
Comparative Mitogenomic Analysis Reveals Gene and Intron Dynamics in Rubiaceae and Intra-Specific Diversification in Damnacanthus indicus


Abstract
:1. Introduction
2. Results
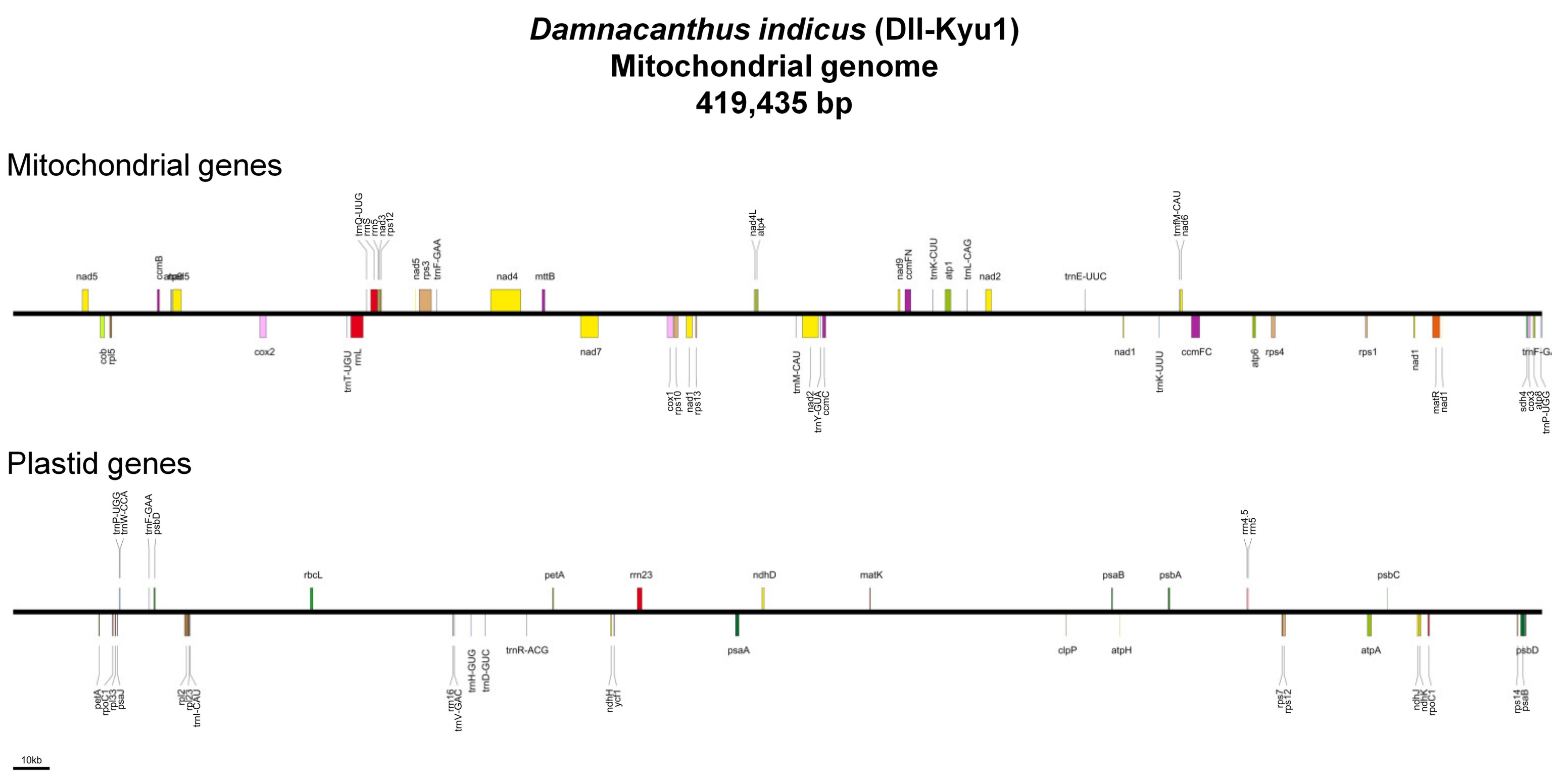
2.1. Mitogenome Assemblies and Features
2.2. Gene Content Comparison with Representative Angiosperms
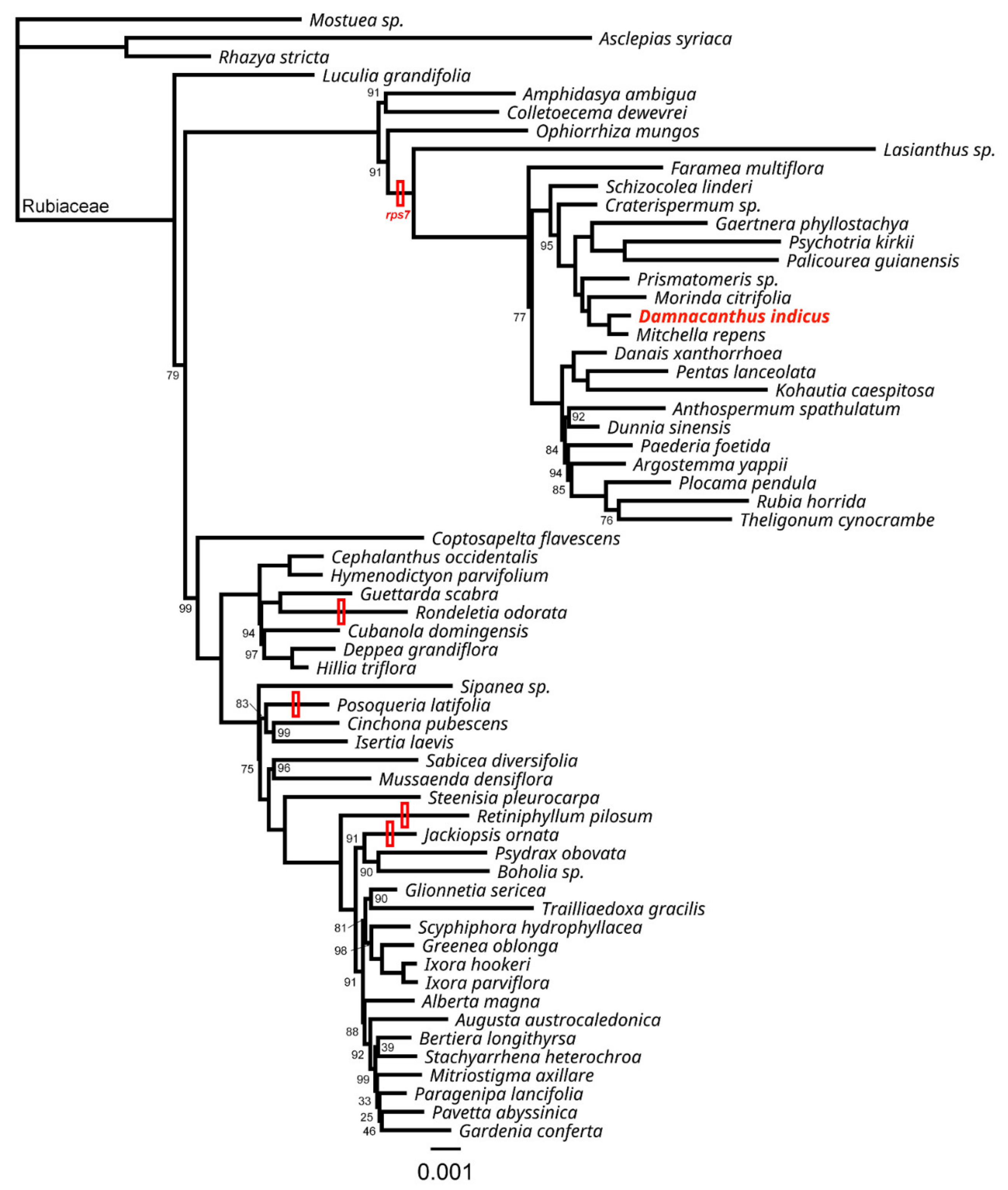
2.3. Losses of rps7 in Rubiaceae
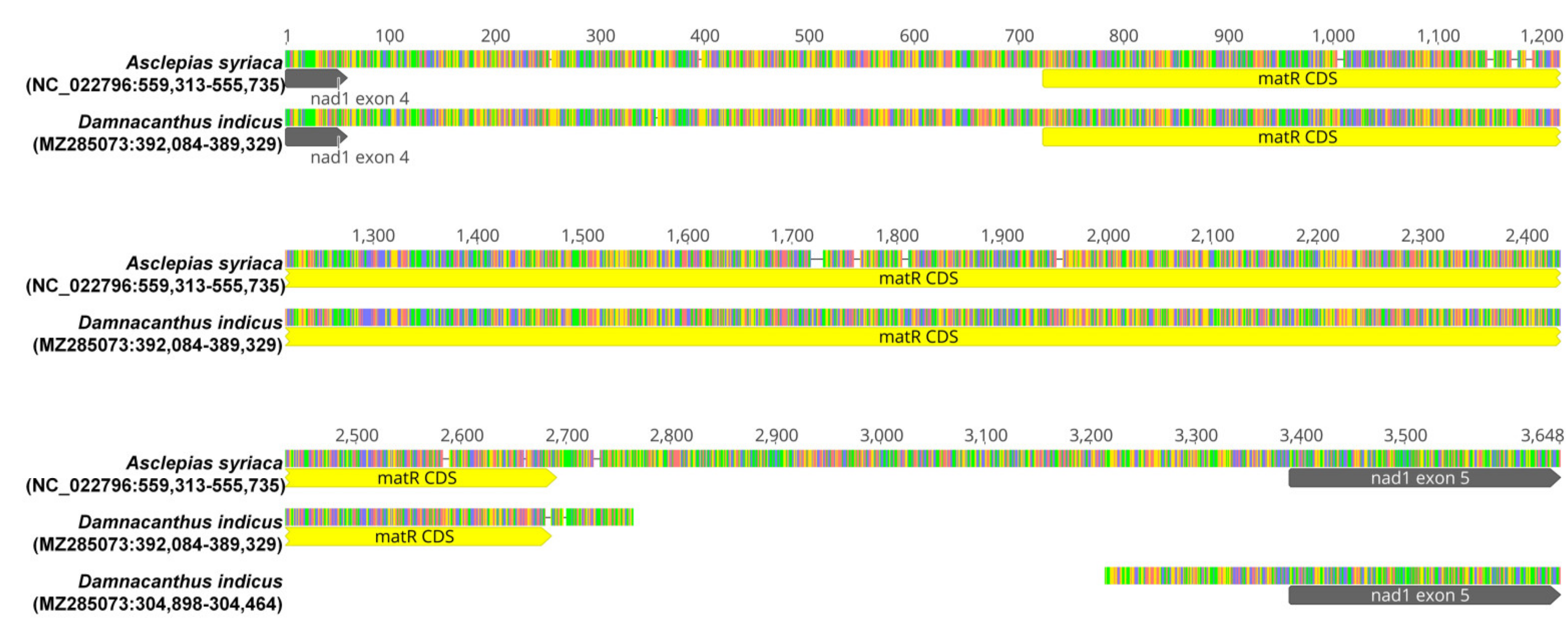
2.4. Trans-Splicing of nad1i728 in Rubiaceae
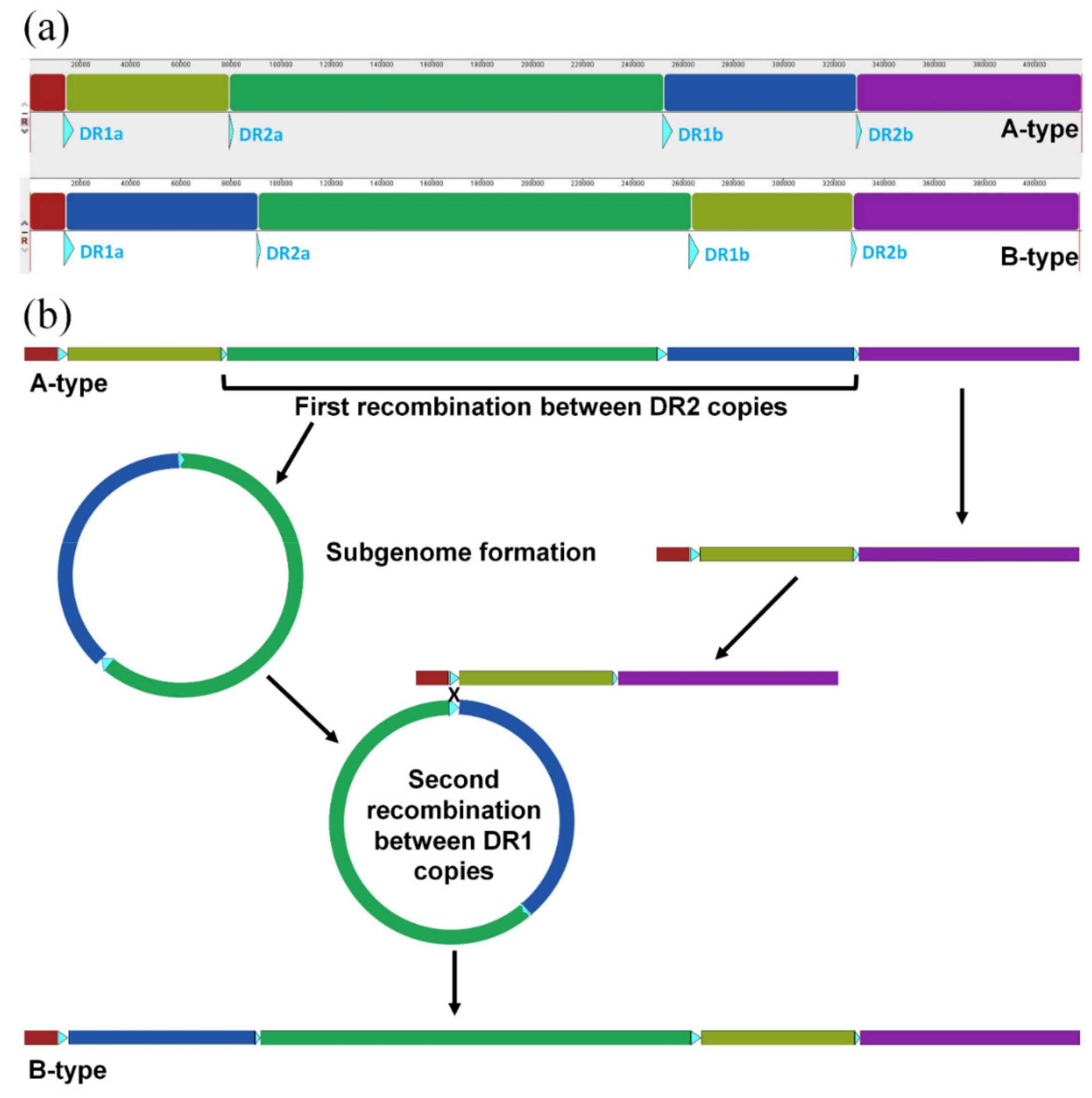
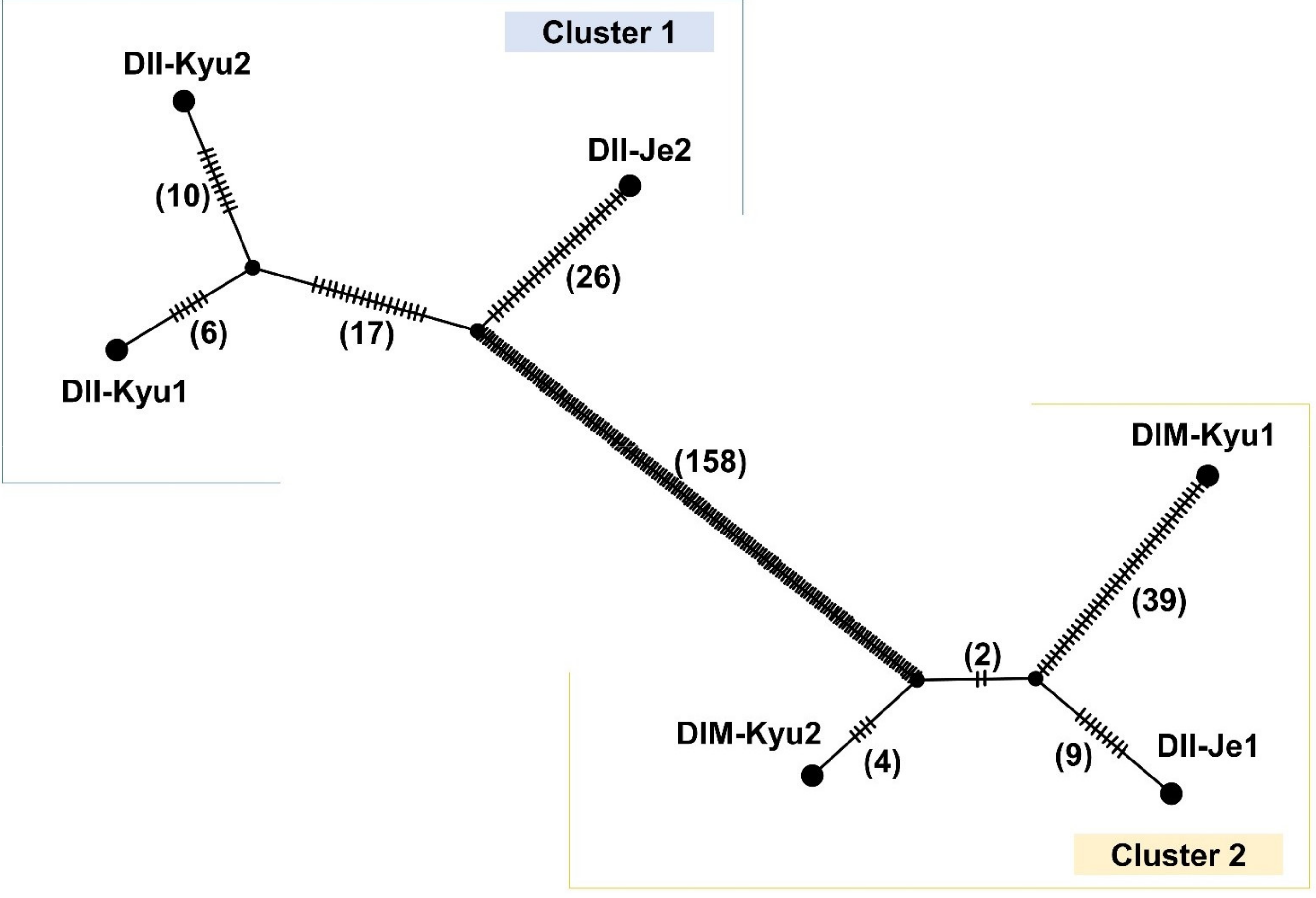
2.5. Intra-Specific Mitogenome Rearrangement and Divergence of Damnacanthus Indicus
3. Discussion
4. Materials and Methods
4.1. Sampling and Sequencing
4.2. Mitogenome Assembly and Annotation
4.3. Gene and Intron Content Comparison with Representative Angiosperms
4.4. Phylogenetic Analysis and rps7 Investigation in Rubiaceae
4.5. Investigation on the Configuration of nad1i728 in Rubiaceae
4.6. Mitogenome Sequence and Structural Divergence in Damnacanthus Indicus
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oh, S.-H. A taxonomic note of Ageratina altissima and Eupatorium rugosum (Asteraceae: Eupatorieae). Korean J. Plant Taxon. 2019, 49, 261–264. [Google Scholar] [CrossRef] [Green Version]
- Jin, D.-P.; Park, J.-W.; Choi, B.-H. Taxonomic study on infraspecific taxa of Lespedeza maximowiczii and hybrids with related species. Korean J. Plant Taxon. 2019, 49, 300–318. [Google Scholar] [CrossRef]
- Khan, N.A.; Singhal, V.K.; Tantray, Y.R.; Gupta, R.C. Report of intraspecific polyploidy (2x, 6x) in Rorippa palustris, Brassicaceae from cold deserts of Ladakh division (J & K), India. Cytologia 2019, 84, 207–210. [Google Scholar]
- Hoang, P.T.N.; Schubert, V.; Meister, A.; Fuchs, J.; Schubert, I. Variation in genome size, cell and nucleus volume, chromosome number and rDNA loci among duckweeds. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Lee, J.-H.; Lee, D.-H.; Choi, B.-H. Phylogeography and genetic diversity of East Asian Neolitsea sericea (Lauraceae) based on variations in chloroplast DNA sequences. J. Plant Res. 2013, 126, 193–202. [Google Scholar] [CrossRef]
- Tamaki, I.; Kawashima, N.; Setsuko, S.; Lee, J.-H.; Itaya, A.; Yukitoshi, K.; Tomaru, N. Population genetic structure and de-mography of Magnolia kobus: Variety borealis is not supported genetically. J. Plant Res. 2019, 132, 741–758. [Google Scholar] [CrossRef]
- Yamazaki, T. Damnacanthus. In Flora of Japan; Iwatsuki, K.Y., T. Boufford, D.E., Ohba, H., Eds.; Kodansha: Tokyo, Japan, 1993; Volume 3, pp. 224–225. [Google Scholar]
- Lee, T.B. Colored Flora of Korea (in Korean); Hyangmonsa Pulishing Co.: Seoul, Korea, 2003; p. 213. [Google Scholar]
- Chen, T.; Taylor, C.M. Damnacanthus. In Flora of China; Wu, C.Y., Raven, P.H., Eds.; Science Press: Beijing, China, 2011; Volume 19, pp. 93–97. [Google Scholar]
- Robbrecht, E.; Puff, C.; Igersheim, A. The genera Mitchella and Damnacanthus, evidence for their close alliance; comments on the campylotropy in the Rubiaceae and the circumscription of the Morindeae. Blumea 1991, 35, 307–345. [Google Scholar]
- Naiki, A.; Nagamasu, H. Distyly and pollen dimorphism in Damnacanthus (Rubiaceae). J. Plant Res. 2003, 116, 105–113. [Google Scholar] [CrossRef]
- Naiki, A.; Nagamasu, H. Correlation between distyly and ploidy level in Damnacanthus (Rubiaceae). Am. J. Bot. 2004, 91, 664–671. [Google Scholar] [CrossRef]
- Huang, W.-P.; Sun, H.; Deng, T.; Razafimandimbison, S.G.; Nie, Z.-L.; Wen, J. Molecular phylogenetics and biogeography of the eastern Asian–eastern North American disjunct Mitchella and its close relative Damnacanthus (Rubiaceae, Mitchelleae). Bot. J. Linnean Soc. 2013, 171, 395–412. [Google Scholar] [CrossRef] [Green Version]
- Knoop, V. The mitochondrial DNA of land plants: Peculiarities in phylogenetic perspective. Curr. Genet. 2004, 46, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Knoop, V. Seed Plant Mitochondrial Genomes: Complexity Evolving. In Genomics of Chloroplasts and Mitochondria; Springer: Dordrecht, The Netherlands, 2012; pp. 175–200. [Google Scholar]
- Putintseva, Y.A.; Bondar, E.I.; Simonov, E.P.; Sharov, V.V.; Oreshkova, N.V.; Kuzmin, D.A.; Konstantinov, Y.M.; Shmakov, V.N.; Belkov, V.I.; Sadovsky, M.G.; et al. Siberian larch (Larix sibirica Ledeb.) mitochondrial genome assembled using both short and long nucleotide sequence reads is currently the largest known mitogenome. BMC Genom. 2020, 21, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.L.; Qiu, Y.-L.; Stoutemyer, M.; Palmer, J.D. Punctuated evolution of mitochondrial gene content: High and variable rates of mitochondrial gene loss and transfer to the nucleus during angiosperm evolution. Proc. Natl. Acad. Sci. USA 2002, 99, 9905–9912. [Google Scholar] [CrossRef] [Green Version]
- Richardson, O.A.; Rice, D.W.; Young, G.J.; Alverson, A.J.; Palmer, J.D. The “fossilized” mitochondrial genome of Liriodendron tulipifera: Ancestral gene content and order, ancestral editing sites, and extraordinarily low mutation rate. BMC Biol. 2013, 11, 29. [Google Scholar] [CrossRef] [Green Version]
- Mower, J.P. Variation in protein gene and intron content among land plant mitogenomes. Mitochondrion 2020, 53, 203–213. [Google Scholar] [CrossRef]
- Choi, I.-S.; Schwarz, E.N.; Ruhlman, T.A.; Khiyami, M.A.; Sabir, J.S.M.; Hajarah, N.H.; Sabir, M.J.; Rabah, S.O.; Jansen, R.K. Fluctuations in Fabaceae mitochondrial genome size and content are both ancient and recent. BMC Plant. Biol. 2019, 19, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rydin, C.; Wikström, N.; Bremer, B. Conflicting results from mitochondrial genomic data challenge current views of Rubiaceae phylogeny. Am. J. Bot. 2017, 104, 1522–1532. [Google Scholar] [CrossRef] [Green Version]
- Johnston, I.G. Tension and resolution: Dynamic, evolving populations of organelle genomes within plant cells. Mol. Plant 2019, 12, 764–783. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.H.; Li, W.-H.; Sharp, P. Rates of nucleotide substitution vary greatly among plant mitochondrial, chloroplast, and nuclear DNAs. Proc. Natl. Acad. Sci. USA 1987, 84, 9054–9058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drouin, G.; Daoud, H.; Xia, J. Relative rates of synonymous substitutions in the mitochondrial, chloroplast and nuclear genomes of seed plants. Mol. Phylogenet. Evol. 2008, 49, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.K.; Palmer, J.D. A chloroplast DNA inversion marks an ancient evolutionary split in the sunflower family (Asteraceae). Proc. Natl. Acad. Sci. USA 1987, 84, 5818–5822. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.J.; Doyle, J.L.; Ballenger, J.; Palmer, J. The distribution and phylogenetic significance of a 50-kb chloroplast DNA in-version in the flowering plant family Leguminosae. Mol. Phylogenet. Evol. 1996, 5, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.-S.; Choi, B.-H. The distinct plastid genome structure of Maackia fauriei (Fabaceae: Papilionoideae) and its systematic implications for genistoids and tribe Sophoreae. PLoS ONE 2017, 12, e0173766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, D.-P.; Choi, I.-S.; Choi, B.-H. Plastid genome evolution in tribe Desmodieae (Fabaceae: Papilionoideae). PLoS ONE 2019, 14, e0218743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, L.W.; Guo, W.; Mower, J.P.; Palmer, J.D. High and variable rates of repeat-mediated mitochondrial genome rearrange-ment in a genus of plants. Mol. Biol. Evol. 2018, 35, 2773–2785. [Google Scholar] [PubMed]
- Gualberto, J.M.; Mileshina, D.; Wallet, C.; Niazi, A.K.; Weber-Lotfi, F.; Dietrich, A. The plant mitochondrial genome: Dynamics and maintenance. Biochimie 2014, 100, 107–120. [Google Scholar] [CrossRef]
- Gualberto, J.M.; Newton, K.J. Plant mitochondrial genomes: Dynamics and mechanisms of mutation. Annu. Rev. Plant. Biol. 2017, 68, 225–252. [Google Scholar] [CrossRef]
- Woloszynska, M. Heteroplasmy and stoichiometric complexity of plant mitochondrial genomes—Though this be madness, yet there’s method in’t. J. Exp. Bot. 2010, 61, 657–671. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Zhu, A.; Fan, W.; Adams, R.P.; Mower, J.P. Extensive shifts from cis- to trans-splicing of gymnosperm mitochondrial introns. Mol. Biol. Evol. 2020, 37, 1615–1620. [Google Scholar] [CrossRef]
- Qiu, Y.-L.; Palmer, J.D. Many independent origins of trans splicing of a plant mitochondrial group II intron. J. Mol. Evol. 2004, 59, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Mower, J.P.; Jain, K.; Hepburn, N.J. The role of horizontal transfer in shaping the plant mitochondrial genome. Adv. Bot. Res. 2012, 63, 41–69. [Google Scholar] [CrossRef]
- Ellis, J. Promiscuous DNA—Chloroplast genes inside plant mitochondria. Nat. Cell Biol. 1982, 299, 678–679. [Google Scholar] [CrossRef]
- Gandini, C.; Sanchez-Puerta, M.V. Foreign plastid sequences in plant mitochondria are frequently acquired via mitochondrion-to-mitochondrion horizontal transfer. Sci. Rep. 2017, 7, 43402. [Google Scholar] [CrossRef]
- Warren, J.M.; Sloan, D.B. Interchangeable parts: The evolutionarily dynamic tRNA population in plant mitochondria. Mitochondrion 2020, 52, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Dugua, X.; Castellanos-Morales, G.; Paredes-Torres, L.M.; Hernández-Rosales, H.S.; Barrera-Redondo, J.; La Vega, G.S.-D.; Tapia-Aguirre, F.; Ruiz-Mondragón, K.Y.; Scheinvar, E.; Hernández, P.; et al. Evolutionary dynamics of transferred sequences between organellar genomes in Cucurbita. J. Mol. Evol. 2019, 87, 327–342. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.-L.; Li, L.; Wang, B.; Xue, J.-Y.; Hendry, T.A.; Li, R.-Q.; Brown, J.W.; Liu, Y.; Hudson, G.T.; Chen, Z.-D. Angiosperm phylogeny inferred from sequences of four mitochondrial genes. J. Syst. Evol. 2010, 48, 391–425. [Google Scholar] [CrossRef]
- Dong, S.; Chen, L.; Liu, Y.; Wang, Y.; Zhang, S.; Yang, L.; Lang, X. The draft mitochondrial genome of Magnolia biondii and mitochondrial phylogenomics of angiosperms. PLoS ONE 2020, 15, e0231020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, J.Y.; Dong, S.; Wang, M.Q.; Song, T.Q.; Zhou, G.C.; Li, Z.; Van de Peer, Y.; Shao, Z.Q.; Wang, W.; Chen, M. Mitochondrial genes from eighteen angiosperms fill sampling gaps for phylogenomic inferences of the early diversification of flowering plants. J. Syst. Evol. 2020. Early View. [Google Scholar] [CrossRef]
- Van De Paer, C.; Bouchez, O.; Besnard, G. Prospects on the evolutionary mitogenomics of plants: A case study on the olive family (Oleaceae). Mol. Ecol. Resour. 2018, 18, 407–423. [Google Scholar] [CrossRef]
- Duminil, J.; Besnard, G. Utility of the Mitochondrial Genome in Plant Taxonomic Studies. In Methods in Molecular Biology; Springer: New York, NY, USA, 2021; Volume 2222, pp. 107–118. [Google Scholar]
- Bogdanova, V.S.; Shatskaya, N.V.; Mglinets, A.V.; Kosterin, O.E.; Vasiliev, G.V. Discordant evolution of organellar genomes in peas (Pisum L.). Mol. Phylogenet. Evol. 2021, 160, 107136. [Google Scholar] [CrossRef]
- Burge, D. Conservation genomics and pollination biology of an endangered, edaphic-endemic, octoploid herb: El Dorado bedstraw (Galium californicum subsp. sierrae; Rubiaceae). PeerJ 2020, 8, e10042. [Google Scholar] [CrossRef]
- Rai, A.; Hirakawa, H.; Nakabayashi, R.; Kikuchi, S.; Hayashi, K.; Rai, M.; Tsugawa, H.; Nakaya, T.; Mori, T.; Nagasaki, H.; et al. Chromosome-level genome assembly of Ophiorrhiza pumila reveals the evolution of camptothecin biosynthesis. Nat. Commun. 2021, 12, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Bautista, M.A.C.; Tao, W.; Zheng, Y.; Deng, Y.; Chen, T.; Miao, S. Chloroplast genome organization and phylogeny of Gy-nchthodes cochinchinensis (DC.) Razafim. & B. Bremer (Rubiaceae). Mitochondrial DNA Part B 2021, 6, 261–262. [Google Scholar] [PubMed]
- Zhang, Y.; Chen, S.; Xu, X.; Wang, R. The complete chloroplast genome of Foonchewia coriacea (Rubioideae: Rubiaceae): A monotypic species endemic to Guangdong, China. Mitochondrial DNA Part. B 2021, 6, 156–157. [Google Scholar] [CrossRef] [PubMed]
- Brose, J.; Lau, K.H.; Dang, T.T.T.; Hamilton, J.P.; Martins, L.D.V.; Hamberger, B.; Hamberger, B.; Jiang, J.; E O’Connor, S.; Buell, C.R. The Mitragyna speciosa (Kratom) Genome: A resource for data-mining potent pharmaceuticals that impact human health. G3 Genes Genomes Genet. 2021, 11, 58. [Google Scholar] [CrossRef]
- Lau, K.H.; Bhat, W.W.; Hamilton, J.P.; Wood, J.; Vaillancourt, B.; Wiegert-Rininger, K.; Newton, L.; Hamberger, B.; Holmes, D.; Hamberger, B.; et al. Genome assembly of Chiococca alba uncovers key enzymes involved in the biosynthesis of unusual terpenoids. DNA Res. 2020, 27, 13. [Google Scholar] [CrossRef]
- Erickson, D.L. DNA testing methodologies in quality control programs. HerbalGram 2020, 128, 58–63. [Google Scholar]
- Liu, H.; Wei, J.; Yang, T.; Mu, W.; Song, B.; Yang, T.; Fu, Y.; Wang, X.; Hu, G.; Li, W.; et al. Molecular digitization of a botanical garden: High-depth whole-genome sequencing of 689 vascular plant species from the Ruili Botanical Garden. GigaScience 2019, 8, 7. [Google Scholar] [CrossRef]
- Tran, H.T.M.; Furtado, A.; Vargas, C.A.C.; Smyth, H.; Lee, L.S.; Henry, R. SNP in the Coffea arabica genome associated with coffee quality. Tree Genet. Genomes 2018, 14, 72. [Google Scholar] [CrossRef]
- Rice, D.W.; Alverson, A.; Richardson, A.O.; Young, G.J.; Sanchez-Puerta, M.V.; Munzinger, J.; Barry, K.; Boore, J.L.; Zhang, Y.; Depamphilis, C.W.; et al. Horizontal transfer of entire genomes via mitochondrial fusion in the angiosperm Amborella. Science 2013, 342, 1468–1473. [Google Scholar] [CrossRef] [Green Version]
- Choi, I.-S.; Ruhlman, T.A.; Jansen, R.K. Comparative mitogenome analysis of the genus Trifolium reveals independent gene fission of ccmFn and intracellular gene transfers in Fabaceae. Int. J. Mol. Sci. 2020, 21, 1959. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-C.; Chen, H.; Yang, D.; Liu, C. Diversity of mitochondrial plastid DNAs (MTPTs) in seed plants. Mitochondrial DNA Part. A 2017, 29, 635–642. [Google Scholar] [CrossRef]
- Warren, J.M.; Salinas-Giegé, T.; Hummel, G.; Coots, N.L.; Svendsen, J.M.; Brown, K.C.; Drouard, L.; Sloan, D.B. Combining tRNA sequencing methods to characterize plant tRNA expression and post-transcriptional modification. RNA Biol. 2021, 18, 64–78. [Google Scholar] [CrossRef]
- Scarcelli, N.; Mariac, C.; Couvreur, T.; Faye, A.; Richard, D.; Sabot, F.; Berthouly-Salazar, C.; Vigouroux, Y. Intra-individual polymorphism in chloroplasts from NGS data: Where does it come from and how to handle it? Mol. Ecol. Resour. 2016, 16, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-S.; Jayakodi, M.; Lee, S.H.; Jeon, J.-H.; Lee, H.-O.; Park, J.Y.; Moon, B.C.; Kim, C.-K.; Wing, R.A.; Newmaster, S.G.; et al. Mitochondrial plastid DNA can cause DNA barcoding paradox in plants. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, K.L.; Song, K.; Roessler, P.G.; Nugent, J.M.; Doyle, J.L.; Palmer, J.D. Intracellular gene transfer in action: Dual transcription and multiple silencings of nuclear and mitochondrial cox2 genes in legumes. Proc. Natl. Acad. Sci. USA 1999, 96, 13863–13868. [Google Scholar] [CrossRef] [Green Version]
- Chapdelaine, Y.; Bonen, L. The wheat mitochondrial gene for subunit I of the NADH dehydrogenase complex: A trans-splicing model for this gene-in-pieces. Cell 1991, 65, 465–472. [Google Scholar] [CrossRef]
- Conklin, P.L.; Wilson, R.K.; Hanson, M. Multiple trans-splicing events are required to produce a mature nad1 transcript in a plant mitochondrion. Genes Dev. 1991, 5, 1407–1415. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.-E.; Meng, Y.; Peng, D.-L.; Nie, Z.-L.; Sun, H. Molecular phylogeny of Galium L. of the tribe Rubieae (Rubia-ceae)–Emphasis on Chinese species and recognition of a new genus Pseudogalium. Mol. Phylogenet. Evol. 2018, 126, 221–232. [Google Scholar] [CrossRef]
- Arrieta-Montiel, M.P.; Shedge, V.; Davila, J.; Christensen, A.C.; A Mackenzie, S. Diversity of the Arabidopsis mitochondrial genome occurs via nuclear-controlled recombination activity. Genetics 2009, 183, 1261–1268. [Google Scholar] [CrossRef] [Green Version]
- André, C.; Levy, A.; Walbot, V. Small repeated sequences and the structure of plant mitochondrial genomes. Trends Genet. 1992, 8, 128–132. [Google Scholar] [CrossRef]
- Takada, M.; Asada, M.; Miyashita, T. Regional differences in the morphology of a shrub Damnacanthus indicus: An induced resistance to deer herbivory? Ecol. Res. 2001, 16, 809–813. [Google Scholar] [CrossRef]
- Lee, D.-J.; Jeon, S. Estimating changes in habitat quality through land-use predictions: Case study of roe deer (Capreolus pygargus tianschanicus) in Jeju Island. Sustainability 2020, 12, 10123. [Google Scholar] [CrossRef]
- Kabemura, Y.; Enoki, T.; Osaki, S.; Yamauchi, K.; Ohgi, D.; Koga, S.; Hishi, T.; Inoue, S.; Yasuda, Y.; Utsumi, Y. Increase of sika deer population on plantation trees and understory in Kasuya Research forest of Kyushu University in Fukuoka Prefecture, Japan. Bull. Kyushu Univ. For. 2018, 99, 18–21. [Google Scholar]
- Lou, S.-M.; Li, H.; Qin, X.-M.; Huang, Z.-P.; Jiang, S.-Y.; Huang, X.-Y. The complete chloroplast genome of Damnacanthus indicus CF Gaertn (Rubiaceae). Mitochondrial DNA Part B 2021, 6, 1251–1252. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq–versatile and accurate an-notation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Ruhlman, T.A.; Sabir, J.S.; Mutwakil, M.H.; Baeshen, M.N.; Sabir, M.J.; Baeshen, N.A.; Jansen, R.K. Complete sequences of organelle genomes from the medicinal plant Rhazya stricta (Apocynaceae) and contrasting patterns of mitochondrial genome evolution across asterids. BMC Genom. 2014, 15, 405. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leigh, J.W.; Bryant, D. popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acronym | Infraspecific Classification | Origin | Sequencing Platform | Number of Raw Reads |
|---|---|---|---|---|
| DII-Kyu1 | var. indicus | Kyushu, Japan | DNB (MGI) | 49,788,516 |
| DII-Kyu2 | var. indicus | Kyushu, Japan | Miseq (Illumina) | 7,603,662 |
| DII-Je1 | var. indicus | Jeju, Korea | DNB (MGI) | 47,817,598 |
| DII-Je2 | var. indicus | Jeju, Korea | Miseq (Illumina) | 8,196,070 |
| DIM-Kyu1 | var. microphyllus | Kyushu, Japan | DNB (MGI) | 44,651,432 |
| DIM-Kyu2 | var. microphyllus | Kyushu, Japan | Miseq (Illumina) | 7,127,138 |
| Acronym | Length (bp) | Coverage | Repeat (bp) | Plastid Origin (bp) | GenBank Accession |
|---|---|---|---|---|---|
| DII-Kyu1 | 419,435 | 275 | 17,822 | 22,689 | MZ285073 |
| DII-Kyu2 | 419,429 | 71 | 17,822 | 22,688 | MZ285074 |
| DII-Je1 | 417,815 | 300 | 17,832 | 22,705 | MZ285071 |
| DII-Je2 | 419,010 | 66 | 18,082 | 22,487 | MZ285072 |
| DIM-Kyu1 | 417,661 | 250 | 17,750 | 22,706 | MZ285075 |
| DIM-Kyu2 | 417,816 | 82 | 17,832 | 22,706 | MZ285076 |
| Species | Gene Status a | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| rpl2 | rpl5 | rpl10 | rpl16 | rps1 | rps2 | rps3 | rps4 | rps7 | rps10 | rps11 | rps12 | rps13 | rps14 | rps19 | sdh3 | sdh4 | |
| Damnacanthus indicus | ● | ● | - | ● | ● | - | ● | ● | ● | ● | - | ● | ● | ● | - | - | ● |
| Rhazya stricta | ● | ● | ● | ● | ● | - | ● | ● | ● | ● | - | ● | ● | ● | ● | ● | ● |
| Asclepias syriaca | ● | ● | ● | ● | ● | - | ● | ● | ● | ● | - | ● | ● | - | ● | ● | ● |
| Nicotiana tabacum | ● | ● | ● | ● | ● | - | ● | ● | ● | ● | - | ● | ● | ● | - | ● | ● |
| Mimulus guttatus | ● | ● | ● | ● | - | - | ● | ● | - | ● | - | ● | ● | ● | - | ● | ● |
| Boea hygrometrica | ● | ● | ● | ● | - | - | ● | ● | ● | ● | - | ● | ● | ● | - | ● | ● |
| Daucus carota | - | ● | ● | ● | ● | - | ● | ● | ● | - | - | ● | ● | - | - | - | - |
| Helianthus annuus | - | ● | ● | ● | - | - | ● | ● | - | - | - | ● | ● | - | ● | - | ● |
| Vaccinium macrocarpon | ● | ● | ● | ● | ● | - | ● | ● | - | ● | - | ● | ● | ● | ● | ● | ● |
| Liriodendron tulipifera | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● |
| Species | Subfamily | Nad1i728 (cis/trans) | GenBank Accession | SRA a Run Number | Original Publication of SRA Data |
|---|---|---|---|---|---|
| Galium porrigens var. tenue | Rubioideae | Trans | MZ292736 and MZ292737 | SRR9961329 | Burge (2020) [46] |
| Ophiorrhiza pumila | Rubioideae | Trans | MZ292738 and MZ292739 | DRR194739 | Rai et al. (2021) [47] |
| Gynochthodes cochinchinensis | Rubioideae | Trans | MZ292740 and MZ292741 | SRR12903483 | Bautista et al. (2021) [48] |
| Foonchewia coriacea | Rubioideae | Trans | MZ292742 and MZ292743 | SRR12917150 | Zhang et al. (2021) [49] |
| Mitragyna speciosa | Cinchonoideae | Cis | MZ292744 | SRR12673030 | Brose et al. (2021) [50] |
| Chiococca alba | Cinchonoideae | Cis | MZ292745 | SRR9087163 | Lau et al. (2020) [51] |
| Corynanthe mayumbensis | Cinchonoideae | Cis | MZ292746 | SRR8690411 | Erickson (2020) [52] |
| Catunaregam spinosa | Ixoroideae | Cis | MZ292747 | SRR7121945 | Liu et al. (2019) [53] |
| Coffea arabica | Ixoroideae | Cis | MZ292748 | SRR7637601 | Tran et al. (2018) [54] |
| Diplospora mollissima | Ixoroideae | Cis | MZ292749 | SRR7121910 | Liu et al. (2019) [53] |
| Acronym | Analyzed Reads (Number) | A-Type (Number) | A-Type (%) | B-Type (Number) | B-Type (%) |
|---|---|---|---|---|---|
| DII-Kyu1 | 114 | 113 | 99.1 | 1 | 0.9 |
| DII-Kyu2 | 78 | 77 | 98.7 | 1 | 1.3 |
| DII-Je1 | 116 | 116 | 100.0 | 0 | 0.0 |
| DII-Je2 | 70 | 70 | 100.0 | 0 | 0.0 |
| DIM-Kyu1 | 101 | 101 | 100.0 | 0 | 0.0 |
| DIM-Kyu2 | 121 | 6 | 5.0 | 115 | 95.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, E.-K.; Cho, W.-B.; Tamaki, I.; Choi, I.-S.; Lee, J.-H. Comparative Mitogenomic Analysis Reveals Gene and Intron Dynamics in Rubiaceae and Intra-Specific Diversification in Damnacanthus indicus. Int. J. Mol. Sci. 2021, 22, 7237. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137237
Han E-K, Cho W-B, Tamaki I, Choi I-S, Lee J-H. Comparative Mitogenomic Analysis Reveals Gene and Intron Dynamics in Rubiaceae and Intra-Specific Diversification in Damnacanthus indicus. International Journal of Molecular Sciences. 2021; 22(13):7237. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137237
Chicago/Turabian StyleHan, Eun-Kyeong, Won-Bum Cho, Ichiro Tamaki, In-Su Choi, and Jung-Hyun Lee. 2021. "Comparative Mitogenomic Analysis Reveals Gene and Intron Dynamics in Rubiaceae and Intra-Specific Diversification in Damnacanthus indicus" International Journal of Molecular Sciences 22, no. 13: 7237. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137237