
Transamidation Down-Regulates Intestinal Immunity of Recombinant α-Gliadin in HLA-DQ8 Transgenic Mice

 ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
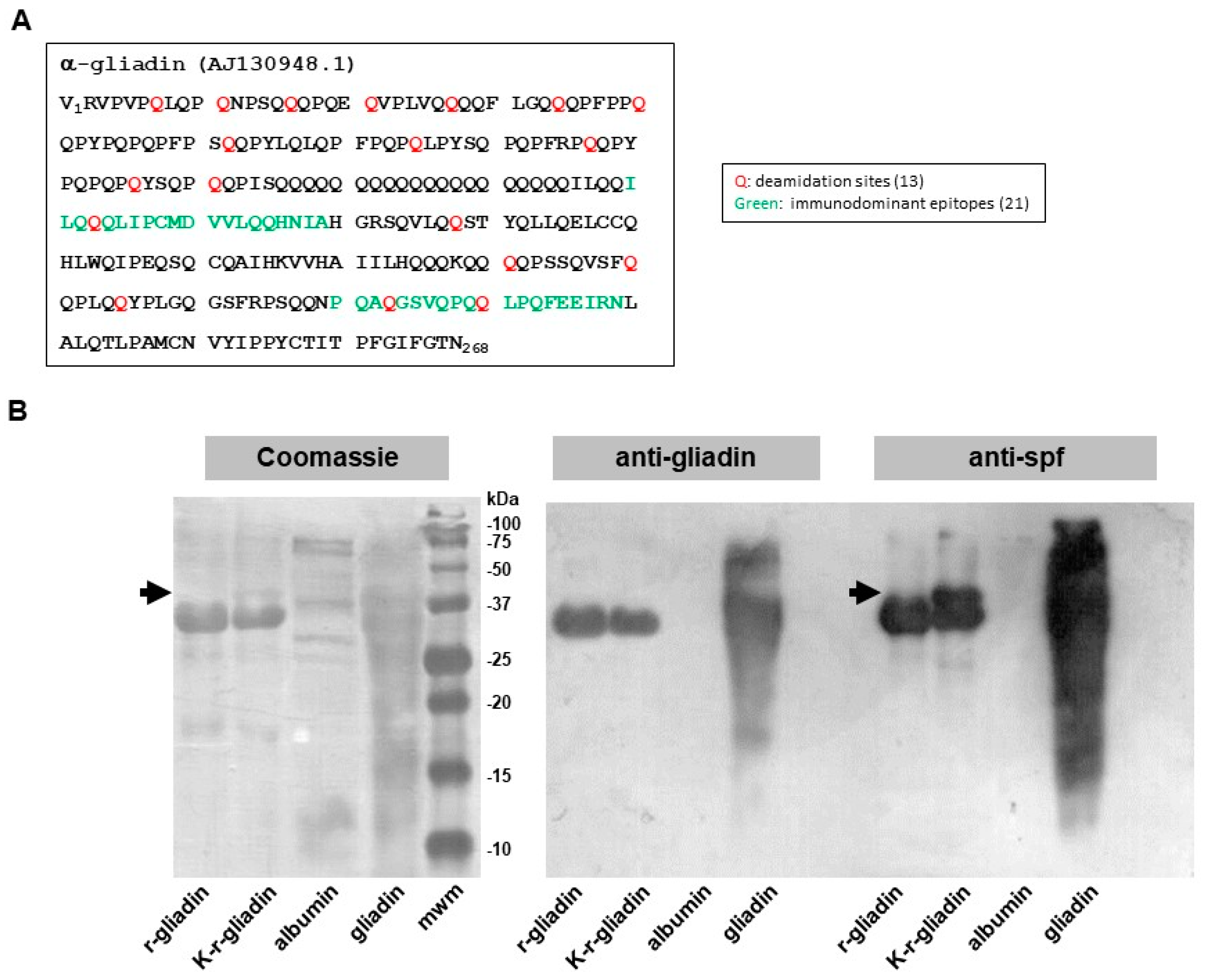
2.1. Biochemical Assessment of Transamidated Recombinant Alpha-Gliadin
2.2. Analysis of Transamidated Gliadin in DQ8 tg Mice
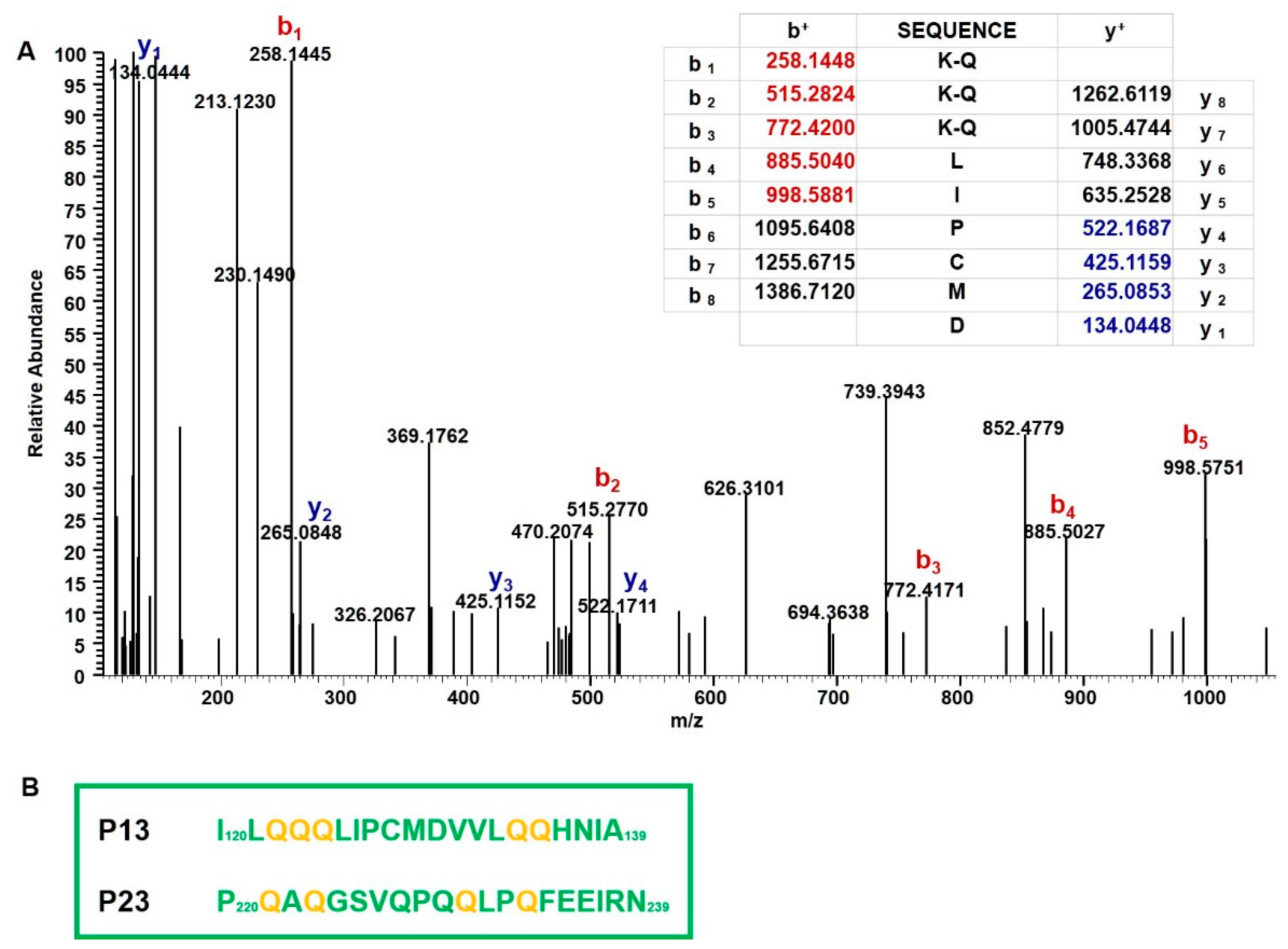
2.3. Mass Spectrometry Analysis of Immunodominant Peptides of r-Gliadin
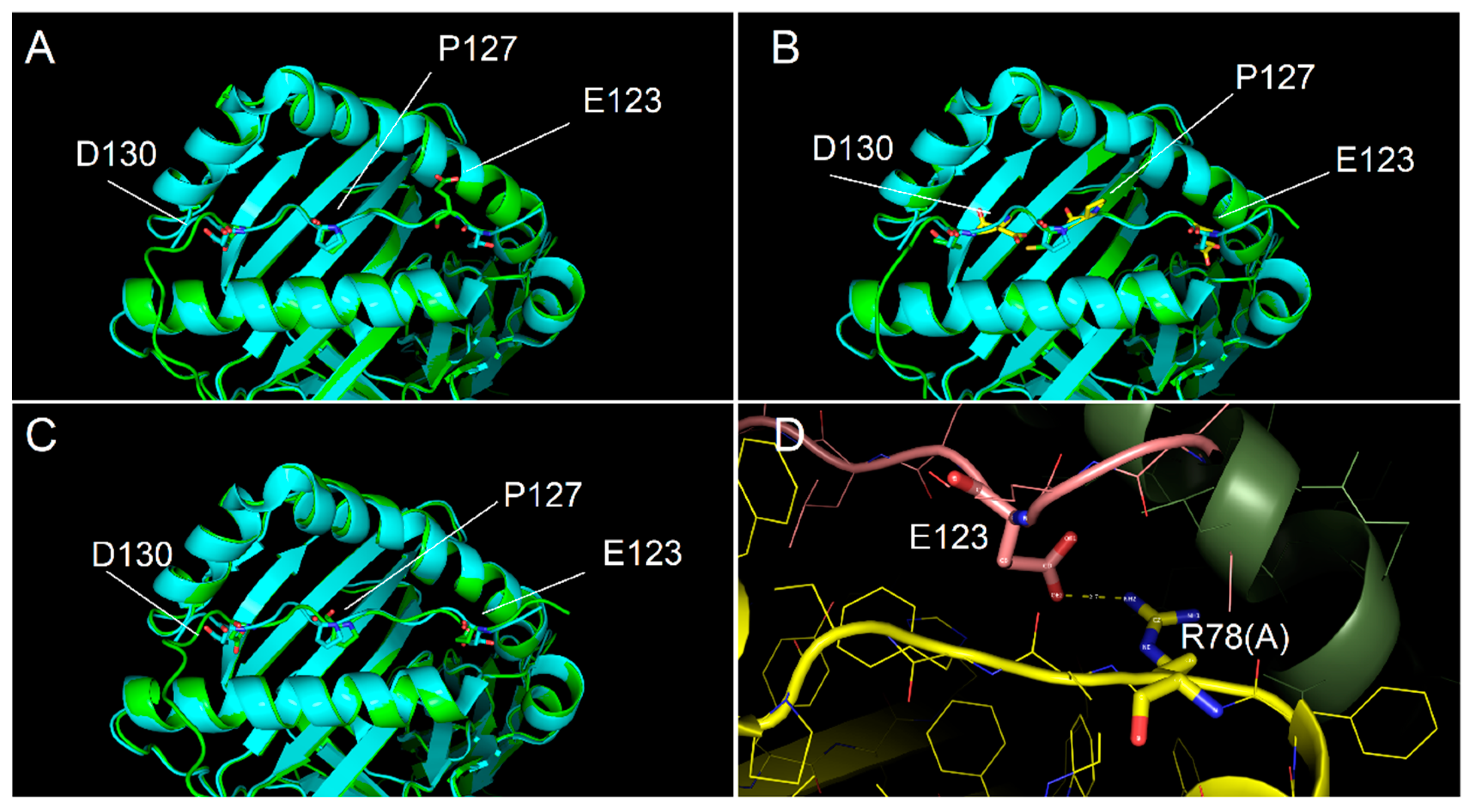
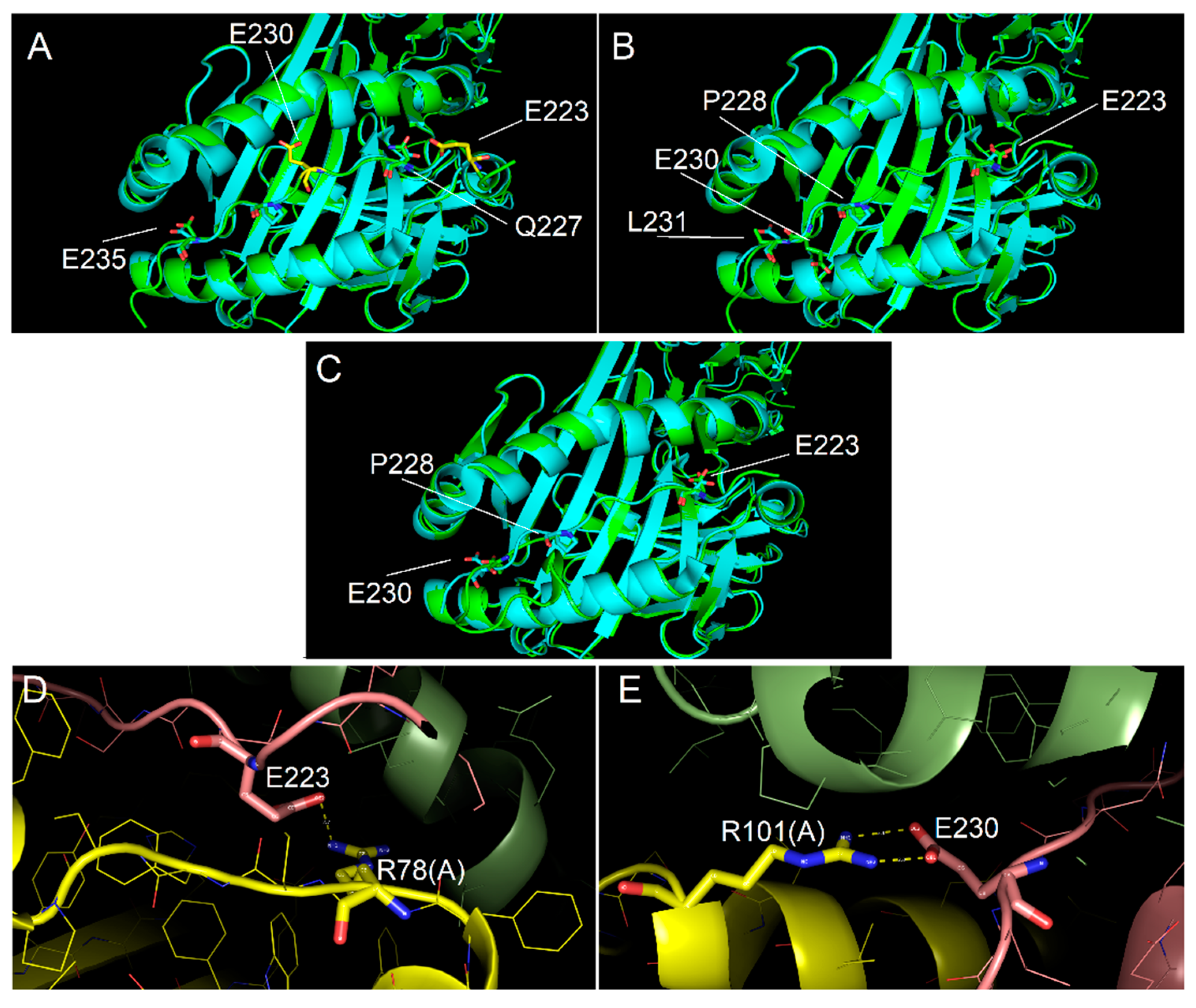
2.4. Ability of Immunodominant Peptides to Interact with HLA-DQ8 Heterodimer
3. Discussion
4. Materials and Methods
4.1. Antigens
4.2. Mice and Treatments
4.3. Western Blotting
4.4. In Vitro Culture of Mesenteric Lymph Node Cells
4.5. Cytokine Analysis
4.6. Identification of Transamidated Residues in K-r-Gliadin by Tandem Mass Spectrometry
4.7. Molecular Modelling Procedures
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jabri, B.; Chen, X.; Sollid, L.M. How T cells taste gluten in celiac disease. Nat. Struct. Mol. Biol. 2014, 21, 429–431. [Google Scholar] [CrossRef] [PubMed]
- Sollid, L.M. Coeliac disease: Dissecting a complex inflammatory disorder. Nat. Rev. Immunol. 2002, 2, 647–655. [Google Scholar] [CrossRef]
- Molberg, O.; McAdam, S.N.; Korner, R.; Quarsten, H.; Kristiansen, C.; Madsen, L.; Fugger, L.; Scott, H.; Noren, O.; Roepstorff, P.; et al. Tissue transglutaminase selectively modifies gliadin peptides that are recognized by gut-derived T cells in celiac disease. Nat. Med. 1988, 4, 713–717. [Google Scholar] [CrossRef]
- Arentz-Hansen, H.; Corner, R.; Molberg, O.; Quarsten, H.; Vader, W.; Kooy, Y.M.; Lundin, K.E.; Koning, F.; Roepstorff, P.; Sollid, L.M.; et al. The intestinal T cell response to alpha-gliadin in adult celiac disease is focused on a single deamidated glutamine targeted by tissue transglutaminase. J. Exp. Med. 2000, 91, 603–612. [Google Scholar] [CrossRef]
- Sjostrom, H.; Lundin, K.E.; Molberg, O.; Corner, R.; McAdam, S.N.; Anthonsen, D.; Quarsten, H.; Noren, O.; Roepstorff, P.; Thorsby, E.; et al. Identification of a gliadin T-cell epitope in coeliac disease: General importance of gliadin deamidation for intestinal T-cell recognition. Scand. J. Immunol. 1998, 48, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Vader, W.; Kooy, Y.; Van Veelen, P.; De Ru, A.; Harris, D.; Benckhuijsen, W.; Pena, S.; Mearin, L.; Drijfhout, J.W.; Koning, F. The gluten response in children with celiac disease is directed toward multiple gliadin and glutenin peptides. Gastroenterology 2002, 122, 1729–1737. [Google Scholar] [CrossRef] [Green Version]
- Van de Wal, Y.; Kooy, Y.; van Veelen, P.; Pena, S.; Mearin, L.; Papadopoulos, G.; Koning, F. Selective deamidation by tissue transglutaminase strongly enhances gliadin-specific T cell reactivity. J. Immunol. 1998, 161, 1585–1588. [Google Scholar] [PubMed]
- Shan, L.; Molberg, O.; Parrot, I.; Hausch, F.; Filiz, F.; Gray, G.M.; Sollid, L.M.; Khosla, C. Structural basis for gluten intolerance in celiac sprue. Science 2002, 297, 2275–2279. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Siegel, M.; Bergseng, E.; Sollid, L.M.; Khosla, C. Inhibition of HLA-DQ2 Mediated Antigen Presentation by Analogues of a High Affinity 33-Residue Peptide from α2-Gliadin. J. Am. Chem. Soc. 2006, 128, 1859–1867. [Google Scholar] [CrossRef] [Green Version]
- Gianfrani, C.; Siciliano, R.A.; Facchiano, A.M.; Camarca, A.; Mazzeo, M.F.; Costantini, S.; Salvati, V.M.; Maurano, F.; Mazzarella, G.; Iaquinto, G.; et al. Transamidation inhibits the intestinal immune response to gliadin in vitro. Gastroenterology 2007, 133, 780–789. [Google Scholar] [CrossRef]
- Kanaji, T.; Ozaki, H.; Takao, T.; Kawajiri, H.; Ide, H.; Motoki, M.; Shimonishi, Y. Primary structure of microbial transglutaminase from Streptoverticillium sp. strain s-8112. J. Biol. Chem. 1993, 668, 11565–11572. [Google Scholar] [CrossRef]
- Yokoyama, K.; Nio, N.; Kikuchi, Y. Properties and applications of microbial transglutaminase. Appl. Microbiol. Biotechnol. 2004, 64, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Mazzeo, M.F.; De Giulio, B.; Senger, S.; Rossi, M.; Malorni, A.; Siciliano, R.A. Identification of transglutaminase-mediated deamidation sites in a recombinant alpha-gliadin by advanced mass-spectrometric methodologies. Protein Sci. 2003, 12, 2434–2442. [Google Scholar] [CrossRef] [PubMed]
- Mazzeo, M.F.; Bonavita, R.; Maurano, F.; Bergamo, P.; Siciliano, R.A.; Rossi, M. Biochemical modifications of gliadins induced by microbial transglutaminase on wheat flour. Biochim. Biophys. Acta. 2013, 1830, 5166–5174. [Google Scholar] [CrossRef]
- Lombardi, E.; Maurano, F.; Bergamo, P.; Bozzella, G.; Luongo, D.; Mazzarella, G.; Capobianco, F.; Rotondi Aufiero, V.; Iaquinto, G.; Rossi, M. Selective inhibition of gliadin immune reactivity by transamidation of wheat flour with microbial transglutaminasel. J. Leukoc. Biol. 2013, 93, 479–488. [Google Scholar] [CrossRef]
- Luongo, D.; Bonavita, R.; Rossi, S.; Aufiero, V.R.; Feliciello, N.R.; Maurano, F.; Iaquinto, G.; Mazzarella, G.; Rossi, M. Tailoring the immune response to wheat gliadin by enzymatic transamidation. Cytokine 2019, 117, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Senger, S.; Maurano, F.; Mazzeo, M.F.; Gaita, M.; Fierro, O.; David, C.S.; Troncone, R.; Auricchio, S.; Siciliano, R.A.; Rossi, M. Identification of immunodominant epitopes of alpha-gliadin in HLA-DQ8 transgenic mice following oral immunization. J. Immunol. 2005, 175, 8087–8095. [Google Scholar] [CrossRef]
- Zilić, S.; Barać, M.; Pešić, M.; Dodig, D.; Ignjatović-Micić, D. Characterization of proteins from grain of different bread and durum wheat genotypes. Int. J. Mol. Sci. 2011, 12, 5878–5894. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Black, K.E.; Murray, J.A.; David, C.S. HLA-DQ determines the response to exogenous wheat proteins: A model of gluten sensitivity in transgenic knockout mice. J. Immunol. 2002, 169, 5595–5600. [Google Scholar] [CrossRef] [Green Version]
- Senger, S.; Luongo, D.; Maurano, F.; Mazzeo, M.F.; Siciliano, R.A.; Gianfrani, C.; David, C.; Troncone, R.; Auricchio, S.; Rossi, M. Intranasal administration of a recombinant alpha-gliadin down-regulates the immune response to wheat gliadin in DQ8 transgenic mice. Immunol. Lett. 2003, 88, 127–134. [Google Scholar] [CrossRef]
- La Sala, A.; He, J.; Laricchia-Robbio, L.; Gorini, S.; Iwasaki, A.; Braun, M.; Yap, G.S.; Sher, A.; Ozato, K.; Kelsall, B. Cholera toxin inhibits IL-12 production and CD8alpha+ dendritic cell differentiation by cAMP-mediated inhibition of IRF8 function. J. Exp. Med. 2009, 206, 1227–1235. [Google Scholar] [CrossRef]
- Dekking, E.H.A.; Van Veelen, P.A.; de Ru, A.; Kooy-Winkelaar, E.M.C.; Gröneveld, T.; Nieuwenhuizen, W.F.; Koning, F. Microbial transglutaminases generate T cell stimulatory epitopes involved in celiac disease. J. Cereal Sci. 2008, 47, 339–346. [Google Scholar] [CrossRef]
- Costantini, S.; Colonna, G.; Rossi, M.; Facchiano, A.M. Modelling of HLA-DQ2 and simulations of its interaction with gluten peptides to explain molecular recognition in celiac disease. J. Mol. Graph. Model. 2005, 23, 419–431. [Google Scholar] [CrossRef]
- Nakae, S.; Iwakura, Y.; Suto, H.; Galli, S.J. Phenotypic differences between Th1 and Th17 cells and negative regulation of Th1 cell differentiation by IL-17. J. Leukoc. Biol. 2007, 81, 1258–1268. [Google Scholar] [CrossRef] [PubMed]
- Gagliani, N.; Amezcua Vesely, M.C.; Iseppon, A.; Brockmann, L.; Xu, H.; Palm, N.W.; de Zoete, M.R.; Licona-Limón, P.; Paiva, R.S.; Ching, T.; et al. Th17 cells transdifferentiate into regulatory T cells during resolution of inflammation. Nature 2015, 523, 221–225. [Google Scholar] [CrossRef]
- Weegels, P.L.; Hamer, R.J.; Schofield, J.D. RP-HPLC and capillary electrophoresis of subunits from glutenin isolated by SDS and Osborne fractionation. J. Cereal Sci. 1995, 22, 211–224. [Google Scholar] [CrossRef]
- Rossi, S.; Luongo, D.; Maurano, F.; Bergamo, P.; Rossi, M. Immunomodulatory activity of recombinant α-gliadin conjugated to cholera toxin in DQ8 transgenic mice. Immunol. Lett. 2017, 187, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cheng, S.; Baisch, J.; Krco, C.; Savarirayan, S.; Hanson, J.; Hodgson, K.; Smart, M.; David, C. Expression and function of HLA-DQ8 (DQA1*0301/DQB1*0302) genes in transgenic mice. Eur. J. Immunogenet. 1996, 23, 15–20. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sollid, L.M. Molecular Basis of Celiac Disease. Annu. Rev. Immunol. 2000, 18, 53–81. [Google Scholar] [CrossRef] [Green Version]
- Sali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef]
- Buonocore, F.; Castro, R.; Randelli, E.; Lefranc, M.P.; Six, A.; Kuhl, H.; Reinhardt, R.; Facchiano, A.; Boudinot, P.; Scapigliati, G. Diversity, Molecular Characterization and Expression of T Cell Receptor γ in a Teleost Fish, the Sea Bass (Dicentrarchus labrax, L). PLoS ONE 2012, 7, e47957. [Google Scholar] [CrossRef] [PubMed]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, S.; Giordano, D.; Mazzeo, M.F.; Maurano, F.; Luongo, D.; Facchiano, A.; Siciliano, R.A.; Rossi, M. Transamidation Down-Regulates Intestinal Immunity of Recombinant α-Gliadin in HLA-DQ8 Transgenic Mice. Int. J. Mol. Sci. 2021, 22, 7019. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137019
Rossi S, Giordano D, Mazzeo MF, Maurano F, Luongo D, Facchiano A, Siciliano RA, Rossi M. Transamidation Down-Regulates Intestinal Immunity of Recombinant α-Gliadin in HLA-DQ8 Transgenic Mice. International Journal of Molecular Sciences. 2021; 22(13):7019. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137019
Chicago/Turabian StyleRossi, Stefano, Deborah Giordano, Maria Fiorella Mazzeo, Francesco Maurano, Diomira Luongo, Angelo Facchiano, Rosa Anna Siciliano, and Mauro Rossi. 2021. "Transamidation Down-Regulates Intestinal Immunity of Recombinant α-Gliadin in HLA-DQ8 Transgenic Mice" International Journal of Molecular Sciences 22, no. 13: 7019. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137019