
5-HT3 Signaling Alters Development of Sacral Neural Crest Derivatives That Innervate the Lower Urinary Tract

 , and
, and
Abstract
:1. Introduction
2. Results
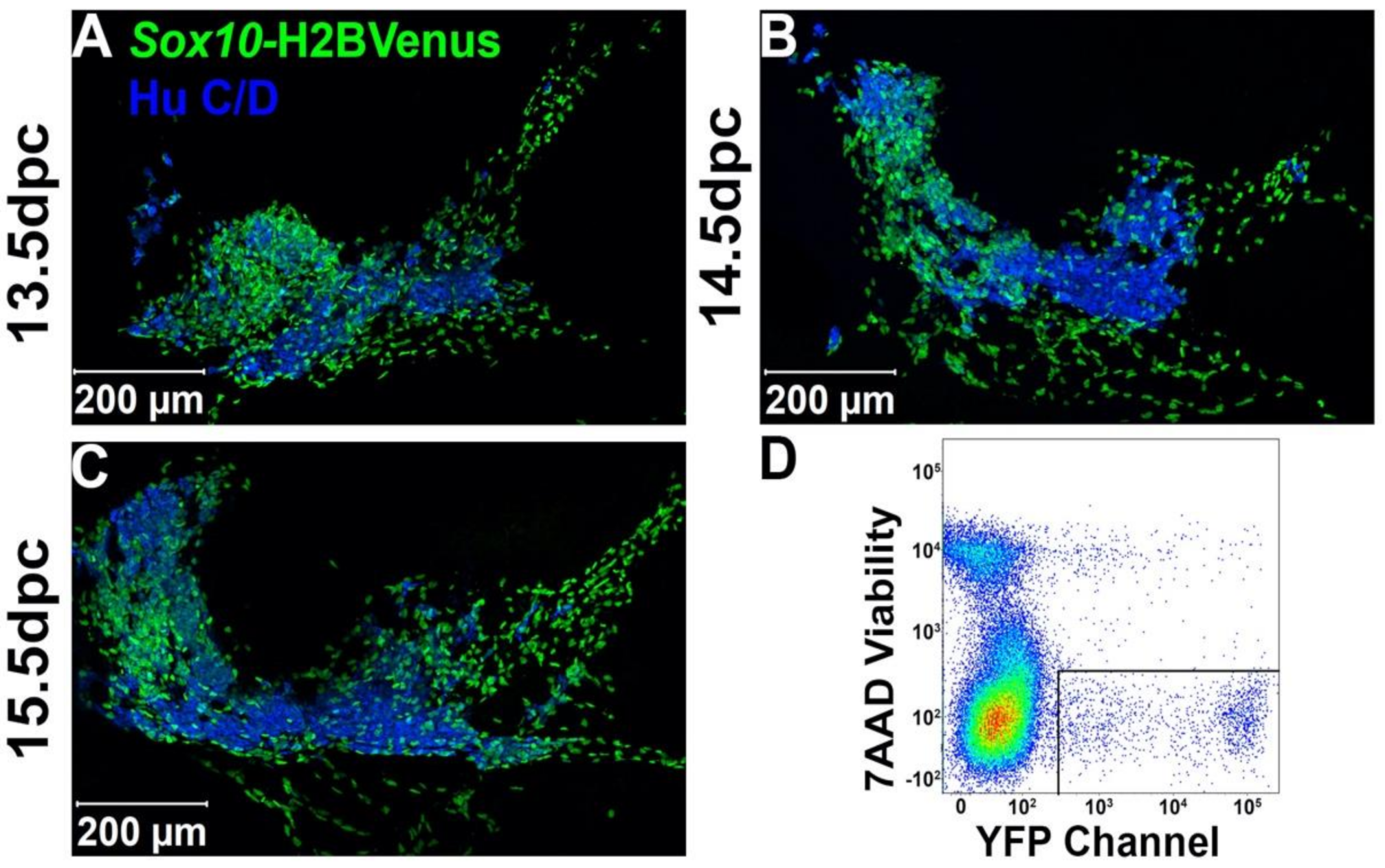
2.1. Sox10+ Neural Crest-Derived Progenitors Are Intermingled with Differentiating Neurons in the Early Pelvic Ganglia
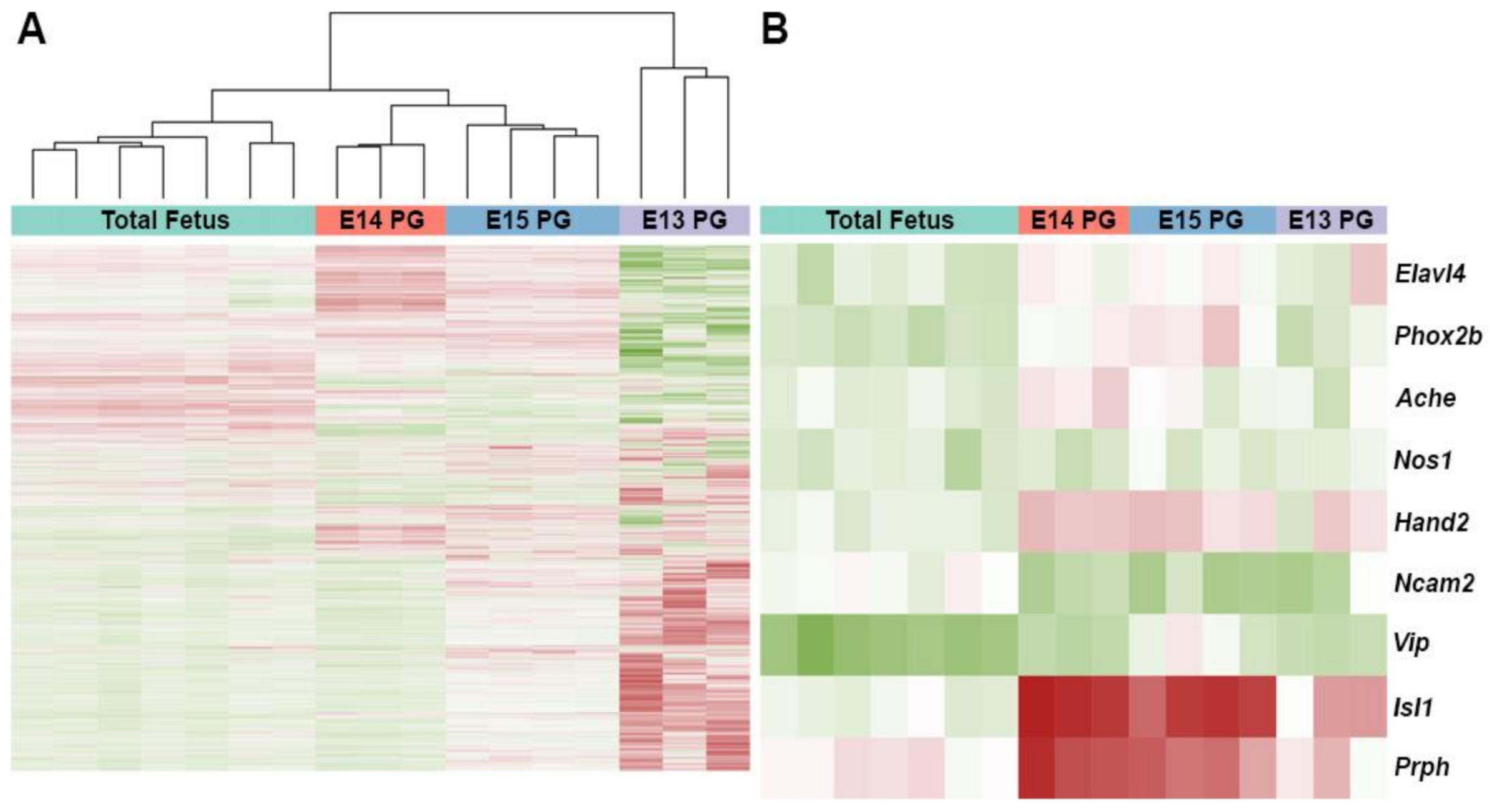
2.2. Expression Profiles of Differentiating Pelvic Ganglia Identify Up-Regulation of Subunits for the Serotonin Receptor 5-HT3
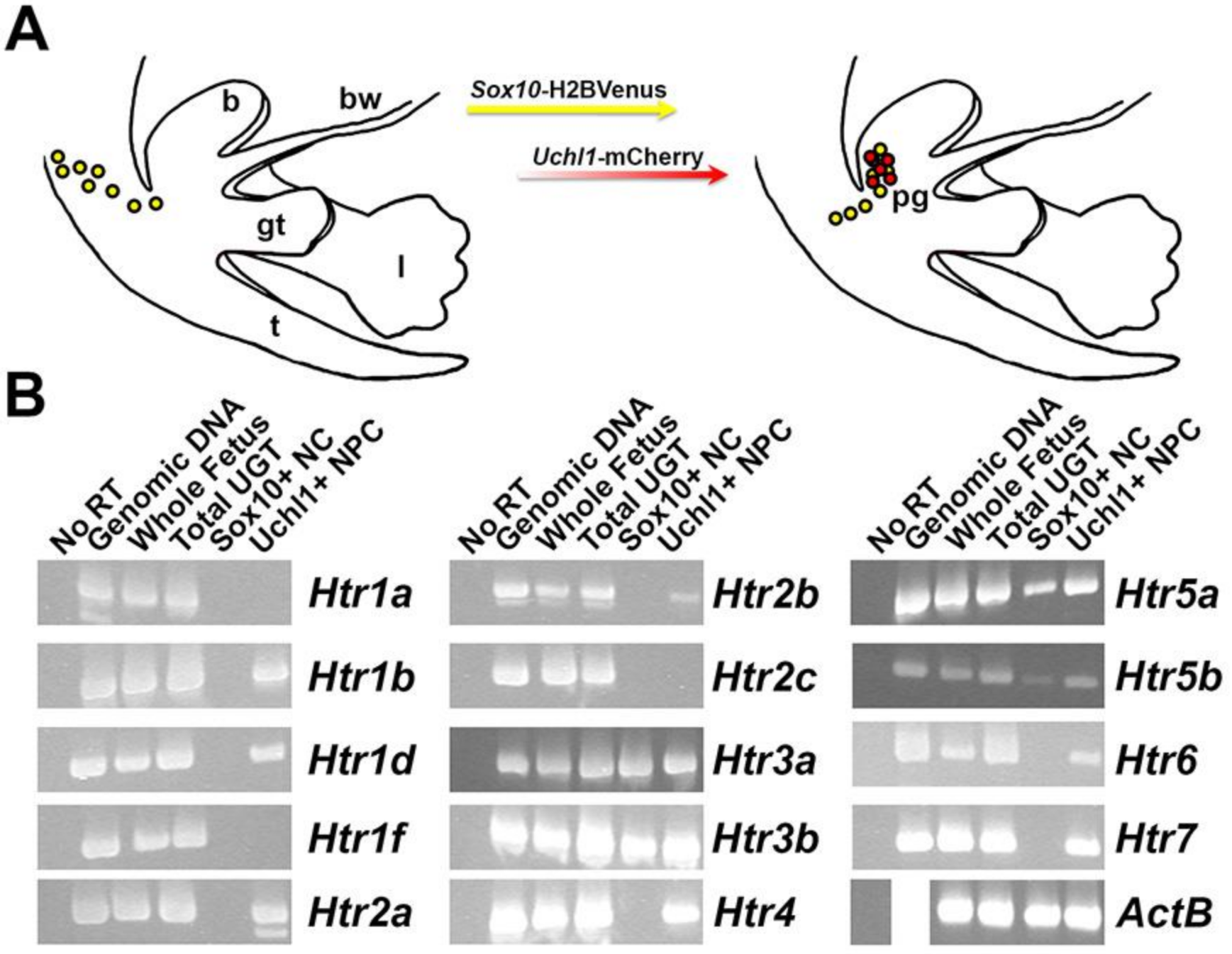
2.3. Multiple Serotonin Receptors Are Expressed during Differentiation of Pelvic Autonomic Neurons
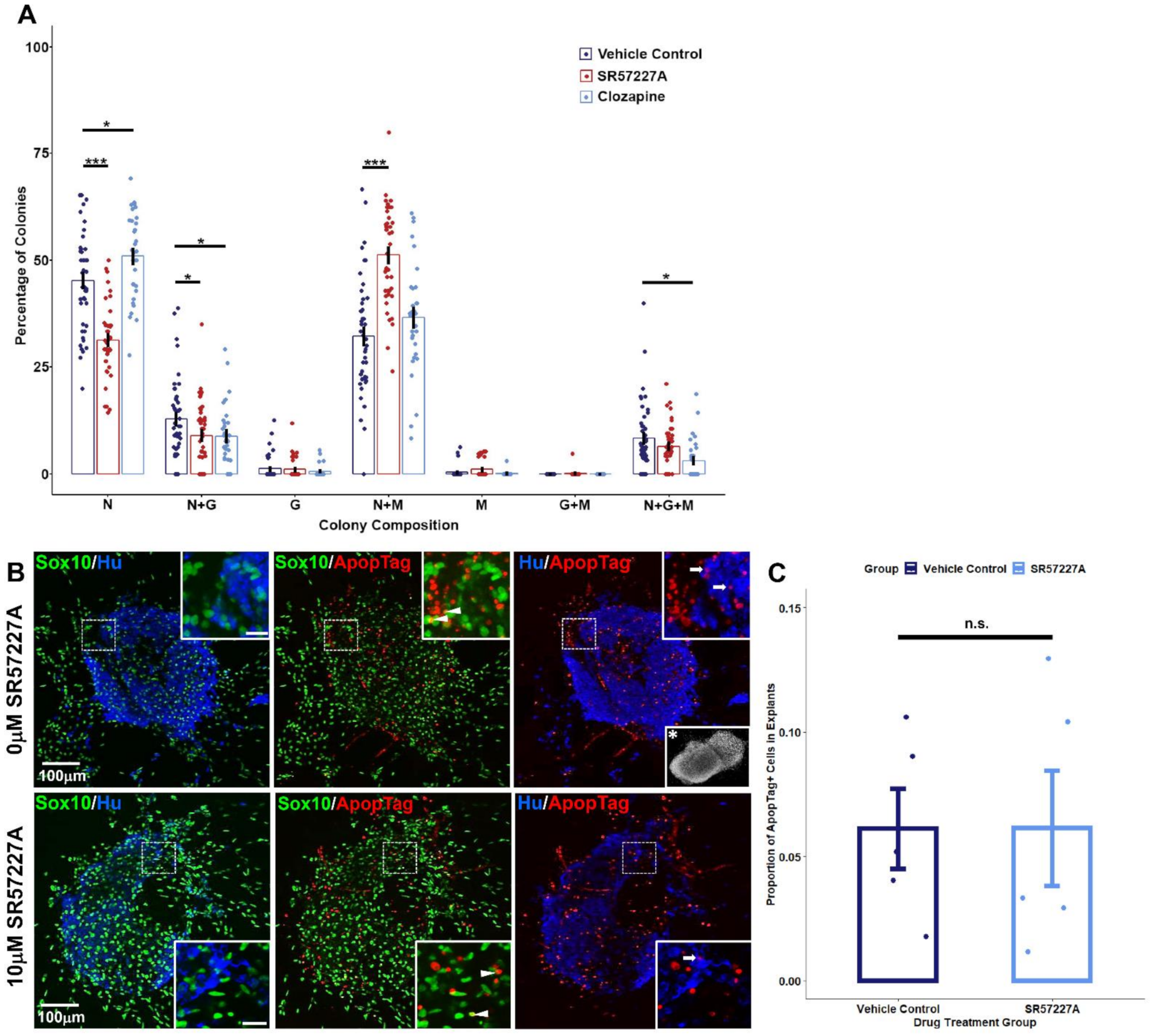
2.4. Perturbing Serotonin Receptor Signaling Alters Differentiation Outcomes of Sox10+ Sacral Neural Crest Progenitors In Vitro
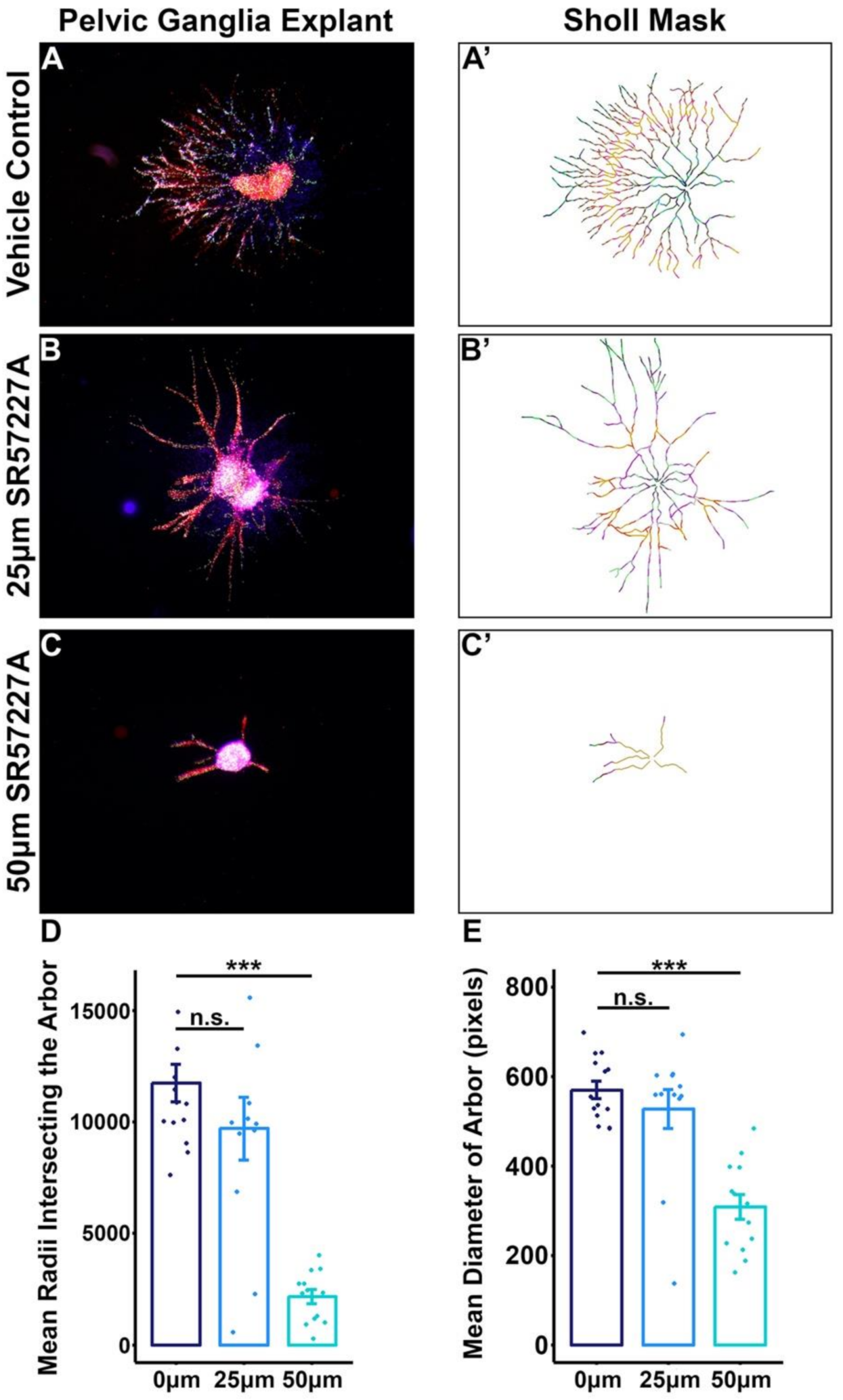
2.5. Over-Stimulating 5-HT3 in Pelvic Ganglia Explants Attenuates Neurite Outgrowth
3. Discussion
4. Materials and Methods
4.1. Mouse Husbandry
4.2. Isolation of mRNA from Fetal Mouse Lower Urinary Tract
4.3. Immunohistochemistry (IHC)
4.4. Microarray Hybridizations and Data Analysis
4.5. Reverse Transcription (RT) PCR
4.6. Assessment of Developmental Potential in Cultures of Sacral Neural Crest Progenitors
4.7. Pelvic Ganglia Explant Cultures
4.8. ApopTag Assay for Apoptosis in Pelvic Ganglia Explant Cultures
4.9. Sholl Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Animal Ethics Statement
References
- Shaban, A.; Drake, M.J.; Hashim, H. The medical management of urinary incontinence. Auton. Neurosci. 2010, 152, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Coyne, K.S.; Wein, A.; Nicholson, S.; Kvasz, M.; Chen, C.-I.; Milsom, I. Economic Burden of Urgency Urinary Incontinence in the United States: A Systematic Review. J. Manag. Care Pharm. 2014, 20, 130–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, S.C.; Belleville, P.J.; Keast, J.R. Regeneration of sensory but not motor axons following visceral nerve injury. Exp. Neurol. 2015, 266, 127–142. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, S.-R.; Song, Y.S.; Lee, H.J. Stem Cell Therapy in Bladder Dysfunction: Where Are We? And Where Do We Have to Go? BioMed Res. Int. 2013, 2013, 930713. [Google Scholar] [CrossRef] [Green Version]
- Wiese, C.B.; Ireland, S.S.; Fleming, N.L.; Yu, J.; Valerius, M.T.; Georgas, K.; Chiu, H.S.; Brennan, J.; Armstrong, J.; Little, M.H.; et al. A Genome-Wide Screen to Identify Transcription Factors Expressed in Pelvic Ganglia of the Lower Urinary Tract. Front. Behav. Neurosci. 2012, 6, 130. [Google Scholar] [CrossRef] [Green Version]
- Stewart, A.L.; Anderson, R.B.; Kobayashi, K.; Young, H.M. Effects of NGF, NT-3 and GDNF family members on neurite outgrowth and migration from pelvic ganglia from embryonic and newborn mice. BMC Dev. Biol. 2008, 8, 73. [Google Scholar] [CrossRef] [Green Version]
- Wanigasekara, Y.; Airaksinen, M.S.; Heuckeroth, R.O.; Milbrandt, J.; Keast, J.R. Neurturin signalling via GFRalpha2 is essential for innervation of glandular but not muscle targets of sacral parasympathetic ganglion neurons. Mol. Cell Neurosci. 2004, 25, 288–300. [Google Scholar] [CrossRef]
- Yan, H.; Keast, J.R. Neurturin regulates postnatal differentiation of parasympathetic pelvic ganglion neurons, initial axonal projections, and maintenance of terminal fields in male urogenital organs. J. Comp. Neurol. 2008, 507, 1169–1183. [Google Scholar] [CrossRef]
- Lake, J.I.; Heuckeroth, R.O. Enteric nervous system development: Migration, differentiation, and disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G1–G24. [Google Scholar] [CrossRef] [Green Version]
- Bondurand, N.; Southard-Smith, E.M. Mouse models of Hirschsprung disease and other developmental disorders of the enteric nervous system: Old and new players. Dev. Biol. 2016, 417, 139–157. [Google Scholar] [CrossRef]
- Young, H.M.; Stamp, L.A.; McKeown, S.J. ENS Development Research Since 1983: Great Strides but Many Remaining Challenges. Adv. Exp. Med. Biol. 2016, 891, 53–62. [Google Scholar] [CrossRef]
- Nagy, N.; Goldstein, A.M. Enteric nervous system development: A crest cell’s journey from neural tube to colon. Semin. Cell Dev. Biol. 2017, 66, 94–106. [Google Scholar] [CrossRef]
- Kasemeier-Kulesa, J.C.; Kulesa, P.M.; Lefcort, F. Imaging neural crest cell dynamics during formation of dorsal root ganglia and sympathetic ganglia. Development 2005, 132, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Kasemeier-Kulesa, J.C.; McLennan, R.; Romine, M.H.; Kulesa, P.M.; Lefcort, F. CXCR4 Controls Ventral Migration of Sympathetic Precursor Cells. J. Neurosci. 2010, 30, 13078–13088. [Google Scholar] [CrossRef]
- Kulesa, P.M.; Gammill, L.S. Neural crest migration: Patterns, phases and signals. Dev. Biol. 2010, 344, 566–568. [Google Scholar] [CrossRef] [Green Version]
- Morrison, J.A.; McLennan, R.; Wolfe, L.A.; Gogol, M.M.; Meier, S.; McKinney, M.C.; Teddy, J.M.; Holmes, L.; Semerad, C.L.; Box, A.C.; et al. Single-cell transcriptome analysis of avian neural crest migration reveals signatures of invasion and molecular transitions. eLife 2017, 6, e28415. [Google Scholar] [CrossRef]
- Saito, D.; Takase, Y.; Murai, H.; Takahashi, Y. The Dorsal Aorta Initiates a Molecular Cascade That Instructs Sympatho-Adrenal Specification. Science 2012, 336, 1578–1581. [Google Scholar] [CrossRef]
- Kameda, Y. Signaling molecules and transcription factors involved in the development of the sympathetic nervous system, with special emphasis on the superior cervical ganglion. Cell Tissue Res. 2014, 357, 527–548. [Google Scholar] [CrossRef]
- Heanue, T.A.; Pachnis, V. Prospective Identification and Isolation of Enteric Nervous System Progenitors Using Sox2. Stem Cells 2011, 29, 128–140. [Google Scholar] [CrossRef] [Green Version]
- Vohra, B.P.; Tsuji, K.; Nagashimada, M.; Uesaka, T.; Wind, D.; Fu, M.; Armon, J.; Enomoto, H.; Heuckeroth, R.O. Differential gene expression and functional analysis implicate novel mechanisms in enteric nervous system precursor migration and neuritogenesis. Dev. Biol. 2006, 298, 259–271. [Google Scholar] [CrossRef] [Green Version]
- Neckel, P.H.; Mohr, R.; Zhang, Y.; Hirt, B.; Just, L. Comparative Microarray Analysis of Proliferating and Differentiating Murine ENS Progenitor Cells. Stem Cells Int. 2016, 2016, 9695827. [Google Scholar] [CrossRef] [Green Version]
- Corpening, J.C.; Deal, K.K.; Cantrell, V.A.; Skelton, S.B.; Buehler, D.P.; Southard-Smith, E.M. Isolation and live imaging of enteric progenitors based on Sox10-Histone2BVenus transgene expression. Genesis 2011, 49, 599–618. [Google Scholar] [CrossRef] [Green Version]
- Wiese, C.B.; Deal, K.K.; Ireland, S.J.; Cantrell, V.A.; Southard-Smith, E.M. Migration pathways of sacral neural crest during development of lower urogenital tract innervation. Dev. Biol. 2017, 429, 356–369. [Google Scholar] [CrossRef]
- Wiese, C.B.; Fleming, N.L.; Buehler, D.P.; Southard-Smith, E.M. A Uchl1-Histone2BmCherry: GFP-gpi BAC transgene for imaging neuronal progenitors. Genesis 2013, 51, 852–861. [Google Scholar] [CrossRef] [Green Version]
- White, P.M.; Morrison, S.; Orimoto, K.; Kubu, C.J.; Verdi, J.M.; Anderson, D.J. Neural Crest Stem Cells Undergo Cell-Intrinsic Developmental Changes in Sensitivity to Instructive Differentiation Signals. Neuron 2001, 29, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Bixby, S.; Kruger, G.M.; Mosher, J.T.; Joseph, N.M.; Morrison, S.J. Cell-Intrinsic Differences between Stem Cells from Different Regions of the Peripheral Nervous System Regulate the Generation of Neural Diversity. Neuron 2002, 35, 643–656. [Google Scholar] [CrossRef] [Green Version]
- Kruger, G.M.; Mosher, J.T.; Bixby, S.; Joseph, N.; Iwashita, T.; Morrison, S.J. Neural Crest Stem Cells Persist in the Adult Gut but Undergo Changes in Self-Renewal, Neuronal Subtype Potential, and Factor Responsiveness. Neuron 2002, 35, 657–669. [Google Scholar] [CrossRef] [Green Version]
- Walters, L.C.; Cantrell, V.A.; Weller, K.P.; Mosher, J.T.; Southard-Smith, E.M. Genetic background impacts developmental potential of enteric neural crest-derived progenitors in the Sox10Dom model of Hirschsprung disease. Hum. Mol. Genet. 2010, 19, 4353–4372. [Google Scholar] [CrossRef] [Green Version]
- Takimoto, Y.; Ishida, Y.; Nakamura, Y.; Kamakura, T.; Yamada, T.; Kondo, M.; Kitahara, T.; Uno, A.; Imai, T.; Horii, A.; et al. 5-HT3 receptor expression in the mouse vestibular ganglion. Brain Res. 2014, 1557, 74–82. [Google Scholar] [CrossRef]
- Edwards, E.; Hampton, E.; Ashby, C.R.; Zhang, J.; Wang, R.Y. 5-HT3-like receptors in the rat medial prefrontal cortex: Further pharmacological characterization. Brain Res. 1996, 733, 21–30. [Google Scholar] [CrossRef]
- Abdel-Aziz, H.; Windeck, T.; Ploch, M.; Verspohl, E.J. Mode of action of gingerols and shogaols on 5-HT3 receptors: Binding studies, cation uptake by the receptor channel and contraction of isolated guinea-pig ileum. Eur. J. Pharmacol. 2006, 530, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Cornu, J.-N.; Melot, C.; Haab, F. A pragmatic approach to the characterization and effective treatment of male patients with postprostatectomy incontinence. Curr. Opin. Urol. 2014, 24, 566–570. [Google Scholar] [CrossRef] [PubMed]
- Laterza, R.M.; Sievert, K.-D.; de Ridder, D.; Vierhout, M.E.; Haab, F.; Cardozo, L.; van Kerrebroeck, P.; Cruz, F.; Kelleher, C.; Chapple, C.; et al. Bladder function after radical hysterectomy for cervical cancer. Neurourol. Urodyn. 2015, 34, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Wit, E.M.K.; Horenblas, S. Urological complications after treatment of cervical cancer. Nat. Rev. Urol. 2014, 11, 110–117. [Google Scholar] [CrossRef]
- Boudes, M.; Uvin, P.; Pinto, S.; Freichel, M.; Birnbaumer, L.; Voets, T.; De Ridder, D.; Vennekens, R. Crucial Role of TRPC1 and TRPC4 in Cystitis-Induced Neuronal Sprouting and Bladder Overactivity. PLoS ONE 2013, 8, e69550. [Google Scholar] [CrossRef]
- Dickson, A.; Avelino, A.; Cruz, F.; Ribeiro-Da-Silva, A. Peptidergic sensory and parasympathetic fiber sprouting in the mucosa of the rat urinary bladder in a chronic model of cyclophosphamide-induced cystitis. Neuroscience 2006, 141, 1633–1647. [Google Scholar] [CrossRef]
- Anderson, R.B.; Stewart, A.L.; Young, H.M. Phenotypes of neural-crest-derived cells in vagal and sacral pathways. Cell Tissue Res. 2005, 323, 11–25. [Google Scholar] [CrossRef]
- Wang, X.; Chan, A.K.; Sham, M.H.; Burns, A.J.; Chan, W.Y. Analysis of the Sacral Neural Crest Cell Contribution to the Hindgut Enteric Nervous System in the Mouse Embryo. Gastroenterology 2011, 141, 992–1002.e6. [Google Scholar] [CrossRef] [Green Version]
- McKeown, S.; Lee, V.M.; Bronner-Fraser, M.; Newgreen, D.F.; Farlie, P.G. Sox10 overexpression induces neural crest-like cells from all dorsoventral levels of the neural tube but inhibits differentiation. Dev. Dyn. 2005, 233, 430–444. [Google Scholar] [CrossRef]
- Martik, M.L.; Bronner, M.E. Regulatory Logic Underlying Diversification of the Neural Crest. Trends Genet. 2017, 33, 715–727. [Google Scholar] [CrossRef] [Green Version]
- Diez-Roux, G.; Banfi, S.; Sultan, M.; Geffers, L.; Anand, S.; Rozado, D.; Magen, A.; Canidio, E.; Pagani, M.; Peluso, I.; et al. A High-Resolution Anatomical Atlas of the Transcriptome in the Mouse Embryo. PLoS Biol. 2011, 9, e1000582. [Google Scholar] [CrossRef]
- Harding, S.D.; Armit, C.; ARmstrong, J.; Brennan, J.; Cheng, Y.; Haggarty, B.; Houghton, D.; Lloyd-MacGilp, S.; Pi, X.; Roochun, Y.; et al. The GUDMAP database--an online resource for genitourinary research. Development 2011, 138, 2845–2853. [Google Scholar] [CrossRef] [Green Version]
- Lauder, J.M.; Krebs, H. Serotonin as a Differentiation Signal in Early Neurogenesis. Dev. Neurosci. 1978, 1, 15–30. [Google Scholar] [CrossRef]
- Bonnin, A.; Torii, M.; Wang, L.; Rakic, P.; Levitt, P. Serotonin modulates the response of embryonic thalamocortical axons to netrin-1. Nat. Neurosci. 2007, 10, 588–597. [Google Scholar] [CrossRef]
- Mazer, C.; Muneyyirci, J.; Taheny, K.; Raio, N.; Borella, A.; Whitaker-Azmitia, P. Serotonin depletion during synaptogenesis leads to decreased synaptic density and learning deficits in the adult rat: A possible model of neurodevelopmental disorders with cognitive deficits. Brain Res. 1997, 760, 68–73. [Google Scholar] [CrossRef]
- Gaspar, P.; Cases, O.; Maroteaux, L. The developmental role of serotonin: News from mouse molecular genetics. Nat. Rev. Neurosci. 2003, 4, 1002–1012. [Google Scholar] [CrossRef]
- Oostland, M.; Sellmeijer, J.; Van Hooft, J.A. Transient expression of functional serotonin 5-HT3receptors by glutamatergic granule cells in the early postnatal mouse cerebellum. J. Physiol. 2011, 589 Pt 20, 4837–4846. [Google Scholar] [CrossRef]
- Oostland, M.; Buijink, M.R.; van Hooft, J.A. Serotonergic control of Purkinje cell maturation and climbing fibre elimination by 5-HT3 receptors in the juvenile mouse cerebellum. J. Physiol. 2013, 591 Pt 7, 1793–1807. [Google Scholar] [CrossRef]
- Van der Velden, L.; van Hooft, J.A.; Chameau, P. Altered dendritic complexity affects firing properties of cortical layer 2/3 pyramidal neurons in mice lacking the 5-HT3A receptor. J. Neurophysiol. 2012, 108, 1521–1528. [Google Scholar] [CrossRef] [Green Version]
- Chameau, P.; Inta, D.; Vitalis, T.; Monyer, H.; Wadman, W.J.; van Hooft, J.A. The N-terminal region of reelin regulates postnatal dendritic maturation of cortical pyramidal neurons. Proc. Natl. Acad. Sci. USA 2009, 106, 7227–7232. [Google Scholar] [CrossRef] [Green Version]
- García-González, D.; Khodosevich, K.; Watanabe, Y.; Rollenhagen, A.; Lübke, J.H.; Monyer, H. Serotonergic Projections Govern Postnatal Neuroblast Migration. Neuron 2017, 94, 534–549.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritter, E.; Wang, Z.; Vezina, C.; Bjorling, D.E.; Southard-Smith, E.M. Serotonin Receptor 5-HT3A Affects Development of Bladder Innervation and Urinary Bladder Function. Front. Neurosci. 2017, 11, 690. [Google Scholar] [CrossRef] [PubMed]
- Derkach, V.; Surprenant, A.; North, R.A. 5-HT3 receptors are membrane ion channels. Nature 1989, 339, 706–709. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, S.S.; Spitzer, N.C. Calcium Signaling in Neuronal Development. Cold Spring Harb. Perspect. Biol. 2011, 3, a004259. [Google Scholar] [CrossRef] [Green Version]
- Deisseroth, K.; Singla, S.; Toda, H.; Monje, M.; Palmer, T.D.; Malenka, R.C. Excitation-Neurogenesis Coupling in Adult Neural Stem/Progenitor Cells. Neuron 2004, 42, 535–552. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Lee, J.; Song, Y.M.; Park, C.; Hwang, S.; Kim, Y.; Kaang, B.; Son, H. Overexpression of calbindin-D 28K in hippocampal progenitor cells increases neuronal differentiation and neurite outgrowth. FASEB J. 2006, 20, 109–111. [Google Scholar] [CrossRef]
- Demarque, M.; Spitzer, N.C. Activity-dependent expression of Lmx1b regulates specification of serotonergic neurons modulating swimming behavior. Neuron 2010, 67, 321–334. [Google Scholar] [CrossRef] [Green Version]
- Guemez-Gamboa, A.; Xu, L.; Meng, D.; Spitzer, N.C. Non-Cell-Autonomous Mechanism of Activity-Dependent Neurotransmitter Switching. Neuron 2014, 82, 1004–1016. [Google Scholar] [CrossRef] [Green Version]
- Lohmann, C.; Wong, R.O. Regulation of dendritic growth and plasticity by local and global calcium dynamics. Cell Calcium 2005, 37, 403–409. [Google Scholar] [CrossRef]
- Wu, G.-Y.; Cline, H.T. Stabilization of Dendritic Arbor Structure in Vivo by CaMKII. Science 1998, 279, 222–226. [Google Scholar] [CrossRef] [Green Version]
- Meltzer, H.Y. An overview of the mechanism of action of clozapine. J. Clin. Psychiatry 1994, 55 (Suppl. B), 47–52. [Google Scholar]
- Schmidt, C.J.; Sorensen, S.M.; Kenne, J.H.; Carr, A.A.; Palfreyman, M.G. The role of 5-HT2A receptors in antipsychotic activity. Life Sci. 1995, 56, 2209–2222. [Google Scholar] [CrossRef]
- Yadav, P.N.; Kroeze, W.K.; Farrell, M.S.; Roth, B.L. Antagonist Functional Selectivity: 5-HT2A Serotonin Receptor Antagonists Differentially Regulate 5-HT2A Receptor Protein Level In Vivo. J. Pharmacol. Exp. Ther. 2011, 339, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Fiorica-Howells, E.; Hen, R.; Gingrich, J.; Li, Z.; Gershon, M.D. 5-HT2A receptors: Location and functional analysis in intestines of wild-type and 5-HT2A knockout mice. Am. J. Physiol. Liver Physiol. 2002, 282, G877–G893. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Dang, H.; Zhu, Q.-M.; Schnegelsberg, B.; Rozengurt, N.; Cain, G.; Prantil, R.; Vorp, D.A.; Guy, N.; Julius, D.; et al. Uropathic Observations in Mice Expressing a Constitutively Active Point Mutation in the 5-HT3A Receptor Subunit. J. Neurosci. 2004, 24, 5537–5548. [Google Scholar] [CrossRef] [Green Version]
- Ansorge, M.S.; Zhou, M.; Lira, A.; Jen, R.; Gingrich, J.A. Early-life blockage of the 5-HT transporter alters emotional behavior in adult mice. Science 2004, 306, 879–881. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Maejima, T.; Wyler, S.C.; Casadesus, G.; Herlitze, S.; Deneris, E.S. Pet-1 is required across different stages of life to regulate serotonergic function. Nat. Neurosci. 2010, 13, 1190–1198. [Google Scholar] [CrossRef]
- Veenstra-VanderWeele, J.; Muller, C.L.; Iwamoto, H.; Sauer, J.E.; Owens, W.A.; Shah, C.R.; Cohen, J.; Mannangatti, P.; Jessen, T.; Thompson, B.J.; et al. Autism gene variant causes hyperserotonemia, serotonin receptor hypersensitivity, social impairment and repetitive behavior. Proc. Natl. Acad. Sci. USA 2012, 109, 5469–5474. [Google Scholar] [CrossRef] [Green Version]
- Garcia, L.P.; Witteveen, J.S.; Middelman, A.; van Hulten, J.A.; Martens, G.J.M.; Homberg, J.R.; Kolk, S.M. Perturbed developmental serotonin signaling affects prefrontal catecholaminergic innervation and cortical integrity. Mol. Neurobiol. 2018, 56, 1405–1420. [Google Scholar] [CrossRef] [Green Version]
- Margolis, K.G.; Li, Z.; Stevanovic, K.; Saurman, V.; Israelyan, N.; Anderson, G.M.; Snyder, I.; Veenstra-VanderWeele, J.; Blakely, R.D.; Gershon, M.D. Serotonin transporter variant drives preventable gastrointestinal abnormalities in development and function. J. Clin. Investig. 2016, 126, 2221–2235. [Google Scholar] [CrossRef] [Green Version]
- Oberlander, T.F.; Gingrich, J.A.; Ansorge, M.S. Sustained Neurobehavioral Effects of Exposure to SSRI Antidepressants During Development: Molecular to Clinical Evidence. Clin. Pharmacol. Ther. 2009, 86, 672–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics 2003, 4, 249–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diboun, I.; Wernisch, L.; Orengo, C.A.; Koltzenburg, M. Microarray analysis after RNA amplification can detect pronounced differences in gene expression using limma. BMC Genom. 2006, 7, 252. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Guo, Y.; Sheng, Q.; Shyr, Y. Advanced Heat Map and Clustering Analysis Using Heatmap3. BioMed Res. Int. 2014, 2014, 986048. [Google Scholar] [CrossRef] [Green Version]
- Meijering, E.; Jacob, M.; Sarria, J.-C.; Steiner, P.; Hirling, H.; Unser, M.; Meijering, E.; Jacob, M.; Sarria, J.-C.; Steiner, P.; et al. Design and validation of a tool for neurite tracing and analysis in fluorescence microscopy images. Cytom. Part A 2004, 58A, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, T.A.; Blackman, A.V.; Oyrer, J.; Jayabal, S.; Chung, A.J.; Watt, A.; Sjostrom, P.; Van Meyel, D.J. Neuronal morphometry directly from bitmap images. Nat. Methods 2014, 11, 982–984. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 13.5 dpc NCP | 14.5 dpc NCP | 15.5 dpc NCP | |||||
|---|---|---|---|---|---|---|---|
| Gene Symbol | mRNA Accession Number | Fold Change | Adjusted p-Value | Fold Change | Adjusted p-Value | Fold Change | Adjusted p-Value |
| Htr1a | NM_008308 | −0.16 | 0.5054 | −0.48 | 0.1293 | −0.07 | 0.8173 |
| Htr1d | ENSMUST00000121571 | 0.21 | 0.4659 | −0.24 | 0.6965 | 0.08 | 0.8384 |
| Htr1f | NM_008310 | 0.73 | 0.0116 | 0.02 | 0.9902 | 0.56 | 0.1074 |
| Htr2a | NM_172812 | 0.47 | 0.1181 | −0.20 | 0.7986 | −0.29 | 0.4381 |
| Htr2b | NM_008311 | −1.41 | 0.0006 | −0.97 | 0.0475 | −1.44 | 0.0011 |
| Htr2c | ENSMUST00000096299 | 1.39 | 2.08 × 10−5 | −0.51 | 0.2132 | −0.06 | 0.8892 |
| Htr3a | NM_013561 | 1.21 | 0.0008 | 2.33 | 2.18 × 10−7 | 2.55 | 4.16 × 10−9 |
| Htr3b | NM_020274 | 0.61 | 0.2917 | 1.79 | 0.0112 | 2.15 | 0.0010 |
| Htr4 | NM_008313 | 0.81 | 0.0047 | 0.08 | 0.9422 | 0.48 | 0.1593 |
| Htr5a | NM_008314 | 0.43 | 0.1824 | −0.07 | 0.9622 | 0.29 | 0.4627 |
| Htr5b | NM_010483 | 1.17 | 1.72 × 10−5 | 0.05 | 0.9626 | 0.24 | 0.4069 |
| Htr6 | NM_021358 | 0.52 | 0.0398 | −0.25 | 0.6064 | 0.44 | 0.1555 |
| Htr7 | NM_008315 | 1.46 | 3.77 × 10−5 | −0.05 | 0.9748 | 0.35 | 0.3744 |
| Genotype | 5′ to 3′ Sequence | Expected Product Size |
|---|---|---|
| Sox10 BAC vector arm insert junction | Sox10 BAC Sp6 Forward: GTTTTTTGCGATCTGCCGTTTC Sox10 BAC Sp6 Reverse: GGCACTTTCATGTTATCTGAGG | 227 bp |
| Sox10 BAC T7 Forward: TCGAGCTTGACATTGTAGGAC Sox10 BAC T7 Reverse: AAGAGCAAGCCTTGGAACTG | 202 bp | |
| Sox10-H2BVenus | Internal Forward: CTGGTCGAGCTCGACGGCGACGTA Internal Reverse: AGTCGCGGCCGCTTTACTTG | 580 bp |
| Uchl1 BAC vector arm insert junction | Uchl1 BAC Sp6 Forward: GCCGTCGACATTTAGGTG Uchl1 BAC Sp6 Reverse: CCTACCCTTCGTCTTCTTTTG | 200 bp |
| Uchl1 BAC T7 Forward: GTGTTGCTTTCTTTGAGTGG Uchl1 BAC T7 Reverse: TACTCAGGATGCTGAAACAGG | 257 bp | |
| Uchl1-H2B | Internal Forward: GTACTAAGGCCGTCACCAAG Internal Reverse: GTACATGAACTGAGGGGACAG | 263 bp |
| Antigen | Host | Vendor, Catalog Number | Dilution | RRID |
|---|---|---|---|---|
| PGP9.5 | Rabbit, polyclonal | AbD Serotec, #7863-0504 | 1:4000 | AB_2210505 |
| Peripherin | Rabbit, polyclonal | Millipore, #AB1530 | 1:1000 | AB_90725 |
| FITC-conjugated FITC | Mouse, monoclonal | Sigma Aldrich, #F3777 | 1:800 | AB_476977 |
| Cy3-conjugated GFAP | Mouse, monoclonal | Sigma Aldrich, #C9205 | 1:800 | AB_476889 |
| HuC/D | Human | Gift of Dr. Vanda Lennon, Mayo Clinic | 1:10,000 | N/A |
| DAPI | N/A | Invitrogen, #D1306 | 1:50,000 | N/A |
| Antigen | Vendor, Catalog Number | Dilution | RRID |
|---|---|---|---|
| Donkey anti-Rabbit AlexaFluor 647 | Jackson ImmunoResearch, #711-605-152 | 1:250 | AB_2492288 |
| Donkey anti-Human AlexaFluor 647 | Jackson ImmunoResearch #709-605-149 | 1:200 | AB_2340578 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ritter, K.E.; Buehler, D.P.; Asher, S.B.; Deal, K.K.; Zhao, S.; Guo, Y.; Southard-Smith, E.M. 5-HT3 Signaling Alters Development of Sacral Neural Crest Derivatives That Innervate the Lower Urinary Tract. Int. J. Mol. Sci. 2021, 22, 6838. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136838
Ritter KE, Buehler DP, Asher SB, Deal KK, Zhao S, Guo Y, Southard-Smith EM. 5-HT3 Signaling Alters Development of Sacral Neural Crest Derivatives That Innervate the Lower Urinary Tract. International Journal of Molecular Sciences. 2021; 22(13):6838. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136838
Chicago/Turabian StyleRitter, K. Elaine, Dennis P. Buehler, Stephanie B. Asher, Karen K. Deal, Shilin Zhao, Yan Guo, and E Michelle Southard-Smith. 2021. "5-HT3 Signaling Alters Development of Sacral Neural Crest Derivatives That Innervate the Lower Urinary Tract" International Journal of Molecular Sciences 22, no. 13: 6838. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136838