
New Partners Identified by Mass Spectrometry Assay Reveal Functions of NCAM2 in Neural Cytoskeleton Organization

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
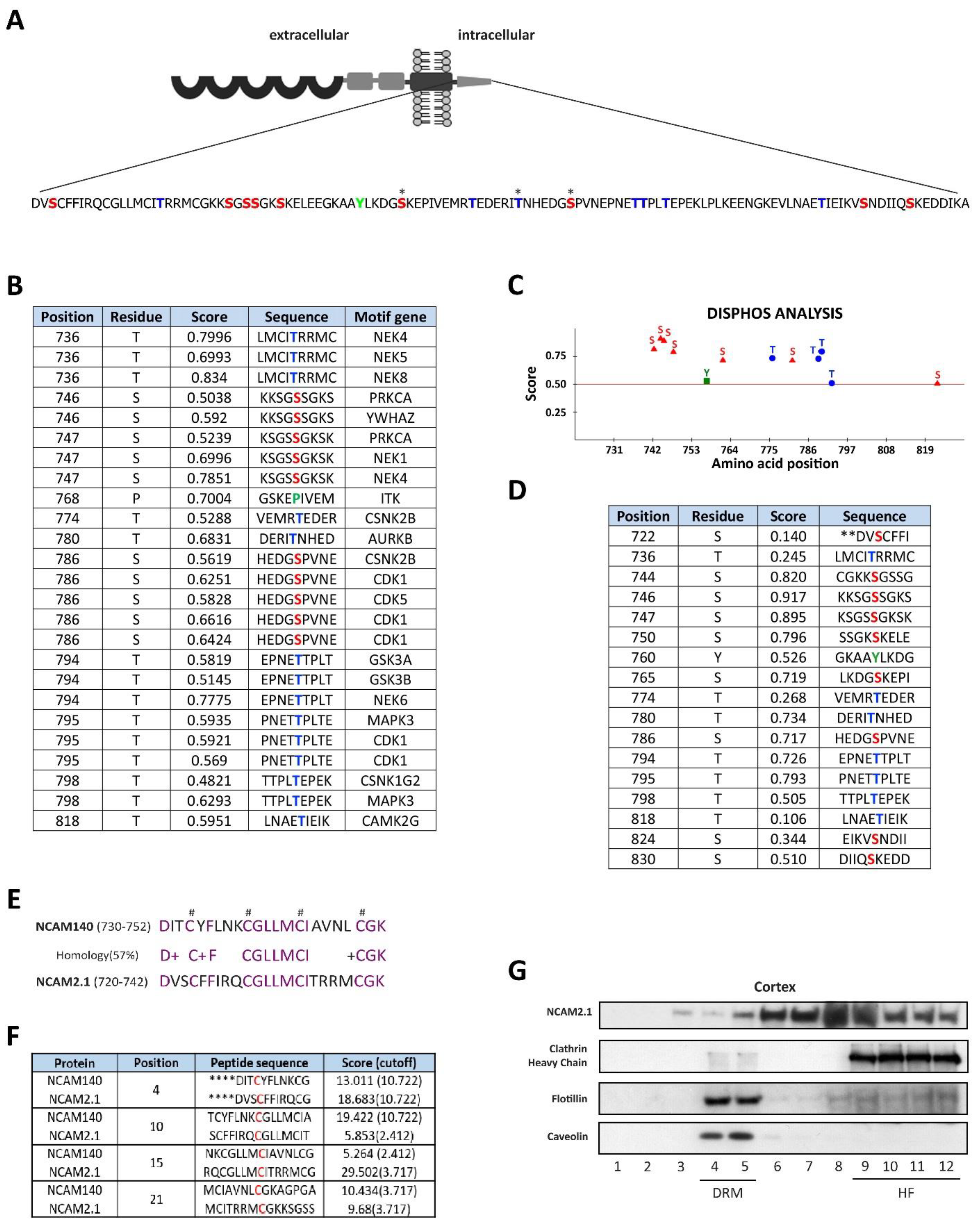
2.1. In Silico Analysis of the Cytoplasmic Domains of NCAM2.1
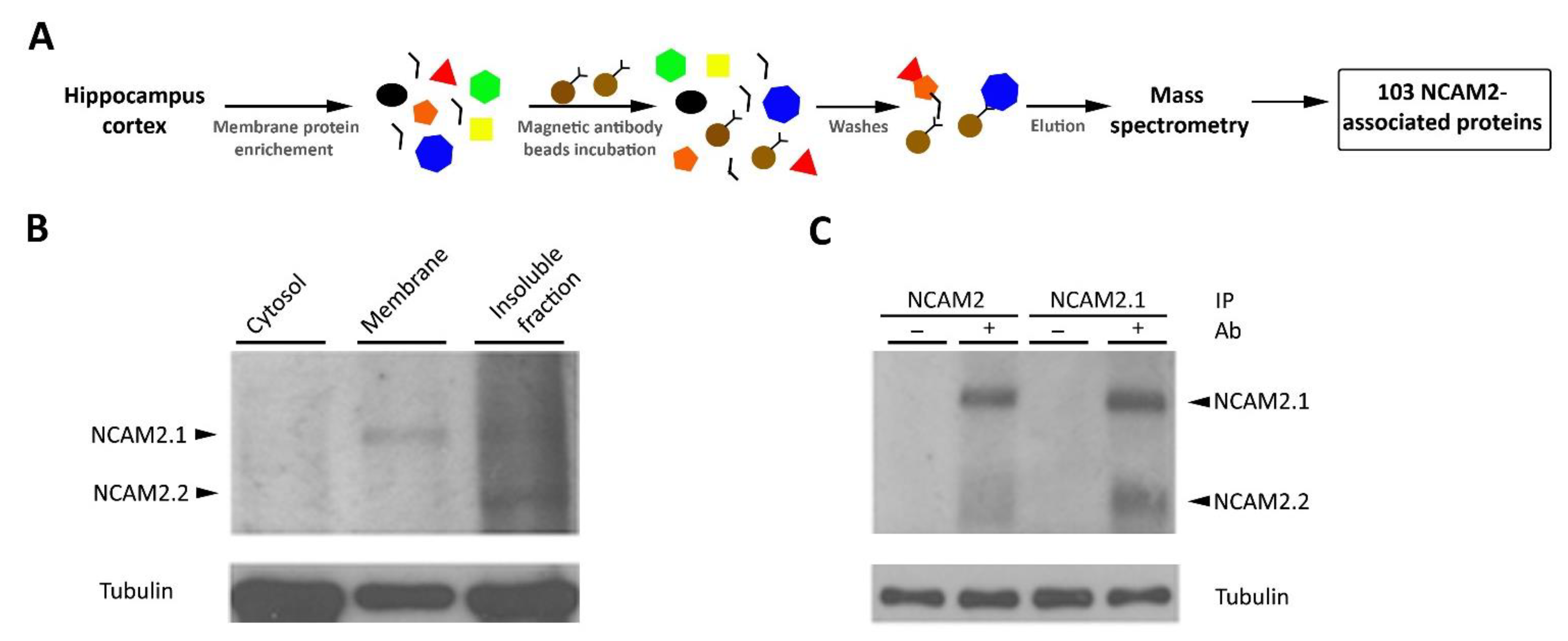
2.2. NCAM2 Interactome in Postnatal Cerebral Cortex
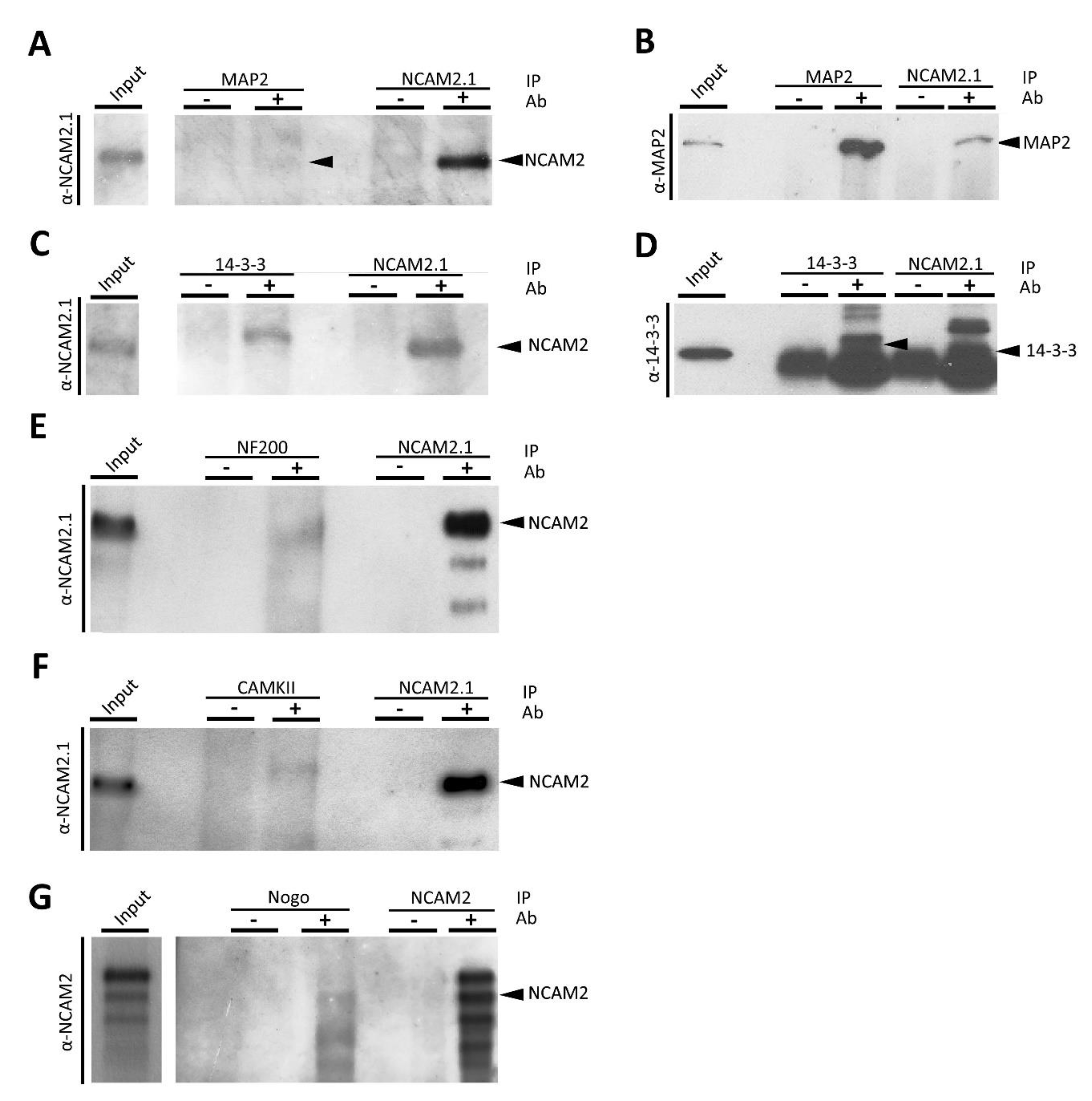
2.3. NCAM2 Interacts with Cytoskeleton and Cytoskeleton-Associated Proteins
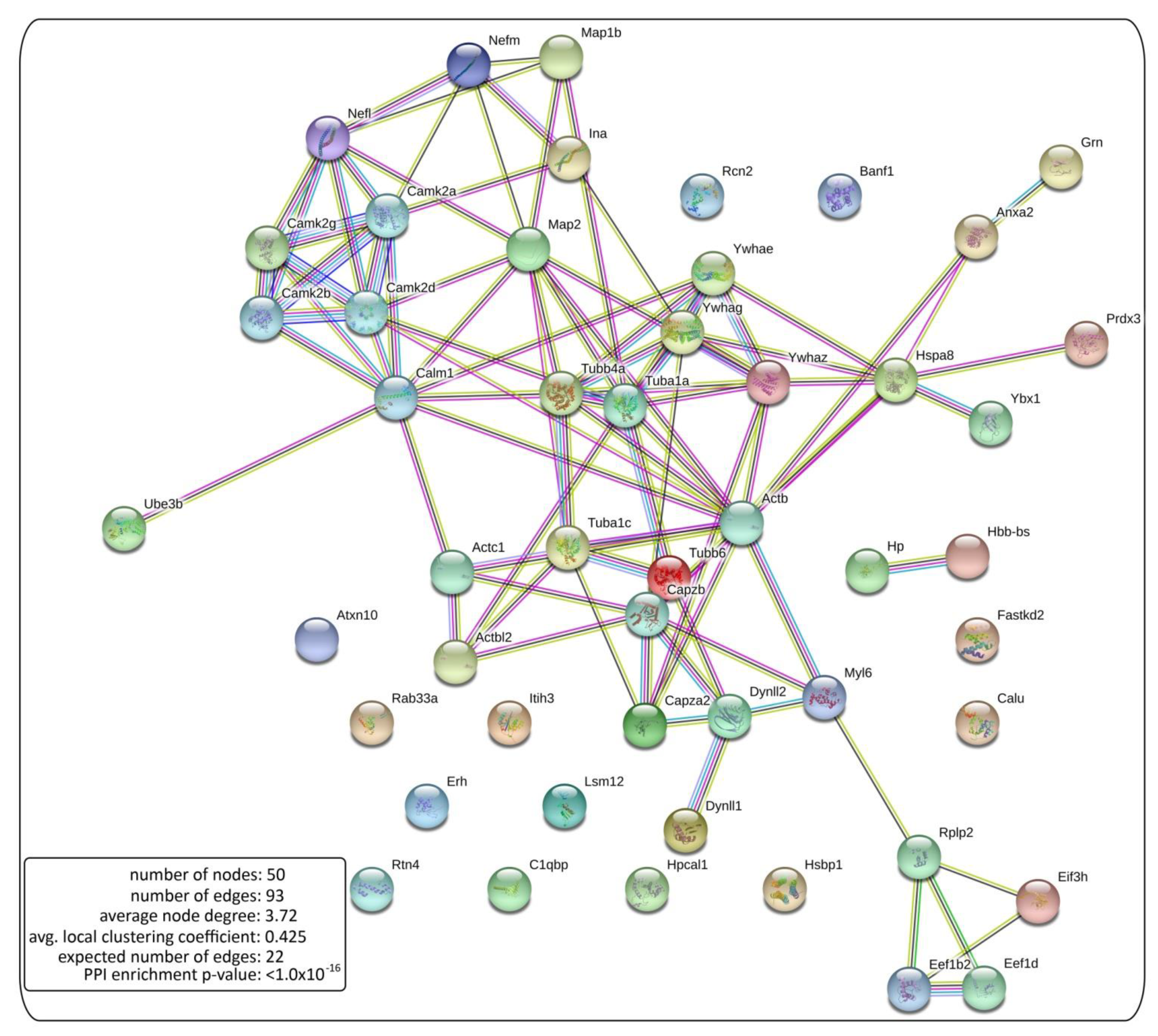
2.4. Bioinformatic Analysis of the NCAM2 Interactome
2.4.1. Bit Map
2.4.2. Gene Ontology Terms
2.4.3. Pathways Analysis
3. Discussion
4. Materials and Methods
4.1. Antibodies
4.2. Mass Spectrometry Assay
4.3. Immunoprecipitation for Western Blotting
4.4. Sucrose Gradient for Lipid Raft Isolation
4.5. Bioinformatic Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Namba, T.; Funahashi, Y.; Nakamuta, S.; Xu, C.; Takano, T.; Kaibuchi, K. Extracellular and intracellular signaling for neuronal polarity. Physiol. Rev. 2015, 95, 995–1024. [Google Scholar] [CrossRef] [Green Version]
- Takano, T.; Xu, C.; Funahashi, Y.; Namba, T.; Kaibuchi, K. Neuronal polarization. Development 2015, 142, 2088–2093. [Google Scholar] [CrossRef] [Green Version]
- Zinn, K.; Özkan, E. Neural immunoglobulin superfamily interaction networks. Curr. Opin. Neurobiol. 2017, 45, 99–105. [Google Scholar] [CrossRef]
- Missaire, M.; Hindges, R. The role of cell adhesion molecules in visual circuit formation: From neurite outgrowth to maps and synaptic specificity. Dev. Neurobiol. 2015, 75, 569–583. [Google Scholar] [CrossRef] [Green Version]
- Sytnyk, V.; Leshchyns’ka, I.; Schachner, M. Neural Cell Adhesion Molecules of the Immunoglobulin Superfamily Regulate Synapse Formation, Maintenance, and Function. Trends Neurosci. 2017, 40, 295–308. [Google Scholar] [CrossRef]
- Gu, Z.; Imai, F.; Kim, I.J.; Fujita, H.; Katayama, K.I.; Mori, K.; Yoshihara, Y.; Yoshida, Y. Expression of the immunoglobulin superfamily cell adhesion molecules in the developing spinal cord and dorsal root ganglion. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, L.; Love, J.; Colman, D.R. Adhesion molecules in the nervous system: Structural insights into function and diversity. Annu. Rev. Neurosci. 2007, 30, 451–474. [Google Scholar] [CrossRef] [PubMed]
- Leshchyns’ka, I.; Sytnyk, V. Reciprocal interactions between cell adhesion molecules of the immunoglobulin superfamily and the cytoskeleton in neurons. Front. Cell Dev. Biol. 2016, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Frei, J.A.; Stoeckli, E.T. SynCAMs—From axon guidance to neurodevelopmental disorders. Mol. Cell. Neurosci. 2017, 81, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Makino, T.; McLysaght, A. Ohnologs in the human genome are dosage balanced and frequently associated with disease. Proc. Natl. Acad. Sci. USA 2010, 107, 9270–9274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pébusque, M.J.; Coulier, F.; Birnbaum, D.; Pontarotti, P. Ancient large-scale genome duplications: Phylogenetic and linkage analyses shed light on chordate genome evolution. Mol. Biol. Evol. 1998, 15, 1145–1159. [Google Scholar] [CrossRef] [Green Version]
- Alenius, M.; Bohm, S. Identification of a novel neural cell adhesion molecule-related gene with a potential role in selective axonal projection. J. Biol. Chem. 1997, 272, 26083–26086. [Google Scholar] [CrossRef] [Green Version]
- Yoshihara, Y.; Kawasaki, M.; Tamada, A.; Fujita, H.; Hayashi, H.; Kagamiyama, H.; Mori, K. OCAM: A new member of the neural cell adhesion molecule family related to zone-to-zone projection of olfactory and vomeronasal axons. J. Neurosci. 1997, 17, 5830–5842. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, O.S.; Bock, E. Brain specific synaptosomal membrane proteins demonstrated by crossed immunoelectrophoresis. J. Neurochem. 1974, 23, 879–880. [Google Scholar] [CrossRef]
- Sheng, L.; Leshchyns’Ka, I.; Sytnyk, V. Cell adhesion and intracellular calcium signaling in neurons. Cell Commun. Signal. 2013, 11, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, C.; Berezin, V.; Bock, E. Neural cell adhesion molecule differentially interacts with isoforms of the fibroblast growth factor receptor. Neuroreport 2011, 22, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Ramser, E.M.; Buck, F.; Schachner, M.; Tilling, T. Binding of αII spectrin to 14-3-3β is involved in NCAM-dependent neurite outgrowth. Mol. Cell. Neurosci. 2010, 45, 66–74. [Google Scholar] [CrossRef]
- Li, J.; Zhang, W.; Yang, H.; Howrigan, D.P.; Wilkinson, B.; Souaiaia, T.; Evgrafov, O.V.; Genovese, G.; Clementel, V.A.; Tudor, J.C.; et al. Spatiotemporal profile of postsynaptic interactomes integrates components of complex brain disorders. Nat. Neurosci. 2017, 20, 1150–1161. [Google Scholar] [CrossRef] [PubMed]
- Visser, J.J.; Cheng, Y.; Perry, S.C.; Chastain, A.B.; Parsa, B.; Masri, S.S.; Ray, T.A.; Kay, J.N.; Wojtowicz, W.M. An extracellular biochemical screen reveals that FLRTs and Unc5s mediate neuronal subtype recognition in the retina. eLife 2015, 4. [Google Scholar] [CrossRef]
- Kleene, R.; Mzoughi, M.; Joshi, G.; Kalus, I.; Bormann, U.; Schulze, C.; Xiao, M.F.; Dityatev, A.; Schachner, M. NCAM-induced neurite outgrowth depends on binding of calmodulin to NCAM and on nuclear import of NCAM and fak fragments. J. Neurosci. 2010, 30, 10784–10798. [Google Scholar] [CrossRef] [Green Version]
- Alenius, M.; Bohm, S. Differential function of RNCAM isoforms in precise target selection of olfactory sensory neurons. Development 2003, 130, 917–927. [Google Scholar] [CrossRef] [Green Version]
- Von Campenhausen, H.; Yoshihara, Y.; Mori, K. OCAM reveals segregated mitral/tufted cell pathways in developing accessory olfactory bulb. Neuroreport 1997, 8, 2607–2612. [Google Scholar] [CrossRef]
- Kulahin, N.; Walmod, P.S. The neural cell adhesion molecule NCAM2/OCAM/RNCAM, a close relative to NCAM. Adv. Exp. Med. Biol. 2010, 663, 403–420. [Google Scholar] [CrossRef] [PubMed]
- Winther, M.; Berezin, V.; Walmod, P.S. NCAM2/OCAM/RNCAM: Cell adhesion molecule with a role in neuronal compartmentalization. Int. J. Biochem. Cell Biol. 2012, 44, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Parcerisas, A.; Pujadas, L.; Ortega-Gascó, A.; Perelló-Amorós, B.; Viais, R.; Hino, K.; Figueiro-Silva, J.; La Torre, A.; Trullás, R.; Simó, S.; et al. NCAM2 Regulates Dendritic and Axonal Differentiation through the Cytoskeletal Proteins MAP2 and 14-3-3. Cereb. Cortex 2020, 30, 3781–3799. [Google Scholar] [CrossRef] [PubMed]
- Sheng, L.; Leshchyns’Ka, I.; Sytnyk, V. Neural cell adhesion molecule 2 promotes the formation of filopodia and neurite branching by inducing submembrane increases in Ca2+ levels. J. Neurosci. 2015, 35, 1739–1752. [Google Scholar] [CrossRef] [Green Version]
- Sheng, L.; Leshchyns’ka, I.; Sytnyk, V. Neural Cell Adhesion Molecule 2 (NCAM2)-Induced c-Src-Dependent Propagation of Submembrane Ca 2+ Spikes Along Dendrites Inhibits Synapse Maturation. Cereb. Cortex 2019, 29, 1439–1459. [Google Scholar] [CrossRef] [PubMed]
- Leshchyns’Ka, I.; Liew, H.T.; Shepherd, C.; Halliday, G.M.; Stevens, C.H.; Ke, Y.D.; Ittner, L.M.; Sytnyk, V. Aβ-dependent reduction of NCAM2-mediated synaptic adhesion contributes to synapse loss in Alzheimer’s disease. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, C.; Steinemann, D.; Mälzer, M.; Roy, M.; Arslan-Kirchner, M.; Illig, T.; Schmidtke, J.; Stuhrmann, M. NCAM2 deletion in a boy with macrocephaly and autism: Cause, association or predisposition? Eur. J. Med. Genet. 2016, 59, 493–498. [Google Scholar] [CrossRef]
- Molloy, C.A.; Keddache, M.; Martin, L.J. Evidence for linkage on 21q and 7q in a subset of autism characterized by developmental regression. Mol. Psychiatry 2005, 10, 741–746. [Google Scholar] [CrossRef] [Green Version]
- Hussman, J.P.; Chung, R.H.; Griswold, A.J.; Jaworski, J.M.; Salyakina, D.; Ma, D.; Konidari, I.; Whitehead, P.L.; Vance, J.M.; Martin, E.R.; et al. A noise-reduction GWAS analysis implicates altered regulation of neurite outgrowth and guidance in autism. Mol. Autism 2011, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- Petit, F.; Plessis, G.; Decamp, M.; Cuisset, J.M.; Blyth, M.; Pendlebury, M.; Andrieux, J. 21q21 deletion involving NCAM2: Report of 3 cases with neurodevelopmental disorders. Eur. J. Med. Genet. 2015, 58, 44–46. [Google Scholar] [CrossRef]
- Huttlin, E.L.; Jedrychowski, M.P.; Elias, J.E.; Goswami, T.; Rad, R.; Beausoleil, S.A.; Villén, J.; Haas, W.; Sowa, M.E.; Gygi, S.P. A tissue-specific atlas of mouse protein phosphorylation and expression. Cell 2010, 143, 1174–1189. [Google Scholar] [CrossRef] [Green Version]
- Kunz, S.; Ziegler, U.; Kunz, B.; Sonderegger, P. Intracellular Signaling Is Changed after Clustering of the Neural Cell Adhesion Molecules Axonin-1 and NgCAM during Neurite Fasciculation. J. Cell Biol. 1996, 135, 253–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D. Cdk5 in neuroskeletal dynamics. NeuroSignals 2003, 12, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Lickert, H.; Bauer, A.; Kemler, R.; Stappert, J. Casein kinase II phosphorylation of E-cadherin increases E-cadherin/β- catenin interaction and strengthens cell-cell adhesion. J. Biol. Chem. 2000, 275, 5090–5095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogh, B.S.; Multhaupt, H.A.B.; Couchman, J.R. Protein Kinase C, Focal Adhesions and the Regulation of Cell Migration. J. Histochem. Cytochem. 2014, 62, 172–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, Y.T.; Gupta, A.; Zhou, Y.; Nikolic, M.; Tsai, L.H. Regulation of N-cadherin-mediated adhesion by the p35-Cdk5 kinase. Curr. Biol. 2000, 10, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Li, M.; Vrana, J.; Schaller, M.D. Glycogen Synthase Kinase 3- and Extracellular Signal-Regulated Kinase-Dependent Phosphorylation of Paxillin Regulates Cytoskeletal Rearrangement. Mol. Cell. Biol. 2006, 26, 2857–2868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delling, M.; Wischmeyer, E.; Dityatev, A.; Sytnyk, V.; Veh, R.W.; Karschin, A.; Schachner, M. The neural cell adhesion molecule regulates cell-surface delivery of G-protein-activated inwardly rectifying potassium channels via lipid rafts. J. Neurosci. 2002, 22, 7154–7164. [Google Scholar] [CrossRef] [Green Version]
- Niethammer, P.; Delling, M.; Sytnyk, V.; Dityatev, A.; Fukami, K.; Schachner, M. Cosignaling of NCAM via lipid rafts and the FGF receptor is required for neuritogenesis. J. Cell Biol. 2002, 157, 521–532. [Google Scholar] [CrossRef]
- Kamiguchi, H. The region-specific activities of lipid rafts during axon growth and guidance. J. Neurochem. 2006, 98, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Hérincs, Z.; Corset, V.; Cahuzac, N.; Furne, C.; Castellani, V.; Hueber, A.O.; Mehlen, P. DCC association with lipid rafts is required for netrin-1-mediated axon guidance. J. Cell Sci. 2005, 118, 1687–1692. [Google Scholar] [CrossRef] [Green Version]
- Caldwell, J.E.; Heiss, S.G.; Mermall, V.; Cooper, J.A. Effects of CapZ, an Actin Capping Protein of Muscle, on the Polymerization of Actin. Biochemistry 1989, 28, 8506–8514. [Google Scholar] [CrossRef]
- Wear, M.A.; Yamashita, A.; Kim, K.; Maéda, Y.; Cooper, J.A. How capping protein binds the barbed end of the actin filament. Curr. Biol. 2003, 13, 1531–1537. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.A.; Wilson, M.H.; Giraud, J.; Xie, Z.; Tseng, H.C.; England, C.; Herscovitz, H.; Tsai, L.H.; Delalle, I. Capzb2 interacts with β-tubulin to regulate growth cone morphology and neurite outgrowth. PLoS Biol. 2009, 7, e1000208. [Google Scholar] [CrossRef] [Green Version]
- Sinnar, S.A.; Antoku, S.; Saffin, J.M.; Cooper, J.A.; Halpain, S. Capping protein is essential for cell migration in vivo and for filopodial morphology and dynamics. Mol. Biol. Cell 2014, 25, 2152–2160. [Google Scholar] [CrossRef]
- Yuan, A.; Rao, M.V.; Nixon, R.A. Neurofilaments and neurofilament proteins in health and disease. Cold Spring Harb. Perspect. Biol. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Rao, M.V.; Campbell, J.; Yuan, A.; Kumar, A.; Gotow, T.; Uchiyama, Y.; Nixon, R.A. The neurofilament middle molecular mass subunit carboxyl-terminal tail domains is essential for the radial growth and cytoskeletal architecture of axons but not for regulating neurofilament transport rate. J. Cell Biol. 2003, 163, 1021–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.C.; Redmond, L. Neuronal CaMKII acts as a structural kinase. Commun. Integr. Biol. 2009, 2, 40–41. [Google Scholar] [CrossRef]
- Wu, G.Y.; Cline, H.T. Stabilization of dendritic arbor structure in vivo by CaMKII. Science 1998, 279, 222–226. [Google Scholar] [CrossRef] [Green Version]
- Schwab, M.E. Functions of Nogo proteins and their receptors in the nervous system. Nat. Rev. Neurosci. 2010, 11, 799–811. [Google Scholar] [CrossRef] [PubMed]
- Coyne, A.N.; Lorenzini, I.; Chou, C.C.; Torvund, M.; Rogers, R.S.; Starr, A.; Zaepfel, B.L.; Levy, J.; Johannesmeyer, J.; Schwartz, J.C.; et al. Post-transcriptional Inhibition of Hsc70-4/HSPA8 Expression Leads to Synaptic Vesicle Cycling Defects in Multiple Models of ALS. Cell Rep. 2017, 21, 110–125. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Daniels, C.K.; Cao, S. Comprehensive review on the HSC70 functions, interactions with related molecules and involvement in clinical diseases and therapeutic potential. Pharmacol. Ther. 2012, 136, 354–374. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Zhang, X. Factors Regulating Neurogenesis in the Adult Dentate Gyrus. Hippocampus Plast. Funct. 2018. [Google Scholar] [CrossRef] [Green Version]
- Twelvetrees, A.E.E.; Pernigo, S.; Sanger, A.; Guedes-Dias, P.; Schiavo, G.; Steiner, R.A.A.; Dodding, M.P.P.; Holzbaur, E.L.L.F. The Dynamic Localization of Cytoplasmic Dynein in Neurons Is Driven by Kinesin-1. Neuron 2016, 90, 1000–1015. [Google Scholar] [CrossRef] [Green Version]
- Grabham, P.W.; Seale, G.E.; Bennecib, M.; Goldberg, D.J.; Vallee, R.B. Cytoplasmic dynein and LIS1 are required for microtubule advance during growth cone remodeling and fast axonal outgrowth. J. Neurosci. 2007, 27, 5823–5834. [Google Scholar] [CrossRef]
- Roossien, D.H.; Lamoureux, P.; Miller, K.E. Cytoplasmic dynein pushes the cytoskeletal meshwork forward during axonal elongation. J. Cell Sci. 2014, 127, 3593–3602. [Google Scholar] [CrossRef] [Green Version]
- Maday, S.; Wallace, K.E.; Holzbaur, E.L.F. Autophagosomes initiate distally and mature during transport toward the cell soma in primary neurons. J. Cell Biol. 2012, 196, 407–417. [Google Scholar] [CrossRef]
- Uchida, A.; Alami, N.H.; Brown, A. Tight functional coupling of kinesin-1A and dynein motors in the bidirectional transport of neurofilaments. Mol. Biol. Cell 2009, 20, 4997–5006. [Google Scholar] [CrossRef] [Green Version]
- Griesi-Oliveira, K.; Suzuki, A.M.; Alves, A.Y.; Mafra, A.C.C.N.; Yamamoto, G.L.; Ezquina, S.; Magalhães, Y.T.; Forti, F.L.; Sertie, A.L.; Zachi, E.C.; et al. Actin cytoskeleton dynamics in stem cells from autistic individuals. Sci. Rep. 2018, 8, 11138. [Google Scholar] [CrossRef]
- Moon, H.M.; Wynshaw-Boris, A. Cytoskeleton in action: Lissencephaly, a neuronal migration disorder. Wiley Interdiscip. Rev. Dev. Biol. 2013, 2, 229–245. [Google Scholar] [CrossRef] [Green Version]
- Robinson, R. Loss of microtubule-to-actin linkage disrupts cortical development. PLoS Biol. 2011, 9, e1001175. [Google Scholar] [CrossRef] [PubMed]
- Ichinohe, N.; Yoshihara, Y.; Hashikawa, T.; Rockland, K.S. Developmental study of dendritic bundles in layer 1 of the rat granular retrosplenial cortex with special reference to a cell adhesion molecule, OCAM. Eur. J. Neurosci. 2003, 18, 1764–1774. [Google Scholar] [CrossRef]
- Hell, J.W. CaMKII: Claiming center stage in postsynaptic function and organization. Neuron 2014, 81, 249–265. [Google Scholar] [CrossRef] [Green Version]
- Schlossman, D.M.; Schmid, S.L.; Braell, W.A.; Rothman, J.E. An enzyme that removes clathrin coats: Purification of an uncoating ATPase. J. Cell Biol. 1984, 99, 723–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zinsmaier, K.E.; Bronk, P. Molecular chaperones and the regulation of neurotransmitter exocytosis. Biochem. Pharmacol. 2001, 62, 1–11. [Google Scholar] [CrossRef]
- Tortosa, E.; Montenegro-Venegas, C.; Benoist, M.; Härtel, S.; González-Billault, C.; Esteban, J.A.; Avila, J. Microtubule-associated protein 1B (MAP1B) is required for dendritic spine development and synaptic maturation. J. Biol. Chem. 2011, 286, 40638–40648. [Google Scholar] [CrossRef] [Green Version]
- Edwards, M.; Zwolak, A.; Schafer, D.A.; Sept, D.; Dominguez, R.; Cooper, J.A. Capping protein regulators fine-tune actin assembly dynamics. Nat. Rev. Mol. Cell Biol. 2014, 15, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, Y.; Yamauchi, Y.; Shinkawa, T.; Taoka, M.; Donai, H.; Takahashi, N.; Isobe, T.; Yamauchi, T. Molecular constituents of the postsynaptic density fraction revealed by proteomic analysis using multidimensional liquid chromatography-tandem mass spectrometry. J. Neurochem. 2004, 88, 759–768. [Google Scholar] [CrossRef]
- Kitanishi, T.; Sakai, J.; Kojima, S.; Saitoh, Y.; Inokuchi, K.; Fukaya, M.; Watanabe, M.; Matsuki, N.; Yamada, M.K. Activity-dependent localization in spines of the F-actin capping protein CapZ screened in a rat model of dementia. Genes Cells 2010, 15, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Tang, X.; Vitriol, E.; Chen, G.; Zheng, J.Q. Actin capping protein is required for dendritic spine development and synapse formation. J. Neurosci. 2011, 31, 10228–10233. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, M.; Hikita, T.; Taya, S.; Uraguchi-asaki, J.; Toyo-Oka, K.; Wynshaw-boris, A.; Ujike, H.; Inada, T.; Takao, K.; Miyakawa, T.T.; et al. Identification of YWHAE, a gene encoding 14-3-3epsilon, as a possible susceptibility gene for schizophrenia. Hum. Mol. Genet. 2008, 17, 3212–3222. [Google Scholar] [CrossRef] [Green Version]
- Wachi, T.; Cornell, B.; Toyo-oka, K. Complete ablation of the 14-3-3epsilon protein results in multiple defects in neuropsychiatric behaviors. Behav. Brain Res. 2017, 319, 31–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foote, M.; Qiao, H.; Graham, K.; Wu, Y.; Zhou, Y. Inhibition of 14-3-3 Proteins Leads to Schizophrenia-Related Behavioral Phenotypes and Synaptic Defects in Mice. Biol. Psychiatry 2015, 78, 386–395. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Portela, M.; Janikiewicz, J.; Doig, J.; Abbott, C.M. Characterisation of translation elongation factor eEF1B subunit expression in mammalian cells and tissues and co-localisation with eEF1A2. PLoS ONE 2014, 9, e114117. [Google Scholar] [CrossRef] [Green Version]
- Kuang, L.; Hashimoto, K.; Huang, E.J.; Gentry, M.S.; Zhu, H. Frontotemporal dementia non-sense mutation of progranulin rescued by aminoglycosides. Hum. Mol. Genet. 2020, 29, 624–634. [Google Scholar] [CrossRef]
- Galimberti, D.; Fumagalli, G.G.; Fenoglio, C.; Cioffi, S.M.G.; Arighi, A.; Serpente, M.; Borroni, B.; Padovani, A.; Tagliavini, F.; Masellis, M.; et al. Progranulin plasma levels predict the presence of GRN mutations in asymptomatic subjects and do not correlate with brain atrophy: Results from the GENFI study. Neurobiol. Aging 2018, 62, 245.e9–245.e12. [Google Scholar] [CrossRef]
- Premi, E.; Gazzina, S.; Bozzali, M.; Archetti, S.; Alberici, A.; Cercignani, M.; Bianchetti, A.; Gasparotti, R.; Turla, M.; Caltagirone, C.; et al. Cognitive Reserve in Granulin-Related Frontotemporal Dementia: From Preclinical to Clinical Stages. PLoS ONE 2013, 8, e74762. [Google Scholar] [CrossRef] [Green Version]
- Roberson, E.D.; Filiano, A.J.; Martens, L.H.; Young, A.H.; Warmus, B.A.; Zhou, P.; Diaz-Ramirez, G.; Jiao, J.; Zhang, Z.; Huang, E.J.; et al. Dissociation of frontotemporal dementia-related deficits and neuroinflammation in progranulin haploinsufficient mice. Ann. Intern. Med. 2013, 158, 5352–5362. [Google Scholar] [CrossRef] [Green Version]
- Kao, A.W.; McKay, A.; Singh, P.P.; Brunet, A.; Huang, E.J. Progranulin, lysosomal regulation and neurodegenerative disease. Nat. Rev. Neurosci. 2017, 18, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Petkau, T.L.; Leavitt, B.R. Progranulin in neurodegenerative disease. Trends Neurosci. 2014, 37, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Petkau, T.L.; Neal, S.J.; Milnerwood, A.; Mew, A.; Hill, A.M.; Orban, P.; Gregg, J.; Lu, G.; Feldman, H.H.; Mackenzie, I.R.A.; et al. Synaptic dysfunction in progranulin-deficient mice. Neurobiol. Dis. 2012, 45, 711–722. [Google Scholar] [CrossRef]
- Gass, J.; Lee, W.C.; Cook, C.; Finch, N.; Stetler, C.; Jansen-West, K.; Lewis, J.; Link, C.D.; Rademakers, R.; Nykjær, A.; et al. Progranulin regulates neuronal outgrowth independent of Sortilin. Mol. Neurodegener. 2012, 7, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van de Kooij, B.; Creixell, P.; van Vlimmeren, A.; Joughin, B.A.; Miller, C.J.; Haider, N.; Simpson, C.D.; Linding, R.; Stambolic, V.; Turk, B.E.; et al. Comprehensive substrate specificity profiling of the human nek kinome reveals unexpected signaling outputs. eLife 2019, 8. [Google Scholar] [CrossRef]
- Obenauer, J.C.; Cantley, L.C.; Yaffe, M.B. Scansite 2.0: Proteome-wide prediction of cell signalling interactions using short sequence motifs. Nucleic Acids Res. 2003, 31, 3635–3641. [Google Scholar] [CrossRef] [Green Version]
- Iakoucheva, L.M.; Radivojac, P.; Brown, C.J.; O’Connor, T.R.; Sikes, J.G.; Obradovic, Z.; Dunker, A.K. The importance of intrinsic disorder for protein phosphorylation. Nucleic Acids Res. 2004, 32, 1037–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein (Gene) | Ptd | Function |
|---|---|---|
| Microtubule-associated protein 2 (Map2) | 24 | actin binding, calmodulin binding, cytoskeletal regulatory, microtubule binding and protein kinase binding |
| Neurofilament light polypeptide (Nefl) | 20 | structural constituent of cytoskeleton andstructural constituent of postsynaptic intermediate filament cytoskeleton |
| Actin, cytoplasmic 1 (Actb) | 19 | structural constituent of cytoskeleton and structural constituent of postsynaptic actin cytoskeleton |
| Neurofilament medium polypeptide (Nefm) | 19 | microtubule binding and structural constituent of cytoskeleton |
| Tubulin beta-4A (Tubb4a) | 18 | structural constituent of cytoskeleton and GTPase activity |
| Alpha-internexin (Ina) | 17 | structural constituent of cytoskeleton, structural constituent of postsynaptic actin and intermediate filament cytoskeleton |
| Tubulin alpha-1A (Tuba1a) | 13 | structural constituent of cytoskeleton |
| Tubulin alpha-1C (Tuba1c) | 12 | structural constituent of cytoskeleton |
| Actin, alpha cardiac muscle 1 (Actc1) | 11 | structural constituent of cytoskeleton and myosin binding |
| Calcium/calmodulin-dependent protein kinase type II beta (Camk2b) | 11 | calmodulin-dependent protein kinase activity, protein serine/threonine kinase activity and structural constituent of postsynaptic actin cytoskeleton |
| Calcium/calmodulin-dependent protein kinase type II alpha(Camk2a) | 10 | calmodulin-dependent protein kinase activity, glutamate receptor binding and protein serine/threonine kinase activity |
| Microtubule-associated protein 1B (Map1b) | 9 | actin binding, cytoskeletal regulatory protein and binding |
| Beta-actin-like protein 2 (Actbl2) | 8 | structural constituent of postsynaptic actin cytoskeleton |
| F-actin-capping protein beta (Capzb) | 8 | actin binding and beta-tubulin binding |
| 14-3-3 protein zeta/delta (Ywhaz) | 8 | recognition of a phosphoserine or phosphothreonine motif |
| Tubulin beta-6 chain (Tubb6) | 7 | structural constituent of cytoskeleton |
| Calcium/calmodulin-dependent protein kinase type II delta (Camk2d) | 6 | calmodulin-dependent protein kinase activity and protein serine/threonine kinase activity |
| Calcium/calmodulin-dependent protein kinase type II gamma (Camk2g) | 6 | calmodulin-dependent protein kinase activity and protein serine/threonine kinase activity |
| Enhancer of rudimentary homolog (Erh) | 5 | Cell cycle |
| Thioredoxin-dependent peroxide reductase (Prdx3) | 5 | cysteine-type endopeptidase inhibitor activity involved in apoptotic process |
| Heat shock cognate 71 kDa protein (Hspa8) | 5 | protein quality control system (folding proteins and degredation), co-chaperone, signaling receptor binding and ubiquitin protein ligase binding |
| F-actin-capping protein alpha-2 (Capza2) | 4 | actin filament binding |
| Calumenin (Calu) | 4 | calcium ion binding and enzyme inhibitor activity |
| Heat shock factor-binding protein 1 (Hsbp1) | 3 | transcription corepressor activity |
| Granulins (Grn) | 3 | chaperone binding, cytokine activity, growth factor activity and RNA binding |
| Reticulocalbin-2 (Rcn2) | 3 | calcium ion binding |
| Elongation factor 1-beta (Eef1b) | 3 | guanyl-nucleotide exchange factor activity and translation elongation factor activity |
| 14-3-3 protein epsilon (Ywhae) | 3 | recognition of a phosphoserine or phosphothreonine motif |
| Hemoglobin beta-1 (Hbb-b1) | 3 | oxygen carrier activity |
| Nuclease-sensitive element-binding protein 1 (Ybx1) | 3 | DNA-binding transcription activator, transcription factor binding |
| Elongation factor 1-delta (Eef1d) | 3 | activating transcription factor bindingand translation elongation factor activity |
| Dynein light chain 1, cytoplasmic (Dynll1) | 2 | motor activity and scaffold protein binding |
| Myosin light polypeptide 6 (Myl6) | 2 | motor activity and actin-dependent ATPase activity |
| Barrier-to-autointegration factor (Banf1) | 2 | DNA binding and enzyme binding |
| Dynein light chain 2, cytoplasmic (Dynll2) | 2 | motor activity and scaffold protein binding |
| 60S acidic ribosomal protein P2 (Rplp2) | 2 | structural constituent of ribosome |
| Complement component 1 Q (C1qbp) | 2 | complement component C1q binding, mRNA binding and translation activator |
| Inter-alpha-trypsin inhibitor heavy chain H3 (Itih3) | 2 | binding protein between hyaluronan and other matrix protein |
| Reticulon-4 (Rtn4) | 2 | cadherin binding and neurite growth regulatory factor |
| Ig kappa chain V-III region PC 2413 (Kv3a5) | 1 | adaptive immune response |
| Ataxin-10 (Atxn10) | 1 | neuritogenesis |
| Protein LSM12 homolog (Lsm12) | 1 | interactor/competitor of ATXN2 |
| Eukaryotic translation initiation factor 3 H (Eif3h) | 1 | translation initiation factor activity |
| Annexin A2 (Anxa2) | 1 | calcium-dependent phospholipid and cytoskeletal protein binding |
| Calmodulin (Calm1) | 1 | calcium-dependent protein and protein kinase binding |
| FAST kinase domain-containing protein 2 (Fastkd2) | 1 | protein kinase activity |
| Haptoglobin (Hp) | 1 | hemoglobin binding |
| Hippocalcin-like protein 1 (Hpcal1) | 1 | calcium ion binding |
| Ras-related protein Rab-33A (Rab33a) | 1 | GTPase activity |
| Ubiquitin-protein ligase E3B (Ube3b) | 1 | ubiquitin conjugating enzyme activity |
| GOs Enrichment for Biological Process | ||
|---|---|---|
| GO | Term Description | Proteins |
| GO:0030030 | cell projection organization | Actb,Atxn10,Camk2a,Camk2b,Capzb,Dynll1,Dynll2,Map1b, Map2,Nefl,Nefm,Rtn4,Tubb4a |
| GO:0007010 | cytoskeleton organization | Actb,Actc1,Capzb,Ina,Map1b,Map2,Nefl,Nefm,Tuba1a, Tub1c,Tubb4a,Tubb6 |
| GO:0007017 | microtubule-based process | Dynll1,Dynll2,Map1b,Map2,Nefl,Nefm,Tuba1a,Tuba1c, Tubb4a,Tubb6 |
| GO:0060052 | neurofilament cytoskeleton organization | Ina,Nefl,Nefm |
| GO:0010769 | regulation of cell morphogenesis involved in differentiation | C1qbp,Camk2b,Map1b,Map2,Nefl,Nefm,Rtn4 |
| GO:0007399 | nervous system development | Actb,Atxn10,Camk2a,Camk2b,Camk2d,Camk2g,Capzb,Grn,Ina,Map1b,Map2,Nefl,Nefm,Rtn4,Ywhae,Ywhag |
| GO:0010970 | transport along microtubule | Dynll1,Dynll2,Map1b,Nefl,Nefm |
| GO:0031175 | neuron projection development | Actb,Atxn10,Camk2a,Capzb,Map1b,Map2,Nefl,Nefm,Rtn4 |
| GO:0048699 | generation of neurons | Actb,Atxn10,Camk2a,Camk2b,Capzb,Grn,Map1b,Map2, Nefl, Nefm,Rtn4,Ywhae,Ywhag |
| GO:0006414 | translational elongation | Eef1b2,Eef1d,Rplp2 |
| GO:0050770 | regulation of axonogenesis | Map1b,Map2,Nefl,Nefm,Rtn4 |
| GO:0061564 | axon development | Actb,Map1b,Map2,Nefl,Nefm,Rtn4 |
| GO:0045664 | regulation of neuron differentiation | Camk2b,Grn,Map1b,Map2,Nefl,Nefm,Rtn4,Ywhag |
| GO:0010975 | regulation of neuron projection development | Camk2b,Grn,Map1b,Map2,Nefl,Nefm,Rtn4 |
| GOs enrichment for Molecular Function | ||
| GO | Term Description | Proteins |
| GO:0005200 | structural constituent of cytoskeleton | Actb,Nefl,Tuba1a,Tuba1c,Tubb4a,Tubb6 |
| GO:0005198 | structural molecule activity | Actb,Ina,Myl6,Nefl,Nefm,Rplp2,Tuba1a,Tuba1c,Tubb4a, Tubb6 |
| GO:0004683 | calmodulin-dependent protein kinase activity | Camk2a,Camk2b,Camk2d,Camk2g |
| GO:0008092 | cytoskeletal protein binding | Actb,Actc1,Anxa2,Camk2d,Capza2,Capzb,Dynll1,Dynll2, Map1b,Map2,Ywhag |
| GO:0005519 | cytoskeletal regulatory protein binding | Map1b,Map2 |
| GO:0044325 | ion channel binding | Calm1,Camk2d,Ywhae,Ywhaz |
| GO:0003924 | GTPase activity | Rab33a,Tuba1a,Tuba1c,Tubb4a,Tubb6 |
| GO:0005509 | calcium ion binding | Anxa2,Calm1,Calu,Hpcal1,Myl6,Rcn2,Tubb4a |
| GO:0097110 | scaffold protein binding | Dynll1,Dynll2,Ywhae |
| GO:0003746 | translation elongation factor activity | Eef1b2,Eef1d |
| GOs enrichment for Cellular Component | ||
| GO | Term Description | Proteins |
| GO:0005856 | cytoskeleton | Actb,Actbl2,Actc1,Calm1,Camk2b,Capza2,Capzb,Dynll1, Dynll2,Hsbp1,Hspa8,Ina,Map1b,Map2,Myl6,Nefl,Nefm, Tuba1a,Tuba1c,Tubb4a,Tubb6,Ywhae |
| GO:0099513 | polymeric cytoskeletal fiber | Actc1,Capzb,Dynll1,Dynll2,Hspa8,Ina,Map1b,Map2,Nefl, Nefm, Tuba1a,Tuba1c,Tubb4a,Tubb6 |
| GO:0014069 | postsynaptic density | Actb,Camk2a,Camk2b,Camk2g,Hspa8,Map1b,Map2,Nefm, Rtn4,Ywhaz |
| GO:0043005 | neuron projection | Actb,Atxn10,Calm1,Camk2a,Camk2b,Camk2d,Camk2g, Capzb,Dynll1,Hspa8,Map1b,Map2,Nefl,Nefm,Rtn4,Tubb4a,Ybx1,Ywhae |
| GO:0030424 | axon | Actb,Calm1,Camk2a,Camk2d,Dynll1,Hspa8,Map1b,Map2, Nefl,Nefm,Rtn4,Tubb4a,Ywhae |
| GO:0098794 | postsynapse | Actb,Camk2a,Camk2b,Camk2g,Capzb,Hspa8,Map1b,Map2,Nefm,Rtn4,Ywhaz |
| GO:0005874 | microtubule | Dynll1,Dynll2,Hspa8,Map1b,Map2,Tuba1a,Tuba1c,Tubb4a,Tubb6 |
| GO:0045202 | synapse | Actb,Camk2a,Camk2b,Camk2d,Camk2g,Capzb,Grn,Hspa8,Map1b,Map2,Nefm,Rtn4,Ywhaz |
| GO:0005883 | neurofilament | Ina,Nefl,Nefm |
| GO:0015630 | microtubule cytoskeleton | Calm1,Camk2b,Dynll1,Dynll2,Hspa8,Map1b,Map2,Tuba1a, Tuba1c,Tubb4a,Tubb6,Ywhae |
| GO:0030426 | growth cone | Calm1,Map1b,Map2,Nefl,Rtn4,Ywhae |
| GO:0016529 | sarcoplasmic reticulum | Calu,Camk2b,Camk2d,Camk2g |
| GO:0030425 | dendrite | Atxn10,Camk2a,Camk2b,Capzb,Hspa8,Map1b,Map2,Rtn4, Ybx1 |
| GO:0150034 | distal axon | Calm1,Hspa8,Map1b,Map2,Nefl,Rtn4,Ywhae |
| GO:0015629 | actin cytoskeleton | Actb,Actc1,Capza2,Capzb,Dynll2,Map2,Myl6 |
| GO:0005853 | eukaryotic translation elongation factor 1 complex | Eef1b2,Eef1d |
| GO:0099524 | postsynaptic cytosol | Camk2a,Hspa8 |
| GO:0005882 | intermediate filament | Hspa8,Ina,Nefl,Nefm |
| GO:0008290 | F-actin capping protein complex | Capza2,Capzb |
| GO:0044294 | dendritic growth cone | Map2,Rtn4 |
| GO:0043194 | axon initial segment | Camk2d,Map2 |
| KEGGs Enrichment | ||
|---|---|---|
| KEGG | Term Description | Proteins |
| mmu04722 | Neurotrophin signaling pathway | Calm1,Camk2a,Camk2b,Camk2d,Camk2g,Ywhae |
| mmu04720 | Long-term potentiation | Calm1,Camk2a,Camk2b,Camk2d,Camk2g |
| mmu04012 | ErbB signaling pathway | Camk2a,Camk2b,Camk2d,Camk2g |
| mmu04540 | Gap junction | Tuba1a,Tuba1c,Tubb4a,Tubb6 |
| mmu04020 | Calcium signaling pathway | Calm1,Camk2a,Camk2b,Camk2d,Camk2g |
| mmu04024 | cAMP signaling pathway | Calm1,Camk2a,Camk2b,Camk2d,Camk2g |
| mmu04360 | Axon guidance | Camk2a,Camk2b,Camk2d,Camk2g |
| Reactome pathways enriched | ||
| Reactome | Term Description | Proteins |
| MMU-195258 | RHO GTPase Effectors | Actb,Calm1,Dynll1,Dynll2,Myl6,Tuba1a,Tuba1c,Tubb4a, Tubb6,Ywhae,Ywhag,Ywhaz |
| MMU-442729 | CREB phosphorylation through the activation of CaMKII | Calm1,Camk2a,Camk2b,Camk2d,Camk2g,Nefl |
| MMU-442982 | Ras activation upon Ca2+ influx through NMDA receptor | Calm1,Camk2a,Camk2b,Camk2d,Camk2g,Nefl |
| MMU-199991 | Membrane Trafficking | Actb,Capza2,Capzb,Dynll1,Dynll2,Rab33a,Tuba1a,Tuba1c, Tubb4a,Tubb6,Ywhae,Ywhag,Ywhaz |
| MMU-438066 | Unblocking of NMDA receptors, glutamate binding and activation | Camk2a,Camk2b,Camk2d,Camk2g,Nefl |
| MMU-5576892 | Phase 0-rapid depolarization | Calm1,Camk2a,Camk2b,Camk2d,Camk2g |
| MMU-190828 | Gap junction trafficking | Actb,Tuba1a,Tuba1c,Tubb4a,Tubb6 |
| MMU-190840 | Microtubule-dependent trafficking of connexons from Golgi to the plasma membrane | Tuba1a,Tuba1c,Tubb4a,Tubb6 |
| MMU-399719 | Trafficking of AMPA receptors | Camk2a,Camk2b,Camk2d,Camk2g |
| MMU-5673001 | RAF/MAP kinase cascade | Actb,Calm1,Camk2a,Camk2b,Camk2d,Camk2g,Nefl |
| MMU-1640170 | Cell Cycle | Dynll1,Dynll2,Tuba1a,Tuba1c,Tubb4a,Tubb6,Ywhae, Ywhag,Ywhaz |
| MMU-5620912 | Anchoring of the basal body to the plasma membrane | Dynll1,Tuba1a,Tubb4a,Ywhae,Ywhag |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parcerisas, A.; Ortega-Gascó, A.; Hernaiz-Llorens, M.; Odena, M.A.; Ulloa, F.; de Oliveira, E.; Bosch, M.; Pujadas, L.; Soriano, E. New Partners Identified by Mass Spectrometry Assay Reveal Functions of NCAM2 in Neural Cytoskeleton Organization. Int. J. Mol. Sci. 2021, 22, 7404. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147404
Parcerisas A, Ortega-Gascó A, Hernaiz-Llorens M, Odena MA, Ulloa F, de Oliveira E, Bosch M, Pujadas L, Soriano E. New Partners Identified by Mass Spectrometry Assay Reveal Functions of NCAM2 in Neural Cytoskeleton Organization. International Journal of Molecular Sciences. 2021; 22(14):7404. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147404
Chicago/Turabian StyleParcerisas, Antoni, Alba Ortega-Gascó, Marc Hernaiz-Llorens, Maria Antonia Odena, Fausto Ulloa, Eliandre de Oliveira, Miquel Bosch, Lluís Pujadas, and Eduardo Soriano. 2021. "New Partners Identified by Mass Spectrometry Assay Reveal Functions of NCAM2 in Neural Cytoskeleton Organization" International Journal of Molecular Sciences 22, no. 14: 7404. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147404