
Generating Cloned Goats by Somatic Cell Nuclear Transfer—Molecular Determinants and Application to Transgenics and Biomedicine


Abstract
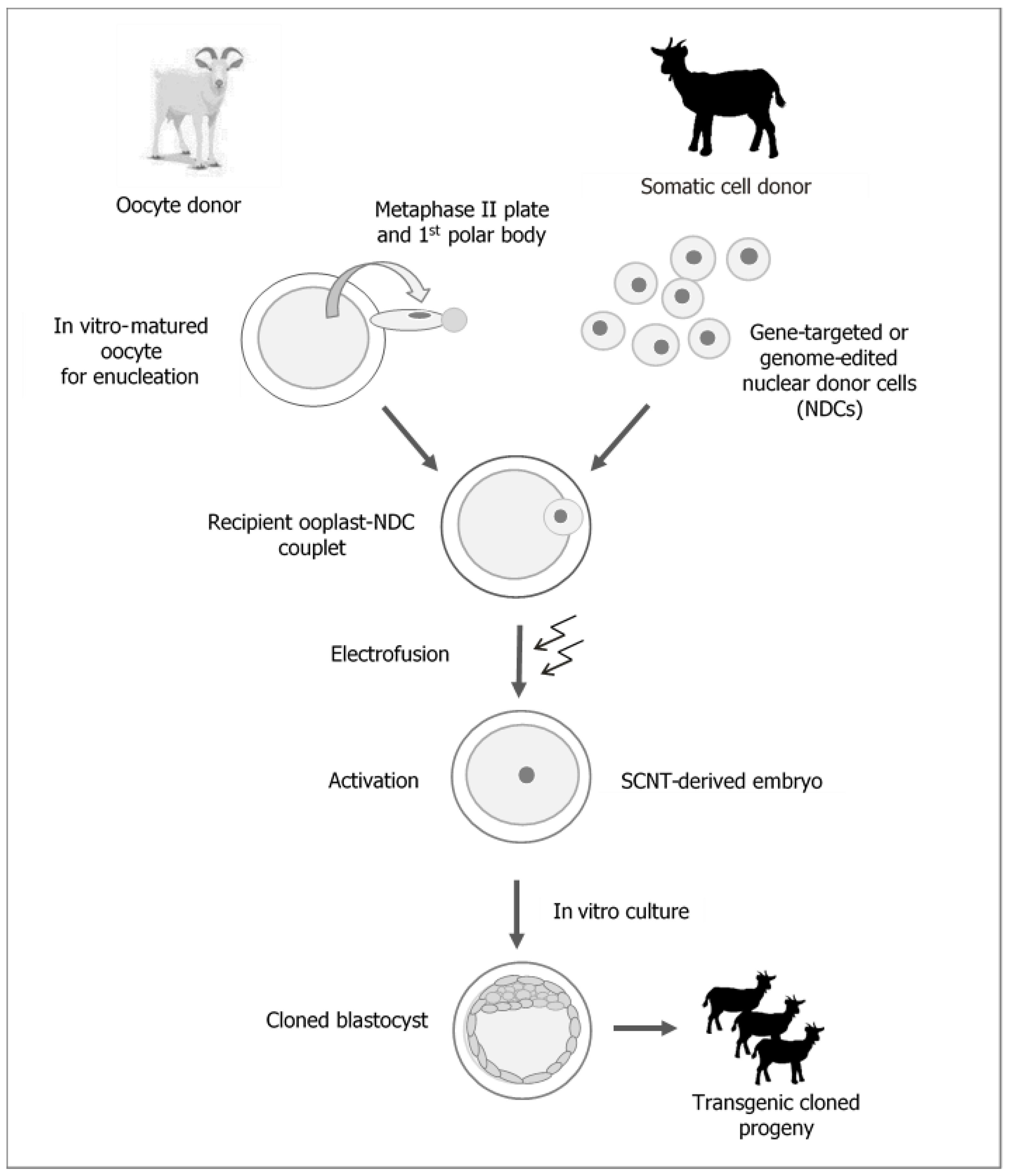
:1. Introduction
2. Key Issues Related to Biological, Molecular, and Epigenetic Determinants Affecting the Efficacy of Somatic Cell Cloning in Goats
3. Species-Specific Advantages of the Goat That Increase the Potential for Its Practical Application in Transgenics, Biopharmacy, Biomedicine, and Biotechnology
4. Transgenic Cloned Goats as Bioreactors That Produce Recombinant Human Therapeutic Proteins
5. Transgenic Cloned Goats as a Source of Valuable Meat for Humans
- –
- Expediting differentiation of the predominant multipotent muscle stem cells (i.e., satellite cells) and their myogenic progenitor cell derivatives into primary myoblasts;
- –
- Accelerating the proliferative growth of mononucleated myoblasts;
- –
- Facilitating cyto- and histophysiological maturation of genetically engineered skeletal muscle tissue by syncytial fusion of myoblasts and their conversion (transformation) into myotubes and the resultant multinucleated myofibers;
- –
- Enlargement of muscle fibers;
- –
- Extension of myofiber lengths and individual sarcomere lengths in whole muscle fibers;
- –
6. Conclusions and Future Goals
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ART | Assisted reproductive technology |
| BLG | β-lactoglobulin |
| Cas9 | CRISPR-associated endonuclease type 9 |
| Cdx2 | A product transcribed from proto-oncogene/oncogene encoding caudal-type homeobox protein 2 that represents the family of intestinal epithelium/adenocarcinoma-specific and DNA-binding homeodomain transcription factors inevitable in intestinal organogenesis |
| c-Myc | Avian myelocytomatosis viral oncogene homolog encoding a DNA-binding |
| proto-oncogenic/oncogenic transcription factor | |
| CpG | 5′-Cytidine-3′-monophopshate-5′-guanosine-3′ |
| CRISPR | Clustered regularly interspaced short palindromic repeat |
| DMR | Differentially methylated region |
| DNMT | DNA methyltransferase |
| eGFP | Enhanced green fluorescent protein |
| GE | Genome-edited |
| GEM | Genetically engineered milk |
| HAT | Histone acetyltransferase |
| HDAC | Histone deacetylase |
| HMT | Histone methyltransferase |
| hLA | Human α-lactalbumin |
| hLF | Human lactoferrin |
| HR | Homologous recombination |
| ICM | Inner cell mass |
| ICR | Imprinting control region |
| ICSI | Intracytoplasmic sperm injection |
| IGF2R | Insulin-like growth factor 2 receptor |
| Klf4 | Krüppel-like factor 4 (also called gut-enriched Krüppel-like factor or GKLF); an evolutionarily conserved zinc finger-containing transcription factor that regulates diverse cellular processes such as cell growth, proliferation, differentiation, apoptosis, and somatic cell reprogramming |
| 5-mC | 5-Methylcytosine |
| MSTN | Myostatin |
| MSTN-KO | Myostatin gene knockout |
| Nanog | Homeobox-containing transcription factor whose name stems from the Celtic/Irish |
| mythical word Tír na nÓg (i.e., Tir Na Nog; The Land of the Ever-Young) | |
| NDC | Nuclear donor cell |
| Oct3/4 | Octamer-binding transcription factor 3/4 (also designated as POU5F1); a member of the family of POU (Pit-Oct-Unc)-domain and homeodomain transcription factors |
| 2-PCPA | Trans-2-phenylcyclopropylamine; Tranylcypromine |
| PGKneo | Neomycin phosphoglycerol kinase; Neomycin glycerol phosphotransferase |
| Rex1 | Reduced expression gene 1 encoding a DNA-binding transcription factor |
| known as reduced expression protein 1 or zinc finger protein 42 homolog | |
| rhAT | Recombinant human antithrombin III |
| rhEPO | Recombinant human erythropoietin |
| SCNT | Somatic cell nuclear transfer |
| Sox2 | Sex-determining region Y (SRY)-box 2; a member of the high mobility group |
| (HMG)-box family of DNA-binding transcription factors | |
| SV-40 | Simian virus-40 |
| TALEN | Transcription activator-like effector nuclease |
| TET3 | Ten-eleven translocation 3 protein; TET 5-methylcytosine dioxygenase 3 |
| Xist | X-inactive specific transcript |
| ZFN | Zinc-finger nuclease |
References
- Samiec, M.; Skrzyszowska, M. Transgenic mammalian species, generated by somatic cell cloning, in biomedicine, biopharmaceutical industry and human nutrition/dietetics—Recent achievements. Pol. J. Vet. Sci. 2011, 14, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Samiec, M.; Skrzyszowska, M. The possibilities of practical application of transgenic mammalian species generated by somatic cell cloning in pharmacology, veterinary medicine and xenotransplantology. Pol. J. Vet. Sci. 2011, 14, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Hall, V.; Hinrichs, K.; Lazzari, G.; Betts, D.H.; Hyttel, P. Early embryonic development, assisted reproductive technologies, and pluripotent stem cell biology in domestic mammals. Vet. J. 2013, 197, 128–142. [Google Scholar] [CrossRef] [Green Version]
- Skrzyszowska, M.; Samiec, M. Generation of monogenetic cattle by different techniques of embryonic cell and somatic cell cloning—Their application to biotechnological, agricultural, nutritional, biomedical and transgenic research—A review. Ann. Anim. Sci. 2021, 21, 1–15. [Google Scholar]
- Martins, L.T.; Neto, S.G.; Tavares, K.C.; Calderón, C.E.; Aguiar, L.H.; Lazzarotto, C.R.; Ongaratto, F.L.; Rodrigues, V.H.; Carneiro Ide, S.; Rossetto, R.; et al. Developmental outcome and related abnormalities in goats: Comparison between somatic cell nuclear transfer- and in vivo-derived concepti during pregnancy through term. Cell. Reprogram. 2016, 18, 264–279. [Google Scholar] [CrossRef] [PubMed]
- Samiec, M.; Skrzyszowska, M. Can reprogramming of overall epigenetic memory and specific parental genomic imprinting memory within donor cell-inherited nuclear genome be a major hindrance for the somatic cell cloning of mammals?—A review. Ann. Anim. Sci. 2018, 18, 623–638. [Google Scholar] [CrossRef] [Green Version]
- Samiec, M.; Skrzyszowska, M. Intrinsic and extrinsic molecular determinants or modulators for epigenetic remodeling and reprogramming of somatic cell-derived genome in mammalian nuclear-transferred oocytes and resultant embryos. Pol. J. Vet. Sci. 2018, 21, 217–227. [Google Scholar] [PubMed]
- Yang, M.; Perisse, I.; Fan, Z.; Regouski, M.; Meyer-Ficca, M.; Polejaeva, I.A. Increased pregnancy losses following serial somatic cell nuclear transfer in goats. Reprod. Fertil. Dev. 2018, 30, 1443–1453. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Liu, Z.; Chen, B.; Wan, Y.; Yang, H.; Zhang, Y.; Cai, Y.; Zhou, J.; Wang, F. Aberrant DNA and histone methylation during zygotic genome activation in goat cloned embryos. Theriogenology 2020, 148, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Zhang, G.; Cai, Y.; Liu, Z.; Zhang, Y.; Meng, F.; Wang, F.; Wan, Y. DNA methylation dynamics during zygotic genome activation in goat. Theriogenology 2020, 156, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Iager, A.E.; Ragina, N.P.; Ross, P.J.; Beyhan, Z.; Cunniff, K.; Rodriguez, R.M.; Cibelli, J.B. Trichostatin A improves histone acetylation in bovine somatic cell nuclear transfer early embryos. Cloning Stem Cells 2008, 10, 371–379. [Google Scholar] [CrossRef]
- Mao, T.; Han, C.; Deng, R.; Wei, B.; Meng, P.; Luo, Y.; Zhang, Y. Treating donor cells with 2-PCPA corrects aberrant histone H3K4 dimethylation and improves cloned goat embryo development. Syst. Biol. Reprod. Med. 2018, 64, 174–182. [Google Scholar] [CrossRef]
- Samiec, M.; Romanek, J.; Lipiński, D.; Opiela, J. Expression of pluripotency-related genes is highly dependent on trichostatin A-assisted epigenomic modulation of porcine mesenchymal stem cells analysed for apoptosis and subsequently used for generating cloned embryos. Anim. Sci. J. 2019, 90, 1127–1141. [Google Scholar] [CrossRef]
- Skrzyszowska, M.; Samiec, M. Enhancement of in vitro developmental outcome of cloned goat embryos after epigenetic modulation of somatic cell-inherited nuclear genome with trichostatin A. Ann. Anim. Sci. 2020, 20, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Wiater, J.; Samiec, M.; Skrzyszowska, M.; Lipiński, D. Trichostatin A-assisted epigenomic modulation affects the expression profiles of not only recombinant human α1,2-fucosyltransferase and α-galactosidase A enzymes but also Galα1→3Gal epitopes in porcine bi-transgenic adult cutaneous fibroblast cells. Int. J. Mol. Sci. 2021, 22, 1386. [Google Scholar] [CrossRef]
- Reggio, B.C.; James, A.N.; Green, H.L.; Gavin, W.G.; Behboodi, E.; Echelard, Y.; Godke, R.A. Cloned transgenic offspring resulting from somatic cell nuclear transfer in the goat: Oocytes derived from both follicle-stimulating hormone-stimulated and nonstimulated abattoir-derived ovaries. Biol. Reprod. 2001, 65, 1528–1533. [Google Scholar] [CrossRef] [Green Version]
- Keefer, C.L.; Baldassarre, H.; Keyston, R.; Wang, B.; Bhatia, B.; Bilodeau, A.S.; Zhou, J.F.; Leduc, M.; Downey, B.R.; Lazaris, A.; et al. Generation of dwarf goat (Capra hircus) clones following nuclear transfer with transfected and nontransfected fetal fibroblasts and in vitro-matured oocytes. Biol. Reprod. 2001, 64, 849–856. [Google Scholar] [CrossRef] [Green Version]
- Keefer, C.L.; Keyston, R.; Lazaris, A.; Bhatia, B.; Begin, I.; Bilodeau, A.S.; Zhou, F.J.; Kafidi, N.; Wang, B.; Baldassarre, H.; et al. Production of cloned goats after nuclear transfer using adult somatic cells. Biol. Reprod. 2002, 66, 199–203. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Wang, Y.; Cheng, Y.; Yang, Y.; Ju, H.; Tang, H.; Shen, Y.; Mu, Z.; Xu, S.; Du, M. Generation of cloned goats (Capra hircus) from transfected foetal fibroblast cells, the effect of donor cell cycle. Mol. Reprod. Dev. 2002, 61, 164–172. [Google Scholar] [CrossRef]
- Baldassarre, H.; Wang, B.; Pierson, J.; Neveu, N.; Lapointe, J.; Cote, F.; Kafidi, N.; Keefer, C.L.; Lazaris, A.; Karatzas, C.N. Prepubertal propagation of transgenic cloned goats by laparoscopic ovum pick-up and in vitro embryo production. Cloning Stem Cells 2004, 6, 25–29. [Google Scholar] [CrossRef]
- Zhu, H.; Hu, L.; Liu, J.; Chen, H.; Cui, C.; Song, Y.; Jin, Y.; Zhang, Y. Generation of β-lactoglobulin-modified transgenic goats by homologous recombination. FEBS J. 2016, 283, 4600–4613. [Google Scholar] [CrossRef]
- Yuan, Y.G.; Song, S.Z.; Zhu, M.M.; He, Z.Y.; Lu, R.; Zhang, T.; Mi, F.; Wang, J.Y.; Cheng, Y. Human lactoferrin efficiently targeted into caprine beta-lactoglobulin locus with transcription activator-like effector nucleases. Asian Australas. J. Anim. Sci. 2017, 30, 1175–1182. [Google Scholar] [CrossRef] [Green Version]
- Behboodi, E.; Memili, E.; Melican, D.T.; Destrempes, M.M.; Overton, S.A.; Williams, J.L.; Flanagan, P.A.; Butler, R.E.; Liem, H.; Chen, L.H.; et al. Viable transgenic goats derived from skin cells. Transgenic Res. 2004, 13, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.J.; Zhang, Y.L.; Zhou, Z.R.; Jia, R.X.; Li, M.; Song, H.; Wang, Z.Y.; Wang, L.Z.; Zhang, G.M.; You, J.H.; et al. Efficiency of donor cell preparation and recipient oocyte source for production of transgenic cloned dairy goats harboring human lactoferrin. Theriogenology 2012, 78, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.R.; Zhong, B.S.; Jia, R.X.; Wan, Y.J.; Zhang, Y.L.; Fan, Y.X.; Wang, L.Z.; You, J.H.; Wang, Z.Y.; Wang, F. Production of myostatin-targeted goat by nuclear transfer from cultured adult somatic cells. Theriogenology 2013, 79, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Sarkhel, B.C. Differential expression pattern of key regulatory developmental genes in pre-implant zona free cloned vs in vitro fertilized goat embryos. Gene Expr. Patterns 2017, 25, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Baldassarre, H.; Wang, B.; Kafidi, N.; Keefer, C.L.; Lazaris, A.; Karatzas, C.N. Advances in the production and propagation of transgenic goats using laparoscopic ovum pick-up and in vitro embryo production technologies. Theriogenology 2002, 57, 275–284. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, Y.G.; Luo, J.P.; Shen, Y.; Yang, Y.F.; Ju, H.M.; Zou, X.G.; Xu, S.F.; Lao, W.D.; Du, M. Cloned goats produced from the somatic cells of an adult transgenic goat. Sheng Wu Gong Cheng Xue Bao (Chin. J. Biotechnol.) 2002, 18, 79–83. (In Chinese) [Google Scholar]
- Ohkoshi, K.; Takahashi, S.; Koyama, S.; Akagi, S.; Adachi, N.; Furusawa, T.; Fujimoto, J.; Takeda, K.; Kubo, M.; Izaike, Y.; et al. In vitro oocyte culture and somatic cell nuclear transfer used to produce a live-born cloned goat. Cloning Stem Cells 2003, 5, 109–115. [Google Scholar] [CrossRef]
- Deng, M.; Liu, Z.; Ren, C.; An, S.; Wan, Y.; Wang, F. Highly methylated Xist in SCNT embryos was retained in deceased cloned female goats. Reprod. Fertil. Dev. 2019, 31, 855–866. [Google Scholar] [CrossRef]
- Wang, F.; Kou, Z.; Zhang, Y.; Gao, S. Dynamic reprogramming of histone acetylation and methylation in the first cell cycle of cloned mouse embryos. Biol. Reprod. 2007, 77, 1007–1016. [Google Scholar] [CrossRef]
- Deng, M.; Ren, C.; Liu, Z.; Zhang, G.; Wang, F.; Wan, Y. Epigenetic status of H19-Igf2 imprinted genes and loss of 5-hydroxymethylcytosine in the brain of cloned goats. Cell. Reprogram. 2017, 19, 199–207. [Google Scholar] [CrossRef]
- Han, C.; Deng, R.; Mao, T.; Luo, Y.; Wei, B.; Meng, P.; Zhao, L.; Zhang, Q.; Quan, F.; Liu, J.; et al. Overexpression of Tet3 in donor cells enhances goat somatic cell nuclear transfer efficiency. FEBS J. 2018, 285, 2708–2723. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Hao, R.; Kessler, B.; Brem, G.; Wolf, E.; Zakhartchenko, V. Rabbit somatic cell cloning: Effects of donor cell type, histone acetylation status and chimeric embryo complementation. Reproduction 2007, 133, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, K.; Sugimura, S.; Wakai, T.; Kawahara, M.; Sato, E. Acetylation level of histone H3 in early embryonic stages affects subsequent development of miniature pig somatic cell nuclear transfer embryos. J. Reprod. Dev. 2009, 55, 638–644. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Deng, M.; Zhang, G.; Ren, C.; Zhang, H.; Zhang, Y.; Wang, L.; Wang, F. Abnormal expression of DNA methyltransferases and genomic imprinting in cloned goat fibroblasts. Cell Biol. Int. 2016, 40, 74–82. [Google Scholar] [CrossRef]
- Xiong, X.; Lan, D.; Li, J.; Zhong, J.; Zi, X.; Ma, L.; Wang, Y. Zebularine and scriptaid significantly improve epigenetic reprogramming of yak fibroblasts and cloning efficiency. Cell. Reprogram. 2013, 15, 293–300. [Google Scholar] [CrossRef]
- Xiong, X.R.; Li, J.; Fu, M.; Gao, C.; Wang, Y.; Zhong, J.C. Oocyte extract improves epigenetic reprogramming of yak fibroblast cells and cloned embryo development. Theriogenology 2013, 79, 462–469. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, J.; Villemoes, K.; Pedersen, A.M.; Purup, S.; Vajta, G. An epigenetic modifier results in improved in vitro blastocyst production after somatic cell nuclear transfer. Cloning Stem Cells 2007, 9, 357–363. [Google Scholar] [CrossRef]
- Wang, Y.S.; Xiong, X.R.; An, Z.X.; Wang, L.J.; Liu, J.; Quan, F.S.; Hua, S.; Zhang, Y. Production of cloned calves by combination treatment of both donor cells and early cloned embryos with 5-aza-2’-deoxycytidine and trichostatin A. Theriogenology 2011, 75, 819–825. [Google Scholar] [CrossRef]
- Dutta, R.; Malakar, D.; Khate, K.; Sahu, S.; Akshey, Y.; Mukesh, M. A comparative study on efficiency of adult fibroblast, putative embryonic stem cell and lymphocyte as donor cells for production of handmade cloned embryos in goat and characterization of putative ntES cells obtained from these embryos. Theriogenology 2011, 76, 851–863. [Google Scholar] [CrossRef]
- Gómez, M.C.; Biancardi, M.N.; Jenkins, J.A.; Dumas, C.; Galiguis, J.; Wang, G.; Earle Pope, C. Scriptaid and 5-aza-2’deoxycytidine enhanced expression of pluripotent genes and in vitro developmental competence in interspecies black-footed cat cloned embryos. Reprod. Domest. Anim. 2012, 47, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Hao, Y.; Ross, J.W.; Spate, L.D.; Walters, E.M.; Samuel, M.S.; Rieke, A.; Murphy, C.N.; Prather, R.S. Histone deacetylase inhibitors improve in vitro and in vivo developmental competence of somatic cell nuclear transfer porcine embryos. Cell. Reprogram. 2010, 12, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Ahn, K.S.; Kim, M.; Shim, H. Comparison of potency between histone deacetylase inhibitors trichostatin A and valproic acid on enhancing in vitro development of porcine somatic cell nuclear transfer embryos. In Vitro Cell. Dev. Biol. Anim. 2011, 47, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Baguisi, A.; Behboodi, E.; Melican, D.T.; Pollock, J.S.; Destrempes, M.M.; Cammuso, C.; Williams, J.L.; Nims, S.D.; Porter, C.A.; Midura, P.; et al. Production of goats by somatic cell nuclear transfer. Nat. Biotechnol. 1999, 17, 456–461. [Google Scholar] [CrossRef]
- Baldassarre, H.; Keefer, C.; Wang, B.; Lazaris, A.; Karatzas, C.N. Nuclear transfer in goats using in vitro matured oocytes recovered by laparoscopic ovum pick-up. Cloning Stem Cells 2003, 5, 279–285. [Google Scholar] [CrossRef] [PubMed]
- An, L.; Yang, L.; Huang, Y.; Cheng, Y.; Du, F. Generating goat mammary gland bioreactors for producing recombinant proteins by gene targeting. Methods Mol. Biol. 2019, 1874, 391–401. [Google Scholar] [PubMed]
- Lu, R.; Zhang, T.; Wu, D.; He, Z.; Jiang, L.; Zhou, M.; Cheng, Y. Production of functional human CuZn-SOD and EC-SOD in bitransgenic cloned goat milk. Transgenic Res. 2018, 27, 343–354. [Google Scholar] [CrossRef]
- Gash, K.K.; Yang, M.; Fan, Z.; Regouski, M.; Rutigliano, H.M.; Polejaeva, I.A. Assessment of microchimerism following somatic cell nuclear transfer and natural pregnancies in goats. J. Anim. Sci. 2019, 97, 3786–3794. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Zhang, G.M.; Wan, Y.J.; Jia, R.X.; Li, P.Z.; Han, L.; Wang, F.; Huang, M.R. Identification of transgenic cloned dairy goats harboring human lactoferrin and methylation status of the imprinted gene IGF2R in their lungs. Genet. Mol. Res. 2015, 14, 11099–11108. [Google Scholar] [CrossRef]
- Meng, L.; Wan, Y.; Sun, Y.; Zhang, Y.; Wang, Z.; Song, Y.; Wang, F. Generation of five human lactoferrin transgenic cloned goats using fibroblast cells and their methylation status of putative differential methylation regions of IGF2R and H19 imprinted genes. PLoS ONE 2013, 8, e77798. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, J.; Yang, W.; Cui, M.; Dai, B.; Dong, Y.; Yang, J.; Zhang, X.; Liu, D.; Liang, H.; et al. Comparison of gene editing efficiencies of CRISPR/Cas9 and TALEN for generation of MSTN knock-out cashmere goats. Theriogenology 2019, 132, 1–11. [Google Scholar] [CrossRef]
- Ni, W.; Qiao, J.; Hu, S.; Zhao, X.; Regouski, M.; Yang, M.; Polejaeva, I.A.; Chen, C. Efficient gene knockout in goats using CRISPR/Cas9 system. PLoS ONE 2014, 9, e106718. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Lu, R.; Yuan, Y.; Zhang, T.; Song, S.; Qi, Z.; Shao, B.; Zhu, M.; Mi, F.; Cheng, Y. Efficient TALEN-mediated myostatin gene editing in goats. BMC Dev. Biol. 2016, 16, 26. [Google Scholar] [CrossRef] [Green Version]
- Baldassarre, H.; Wang, B.; Kafidi, N.; Gauthier, M.; Neveu, N.; Lapointe, J.; Sneek, L.; Leduc, M.; Duguay, F.; Zhou, J.F.; et al. Production of transgenic goats by pronuclear microinjection of in vitro produced zygotes derived from oocytes recovered by laparoscopy. Theriogenology 2003, 59, 831–839. [Google Scholar] [CrossRef]
- Zhang, T.; Yuan, Y.; Lu, R.; Xu, S.; Zhou, M.; Yuan, T.; Lu, Y.; Yan, K.; Cheng, Y. The goat β-casein/CMV chimeric promoter drives the expression of hLF in transgenic goats produced by cell transgene microinjection. Int. J. Mol. Med. 2019, 44, 2057–2064. [Google Scholar] [CrossRef]
- Echelard, Y.; Ziomek, C.A.; Meade, H.M. Production of recombinant therapeutic proteins in the milk of transgenic animals. BioPharm Int. 2006, 19, 36–46. [Google Scholar]
- Niemann, H.; Kues, W.A. Transgenic farm animals: An update. Reprod. Fertil. Dev. 2007, 19, 762–770. [Google Scholar] [CrossRef]
- Shepelev, M.V.; Kalinichenko, S.V.; Deykin, A.V.; Korobko, I.V. Production of recombinant proteins in the milk of transgenic animals: Current state and prospects. Acta Nat. 2018, 10, 40–47. [Google Scholar] [CrossRef]
- Echelard, Y.; Meade, H.M.; Ziomek, C.A. The first biopharmaceutical from transgenic animals: ATryn®. In Modern Biopharmaceuticals: Design, Development and Optimization; Knäblein, J., Ed.; WILEY-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2005; Chapter 11; pp. 995–1020, Print ISBN: 9783527311842, Online ISBN: 9783527620982; ISBN 9783527311842. [Google Scholar] [CrossRef]
- Patnaik, M.M.; Moll, S. Inherited antithrombin deficiency: A review. Haemophilia 2008, 14, 1229–1239. [Google Scholar] [CrossRef]
- Wang, X.; Yu, H.; Lei, A.; Zhou, J.; Zeng, W.; Zhu, H.; Dong, Z.; Niu, Y.; Shi, B.; Cai, B.; et al. Generation of gene-modified goats targeting MSTN and FGF5 via zygote injection of CRISPR/Cas9 system. Sci. Rep. 2015, 5, 13878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Li, W.; Zhang, X.; Zhang, N.; He, S.; Huang, J.; Ge, Y.; Liu, M. Knockdown of endogenous myostatin promotes sheep myoblast proliferation. In Vitro Cell. Dev. Biol. Anim. 2014, 50, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Petersen, B.; Niemann, H. Molecular scissors and their application in genetically modified farm animals. Transgenic Res. 2015, 24, 381–396. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, L.; Ren, G.; Li, Z.; Ren, C.; Zhang, T.; Xu, K.; Zhang, Z. Targeted disruption of the sheep MSTN gene by engineered zinc-finger nucleases. Mol. Biol. Rep. 2014, 41, 209–215. [Google Scholar] [CrossRef]
- Miller, J.C.; Tan, S.; Qiao, G.; Barlow, K.A.; Wang, J.; Xia, D.F.; Meng, X.; Paschon, D.E.; Leung, E.; Hinkley, S.J.; et al. A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. 2011, 29, 143–148. [Google Scholar] [CrossRef]
- Frock, R.L.; Hu, J.; Meyers, R.M.; Ho, Y.J.; Kii, E.; Alt, F.W. Genome-wide detection of DNA double-stranded breaks induced by engineered nucleases. Nat. Biotechnol. 2015, 33, 179–186. [Google Scholar] [CrossRef]
- Hu, J.H.; Davis, K.M.; Liu, D.R. Chemical biology approaches to genome editing: Understanding, controlling, and delivering programmable nucleases. Cell Chem. Biol. 2016, 23, 57–73. [Google Scholar] [CrossRef] [Green Version]
- Hao, F.; Yan, W.; Li, X.; Wang, H.; Wang, Y.; Hu, X.; Liu, X.; Liang, H.; Liu, D. Generation of cashmere goats carrying an EDAR gene mutant using CRISPR-Cas9-mediated genome editing. Int. J. Biol. Sci. 2018, 14, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Fan, Z.; Yang, M.; Regouski, M.; Polejaeva, I.A. Gene knockouts in goats using CRISPR/Cas9 system and somatic cell nuclear transfer. Methods Mol. Biol. 2019, 1874, 373–390. [Google Scholar]
- Kalds, P.; Zhou, S.; Cai, B.; Liu, J.; Wang, Y.; Petersen, B.; Sonstegard, T.; Wang, X.; Chen, Y. Sheep and goat genome engineering: From random transgenesis to the CRISPR era. Front. Genet. 2019, 10, 750. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Target of Genetic Modification | Strategy of Genetic Modification | Method of Somatic Cell Transfection | Genotypic Effect of Genetic Modification | Phenotypic Effect of Genetic Modification | Reference |
|---|---|---|---|---|---|
| Mammary gland (udder) | Gene targeting (HR-mediated targeted mutagenesis) | Electroporation | - HR-induced disruption of caprine the BLG gene by:
| - Functionally inactivating the caprine BLG gene - Targeted expression of the hLA gene in the lactogenic cells of udder-based bioreactors - Synthesis of upgraded or humanized milk without allergenic (BLG-triggered) properties | [21] |
| Lipofection | - HR-dependent targeted incorporation of hLF cDNA into the nuclear genome (under the control of the goat β-casein gene promoter) | - Mammary gland-specific monoallelic expression of the hLF gene - Udder-mediated synthesis of upgraded or humanized milk - Production of genetically engineered milk (GEM) characterized by a broad spectrum of hLF-induced immunotherapeutic properties - GEM properties determined by antimicrobial, immunomodulatory, anti-inflammatory, and anticancer attributes of hLF | [50,51] | ||
| TALEN-mediated gene editing | Electroporation | - TALEN-dependent targeted insertion of hLF cDNA at BLG locus resulting in:
| -Targeted expression of the hLF gene in udder-based bioreactors - Synthesis of hLF-enriched or humanized milk - Production of GEM displaying immunotherapeutic properties | [22] | |
| Skeletal muscles | Gene targeting (HR-mediated targeted mutagenesis) | Lipofection | - Monoallelic knockout (semi-deficiency) of the MSTN gene in SCNT-derived progeny | - Inducing hyperplasia and hypertrophy of striated muscle cells -Remarkably gaining skeletal muscle mass and augmenting meatiness by genetically transforming the muscular system of heterozygous (MSTN+/−) transgenic cloned offspring | [25] |
| CRISPR/Cas9-mediated gene editing | Electroporation | - Monoallelic knockout (semi-deficiency) of the MSTN gene in SCNT-derived progeny | - Expression of cellular hyperplasia and hypertrophy in genome-edited (GE) skeletal muscle tissue of heterozygous (MSTN+/−) transgenic cloned offspring | [52] | |
| Nucleofection | - Biallelic knockout (deficiency) of the MSTN gene in SCNT-derived progeny | - Expression of myofiber hyperplasia and hypertrophy in GE muscular system of homozygous (MSTN−/−) transgenic cloned offspring | [53] | ||
| TALEN-mediated gene editing | Electroporation | - Monoallelically knocking out the MSTN gene - Biallelically knocking out the MSTN gene in SCNT-derived progeny | - Triggering hyperplasia and hypertrophy of skeletal myocytes in the GE muscular system of:
| [54] | |
| Electroporation | - Monoallelic knockout of the MSTN gene (MSTN+/−) in NDCs - Biallelic knockout of the MSTN gene (MSTN−/−) in NDCs | - Onset of the mono- or biallelically transcriptionally silencing MSTN gene in isozygous GE NDCs - Failure in the generation of GE cloned progeny exhibiting phenotypes determined by MSTN mono- or biallelic deletion | [52] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skrzyszowska, M.; Samiec, M. Generating Cloned Goats by Somatic Cell Nuclear Transfer—Molecular Determinants and Application to Transgenics and Biomedicine. Int. J. Mol. Sci. 2021, 22, 7490. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147490
Skrzyszowska M, Samiec M. Generating Cloned Goats by Somatic Cell Nuclear Transfer—Molecular Determinants and Application to Transgenics and Biomedicine. International Journal of Molecular Sciences. 2021; 22(14):7490. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147490
Chicago/Turabian StyleSkrzyszowska, Maria, and Marcin Samiec. 2021. "Generating Cloned Goats by Somatic Cell Nuclear Transfer—Molecular Determinants and Application to Transgenics and Biomedicine" International Journal of Molecular Sciences 22, no. 14: 7490. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147490