossRef]
- Maida, C.D.; Vasto, S.; Di Raimondo, D.; Casuccio, A.; Vassallo, V.; Daidone, M.; Del Cuore, A.; Pacinella, G.; Cirrincione, A.; Simonetta, I.; et al. Inflammatory activation and endothelial dysfunction markers in patients with permanent atrial fibrillation: A cross-sectional study. Aging 2020, 12, 8423–8433. [Google Scholar] [CrossRef]
- Bello, F.; Marchi, A.; Prisco, D.; Olivotto, I.; Emmi, G. Antiarrhythmic efficacy of anakinra in a young patient with autoimmune lymphocytic myocarditis. Rheumatology 2020, 59, keaa207. [Google Scholar] [CrossRef]
- Cohen, C.D.; De Blasio, M.J.; Lee, M.K.S.; Farrugia, G.E.; Prakoso, D.; Krstevski, C.; Deo, M.; Donner, D.G.; Kiriazis, H.; Flynn, M.C.; et al. Diastolic dysfunction in a pre-clinical model of diabetes is associated with changes in the cardiac non-myocyte cellular composition. Cardiovasc. Diabetol. 2021, 20, 116. [Google Scholar] [CrossRef]
- White, P.C.; Adhikari, S.; Grishman, E.K.; Sumpter, K.M. A phase I study of anti-inflammatory therapy with rilonacept in adolescents and adults with type 1 diabetes mellitus. Pediatr. Diabetes 2018, 19, 788–793. [Google Scholar] [CrossRef] [PubMed]
- Moran, A.; Bundy, B.; Becker, D.J.; DiMeglio, L.A.; Gitelman, S.E.; Goland, R.; Greenbaum, C.J.; Herold, K.C.; Marks, J.B.; Raskin, P.; et al. Interleukin-1 antagonism in type 1 diabetes of recent onset: Two multicentre, randomised, double-blind, placebo-controlled trials. Lancet 2013, 381, 1905–1915. [Google Scholar] [CrossRef] [Green Version]
- Chiamvimonvat, N.; Chen-Izu, Y.; Clancy, C.E.; Deschenes, I.; Dobrev, D.; Heijman, J.; Izu, L.; Qu, Z.; Ripplinger, C.M.; Vandenberg, J.I.; et al. Potassium currents in the heart: Functional roles in repolarization, arrhythmia and therapeutics. J. Physiol. 2017, 595, 2229–2252. [Google Scholar] [CrossRef] [Green Version]
- Tande, P.M.; Bjornstad, H.; Yant, G.; Refsum, H. Rate-dependent class III antiarrhythmic action, negative chronotropy, and positive inotropy of a novel IK blocking drug UK-68-798: Potent in guinea-pig but no effect in rat myocardium. J. Cardiovasc. Pharmacol. 1990, 16, 401–410. [Google Scholar] [CrossRef]
- Randeria, S.N.; Thomson, G.J.A.; Nell, T.A.; Roberts, T.; Pretorius, E. Inflammatory cytokines in type 2 diabetes mellitus as facilitators of hypercoagulation and abnormal clot formation. Cardiovasc. Diabetol. 2019, 18, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, J.R.; Pereira, L.; Wang, L.; Han, G.; Ferguson, A.; Dao, K.; Copeland, R.J.; Despa, F.; Hart, G.W.; Ripplinger, C.M.; et al. Diabetic hyperglycaemia activates CaMKII and arrhythmias by O-linked glycosylation. Nature 2013, 502, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Curtis, M.J.; Hancox, J.C.; Farkas, A.; Wainwright, C.L.; Stables, C.L.; Saint, D.A.; Clements-Jewery, H.; Lambiase, P.D.; Billman, G.E.; Janse, M.J.; et al. The Lambeth Conventions (II): Guidelines for the study of animal and human ventricular and supraventricular arrhythmias. Pharmacol. Ther. 2013, 139, 213–248. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RMP (mV) | APA (mV) | APD90 (ms) | APD50 (ms) | APD30 (ms) | |
|---|---|---|---|---|---|
| Control (17/10) | −64.0 ± 6.7 | 72.4 ± 7.8 | 24.7 ± 4.5 | 9.0 ± 2.1 | 6.3 ± 1.2 |
| Diabetic Plasma (8/4) | −63.0 ± 8.2 | 74.7 ± 8.0 | 57.4 ± 14.5 * | 20.7 ± 5.6 * | 13.0 ± 3.9 * |
| Diabetic Plasma Blocked (13/4) | −64.4 ± 2.0 | 68.4 ± 4.4 | 33.3 ± 3.2 | 8.3 ± 1.9 # | 5.2 ± 1.5 # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zayas-Arrabal, J.; Alquiza, A.; Tuncay, E.; Turan, B.; Gallego, M.; Casis, O. Molecular and Electrophysiological Role of Diabetes-Associated Circulating Inflammatory Factors in Cardiac Arrhythmia Remodeling in a Metabolic-Induced Model of Type 2 Diabetic Rat. Int. J. Mol. Sci. 2021, 22, 6827. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136827
Zayas-Arrabal J, Alquiza A, Tuncay E, Turan B, Gallego M, Casis O. Molecular and Electrophysiological Role of Diabetes-Associated Circulating Inflammatory Factors in Cardiac Arrhythmia Remodeling in a Metabolic-Induced Model of Type 2 Diabetic Rat. International Journal of Molecular Sciences. 2021; 22(13):6827. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136827
Chicago/Turabian StyleZayas-Arrabal, Julian, Amaia Alquiza, Erkan Tuncay, Belma Turan, Monica Gallego, and Oscar Casis. 2021. "Molecular and Electrophysiological Role of Diabetes-Associated Circulating Inflammatory Factors in Cardiac Arrhythmia Remodeling in a Metabolic-Induced Model of Type 2 Diabetic Rat" International Journal of Molecular Sciences 22, no. 13: 6827. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136827