
Molecular Basis for the Activation of Human Innate Immune Response by the Flagellin Derived from Plant-Pathogenic Bacterium, Acidovorax avenae



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
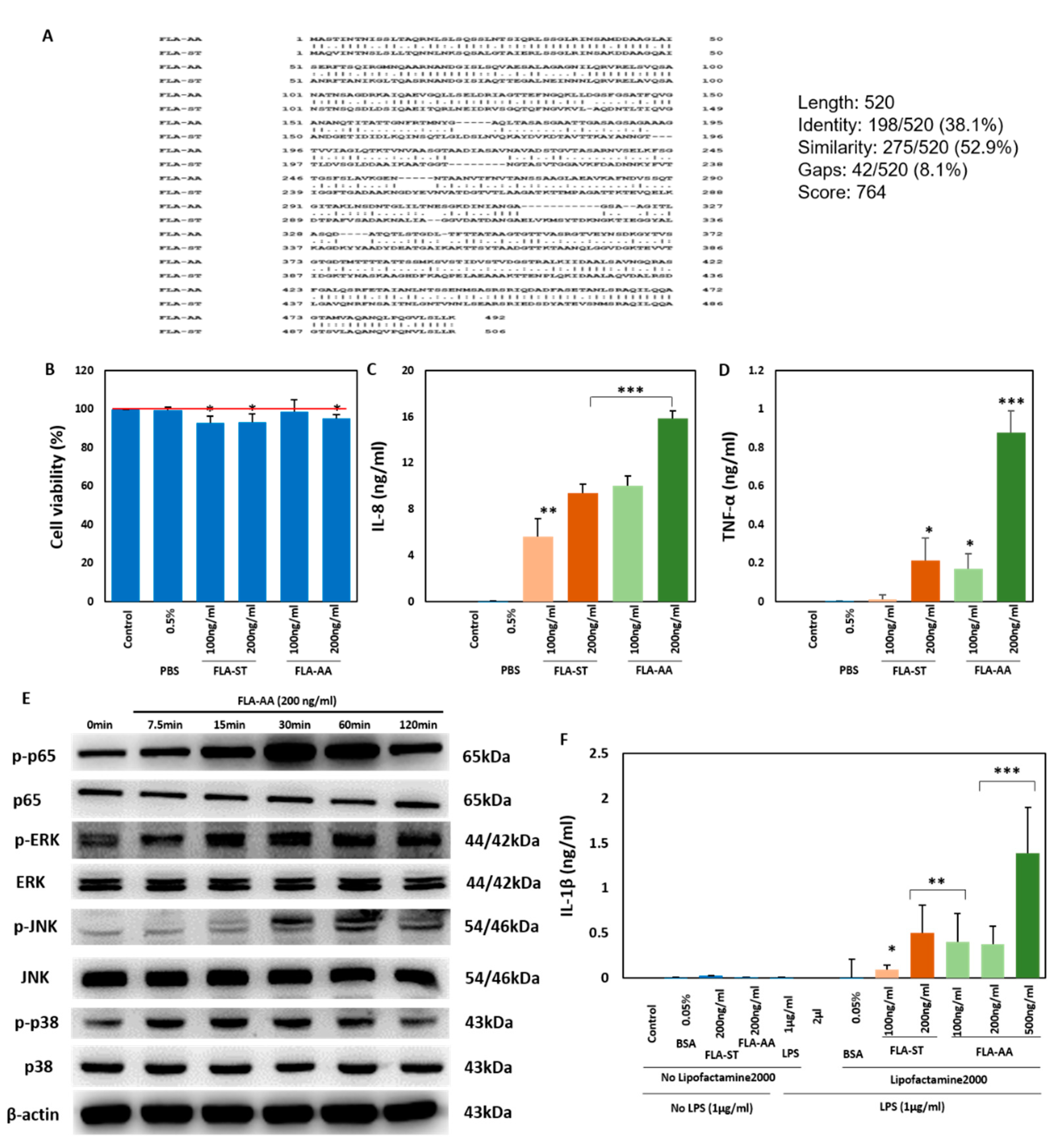
2. Results
2.1. Activation of Immune Signaling in Human Macrophages
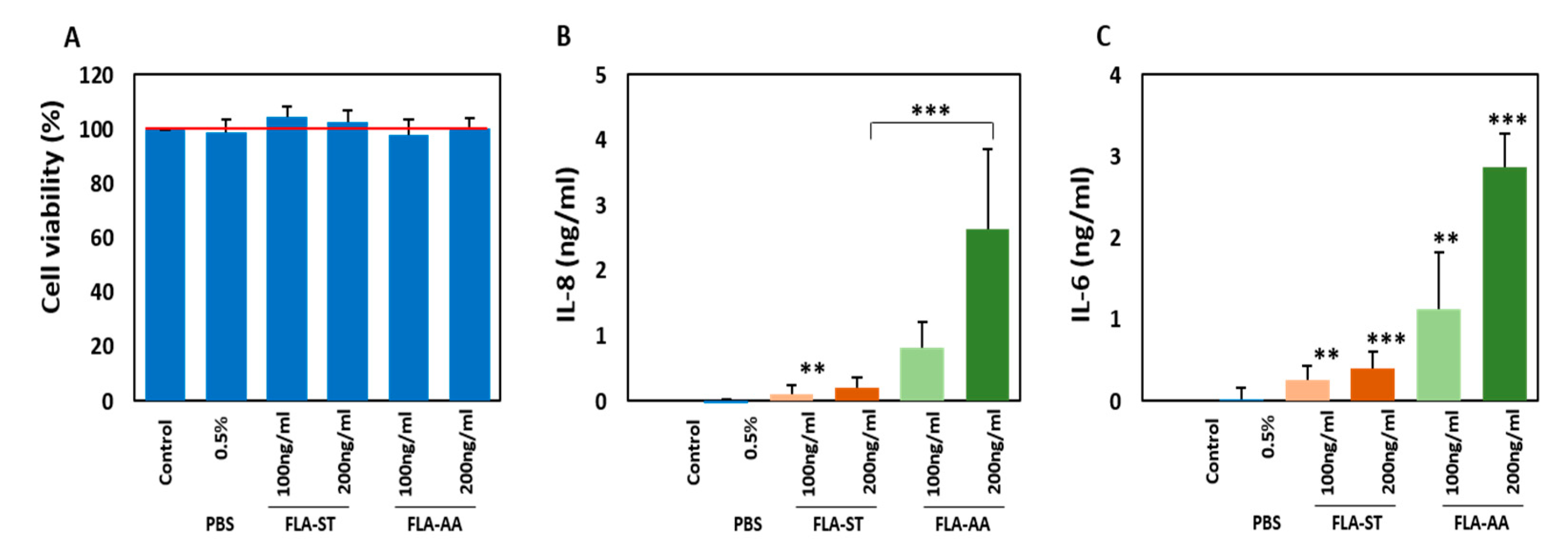
2.2. Secretion of Inflammatory Cytokines by Activated Primary Human Dermal Fibroblasts (HDFs)
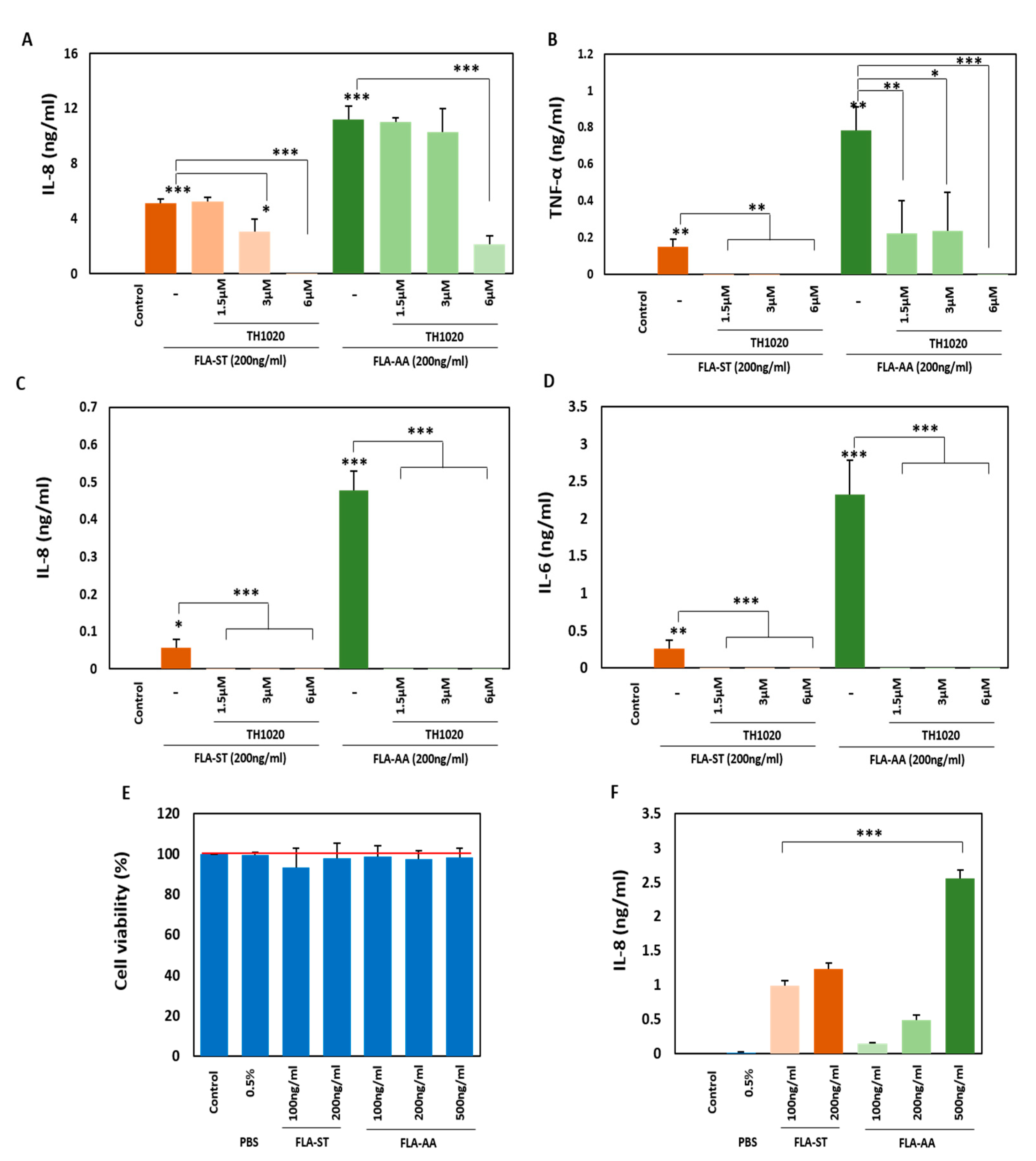
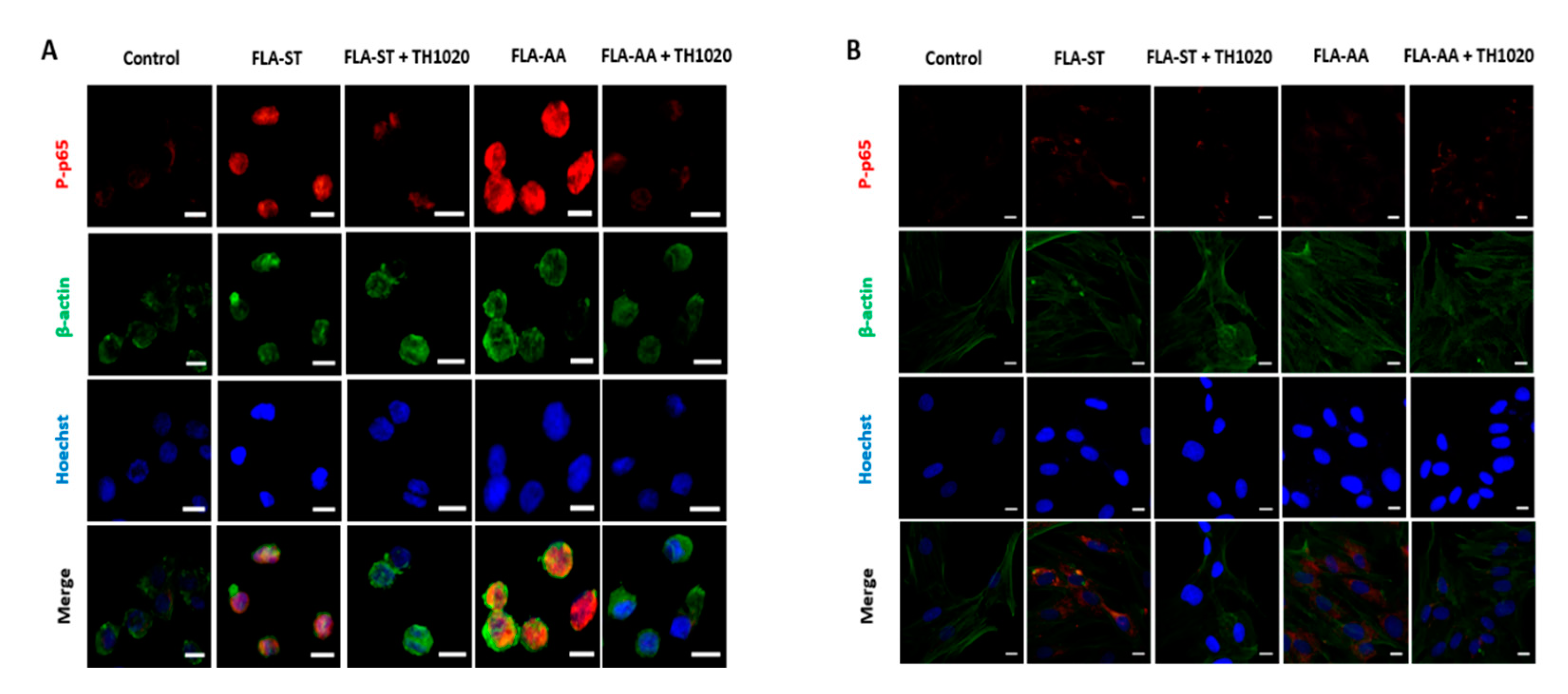
2.3. FLA-AA–Mediated Signaling Is Specific to TLR5
3. Discussion
4. Materials and Methods
4.1. Sequence Alignment
4.2. Cell Lines and Reagents
4.3. Cell Viability Assay
4.4. Western Blot Analyses
4.5. Immunofluorescence
4.6. Cytokine Detection Assays
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Song, W.; Kim, H.; Hwang, C.; Schaad, N. Detection of Acidovorax avenae ssp. avenae in Rice Seeds Using BIO-PCR. J. Phytopathol. 2004, 152, 667–676. [Google Scholar] [CrossRef]
- Lipuma, J.J.; Currie, B.J.; Peacock, S.J.; Vandamme, P.A. Burkholderia, Stenotrophomonas, Ralstonia, Cupriavidus, Pandoraea, Brevundimonas, Comamonas, Delftia, andAcidovorax. In Manual of Clinical Microbiology, 11th ed.; American Society of Microbiology: Washington, DC, USA, 2015; pp. 791–812. [Google Scholar]
- Xie, G.; Sun, X.; Mew, W. Characterization of Acidovorax avenae subsp. avenae from rice seeds. Zhongguo Shuidao Kexue 1998, 12, 165–171. [Google Scholar]
- Xie, G.-L.; Zhang, G.-Q.; Liu, H.; Lou, M.-M.; Tian, W.-X.; Li, B.; Zhou, X.-P.; Zhu, B.; Jin, G.-L. Genome sequence of the rice-pathogenic bacterium Acidovorax avenae subsp. avenae RS-1. J. Bacteriol. 2011, 193, 5013–5014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Moore, J.E.; Millar, B.C.; Alexander, H.D.; McClurg, R.; Morris, T.C.; Rooney, P.J. Improved laboratory diagnosis of bacterial and fungal infections in patients with hematological malignancies using PCR and ribosomal RNA sequence analysis. Leuk. Lymphoma 2004, 45, 1637–1641. [Google Scholar] [CrossRef] [PubMed]
- Shetty, A.; Barnes, R.; Healy, B.; Groves, P. A case of sepsis caused by Acidovorax. J. Infect. 2005, 51, e171–e172. [Google Scholar] [CrossRef] [PubMed]
- Malkan, A.D.; Strollo, W.; Scholand, S.J.; Dudrick, S.J. Implanted-port-catheter-related sepsis caused by Acidovorax avenae and methicillin-sensitive Staphylococcus aureus. J. Clin. Microbiol. 2009, 47, 3358–3361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orsborne, C.; Hardy, A.; Isalska, B.; Williams, S.G.; Muldoon, E.G. Acidovorax oryzae catheter-associated bloodstream infection. J. Clin. Microbiol. 2014, 52, 4421–4424. [Google Scholar] [CrossRef] [Green Version]
- Willems, A.; Falsen, E.; Pot, B.; Jantzen, E.; Hoste, B.; Vandamme, P.; Gillis, M.; Kersters, K.; De Ley, J. Acidovorax, a new genus for Pseudomonas facilis, Pseudomonas delafieldii, E. Falsen (EF) group 13, EF group 16, and several clinical isolates, with the species Acidovorax facilis comb. nov., Acidovorax delafieldii comb. nov., and Acidovorax temperans sp. nov. Int. J. Syst. Evol. Microbiol. 1990, 40, 384–398. [Google Scholar]
- Janeway, C.A. Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harb. Symp. Quant. Biol. 1989, 1–13. [Google Scholar]
- Medzhitov, R.; Preston-Hurlburt, P.; Janeway, C.A., Jr. A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature 1997, 388, 394. [Google Scholar] [CrossRef]
- Uematsu, S.; Akira, S. Toll-Like receptors (TLRs) and their ligands. In Toll-Like Receptors (TLRs) and Innate Immunity; Springer: Berlin/Heidelberg, Germany, 2008; pp. 1–20. [Google Scholar]
- Lien, E.; Sellati, T.J.; Yoshimura, A.; Flo, T.H.; Rawadi, G.; Finberg, R.W.; Carroll, J.D.; Espevik, T.; Ingalls, R.R.; Radolf, J.D. Toll-like receptor 2 functions as a pattern recognition receptor for diverse bacterial products. J. Biol. Chem. 1999, 274, 33419–33425. [Google Scholar] [CrossRef] [Green Version]
- Brightbill, H.D.; Libraty, D.H.; Krutzik, S.R.; Yang, R.-B.; Belisle, J.T.; Bleharski, J.R.; Maitland, M.; Norgard, M.V.; Plevy, S.E.; Smale, S.T. Host defense mechanisms triggered by microbial lipoproteins through toll-like receptors. Science 1999, 285, 732–736. [Google Scholar] [CrossRef] [Green Version]
- Poltorak, A.; He, X.; Smirnova, I.; Liu, M.-Y.; Van Huffel, C.; Du, X.; Birdwell, D.; Alejos, E.; Silva, M.; Galanos, C. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: Mutations in Tlr4 gene. Science 1998, 282, 2085–2088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, J.C.; Young, D.W.; Golenbock, D.T.; Christ, W.J.; Gusovsky, F. Toll-like receptor-4 mediates lipopolysaccharide-induced signal transduction. J. Biol. Chem. 1999, 274, 10689–10692. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Eugene, C.Y.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-κB by Toll-like receptor 3. Nature 2001, 413, 732. [Google Scholar] [CrossRef]
- Diebold, S.S.; Kaisho, T.; Hemmi, H.; Akira, S.; e Sousa, C.R. Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science 2004, 303, 1529–1531. [Google Scholar] [CrossRef]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-specific recognition of single-stranded RNA via toll-like receptor 7 and 8. Science 2004, 303, 1526–1529. [Google Scholar] [CrossRef] [Green Version]
- Jurk, M.; Heil, F.; Vollmer, J.; Schetter, C.; Krieg, A.M.; Wagner, H.; Lipford, G.; Bauer, S. Human TLR7 or TLR8 independently confer responsiveness to the antiviral compound R-848. Nat. Immunol. 2002, 3, 499. [Google Scholar] [CrossRef] [PubMed]
- Hemmi, H.; Takeuchi, O.; Kawai, T.; Kaisho, T.; Sato, S.; Sanjo, H.; Matsumoto, M.; Hoshino, K.; Wagner, H.; Takeda, K. A Toll-like receptor recognizes bacterial DNA. Nature 2000, 408, 740. [Google Scholar] [CrossRef] [PubMed]
- Lund, J.; Sato, A.; Akira, S.; Medzhitov, R.; Iwasaki, A. Toll-like receptor 9–mediated recognition of Herpes simplex virus-2 by plasmacytoid dendritic cells. J. Exp. Med. 2003, 198, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Coban, C.; Ishii, K.J.; Kawai, T.; Hemmi, H.; Sato, S.; Uematsu, S.; Yamamoto, M.; Takeuchi, O.; Itagaki, S.; Kumar, N. Toll-like receptor 9 mediates innate immune activation by the malaria pigment hemozoin. J. Exp. Med. 2005, 201, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ve, T.; Gay, N.J.; Mansell, A.; Kobe, B.; Kellie, S. Adaptors in toll-like receptor signaling and their potential as therapeutic targets. Curr. Drug Targets 2012, 13, 1360–1374. [Google Scholar] [CrossRef]
- Gay, N.J.; Symmons, M.F.; Gangloff, M.; Bryant, C.E. Assembly and localization of Toll-like receptor signalling complexes. Nat. Rev. Immunol. 2014, 14, 546. [Google Scholar] [CrossRef]
- Gómez-Gómez, L.; Boller, T. FLS2: An LRR receptor–like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Mol. Cell 2000, 5, 1003–1011. [Google Scholar] [CrossRef]
- Bauer, Z.; Gómez-Gómez, L.; Boller, T.; Felix, G. Sensitivity of different ecotypes and mutants ofArabidopsis thaliana toward the bacterial elicitor flagellin correlates with the presence of receptor-binding sites. J. Biol. Chem. 2001, 276, 45669–45676. [Google Scholar] [CrossRef] [Green Version]
- Uematsu, S.; Jang, M.H.; Chevrier, N.; Guo, Z.; Kumagai, Y.; Yamamoto, M.; Kato, H.; Sougawa, N.; Matsui, H.; Kuwata, H. Detection of pathogenic intestinal bacteria by Toll-like receptor 5 on intestinal CD11c+ lamina propria cells. Nat. Immunol. 2006, 7, 868. [Google Scholar] [CrossRef]
- Gewirtz, A.T.; Navas, T.A.; Lyons, S.; Godowski, P.J.; Madara, J.L. Cutting edge: Bacterial flagellin activates basolaterally expressed TLR5 to induce epithelial proinflammatory gene expression. J. Immunol. 2001, 167, 1882–1885. [Google Scholar] [CrossRef] [Green Version]
- Miao, E.A.; Alpuche-Aranda, C.M.; Dors, M.; Clark, A.E.; Bader, M.W.; Miller, S.I.; Aderem, A. Cytoplasmic flagellin activates caspase-1 and secretion of interleukin 1β via Ipaf. Nat. Immunol. 2006, 7, 569. [Google Scholar] [CrossRef] [PubMed]
- Amer, A.; Franchi, L.; Kanneganti, T.-D.; Body-Malapel, M.; Özören, N.; Brady, G.; Meshinchi, S.; Jagirdar, R.; Gewirtz, A.; Akira, S. Regulation of Legionella phagosome maturation and infection through flagellin and host Ipaf. J. Biol. Chem. 2006, 281, 35217–35223. [Google Scholar] [CrossRef] [Green Version]
- Miao, E.A.; Leaf, I.A.; Treuting, P.M.; Mao, D.P.; Dors, M.; Sarkar, A.; Warren, S.E.; Wewers, M.D.; Aderem, A. Caspase-1-induced pyroptosis is an innate immune effector mechanism against intracellular bacteria. Nat. Immunol. 2010, 11, 1136. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Batra, S.; Wakamatsu, N.; Pacher, P.; Jeyaseelan, S. NLRC4 inflammasome-mediated production of IL-1β modulates mucosal immunity in the lung against gram-negative bacterial infection. J. Immunol. 2012, 188, 5623–5635. [Google Scholar] [CrossRef] [Green Version]
- Andersen-Nissen, E.; Smith, K.D.; Bonneau, R.; Strong, R.K.; Aderem, A. A conserved surface on Toll-like receptor 5 recognizes bacterial flagellin. J. Exp. Med. 2007, 204, 393–403. [Google Scholar] [CrossRef] [Green Version]
- Keestra, A.M.; de Zoete, M.R.; van Aubel, R.A.; van Putten, J.P. Functional characterization of chicken TLR5 reveals species-specific recognition of flagellin. Mol. Immunol. 2008, 45, 1298–1307. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, J.; Shi, J.; Gong, Y.-N.; Lu, Q.; Xu, H.; Liu, L.; Shao, F. The NLRC4 inflammasome receptors for bacterial flagellin and type III secretion apparatus. Nature 2011, 477, 596–600. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499. [Google Scholar] [CrossRef]
- Goodbourn, S.; Didcock, L.; Randall, R. Interferons: Cell signalling, immune modulation, antiviral response and virus countermeasures. J. Gen. Virol. 2000, 81, 2341–2364. [Google Scholar] [CrossRef]
- Yan, L.; Liang, J.; Yao, C.; Wu, P.; Zeng, X.; Cheng, K.; Yin, H. Pyrimidine Triazole Thioether Derivatives as Toll-Like Receptor 5 (TLR5)/Flagellin Complex Inhibitors. ChemMedChem 2016, 11, 822–826. [Google Scholar] [CrossRef]
- Leifson, E. Atlas of bacterial flagellation. In Atlas of Bacterial Flagellation; Academic Press: Cambridge, MA, USA, 1960. [Google Scholar]
- Ping, L. The asymmetric flagellar distribution and motility of Escherichia coli. J. Mol. Biol. 2010, 397, 906–916. [Google Scholar] [CrossRef]
- Chilcott, G.S.; Hughes, K.T. Coupling of flagellar gene expression to flagellar assembly in Salmonella enterica serovar typhimurium and Escherichia coli. Microbiol. Mol. Biol. Rev. 2000, 64, 694–708. [Google Scholar] [CrossRef] [Green Version]
- Aizawa, S.I.; Kubori, T. Bacterial flagellation and cell division. Genes Cells 1998, 3, 625–634. [Google Scholar] [CrossRef]
- Partridge, J.D.; Harshey, R.M. More than motility: Salmonella flagella contribute to overriding friction and facilitating colony hydration during swarming. J. Bacteriol. 2013, 195, 919–929. [Google Scholar] [CrossRef]
- Duan, Q.; Zhou, M.; Zhu, X.; Bao, W.; Wu, S.; Ruan, X.; Zhang, W.; Yang, Y.; Zhu, J.; Zhu, G. The flagella of F18ab Escherichia coli is a virulence factor that contributes to infection in a IPEC-J2 cell model in vitro. Vet. Microbiol. 2012, 160, 132–140. [Google Scholar] [CrossRef]
- Duan, Q.; Zhou, M.; Zhu, X.; Yang, Y.; Zhu, J.; Bao, W.; Wu, S.; Ruan, X.; Zhang, W.; Zhu, G. Flagella from F18+ Escherichia coli play a role in adhesion to pig epithelial cell lines. Microb. Pathog. 2013, 55, 32–38. [Google Scholar] [CrossRef]
- Arora, S.K.; Ritchings, B.W.; Almira, E.C.; Lory, S.; Ramphal, R. Cloning and characterization of Pseudomonas aeruginosa fliF, necessary for flagellar assembly and bacterial adherence to mucin. Infect. Immun. 1996, 64, 2130–2136. [Google Scholar] [CrossRef] [Green Version]
- Olsen, J.E.; Hoegh-Andersen, K.H.; Casadesús, J.; Rosenkrantz, J.T.; Chadfield, M.S.; Thomsen, L.E. The role of flagella and chemotaxis genes in host pathogen interaction of the host adapted Salmonella enterica serovar Dublin compared to the broad host range serovar S. Typhimurium. BMC Microbiol. 2013, 13, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Crawford, R.W.; Reeve, K.E.; Gunn, J.S. Flagellated but not hyperfimbriated Salmonella enterica serovar Typhimurium attaches to and forms biofilms on cholesterol-coated surfaces. J. Bacteriol. 2010, 192, 2981–2990. [Google Scholar] [CrossRef] [Green Version]
- Allen-Vercoe, E.; Woodward, M.J. The role of flagella, but not fimbriae, in the adherence of Salmonella enterica serotype Enteritidis to chick gut explant. J. Med Microbiol. 1999, 48, 771–780. [Google Scholar] [CrossRef]
- Marchetti, M.; Sirard, J.C.; Sansonetti, P.; Pringault, E.; Kernéis, S. Interaction of pathogenic bacteria with rabbit appendix M cells: Bacterial motility is a key feature in vivo. Microbes Infect. 2004, 6, 521–528. [Google Scholar] [CrossRef]
- La Ragione, R.; Sayers, A.; Woodward, M.J. The role of fimbriae and flagella in the colonization, invasion and persistence of Escherichia coli O78 [ratio] K80 in the day-old-chick model. Epidemiol. Infect. 2000, 124, 351–363. [Google Scholar] [CrossRef]
- Krukonis, E.S.; DiRita, V.J. From motility to virulence: Sensing and responding to environmental signals in Vibrio cholerae. Curr. Opin. Microbiol. 2003, 6, 186–190. [Google Scholar] [CrossRef]
- Arora, S.K.; Neely, A.N.; Blair, B.; Lory, S.; Ramphal, R. Role of motility and flagellin glycosylation in the pathogenesis of Pseudomonas aeruginosa burn wound infections. Infect. Immun. 2005, 73, 4395–4398. [Google Scholar] [CrossRef] [Green Version]
- Lockman, H.A.; Curtiss, R., 3rd. Salmonella typhimurium mutants lacking flagella or motility remain virulent in BALB/c mice. Infect. Immun. 1990, 58, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Gardel, C.L.; Mekalanos, J.J. Alterations in Vibrio cholerae motility phenotypes correlate with changes in virulence factor expression. Infect. Immun. 1996, 64, 2246–2255. [Google Scholar] [CrossRef] [Green Version]
- Food and Drug Administration. Bad bug book, foodborne pathogenic microorganisms and natural toxins. In Gram-positiVe Bacteria, 2nd ed.; Lampel, K., Al-Khaldi, S., Cahill, S., Eds.; Center for Food Safety and Applied Nutrition of the Food and Drug Administration (FDA), US Department of Health and Human Services: Silver Spring, MD, USA, 2012. [Google Scholar]
- Parke, J.L.; Gurian-Sherman, D. Diversity of the Burkholderia cepacia complex and implications for risk assessment of biological control strains. Annu. Rev. Phytopathol. 2001, 39, 225–258. [Google Scholar] [CrossRef]
- Steinkamp, G.; Wiedemann, B.; Rietschel, E.; Krahl, A.; Gielen, J.; Bärmeier, H.; Ratjen, F.; Group, E.B.S. Prospective evaluation of emerging bacteria in cystic fibrosis. J. Cyst. Fibros. 2005, 4, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Vincent, J.-L.; Bihari, D.J.; Suter, P.M.; Bruining, H.A.; White, J.; Nicolas-Chanoin, M.-H.; Wolff, M.; Spencer, R.C.; Hemmer, M. The prevalence of nosocomial infection in intensive care units in Europe: Results of the European Prevalence of Infection in Intensive Care (EPIC) Study. JAMA 1995, 274, 639–644. [Google Scholar] [CrossRef]
- Hirai, H.; Furukawa, T.; Katsuragi, Y.; Che, F.-S. Purification of Flagellin from Acidovorax avenae and Analysis of Plant Immune Responses Induced by the Purified Flagellin. Bio-Protocol 2016, 6. [Google Scholar] [CrossRef]
- Martinon, F.; Burns, K.; Tschopp, J. The inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-β. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef]
- Nagai, H.; Cambronne, E.D.; Kagan, J.C.; Amor, J.C.; Kahn, R.A.; Roy, C.R. A C-terminal translocation signal required for Dot/Icm-dependent delivery of the Legionella RalF protein to host cells. Proc. Natl. Acad. Sci. USA 2005, 102, 826–831. [Google Scholar] [CrossRef] [Green Version]
- Franchi, L.; Amer, A.; Body-Malapel, M.; Kanneganti, T.-D.; Özören, N.; Jagirdar, R.; Inohara, N.; Vandenabeele, P.; Bertin, J.; Coyle, A. Cytosolic flagellin requires Ipaf for activation of caspase-1 and interleukin 1β in salmonella-infected macrophages. Nat. Immunol. 2006, 7, 576–582. [Google Scholar] [CrossRef]
- Sun, Y.-H.; Rolán, H.G.; Tsolis, R.M. Injection of flagellin into the host cell cytosol by Salmonella enterica serotype Typhimurium. J. Biol. Chem. 2007, 282, 33897–33901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, E.A.; Ernst, R.K.; Dors, M.; Mao, D.P.; Aderem, A. Pseudomonas aeruginosa activates caspase 1 through Ipaf. Proc. Natl. Acad. Sci. USA 2008, 105, 2562–2567. [Google Scholar] [CrossRef] [Green Version]
- Warren, S.E.; Mao, D.P.; Rodriguez, A.E.; Miao, E.A.; Aderem, A. Multiple Nod-like receptors activate caspase 1 during Listeria monocytogenes infection. J. Immunol. 2008, 180, 7558–7564. [Google Scholar] [CrossRef] [Green Version]
- Ellenbroek, G.H.; Van Puijvelde, G.H.; Anas, A.A.; Bot, M.; Asbach, M.; Schoneveld, A.; Van Santbrink, P.J.; Foks, A.C.; Timmers, L.; Doevendans, P.A. Leukocyte TLR5 deficiency inhibits atherosclerosis by reduced macrophage recruitment and defective T-cell responsiveness. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Koosakulnirand, S.; Phokrai, P.; Jenjaroen, K.; Roberts, R.A.; Utaisincharoen, P.; Dunachie, S.J.; Brett, P.J.; Burtnick, M.N.; Chantratita, N. Immune response to recombinant Burkholderia pseudomallei FliC. PLoS ONE 2018, 13, e0198906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, E.-J.; Kang, M.-J.; Kim, Y.-S.; Kim, S.-M.; Lee, S.-E.; Kim, C.-H.; Kim, D.-J.; Park, J.-H. Flagellin promotes the proliferation of gastric cancer cells via the Toll-like receptor 5. Int. J. Mol. Med. 2011, 28, 115–119. [Google Scholar]
- Orhan, F.; Bhat, M.; Sandberg, K.; Ståhl, S.; Piehl, F.; Consortium, K.S.P.; Svensson, C.; Erhardt, S.; Schwieler, L.; Farde, L. Tryptophan Metabolism Along the Kynurenine Pathway Downstream of Toll—Like Receptor Stimulation in Peripheral Monocytes. Scand. J. Immunol. 2016, 84, 262–271. [Google Scholar] [CrossRef]
- Weaver, L.K.; Hintz-Goldstein, K.A.; Pioli, P.A.; Wardwell, K.; Qureshi, N.; Vogel, S.N.; Guyre, P.M. Pivotal advance: Activation of cell surface Toll-like receptors causes shedding of the hemoglobin scavenger receptor CD163. J. Leukoc. Biol. 2006, 80, 26–35. [Google Scholar] [CrossRef]
- Zhou, S.-X.; Li, F.-S.; Qiao, Y.-L.; Zhang, X.-Q.; Wang, Z.-D. Toll-like receptor 5 agonist inhibition of growth of A549 lung cancer cells in vivo in a Myd88 dependent manner. Asian Pac. J. Cancer Prev. 2012, 13, 2807–2812. [Google Scholar] [CrossRef] [Green Version]
- Sfondrini, L.; Rossini, A.; Besusso, D.; Merlo, A.; Tagliabue, E.; Mènard, S.; Balsari, A. Antitumor activity of the TLR-5 ligand flagellin in mouse models of cancer. J. Immunol. 2006, 176, 6624–6630. [Google Scholar] [CrossRef]
- Rhee, S.H.; Im, E.; Pothoulakis, C. Toll-like receptor 5 engagement modulates tumor development and growth in a mouse xenograft model of human colon cancer. Gastroenterology 2008, 135, 518–528.e513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdelya, L.G.; Gleiberman, A.S.; Toshkov, I.; Aygun-Sunar, S.; Bapardekar, M.; Manderscheid-Kern, P.; Bellnier, D.; Krivokrysenko, V.I.; Feinstein, E.; Gudkov, A.V. Toll-like receptor 5 agonist protects mice from dermatitis and oral mucositis caused by local radiation: Implications for head-and-neck cancer radiotherapy. Int. J. Radiat. Oncol. 2012, 83, 228–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsuda, A.; Wakui, H.; Iwamoto, K.; Ozawa, M.; Togashi, M.; Masai, R.; Maki, N.; Hatakeyama, T.; Sawada, K. Up-regulated expression of Toll-like receptors mRNAs in peripheral blood mononuclear cells from patients with systemic lupus erythematosus. Clin. Exp. Immunol. 2008, 152, 482–487. [Google Scholar] [CrossRef]
- Bergt, S.; Wagner, N.-M.; Heidrich, M.; Butschkau, A.; Nöldge-Schomburg, G.E.; Vollmar, B.; Roesner, J.P. Hydrocortisone reduces the beneficial effects of toll-like receptor 2 deficiency on survival in a mouse model of polymicrobial sepsis. Shock 2013, 40, 414–419. [Google Scholar] [CrossRef]
- Seung, N.R.; Park, E.J.; Kim, C.W.; Kim, K.H.; Kim, K.J.; Cho, H.J.; Park, H.R. Comparison of expression of heat-shock protein 60, Toll-like receptors 2 and 4, and T-cell receptor γδ in plaque and guttate psoriasis. J. Cutan. Pathol. 2007, 34, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Cavassani, K.A.; Ishii, M.; Wen, H.; Schaller, M.A.; Lincoln, P.M.; Lukacs, N.W.; Hogaboam, C.M.; Kunkel, S.L. TLR3 is an endogenous sensor of tissue necrosis during acute inflammatory events. J. Exp. Med. 2008, 205, 2609–2621. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Yang, Y.; Dai, J.; Medzhitov, R.; Freudenberg, M.A.; Zhang, P.L.; Li, Z. TLR4 up-regulation at protein or gene level is pathogenic for lupus-like autoimmune disease. J. Immunol. 2006, 177, 6880–6888. [Google Scholar] [CrossRef]
- Roger, T.; Froidevaux, C.; Le Roy, D.; Reymond, M.K.; Chanson, A.-L.; Mauri, D.; Burns, K.; Riederer, B.M.; Akira, S.; Calandra, T. Protection from lethal gram-negative bacterial sepsis by targeting Toll-like receptor 4. Proc. Natl. Acad. Sci. USA 2009, 106, 2348–2352. [Google Scholar] [CrossRef] [Green Version]
- Pisitkun, P.; Deane, J.A.; Difilippantonio, M.J.; Tarasenko, T.; Satterthwaite, A.B.; Bolland, S. Autoreactive B cell responses to RNA-related antigens due to TLR7 gene duplication. Science 2006, 312, 1669–1672. [Google Scholar] [CrossRef]
- Leung, P.Y.; Stevens, S.L.; Packard, A.E.; Lessov, N.S.; Yang, T.; Conrad, V.K.; Van Den Dungen, N.N.; Simon, R.P.; Stenzel-Poore, M.P. Toll-like receptor 7 preconditioning induces robust neuroprotection against stroke by a novel type I interferon-mediated mechanism. Stroke 2012, 43, 1383–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilliet, M.; Conrad, C.; Geiges, M.; Cozzio, A.; Thürlimann, W.; Burg, G.; Nestle, F.O.; Dummer, R. Psoriasis triggered by toll-like receptor 7 agonist imiquimod in the presence of dermal plasmacytoid dendritic cell precursors. Arch. Dermatol. 2004, 140, 1490–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, S.W.; Scharping, N.E.; Kolhatkar, N.S.; Khim, S.; Schwartz, M.A.; Li, Q.-Z.; Hudkins, K.L.; Alpers, C.E.; Liggitt, D.; Rawlings, D.J. Opposing impact of B cell–intrinsic TLR7 and TLR9 signals on autoantibody repertoire and systemic inflammation. J. Immunol. 2014, 192, 4525–4532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.; Yang, X.; Xiang, Y.; Li, H.; Yan, H.; Zhou, J.; Caudle, Y.; Zhang, X.; Yin, D. Inhibition of Toll-like receptor 9 attenuates sepsis-induced mortality through suppressing excessive inflammatory response. Cell. Immunol. 2015, 295, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.S.; Sørensen, O.E.; Liu, P.T.; Jalian, H.R.; Eshtiaghpour, D.; Behmanesh, B.E.; Chung, W.; Starner, T.D.; Kim, J.; Sieling, P.A. TGF-α regulates TLR expression and function on epidermal keratinocytes. J. Immunol. 2005, 174, 6137–6143. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Javaid, N.; Hirai, H.; Che, F.-S.; Choi, S. Molecular Basis for the Activation of Human Innate Immune Response by the Flagellin Derived from Plant-Pathogenic Bacterium, Acidovorax avenae. Int. J. Mol. Sci. 2021, 22, 6920. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136920
Javaid N, Hirai H, Che F-S, Choi S. Molecular Basis for the Activation of Human Innate Immune Response by the Flagellin Derived from Plant-Pathogenic Bacterium, Acidovorax avenae. International Journal of Molecular Sciences. 2021; 22(13):6920. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136920
Chicago/Turabian StyleJavaid, Nasir, Hiroyuki Hirai, Fang-Sik Che, and Sangdun Choi. 2021. "Molecular Basis for the Activation of Human Innate Immune Response by the Flagellin Derived from Plant-Pathogenic Bacterium, Acidovorax avenae" International Journal of Molecular Sciences 22, no. 13: 6920. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136920