Chitinases and Chitinase-Like Proteins as Therapeutic Targets in Inflammatory Diseases, with a Special Focus on Inflammatory Bowel Diseases


 , and
, and 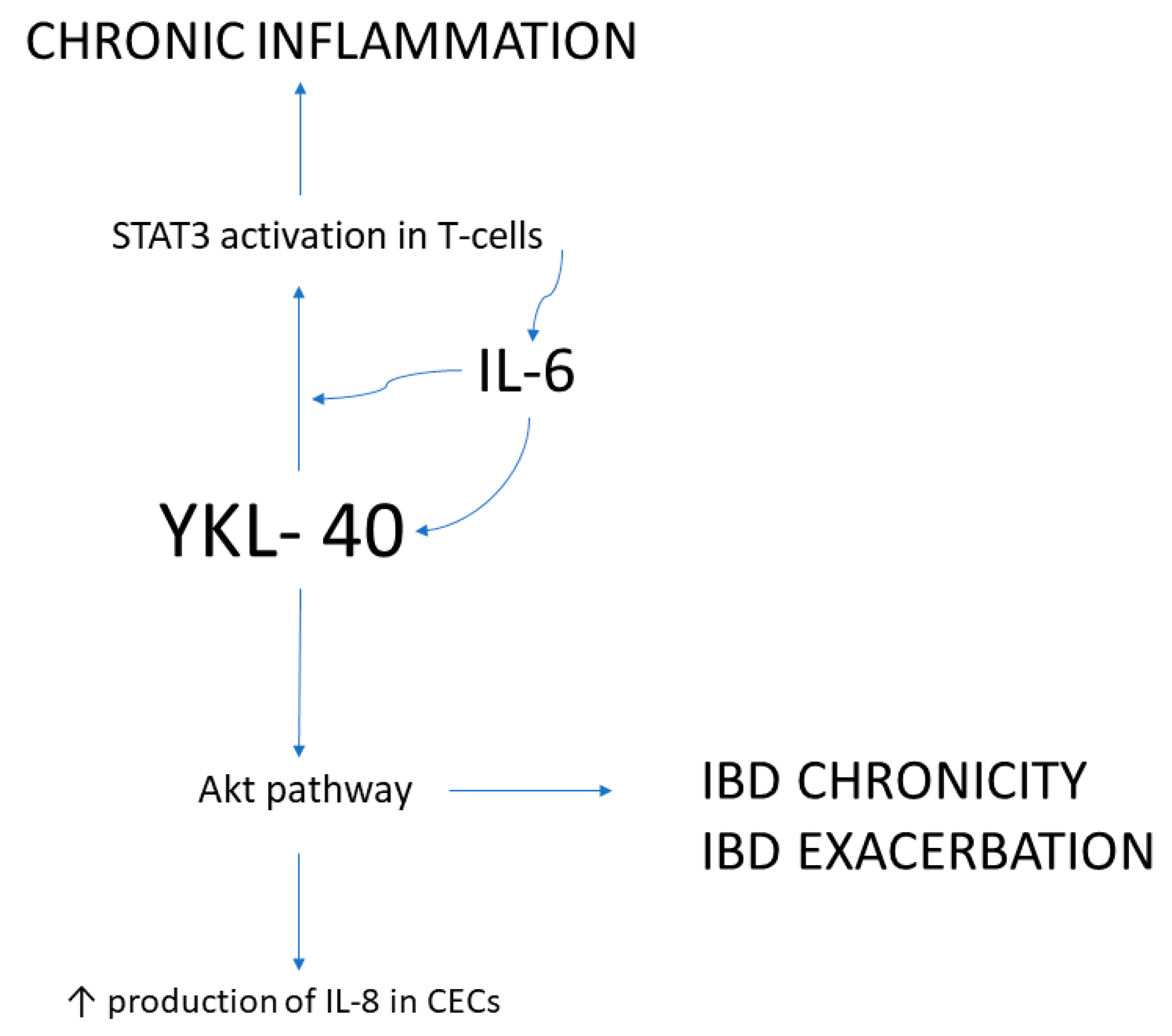{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Chitinases and CLPs
2.1. CHIT1
2.2. AMCase
2.3. CLPs
3. Overview of the Role and Significance of Chitinases in Human Diseases
3.1. Inflammatory Lung Disorders
3.2. Neuroinflammation
3.3. Metabolic Diseases
3.4. Autoinflammatory Disorders
3.5. Gastric Inflammation
4. Pathophysiology of Inflammatory Bowel Diseases (IBDs)
5. Chitinases in IBDs
5.1. CHIT1
5.2. AMCase
5.3. YKL-40
6. A Brief Review of Human Drug-Like Chitinase Inhibitors
7. Chitinase Inhibitors in IBDs and Colorectal Cancer Models
8. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sahai, A.S.; Manocha, M.S. Chitinases of fungi and plants: Their involvement in morphogenesis and host-parasite interaction. FEMS Microbiol. Rev. 1993, 11, 317–338. [Google Scholar] [CrossRef]
- Koga, D.; Sasaki, Y.; Uchiumi, Y.; Hirai, N.; Arakane, Y.; Nagamatsu, Y. Purification and characterization of Bombyx mori chitinases. Insect Biochem. Mol. Biol. 1997, 27, 757–767. [Google Scholar] [CrossRef]
- Fukamizo, T.; Sakai, C.; Tamoi, M. Plant chitinases: Structure-function relationships and their physiology. Foods Food Ingred. J. Jpn. 2003, 208. [Google Scholar]
- Bussink, A.P.; Speijer, D.; Aerts, J.M.F.G.; Boot, R.G. Evolution of Mammalian Chitinase(-Like) Members of Family 18 Glycosyl Hydrolases. Genetics 2007, 177, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, L.; Rosa, M.D.; Zambito, A.M.; Dell’Ombra, N.; Marco, R.D.; Malaguarnera, M. Potential Role of Chitotriosidase Gene in Nonalcoholic Fatty Liver Disease Evolution. Am. J. Gastroenterol. 2006, 101, 2060–2069. [Google Scholar] [CrossRef] [PubMed]
- Steinacker, P.; Verde, F.; Fang, L.; Feneberg, E.; Oeckl, P.; Roeber, S.; Anderl-Straub, S.; Danek, A.; Diehl-Schmid, J.; Fassbender, K.; et al. Chitotriosidase (CHIT1) is increased in microglia and macrophages in spinal cord of amyotrophic lateral sclerosis and cerebrospinal fluid levels correlate with disease severity and progression. J. Neurol. Neurosurg. Psychiatry 2018, 89, 239–247. [Google Scholar] [CrossRef]
- Renkema, H.; Boot, R.; Muijsers, A.O.; Donker-Koopman, W.E.; Aerts, J. Purification and Characterization of Human Chitotriosidase, a Novel Member of the Chitinase Family of Proteins. J. Biol. Chem. 1995, 270, 2198–2202. [Google Scholar] [CrossRef] [Green Version]
- Grosso, S.; Margollicci, M.A.; Bargagli, E.; Buccoliero, Q.R.; Perrone, A.; Galimberti, D.; Morgese, G.; Balestri, P.; Rottoli, P. Serum levels of chitotriosidase as a marker of disease activity and clinical stage in sarcoidosis. Scand. J. Clin. Lab. Investig. 2004, 64, 57–62. [Google Scholar] [CrossRef]
- Létuvé, S.; Kozhich, A.; Humbles, A.; Brewah, Y.; Dombret, M.-C.; Grandsaigne, M.; Adle, H.; Kolbeck, R.; Aubier, M.; Coyle, A.J.; et al. Lung Chitinolytic Activity and Chitotriosidase Are Elevated in Chronic Obstructive Pulmonary Disease and Contribute to Lung Inflammation. Am. J. Pathol. 2010, 176, 638–649. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.; Sa Miranda, M.C.; Amaral, O. Allelic frequency determination of the 24-bp chitotriosidase duplication in the Portuguese population by real-time PCR. Blood Cells Mol. Dis. 2004, 33, 362–364. [Google Scholar] [CrossRef]
- Boot, R.G.; van Achterberg, T.A.E.; van Aken, B.E.; Renkema, G.H.; Jacobs, M.J.H.M.; Aerts, J.M.F.G.; de Vries, C.J.M. Strong induction of members of the chitinase family of proteins in atherosclerosis: Chitotriosidase and human cartilage gp-39 expressed in lesion macrophages. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 687–694. [Google Scholar] [CrossRef] [Green Version]
- Irun, P.; Alfonso, P.; Aznarez, S.; Giraldo, P.; Pocovi, M. Chitotriosidase variants in patients with Gaucher disease. Implications for diagnosis and therapeutic monitoring. Clin. Biochem. 2013, 46, 1804–1807. [Google Scholar] [CrossRef]
- Boot, R.G.; Blommaart, E.F.C.; Swart, E.; der Vlugt, K.G.-V.; Bijl, N.; Moe, C.; Place, A.; Aerts, J. Identification of a Novel Acidic Mammalian Chitinase Distinct from Chitotriosidase. J. Biol. Chem. 2001, 276, 6770–6778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cozzarini, E.; Bellin, M.; Norberto, L.; Polese, L.; Musumeci, S.; Lanfranchi, G.; Paoletti, M.G. CHIT1 and AMCase expression in human gastric mucosa: Correlation with inflammation and Helicobacter pylori infection. Eur. J. Gastroenterol. Hepatol. 2009, 21, 1119–1126. [Google Scholar] [CrossRef]
- Paoletti, M.G.; Norberto, L.; Damini, R.; Musumeci, S. Human Gastric Juice Contains Chitinase That Can Degrade Chitin. Ann. Nutr. Metab. 2007, 51, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Ohno, M.; Togashi, Y.; Tsuda, K.; Okawa, K.; Kamaya, M.; Sakaguchi, M.; Sugahara, Y.; Oyama, F. Quantification of Chitinase mRNA Levels in Human and Mouse Tissues by Real-Time PCR: Species-Specific Expression of Acidic Mammalian Chitinase in Stomach Tissues. PLoS ONE 2013, 8, e67399. [Google Scholar] [CrossRef]
- Okawa, K.; Ohno, M.; Kashimura, A.; Kimura, M.; Kobayashi, Y.; Sakaguchi, M.; Sugahara, Y.; Kamaya, M.; Kino, Y.; Bauer, P.O.; et al. Loss and Gain of Human Acidic Mammalian Chitinase Activity by Nonsynonymous SNPs. Mol. Biol. Evol. 2016, 33, 3183–3193. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.Y.; Rosenthal, P.; Miller, M.; Pham, A.; Aceves, S.; Sakuda, S.; Broide, D.H. Targeting AMCase reduces esophageal eosinophilic inflammation and remodeling in a mouse model of egg induced eosinophilic esophagitis. Int. Immunopharmacol. 2014, 18, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Fadel, F.; Zhao, Y.; Cachau, R.; Cousido-Siah, A.; Ruiz, F.X.; Harlos, K.; Howard, E.; Mitschler, A.; Podjarny, A. New insights into the enzymatic mechanism of human chitotriosidase (CHIT1) catalytic domain by atomic resolution X-ray diffraction and hybrid QM/MM. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 1455–1470. [Google Scholar] [CrossRef] [Green Version]
- Fusetti, F.; Pijning, T.; Kalk, K.H.; Bos, E.; Dijkstra, B.W. Crystal Structure and Carbohydrate-binding Properties of the Human Cartilage Glycoprotein-39. J. Biol. Chem. 2003, 278, 37753–37760. [Google Scholar] [CrossRef] [Green Version]
- Hakala, E.B.; White, C.; Recklies, A.D. Human cartilage gp-39, a major secretory product of articular chondrocytes and synovial cells, is a mammalian member of a chitinase protein family. J. Biol. Chem. 1993, 268, 25803–25810. [Google Scholar] [CrossRef]
- Shackelton, L.M.; Mann, D.M.; Millis, A.J. Identification of a 38-kDa Heparin-binding Glycoprotein (gp38k) in Differentiating Vascular Smooth Muscle Cells as a Member of a Group of Proteins Associated with Tissue Remodeling. J. Biol. Chem. 1995, 270, 13076–13083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, S.; Weisman, M.; O’Dell, J.; Scott, T.; Visor, M.K.J.; Swindlehurst, C. Chondrex: New marker of joint disease. Clin. Chem. 1998, 44, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, A.K.; Singh, G.; Paramasivam, M.; Saravanan, K.; Jabeen, T.; Sharma, S.; Yadav, S.; Kaur, P.; Kumar, P.; Srinivasan, A.; et al. Crystal Structure of a Novel Regulatory 40-kDa Mammary Gland Protein (MGP-40) Secreted during Involution. J. Biol. Chem. 2003, 278, 14451–14460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigg, H.F.; Wait, R.; Rowan, A.D.; Cawston, T.E. The Mammalian Chitinase-like Lectin, YKL-40, Binds Specifically to Type I Collagen and Modulates the Rate of Type I Collagen Fibril Formation. J. Biol. Chem. 2006, 281, 21082–21095. [Google Scholar] [CrossRef] [Green Version]
- Ngernyuang, N.; Yan, W.; Schwartz, L.M.; Oh, D.; Liu, Y.-B.; Chen, H.; Shao, R. A Heparin Binding Motif Rich in Arginine and Lysine is the Functional Domain of YKL-40. Neoplasia 2018, 20, 182–192. [Google Scholar] [CrossRef]
- Johansen, J.S. Studies on serum YKL-40 as a biomarker in diseases with inflammation, tissue remodelling, fibroses and cancer. Dan. Med. Bull. 2006, 53, 172–209. [Google Scholar]
- Johansen, J.S.; Christoffersen, P.; Møller, S.; Price, A.P.; Henriksen, J.H.; Garbarsch, C.; Bendtsen, F. Serum YKL-40 is increased in patients with hepatic fibrosis. J. Hepatol. 2000, 32, 911–920. [Google Scholar] [CrossRef]
- Pelloski, C.E.; Mahajan, A.; Maor, M.H.; Chang, E.L.; Woo, S.; Gilbert, M.R.; Colman, H.; Yang, H.; LeDoux, A.; Blair, H.; et al. YKL-40 Expression is Associated with Poorer Response to Radiation and Shorter Overall Survival in Glioblastoma. Clin. Cancer Res. 2005, 11, 3326–3334. [Google Scholar] [CrossRef] [Green Version]
- Nutt, C.L.; Betensky, R.A.; Brower, M.A.; Batchelor, T.T.; Louis, D.N.; Stemmer-Rachamimov, A.O. YKL-40 Is a Differential Diagnostic Marker for Histologic Subtypes of High-Grade Gliomas. Clin. Cancer Res. 2005, 11, 2258–2264. [Google Scholar] [CrossRef] [Green Version]
- Kawada, M.; Seno, H.; Kanda, K.; Nakanishi, Y.; Akitake, R.; Komekado, H.; Kawada, K.; Sakai, Y.; Mizoguchi, E.; Chiba, T. Chitinase 3-like 1 promotes macrophage recruitment and angiogenesis in colorectal cancer. Oncogene 2012, 31, 3111–3123. [Google Scholar] [CrossRef] [Green Version]
- Connor, J.R.; Dodds, R.A.; Emery, J.G.; Kirkpatrick, R.B.; Rosenberg, M.; Gowen, M. Human cartilage glycoprotein 39 (HC gp-39) mRNA expression in adult and fetal chondrocytes, osteoblasts and osteocytes by in-situ hybridization. Osteoarthr. Cartil. 2000, 8, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Volck, B.; Ostergaard, K.; Johansen, J.; Garbarsch, C.; Price, P.A. The distribution of YKL-40 in osteoarthritic and normal human articular cartilage. Scand. J. Rheumatol. 1999, 28, 171–179. [Google Scholar]
- Kawada, M.; Chen, C.-C.; Arihiro, A.; Nagatani, K.; Watanabe, T.; Mizoguchi, E. Chitinase 3-like-1 enhances bacterial adhesion to colonic epithelial cells through the interaction with bacterial chitin-binding protein. Lab. Investig. 2008, 88, 883–895. [Google Scholar] [CrossRef] [Green Version]
- Mizoguchi, E. Chitinase 3–Like-1 Exacerbates Intestinal Inflammation by Enhancing Bacterial Adhesion and Invasion in Colonic Epithelial Cells. Gastroenterology 2006, 130, 398–411. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Trinh, K.; Figueira, W.F.; Price, P.A. Isolation and Sequence of a Novel Human Chondrocyte Protein Related to Mammalian Members of the Chitinase Protein Family. J. Biol. Chem. 1996, 271, 19415–19420. [Google Scholar] [CrossRef] [Green Version]
- Miyatake, K.; Tsuji, K.; Yamaga, M.; Yamada, J.; Matsukura, Y.; Abula, K.; Sekiya, I.; Muneta, T. Human YKL39 (chitinase 3-like protein 2), an osteoarthritis-associated gene, enhances proliferation and type II collagen expression in ATDC5 cells. Biochem. Biophys. Res. Commun. 2013, 431, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Larionova, I.; Litviakov, N.; Riabov, V.; Zavyalova, M.; Tsyganov, M.; Buldakov, M.; Song, B.; Moganti, K.; Kazantseva, P.; et al. Tumor-associated macrophages in human breast cancer produce new monocyte attracting and pro-angiogenic factor YKL-39 indicative for increased metastasis after neoadjuvant chemotherapy. OncoImmunology 2018, 7, e1436922. [Google Scholar] [CrossRef] [PubMed]
- Malette, B.; Filion, B.; Kan, F.W.K.; Bleau, G.; St-Jacques, S. Hormonal control of the biosynthesis of hamster oviductin. Microsc. Res. Tech. 1995, 31, 470–477. [Google Scholar] [CrossRef]
- Lok, I.H.; Briton-Jones, C.M.; Yuen, P.M.; Haines, C.J. Variable Expression of Oviductin mRNA at Different Stages of Human Reproductive Cycle. J. Assist. Reprod. Genet. 2002, 19, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Arias, E.B.; Verhage, H.G.; Jaffe, R.C. Complementary Deoxyribonucleic Acid Cloning and Molecular Characterization of an Estrogen-Dependent Human Oviductal Glycoprotein1. Biol. Reprod. 1994, 51, 685–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araki, Y.; Nohara, M.; Yoshida-Komiya, H.; Kuramochi, T.; Ito, M.; Hoshi, H.; Shinkai, J.; Sendai, J. Effect of a null mutation of the oviduct-specific glycoprotein gene on mouse fertilization. Biochem. J. 2003, 374, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Kzhyshkowska, J.; Mamidi, S.; Gratchev, A.; Kremmer, E.; Schmuttermaier, C.; Krusell, L.; Haus, G.; Utikal, J.; Schledzewski, K.; Scholtze, J.; et al. Novel stabilin-1 interacting chitinase-like protein (SI-CLP) is up-regulated in alternatively activated macrophages and secreted via lysosomal pathway. Blood 2006, 107, 3221–3228. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhang, J.; Zhu, K.Y. Chitin in Arthropods: Biosynthesis, Modification, and Metabolism. Adv. Exp. Med. Biol. 2019, 1142, 169–207. [Google Scholar] [CrossRef]
- Younes, I.; Rinaudo, M. Chitin and Chitosan Preparation from Marine Sources. Structure, Properties and Applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, T.; Kimoto, H. Bacterial Chitinase System as a Model of Chitin Biodegradation. Adv. Exp. Med. Biol. 2019, 1142, 131–151. [Google Scholar] [CrossRef] [PubMed]
- Nagatani, K.; Wang, S.; Llado, V.; Lau, C.W.; Li, Z.; Mizoguchi, A.; Nagler, C.R.; Shibata, Y.; Reinecker, H.-C.; Mora, R.J.; et al. Chitin microparticles for the control of intestinal inflammation. Inflamm. Bowel Dis. 2012, 18, 1698–1710. [Google Scholar] [CrossRef] [Green Version]
- Van Dyken, S.J.; Liang, H.E.; Naikawadi, R.P.; Woodruff, P.G.; Wolters, P.J.; Erle, D.J.; Locksley, R.M. Spontaneous Chitin Accumulation in Airways and Age-Related Fibrotic Lung Disease. Cell 2017, 169, 497–509. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Zhang, K.Y.J. Human Chitinases: Structure, Function, and Inhibitor Discovery. In Advances in Experimental Medicine and Biology; Springer LLC: New York, NY, USA, 2019; Volume 1142, pp. 221–251. [Google Scholar]
- Ray, A.; Cohn, L. Th2 cells and GATA-3 in asthma: New insights into the regulation of airway inflammation. J. Clin. Investig. 1999, 104, 985–993. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.-J.; Liu, Y.-K.; Liu, C.-L.; Shen, C.-N.; Kuo, M.-L.; Su, C.-C.; Tseng, C.-P.; Yen, T.-C. Inhibition of Acidic Mammalian Chitinase by RNA Interference Suppresses Ovalbumin-Sensitized Allergic Asthma. Hum. Gene Ther. 2009, 20, 1597–1606. [Google Scholar] [CrossRef]
- Zhu, Z.; Zheng, T.; Homer, R.J.; Kim, Y.-K.; Chen, N.Y.; Cohn, L.; Hamid, Q.; Elias, J.A. Acidic Mammalian Chitinase in Asthmatic Th2 Inflammation and IL-13 Pathway Activation. Science 2004, 304, 1678–1682. [Google Scholar] [CrossRef]
- Zhou, Y.; Peng, H.; Sun, H.; Peng, X.; Tang, C.; Gan, Y.; Chen, X.; Mathur, A.; Hu, B.; Slade, M.D.; et al. Chitinase 3-Like 1 Suppresses Injury and Promotes Fibroproliferative Responses in Mammalian Lung Fibrosis. Sci. Transl. Med. 2014, 6. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.G.; Herzog, E.L.; Ahangari, F.; Zhou, Y.; Gulati, I.; Lee, C.-M.; Peng, X.; Feghali-Bostwick, C.; Jimenez, S.A.; Varga, J.; et al. Chitinase 1 is a biomarker for and therapeutic target in scleroderma-associated interstitial lung disease that augments TGF-β1 signaling. J. Immunol. 2012, 189, 2635–2644. [Google Scholar] [CrossRef] [Green Version]
- Kanneganti, M.; Kamba, A.; Mizoguchi, E. Role of Chitotriosidase (Chitinase 1) Under Normal and Disease Conditions. J. Epithel. Biol. Pharmacol. 2012, 5, 1–9. [Google Scholar] [CrossRef]
- Bargagli, E.; Bennett, D.; Maggiorelli, C.; Di Sipio, P.; Margollicci, M.; Bianchi, N.; Rottoli, P. Human Chitotriosidase: A Sensitive Biomarker of Sarcoidosis. J. Clin. Immunol. 2013, 33, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Kzhyshkowska, J.; Gratchev, A.; Goerdt, S. Human Chitinases and Chitinase-Like Proteins as Indicators for Inflammation and Cancer. Biomark. Insights 2007, 2. [Google Scholar] [CrossRef]
- Di Rosa, M.; Dell’Ombra, N.; Zambito, A.M.; Malaguarnera, M.; Nicoletti, F.; Malaguarnera, L. Chitotriosidase and inflammatory mediator levels in Alzheimer’s disease and cerebrovascular dementia. Eur. J. Neurosci. 2006, 23, 2648–2656. [Google Scholar] [CrossRef]
- Hardiman, O.; Al-Chalabi, A.; Chio, A.; Corr, E.M.; Logroscino, G.; Robberecht, W.; Shaw, P.J.; Simmons, Z.; van den Berg, L.H. Amyotrophic lateral sclerosis. Nat. Rev. Dis. Primers 2017, 3, 17071. [Google Scholar] [CrossRef] [PubMed]
- Varghese, A.M.; Ghosh, M.; Bhagat, S.K.; Vijayalakshmi, K.; Preethish-Kumar, V.; Vengalil, S.; Chevula, P.C.R.; Nashi, S.; Polavarapu, K.; Sharma, M.; et al. Chitotriosidase, a biomarker of amyotrophic lateral sclerosis, accentuates neurodegeneration in spinal motor neurons through neuroinflammation. J. Neuroinflamm. 2020, 17, 232. [Google Scholar] [CrossRef]
- Sanfilippo, C.; Longo, A.M.; Lazzara, F.; Cambria, D.; Distefano, G.; Palumbo, M.; Cantarella, A.; Malaguarnera, L.; Di Rosa, M. CHI3L1 and CHI3L2 overexpression in motor cortex and spinal cord of sALS patients. Mol. Cell. Neurosci. 2017, 85, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Żurawska-Płaksej, E.; Knapik-Kordecka, M.; Rorbach-Dolata, A.; Piwowar, A. Increased chitotriosidase activity in plasma of patients with type 2 diabetes. Arch. Med. Sci. 2016, 12, 977–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathcke, C.N.; Johansen, J.S.; Vestergaard, H. YKL-40, a biomarker of inflammation, is elevated in patients with type 2 diabetes and is related to insulin resistance. Inflamm. Res. 2006, 55, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Saitou, Y.; Shiraki, K.; Yamanaka, Y.; Yamaguchi, Y.; Kawakita, T.; Yamamoto, N.; Sugimoto, K.; Murata, K.; Nakano, T. Noninvasive estimation of liver fibrosis and response to interferon therapy by a serum fibrogenesis marker, YKL-40, in patients with HCV-associated liver disease. World J. Gastroenterol. 2005, 11, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, L.; Di Rosa, M.; Zambito, A.M.; Dell’Ombra, N.; Nicoletti, F. Chitotriosidase gene expression in Kupffer cells from patients with non-alcoholic fatty liver disease. Gut 2006, 55, 1313–1320. [Google Scholar] [CrossRef]
- Knorr, T.; Obermayr, F.; Bartnik, E.; Zien, A.; Aigner, T. YKL-39 (chitinase 3-like protein 2), but not YKL-40 (chitinase 3-like protein 1), is up regulated in osteoarthritic chondrocytes. Ann. Rheum. Dis. 2003, 62, 995–998. [Google Scholar] [CrossRef] [Green Version]
- Kamba, A.; Lee, I.-A.; Mizoguchi, E. Potential Association Between TLR4 and Chitinase 3-Like 1 (CHI3L1/YKL-40) Signaling on Colonic Epithelial Cells in Inflammatory Bowel Disease and Colitis-Associated Cancer. Curr. Mol. Med. 2013, 13, 1110–1121. [Google Scholar] [CrossRef] [Green Version]
- Zielińska, A.; Sałaga, M.; Włodarczyk, M.; Fichna, J. Chronic abdominal pain in irritable bowel syndrome—Current and future therapies. Expert Rev. Clin. Pharmacol. 2018, 11, 729–739. [Google Scholar] [CrossRef]
- Deutschmann, C.; Sowa, M.; Murugaiyan, J.; Roesler, U.; Röber, N.; Conrad, K.; Laass, M.W.; Bogdanos, D.; Sipeki, N.; Papp, M.; et al. Identification of Chitinase-3-Like Protein 1 as a Novel Neutrophil Antigenic Target in Crohn’s Disease. J. Crohns Colitics 2019, 13, 894–904. [Google Scholar] [CrossRef] [Green Version]
- Kawada, M.; Hachiya, Y.; Arihiro, A.; Mizoguchi, E. Role of mammalian chitinases in inflammatory conditions. Keio J. Med. 2007, 56, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Wéra, O.; Lancellotti, P.; Oury, C. The Dual Role of Neutrophils in Inflammatory Bowel Diseases. J. Clin. Med. 2016, 5, 118. [Google Scholar] [CrossRef]
- Schulte-Pelkum, J.; Radice, A.; Norman, G.L.; Hoyos, M.L.; Lakos, G.; Buchner, C.; Musset, L.; Miyara, M.; Stinton, L.; Mahler, M. Novel Clinical and Diagnostic Aspects of Antineutrophil Cytoplasmic Antibodies. J. Immunol. Res. 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.-A.; Low, D.; Kamba, A.; Lladó, V.; Mizoguchi, E. Oral caffeine administration ameliorates acute colitis by suppressing chitinase 3-like 1 expression in intestinal epithelial cells. J. Gastroenterol. 2014, 49, 1206–1216. [Google Scholar] [CrossRef] [Green Version]
- Tran, H.T.; Barnich, N.; Mizoguchi, E. Potential role of chitinases and chitin-binding proteins in host-microbial interactions during the development of intestinal inflammation. Histol. Histopathol. 2011, 26, 1453–1464. [Google Scholar] [CrossRef]
- Lee, I.-A.; Kamba, A.; Low, D.; Mizoguchi, E. Novel methylxanthine derivative-mediated anti-inflammatory effects in inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 1127. [Google Scholar] [CrossRef]
- Ziatabar, S.; Zepf, J.; Rich, S.; Danielson, B.T.; Bollyky, P.I.; Stern, R. Chitin, chitinases, and chitin lectins: Emerging roles in human pathophysiology. Pathophysiology 2018, 25, 253–262. [Google Scholar] [CrossRef]
- Vannella, K.M.; Ramalingam, T.R.; Hart, K.M.; Prado, R.D.Q.; Sciurba, J.; Barron, L.; A Borthwick, L.; Smith, A.D.; Mentink-Kane, M.; White, S.; et al. Acidic chitinase primes the protective immune response to gastrointestinal nematodes. Nat. Immunol. 2016, 17, 538–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Rosa, M.; Distefano, G.; Zorena, K.; Malaguarnera, L. Chitinases and immunity: Ancestral molecules with new functions. Immunobiology 2015, 221, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.G.; Hartl, D.; Lee, G.R.; Koller, B.; Matsuura, H.; Da Silva, C.A.; Sohn, M.H.; Cohn, L.; Homer, R.J.; Kozhich, A.A.; et al. Role of breast regression protein 39 (BRP-39)/chitinase 3-like-1 in Th2 and IL-13–induced tissue responses and apoptosis. J. Exp. Med. 2009, 206, 1149–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, H.T.; Lee, I.-A.; Low, D.; Kamba, A.; Mizoguchi, A.; Shi, H.N.; Lee, C.G.; Elias, J.A.; Mizoguchi, E. Chitinase 3-like 1 Synergistically Activates IL6-mediated STAT3 Phosphorylation in Intestinal Epithelial Cells in Murine Models of Infectious Colitis. Inflamm. Bowel Dis. 2014, 20, 835–846. [Google Scholar] [CrossRef]
- Vieira, A.; Fang, C.B.; Rolim, E.G.; Klug, W.A.; Steinwurz, F.; Rossini, L.G.B.; Candelaria, P.A. Inflammatory bowel disease activity assessed by fecal calprotectin and lactoferrin: Correlation with laboratory parameters, clinical, endoscopic and histological indexes. BMC Res. Notes 2009, 2, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vind Johansen, I.J.S.; Price, P.A.; Munkholm, P. Serum YKL-40, a potential new marker of disease activity in patients with inflammatory bowel disease. Scand. J. Gastroenterol. 2003, 38, 599–605. [Google Scholar]
- Limburg, P.J.; Ahlquist, D.A.; Sandborn, W.J.; Mahoney, D.W.; Devens, M.E.; Harrington, J.J.; Zinsmeister, A.R. Fecal calprotectin levels predict colorectal inflammation among patients with chronic diarrhea referred for colonoscopy. Am. J. Gastroenterol. 2000, 95, 2831–2837. [Google Scholar] [CrossRef]
- Aomatsu, T.; Imaeda, H.; Matsumoto, K.; Kimura, E.; Yoden, A.; Tamai, H.; Fujiyama, Y.; Mizoguchi, E.; Andoh, A. Faecal chitinase 3-like-1: A novel biomarker of disease activity in paediatric inflammatory bowel disease. Aliment. Pharmacol. Ther. 2011, 34, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Koutroubakis, I.E.; Petinaki, E.; Dimoulios, P.; Vardas, E.; Roussomoustakaki, M.; Maniatis, A.N.; Kouroumalis, E.A. Increased serum levels of YKL-40 in patients with inflammatory bowel disease. Int. J. Colorectal Dis. 2003, 18, 254–259. [Google Scholar] [CrossRef]
- Von Lampe, B.; Barthel, B.; Coupland, S.E.; Riecken, E.O.; Rosewicz, S. Differential expression of matrix metalloproteinases and their tissue inhibitors in colon mucosa of patients with inflammatory bowel disease. Gut 2000, 47, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Bernardi, D.; Podswiadek, M.; Zaninotto, M.; Punzi, L.; Plebani, M. YKL-40 as a Marker of Joint Involvement in Inflammatory Bowel Disease. Clin. Chem. 2003, 49, 1685–1688. [Google Scholar] [CrossRef] [Green Version]
- Gaballa, H.A.; El-shahawy, E.D.; El-sayed, H.M. Serum YKL-40 as a Marker of Articular Manifestations in Inflammatory Bowel Disease. Coron. Artery Dis. 2007, 16, 293–300. [Google Scholar]
- Chen, C.-C.; Pekow, J.; Llado, V.; Kanneganti, M.; Lau, C.W.; Mizoguchi, A.; Mino-Kenudson, M.; Bissonnette, M.; Mizoguchi, E. Chitinase 3-Like-1 Expression in Colonic Epithelial Cells as a Potentially Novel Marker for Colitis-Associated Neoplasia. Am. J. Pathol. 2011, 179, 1494–1503. [Google Scholar] [CrossRef] [PubMed]
- Catalán, V.; Gómez-Ambrosi, J.; Rodríguez, A.; Ramírez, B.; Izaguirre, M.; Hernández-Lizoain, J.L.; Baixauli, J.; Marti, P.; Valentí, V.; Moncada, R.; et al. Increased Obesity-Associated Circulating Levels of the Extracellular Matrix Proteins Osteopontin, Chitinase-3 Like-1 and Tenascin C Are Associated with Colon Cancer. PLoS ONE 2016, 11, e0162189. [Google Scholar] [CrossRef] [PubMed]
- Eurich, K.; Segawa, M.; Toei-Shimizu, S.; Mizoguchi, E. Potential role of chitinase 3-like-1 in inflammation-associated carcinogenic changes of epithelial cells. World J. Gastroenterol. 2009, 15, 5249–5259. [Google Scholar] [CrossRef]
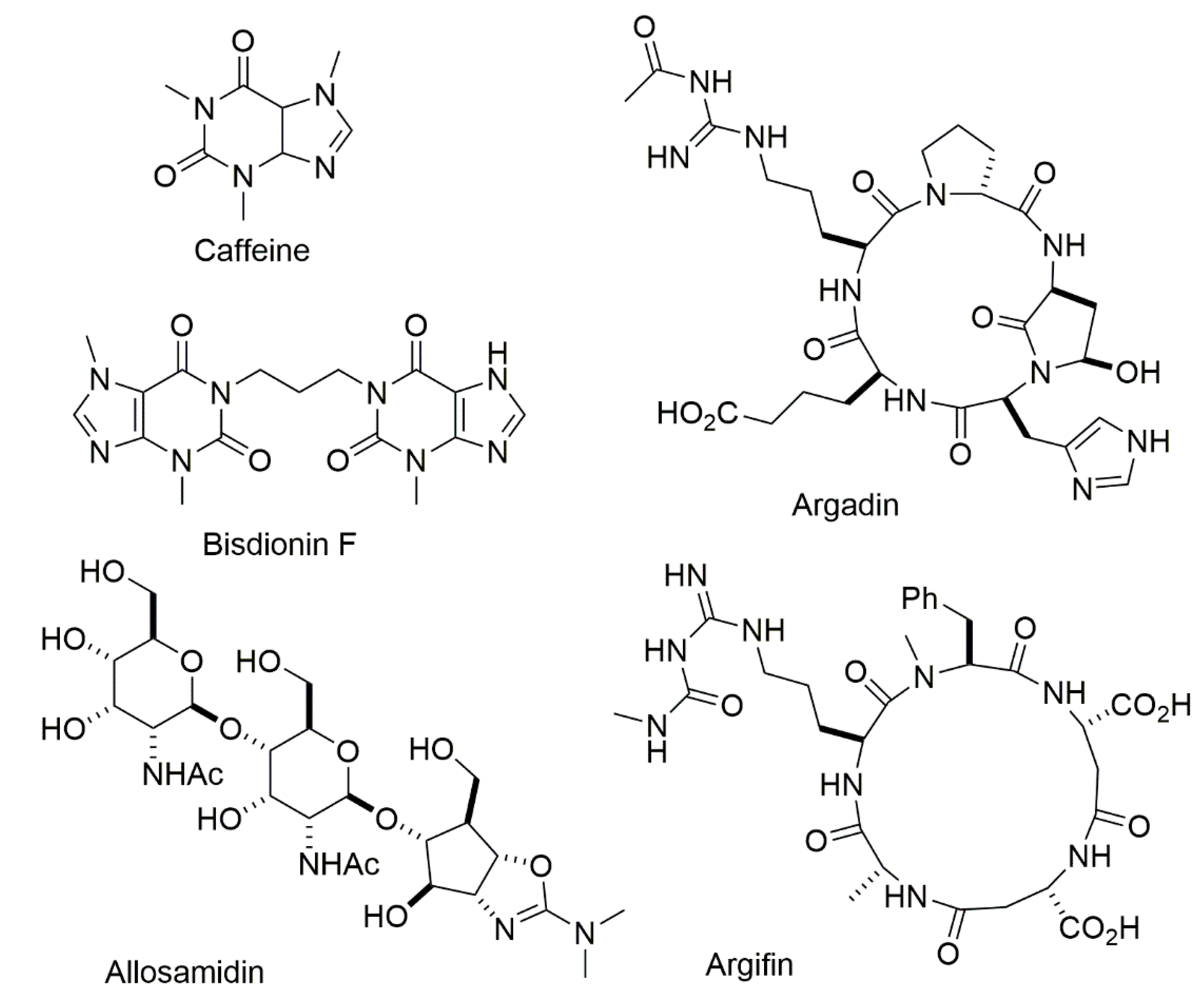
- Huang, G.; Peng, D.; Mei, X.; Chen, X.; Xiao, F.; Tang, Q. High-efficient synthesis and biological activities of allosamidins. J. Enzym. Inhib. Med. Chem. 2015, 30, 863–866. [Google Scholar] [CrossRef]
- Hirose, T.; Sunazuka, T.; Ōmura, S. Recent development of two chitinase inhibitors, Argifin and Argadin, produced by soil microorganisms. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2010, 86, 85–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, F.V.; Andersen, O.A.; Vora, K.A.; De Martino, J.A.; van Aalten, D.M.F. Methylxanthine drugs are chitinase inhibitors: Investigation of inhibition and binding modes. Chem. Biol. 2005, 12, 973–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutherland, T.E.; Andersen, O.A.; Betou, M.; Eggleston, I.M.; Maizels, R.M.; Van Aalten, D.; Allen, J.E. Analyzing Airway Inflammation with Chemical Biology: Dissection of Acidic Mammalian Chitinase Function with a Selective Drug-like Inhibitor. Chem. Biol. 2011, 18, 569–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, D.C.; Olland, A.M.; Jacob, J.; Brooks, J.; Bursavich, M.G.; Czerwinski, R.; Declercq, C.; Johnson, M.; Joseph-McCarthy, D.; Ellingboe, J.W.; et al. Identification and Characterization of Acidic Mammalian Chitinase Inhibitors. J. Med. Chem. 2010, 53, 6122–6128. [Google Scholar] [CrossRef] [PubMed]
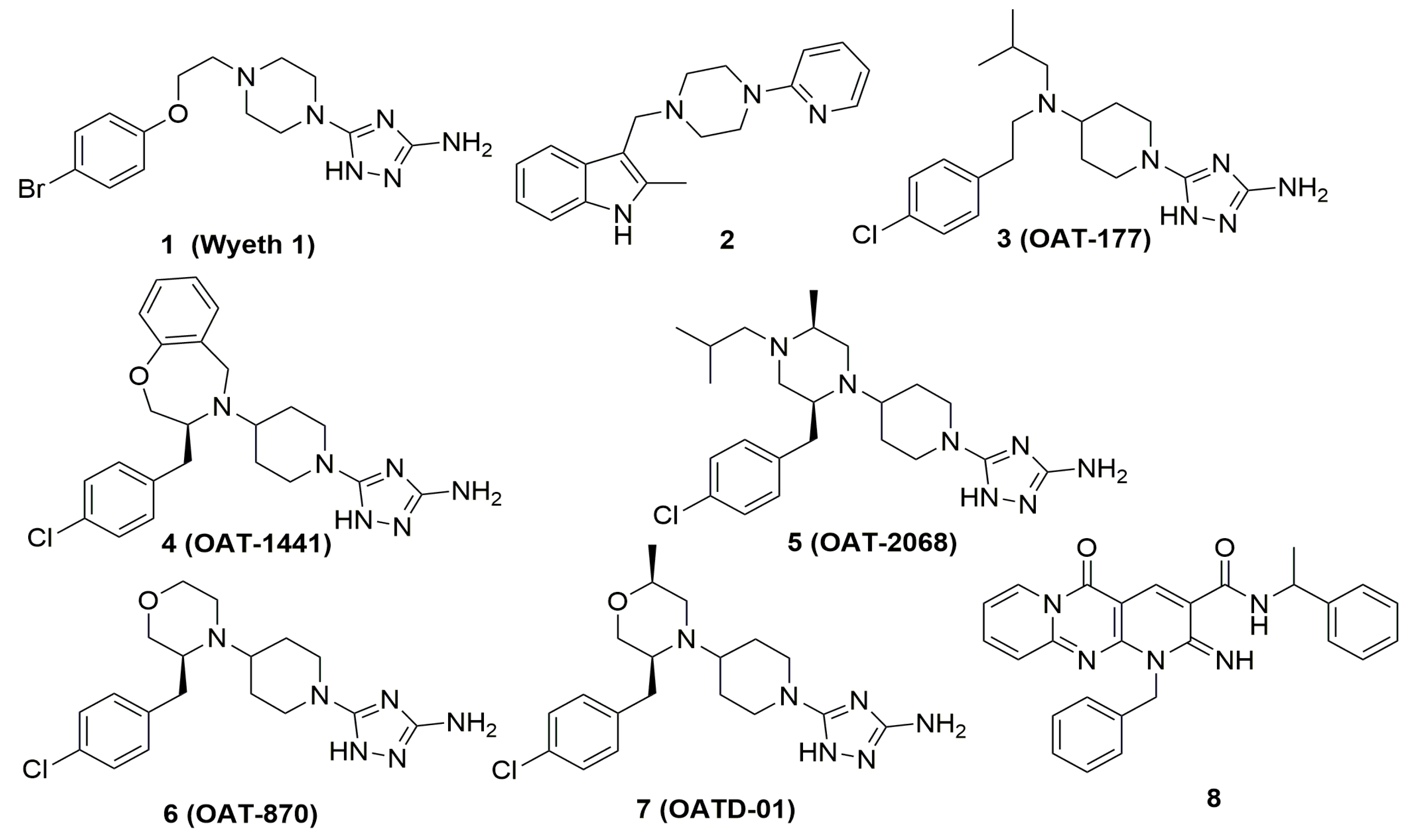
- Mazur, M.; Olczak, J.; Olejniczak, S.; Koralewski, R.; Czestkowski, W.; Jedrzejczak, A.; Golab, J.; Dzwonek, K.; Dymek, B.; Sklepkiewicz, P.L.; et al. Targeting Acidic Mammalian chitinase Is Effective in Animal Model of Asthma. J. Med. Chem. 2018, 61, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Andryianau, G.; Kowalski, M.; Piotrowicz, M.C.; Rajkiewicz, A.A.; Dymek, B.; Sklepkiewicz, P.L.; Pluta, E.; Stefaniak, F.; Czestkowski, W.; Olejniczak, S.; et al. Benzoxazepine-Derived Selective, Orally Bioavailable Inhibitor of Human Acidic Mammalian Chitinase. ACS Med. Chem. Lett. 2020, 11, 1228–1235. [Google Scholar] [CrossRef] [PubMed]
- Mazur, M.; Bartoszewicz, A.; Dymek, B.; Salamon, M.; Andryianau, G.; Kowalski, M.; Olejniczak, S.; Matyszewski, K.; Pluta, E.; Borek, B.; et al. Discovery of selective, orally bioavailable inhibitor of mouse chitotriosidase. Bioorg. Med. Chem. Lett. 2018, 28, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Mazur, M.; Dymek, B.; Koralewski, R.; Sklepkiewicz, P.; Olejniczak, S.; Mazurkiewicz, M.; Piotrowicz, M.; Salamon, M.; Jędrzejczak, K.; Zagozdzon, A.; et al. Development of Dual Chitinase Inhibitors as Potential New Treatment for Respiratory System Diseases. J. Med. Chem. 2019, 62, 7126–7145. [Google Scholar] [CrossRef]
- Koralewski, R.; Dymek, B.; Mazur, M.; Sklepkiewicz, P.; Olejniczak, S.; Czestkowski, W.; Matyszewski, K.; Andryianau, G.; Niedziejko, P.; Kowalski, M.; et al. Discovery of OATD-01, a First-in-Class Chitinase Inhibitor as Potential New Therapeutics for Idiopathic Pulmonary Fibrosis. J. Med. Chem. 2020, 63. [Google Scholar] [CrossRef]
- Jiang, X.; Kumar, A.; Motomura, Y.; Liu, T.; Zhou, Y.; Moro, K.; Zhang, K.Y.J.; Yang, Q. A Series of Compounds Bearing a Dipyrido-Pyrimidine Scaffold Acting as Novel Human and Insect Pest Chitinase Inhibitors. J. Med. Chem. 2020, 63, 987–1001. [Google Scholar] [CrossRef]
- Johansen, J.S.; Christensen, I.J.; Jorgensen, L.N.; Olsen, J.; Rahr, H.B.; Nielsen, K.T.; Laurberg, S.; Brünner, N.; Nielsen, H.J. Serum YKL-40 in Risk Assessment for Colorectal Cancer: A Prospective Study of 4,496 Subjects at Risk of Colorectal Cancer. Cancer Epidemiol. Biomark. Prev. 2015, 24, 621–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-C.; Llado, V.; Eurich, K.; Tran, H.T.; Mizoguchi, E. Carbohydrate-binding motif in chitinase 3-like 1 (CHI3L1/YKL-40) specifically activates Akt signaling pathway in colonic epithelial cells. Clin. Immunol. 2011, 140, 268–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, H.; Su, Q.; Lin, Z.; Zhu, X.; Peng, M.; Lv, Z. Potential suppressive effects of theophylline on human rectal cancer SW480 cells in vitro by inhibiting YKL-40 expression. Oncol. Lett. 2018, 15, 7403–7408. [Google Scholar] [CrossRef] [PubMed]
- Koyama, R.; Kataoka, H.; Tanaka, Y.; Nakatsugi, S.; Furukawa, M. Effect of Caffeine on Ibuprofen-induced Gastric Mucosal Damage in Rats. J. Pharm. Pharmacol. 1999, 51, 817–824. [Google Scholar] [CrossRef]
- Peterson, T.C.; Peterson, M.R.; Raoul, J.M. The effect of pentoxifylline and its metabolite-1 on inflammation and fibrosis in the TNBS model of colitis. Eur. J. Pharmacol. 2011, 662, 47–54. [Google Scholar] [CrossRef]
- Peterson, T.C.; Davey, K. Effect of acute pentoxifylline treatment in an experimental model of colitis. Aliment. Pharmacol. Ther. 1997, 11, 575–580. [Google Scholar] [CrossRef] [Green Version]
- Murthy, S.; Cooper, H.S.; Yoshitake, H.; Meyer, J.; Meyer, C.J.; Murthy, M.S. Combination therapy of pentoxifylline and TNFalpha monoclonal antibody in dextran sulphate-induced mouse colitis. Aliment. Pharmacol. Ther. 1999, 13, 251–260. [Google Scholar] [CrossRef]
- Reimund, J.M.; Dumont, S.; Muller, C.; Kenney, J.S.; Kedinger, M.; Baumann, R.; Poindron, P.; Duclos, B. In vitro effects of oxpentifylline on inflammatory cytokine release in patients with inflammatory bowel disease. Gut 1997, 40, 475–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzeminski, L.; Czestkowski, W.; Mazur, M.; Bartoszewicz, A.; Olejniczak, S.; Siwinska, A.; Krysztofiak, K.; Belczyk-Ciesielska, A.; Koziel, R.; Papiernik, D.; et al. Discovery of small molecule compounds interfering with YKL-40 carbohydrate binding as potential therapeutics for cancer. In Proceedings of the Abstracts of Papers of the American Chemical Society, San Diego, CA, USA, 25–29 August 2019; Volume 258. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazur, M.; Zielińska, A.; Grzybowski, M.M.; Olczak, J.; Fichna, J. Chitinases and Chitinase-Like Proteins as Therapeutic Targets in Inflammatory Diseases, with a Special Focus on Inflammatory Bowel Diseases. Int. J. Mol. Sci. 2021, 22, 6966. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136966
Mazur M, Zielińska A, Grzybowski MM, Olczak J, Fichna J. Chitinases and Chitinase-Like Proteins as Therapeutic Targets in Inflammatory Diseases, with a Special Focus on Inflammatory Bowel Diseases. International Journal of Molecular Sciences. 2021; 22(13):6966. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136966
Chicago/Turabian StyleMazur, Marzena, Anna Zielińska, Marcin M. Grzybowski, Jacek Olczak, and Jakub Fichna. 2021. "Chitinases and Chitinase-Like Proteins as Therapeutic Targets in Inflammatory Diseases, with a Special Focus on Inflammatory Bowel Diseases" International Journal of Molecular Sciences 22, no. 13: 6966. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136966