
Genome-Wide Identification of GRAS Gene Family and Their Responses to Abiotic Stress in Medicago sativa

 and
and
Abstract
:1. Introduction
2. Results
2.1. Identification and Multiple Sequence Analysis of GRAS Genes in Alfalfa
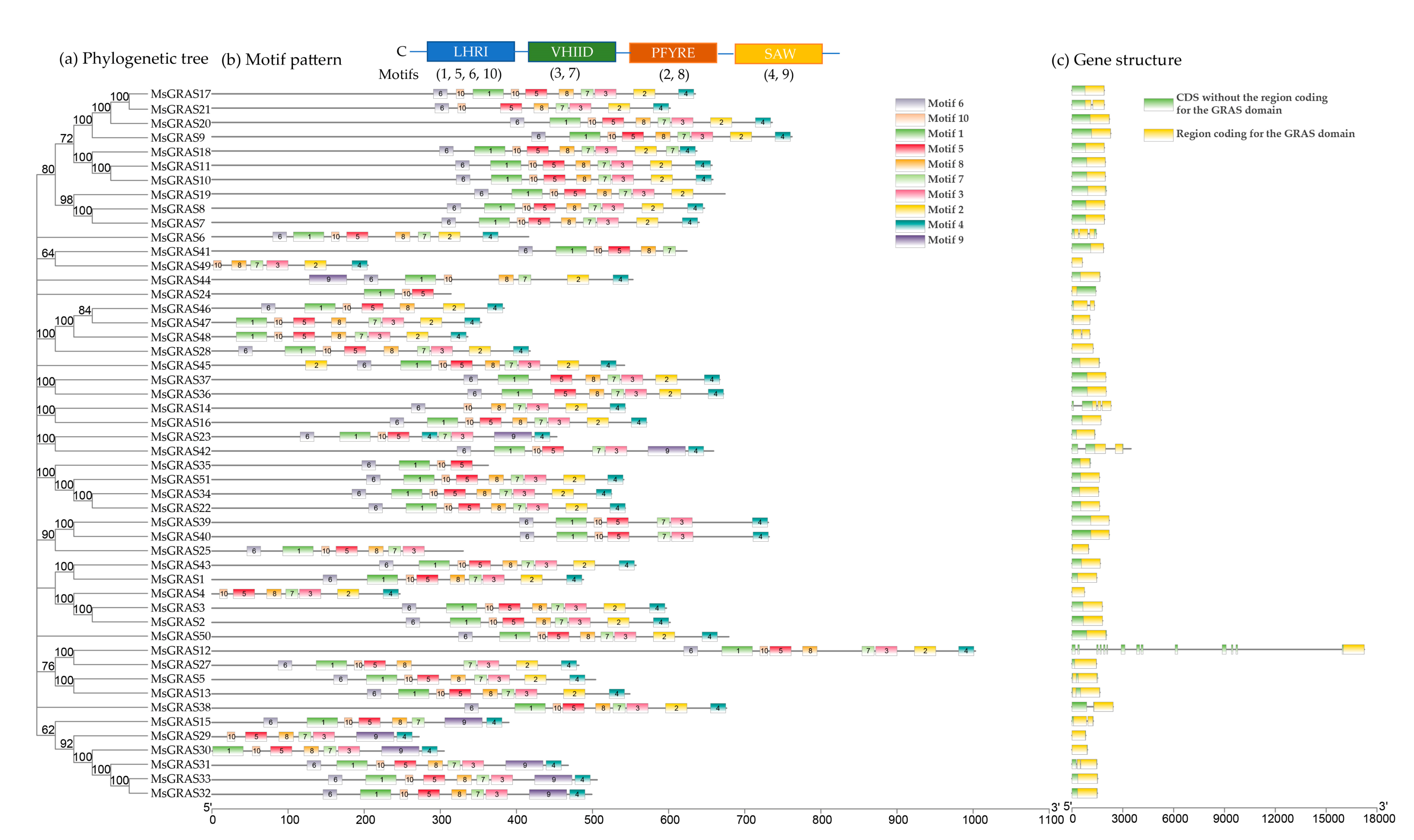
2.2. Phylogenetic Analysis and Classification of GRAS Members
2.3. Transcriptome Analysis of the Alfalfa Leaves under Abiotic Stress
2.4. Validation of Expression Profiles of MsGRAS Genes under Abiotic Stress and Hormone Treatments
3. Discussion
3.1. Duplication of MsGRAS Genes
3.2. Expression and Function of MsGRAS Genes
3.3. Possible Functions of MsGRAS Members
4. Materials and Methods
4.1. Plant Materials
4.2. RNA Extraction and Gene Expression Analysis
4.3. Identification of GRAS Genes in M. sativa
4.3.1. Chromosomal Distribution and Gene Duplication Events Analysis
4.3.2. Sequence Analyses and Structural Characterization of MsGRAS Genes
4.4. Phylogenetic and Collinearity Analysis of MsGRAS Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Riano-Pachon, D.M.; Ruzicic, S.; Dreyer, I.; Mueller-Roeber, B. PlnTFDB: An integrative plant transcription factor database. BMC Bioinform. 2007, 8, 42. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, S.; Oldroyd, G.E. GRAS-domain transcription factors that regulate plant development. Plant Signal. Behav. 2009, 4, 698–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Laurenzio, L.; Wysocka-Diller, J.; Malamy, J.E.; Pysh, L.; Helariutta, Y.; Freshour, G.; Hahn, M.G.; Feldman, K.A. The SCARECROW gene regulates an asymmetric cell division that is essential for generating the radial organization of the Arabidopsis root. Cell 1996, 86, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Silverstone, A.L.; Ciampaglio, C.N.; Sun, T.P. The Arabidopsis RGA gene encodes a transcriptional regulator repressing the gibberellin signal transduction pathway. Plant Cell 1998, 10, 156–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pysh, L.D.; Wysocka-Diller, J.W.; Camilleri, C.; Bouchez, D.; Benfey, P.N. The GRAS gene family in Arabidopsis: Sequence characterization and basic expression analysis of the SCARECROW-LIKE genes. Plant J. 1999, 18, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Bolle, C. The role of GRAS proteins in plant signal transduction and development. Planta 2004, 218, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Jones, W.T.; Rikkerink, E.H. GRAS proteins: The versatile roles of intrinsically disordered proteins in plant signalling. Biochem. J. 2012, 442, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-H.; Kim, B.; Song, S.-K.; Heo, J.-O.; Yu, N.-I.; Lee, S.A.; Kim, M.; Kim, D.G.; Sohn, S.O.; Lim, C.E. Large-scale analysis of the GRAS gene family in Arabidopsis thaliana. Plant Mol. Biol. 2008, 67, 659–670. [Google Scholar] [CrossRef] [Green Version]
- Tian, C.G.; Wan, P.; Sun, S.H.; Li, J.Y.; Chen, M.S. Genome-wide analysis of the GRAS gene family in rice and Arabidopsis. Plant Mol. Biol. 2004, 54, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ding, X.; Gao, Y.; Yang, S. Genome-wide identification and characterization of GRAS genes in soybean (Glycine max). BMC Plant Biol. 2020, 20, 415. [Google Scholar] [CrossRef]
- Huang, W.; Xian, Z.; Kang, X.; Tang, N.; Li, Z. Genome-wide identification, phylogeny and expression analysis of GRAS gene family in tomato. BMC Plant Biol. 2015, 15, 209. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Sun, Y.; Xue, J.; Jia, X.; Li, R. Genome-wide characterization and expression analysis of GRAS gene family in pepper (Capsicum annuum L.). PeerJ 2018, 6, e4796. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Cao, Y.; Shang, C.; Li, J.; Wang, J.; Wu, Z.; Ma, L.; Qi, T.; Fu, C.; Bai, Z.; et al. Genome-wide characterization of GRAS family genes in Medicago truncatula reveals their evolutionary dynamics and functional diversification. PLoS ONE 2017, 12, e0185439. [Google Scholar] [CrossRef]
- Wang, Y.X.; Liu, Z.W.; Wu, Z.J.; Li, H.; Wang, W.L.; Cui, X.; Zhuang, J. Genome-wide identification and expression analysis of GRAS family transcription factors in tea plant (Camellia sinensis). Sci. Rep. 2018, 8, 3949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Huang, L.; Ma, Z.; Sun, W.; Wu, Q.; Tang, Z.; Bu, T.; Li, C.; Chen, H. Genome-wide identification, expression analysis and functional study of the GRAS gene family in Tartary buckwheat (Fagopyrum tataricum). BMC Plant Biol. 2019, 19, 342. [Google Scholar] [CrossRef] [Green Version]
- Bolle, C.; Koncz, C.; Chua, N.H. PAT1, a new member of the GRAS family, is involved in phytochrome a signal transduction. Genes Dev. 2000, 14, 1269–1278. [Google Scholar]
- Song, X.M.; Liu, T.K.; Duan, W.K.; Ma, Q.H.; Ren, J.; Wang, Z.; Li, Y.; Hou, X.L. Genome-wide analysis of the GRAS gene family in Chinese cabbage (Brassica rapa ssp. pekinensis). Genomics 2014, 103, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Liu, J.; Yang, Z.E.; Chen, E.Y.; Zhang, C.J.; Zhang, X.Y.; Li, F.G. Genome-wide analysis of GRAS transcription factor gene family in Gossypium hirsutum L. BMC Genom. 2018, 19, 348. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, N.S.; Pruthi, G.; Singh, S.; Bishnoi, R.; Singla, D. Genome-wide identification and analysis of GRAS transcription factors in the bottle gourd genome. Sci. Rep. 2020, 10, 14338. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Ueguchi-Tanaka, M. DELLA and SCL3 balance gibberellin feedback regulation by utilizing indeterminate domain proteins as transcriptional scaffolds. Plant Signal. Behav. 2014, 9, e29726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.C.; Cheng, H.; King, K.E.; Wang, W.F.; He, Y.W.; Hussain, A.; Lo, J.; Harberd, N.P.; Peng, J.R. Gibberellin regulates Arabidopsis seed germination via RGL2, a GAI/RGA-like gene whose expression is up-regulated following imbibition. Genes Dev. 2002, 16, 646–658. [Google Scholar] [CrossRef] [Green Version]
- Wen, C.K.; Chang, C. Arabidopsis RGL1 encodes a negative regulator of gibberellin responses. Plant Cell 2002, 14, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Carol, P.; Richards, D.E.; King, K.E.; Cowling, R.J.; Murphy, G.P.; Harberd, N.P. The Arabidopsis GAI gene defines a signaling pathway that negatively regulates gibberellin responses. Genes Dev. 1997, 11, 3194–3205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greb, T.; Clarenz, O.; Schafer, E.; Muller, D.; Herrero, R.; Schmitz, G.; Theres, K. Molecular analysis of the lateral suppressor gene in Arabidopsis reveals a conserved control mechanism for axillary meristem formation. Genes Dev. 2003, 17, 1175–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Wang, X.; Li, C.; Li, H.; Zhang, J.; Ye, Z. Silencing GRAS2 reduces fruit weight in tomato. J. Integr. Plant Biol. 2018, 60, 498–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuurman, J.; Jaggi, F.; Kuhlemeier, C. Shoot meristem maintenance is controlled by a GRAS-gene mediated signal from differentiating cells. Genes Dev. 2002, 16, 2213–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Zhu, M.; Ren, L.; Li, A.; Chen, G.; Hu, Z. The SlFSR gene controls fruit shelf-life in tomato. J. Exp. Bot. 2018, 69, 2897–2909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Peng, S.; Xian, Z.; Lin, D.; Hu, G.; Yang, L.; Ren, M.; Li, Z. Overexpression of a tomato miR171 target gene SlGRAS24 impacts multiple agronomical traits via regulating gibberellin and auxin homeostasis. Plant Biotechnol. J. 2017, 15, 472–488. [Google Scholar] [CrossRef]
- Ma, H.S.; Liang, D.; Shuai, P.; Xia, X.L.; Yin, W.L. The salt- and drought-inducible poplar GRAS protein SCL7 confers salt and drought tolerance in Arabidopsis thaliana. J. Exp. Bot. 2010, 61, 4011–4019. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Zhang, B.; Su, T.; Li, P.; Xin, X.; Wang, W.; Zhao, X.; Yu, Y.; Zhang, D.; Yu, S.; et al. BrLAS, a GRAS transcription factor from Brassica rapa, is involved in drought stress tolerance in transgenic Arabidopsis. Front. Plant Sci. 2018, 9, 1792. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zeng, Y.; Yang, Y.; Huang, L.; Tang, B.; Zhang, H.; Hao, F.; Liu, W.; Li, Y.; Liu, Y.; et al. Allele-aware chromosome-level genome assembly and efficient transgene-free genome editing for the autotetraploid cultivated alfalfa. Nat. Commun. 2020, 11, 2494. [Google Scholar] [CrossRef]
- Li, A.; Liu, A.; Du, X.; Chen, J.-Y.; Yin, M.; Hu, H.-Y.; Shrestha, N.; Wu, S.-D.; Wang, H.-Q.; Dou, Q.-W. A chromosome-scale genome assembly of a diploid alfalfa, the progenitor of autotetraploid alfalfa. Hortic. Res. 2020, 7, 1–12. [Google Scholar] [CrossRef]
- Shen, C.; Du, H.; Chen, Z.; Lu, H.; Zhu, F.; Chen, H.; Meng, X.; Liu, Q.; Liu, P.; Zheng, L.; et al. The chromosome-level genome sequence of the autotetraploid alfalfa and resequencing of core germplasms provide genomic resources for alfalfa research. Mol. Plant 2020, 13, 1250–1261. [Google Scholar] [CrossRef]
- Sudhir, K.; Glen, S.; Li, M.; Christina, K.; Koichiro, T. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar]
- Gruber, M.Y.; Xia, J.; Yu, M.; Steppuhn, H.; Wall, K.; Messer, D.; Sharpe, A.G.; Acharya, S.N.; Wishart, D.S.; Johnson, D.; et al. Transcript analysis in two alfalfa salt tolerance selected breeding populations relative to a non-tolerant population. Genome 2017, 60, 104–127. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Liu, W.; Xiang, C.; Li, X.; Wang, Q.; Wang, T.; Liu, Z.; Zhang, J.; Gao, L.; Zhang, W. Genome-wide characterization of GRAS family and their potential roles in cold tolerance of cucumber (Cucumis sativus L.). Int. J. Mol. Sci. 2020, 21, 3857. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, X.; Zhang, H.; Ma, L.; Zhao, H.; Jones, C.S.; Chen, J.; Liu, G. A genome-wide association study approach to the identification of candidate genes underlying agronomic traits in alfalfa (Medicago sativa L.). Plant Biotechnol. J. 2020, 18, 611–613. [Google Scholar] [CrossRef]
- Li, X.; Brummer, E.C. Applied genetics and genomics in alfalfa breeding. Agronomy 2012, 2, 40–61. [Google Scholar] [CrossRef] [Green Version]
- Gao, T.W.; Zhang, W.W.; Song, A.P.; An, C.; Xin, J.J.; Jiang, J.F.; Guan, Z.Y.; Chen, F.D.; Chen, S.M. Phylogenetic and transcriptional analysis of chrysanthemum GRAS transcription factors. Biol. Plant. 2018, 62, 711–720. [Google Scholar] [CrossRef]
- Lu, J.; Wang, T.; Xu, Z.; Sun, L.; Zhang, Q. Genome-wide analysis of the GRAS gene family in Prunus mume. Mol. Genet. Genom. 2015, 290, 303–317. [Google Scholar] [CrossRef]
- Xu, W.; Chen, Z.; Ahmed, N.; Han, B.; Cui, Q.; Liu, A. Genome-wide identification, evolutionary analysis, and stress responses of the GRAS gene family in castor beans. Int. J. Mol. Sci. 2016, 17, 1004. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Widmer, A. Genome-wide comparative analysis of the GRAS gene family in Populus, Arabidopsis and Rice. Plant Mol. Biol. Report. 2014, 32, 1129–1145. [Google Scholar] [CrossRef]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, H.; Wang, X.; Bowers, J.E.; Ming, R.; Alam, M.; Paterson, A.H. Unraveling ancient hexaploidy through multiply-aligned angiosperm gene maps. Genome Res. 2008, 18, 1944–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, M.; Guo, B.; He, S. The roles and evolutionary patterns of intronless genes in deuterostomes. Comp. Funct. Genom. 2011, 2011, 8. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Nijhawan, A.; Arora, R.; Agarwal, P.; Ray, S.; Sharma, P.; Kapoor, S.; Tyagi, A.K.; Khurana, J.P. F-box proteins in rice. Genome-wide analysis, classification, temporal and spatial gene expression during panicle and seed development, and regulation by light and abiotic stress. Plant Physiol. 2007, 143, 1467–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, M.; Tyagi, A.K.; Khurana, J.P. Genome-wide analysis, evolutionary expansion, and expression of early auxin-responsive SAUR gene family in rice (Oryza sativa). Genomics 2006, 88, 360–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aubourg, S.; Kreis, M.; Lecharny, A. The DEAD box RNA helicase family in Arabidopsis thaliana. Nucleic Acids Res. 1999, 27, 628–636. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Iyer, L.M.; Aravind, L. Bacterial GRAS domain proteins throw new light on gibberellic acid response mechanisms. Bioinformatics 2012, 28, 2407–2411. [Google Scholar] [CrossRef] [Green Version]
- Li, M.Y.; Xu, Z.S.; Tian, C.; Huang, Y.; Wang, F.; Xiong, A.S. Genomic identification of WRKY transcription factors in carrot (Daucus carota) and analysis of evolution and homologous groups for plants. Sci. Rep. 2016, 6, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dill, A.; Jung, H.S.; Sun, T.P. The DELLA motif is essential for gibberellin-induced degradation of RGA. Proc. Natl. Acad. Sci. USA 2001, 98, 14162–14167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, K.E.; Moritz, T.; Harberd, N.P. Gibberellins are not required for normal stem growth in Arabidopsis thaliana in the absence of GAI and RGA. Genetics 2001, 159, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Harberd, N.P. Relieving DELLA restraint. Science 2003, 299, 1853–1854. [Google Scholar] [CrossRef]
- Achard, P.; Vriezen, W.H.; Van Der Straeten, D.; Harberd, N.P. Ethylene regulates Arabidopsis development via the modulation of DELLA protein growth repressor function. Plant Cell 2003, 15, 2816–2825. [Google Scholar] [CrossRef] [Green Version]
- Aoyanagi, T.; Ikeya, S.; Kobayashi, A.; Kozaki, A. Gene regulation via the combination of transcription factors in the indeterminate domain and GRAS families. Genes 2020, 11, 613. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.O.; Chang, K.S.; Kim, I.A.; Lee, M.H.; Lee, S.A.; Song, S.K.; Lee, M.M.; Lim, J. Funneling of gibberellin signaling by the GRAS transcription regulator scarecrow-like 3 in the Arabidopsis root. Proc. Natl. Acad. Sci. USA 2011, 108, 2166–2171. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.H.; Shang, F.; Lin, Q.T.; Lou, C.; Zhang, J. Tillering and panicle branching genes in rice. Gene 2014, 537, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, K.; Schmitt, T.; Rossberg, M.; Schmitz, G.; Theres, K. The Lateral suppressor (Ls) gene of tomato encodes a new member of the VHIID protein family. Proc. Natl. Acad. Sci. USA 1999, 96, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, P.; Jha, B. Transcription factors in plants and ABA dependent and independent abiotic stress signalling. Biol. Plant. 2010, 54, 201–212. [Google Scholar] [CrossRef]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Navarro, L.; Bari, R.; Achard, P.; Lison, P.; Nemri, A.; Harberd, N.P.; Jones, J.D. DELLAs control plant immune responses by modulating the balance of jasmonic acid and salicylic acid signaling. Curr. Biol. 2008, 18, 650–655. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Chen, S.; Li, T.; Ma, X.; Liang, X.; Ding, X.; Liu, H.; Luo, L. OsGRAS23, a rice GRAS transcription factor gene, is involved in drought stress response through regulating expression of stress-responsive genes. BMC Plant Biol. 2015, 15, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Id in “Zhongmu No.1” Assembly | Isoelectric Point | Molecular Weight (Da) | Amino Acids | Subcellular Localization | Expressed in Root | Expressed in Leaf |
|---|---|---|---|---|---|---|---|
| MsGRAS1 | MsG0180004788.01.T01 | 6.41 | 55,671.46 | 487 | Nucleus | No | No |
| MsGRAS2 | MsG0280007861.01.T01 | 5.14 | 68,709.99 | 601 | Nucleus | Yes | No |
| MsGRAS3 | MsG0280007862.01.T01 | 5.16 | 68,262.82 | 596 | Nucleus | Yes | No |
| MsGRAS4 | MsG0280007863.01.T01 | 5.37 | 27,925.1 | 246 | Nucleus | No | No |
| MsGRAS5 | MsG0280009128.01.T01 | 5.47 | 56,013.31 | 503 | Cytoplasmic | Yes | Yes |
| MsGRAS6 | MsG0280010628.01.T01 | 5.96 | 46,813.87 | 415 | Cytoplasmic | Yes | No |
| MsGRAS7 | MsG0280011038.01.T01 | 5.42 | 73,172.09 | 639 | Nucleus | No | Yes |
| MsGRAS8 | MsG0280011039.01.T01 | 5.14 | 73,950.9 | 646 | Nucleus | Yes | Yes |
| MsGRAS9 | MsG0280011040.01.T01 | 5.47 | 86,165.78 | 761 | Nucleus | Yes | Yes |
| MsGRAS10 | MsG0280011041.01.T01 | 5.26 | 74,730.24 | 657 | Nucleus | Yes | Yes |
| MsGRAS11 | MsG0280011042.01.T01 | 5.51 | 74,363.63 | 656 | Nucleus | Yes | No |
| MsGRAS12 | MsG0380012325.01.T01 | 6.16 | 112,665.07 | 1002 | Nucleus | No | No |
| MsGRAS13 | MsG0380015272.01.T01 | 5 | 60,086.79 | 548 | Cytoplasmic | Yes | Yes |
| MsGRAS14 | MsG0380015408.01.T01 | 4.93 | 61,268.48 | 542 | Nucleus | Yes | Yes |
| MsGRAS15 | MsG0380015569.01.T01 | 9.24 | 42,996.71 | 389 | Nucleus | Yes | Yes |
| MsGRAS16 | MsG0380016451.01.T01 | 5.09 | 64,121.06 | 570 | Nucleus | Yes | Yes |
| MsGRAS17 | MsG0480020975.01.T01 | 5.9 | 72,561.93 | 634 | Nucleus | Yes | Yes |
| MsGRAS18 | MsG0480020976.01.T01 | 5.99 | 71,593.66 | 636 | Nucleus | Yes | Yes |
| MsGRAS19 | MsG0480021018.01.T01 | 5.62 | 76,258.96 | 673 | Nucleus | Yes | Yes |
| MsGRAS20 | MsG0480021019.01.T01 | 5.24 | 83,755.65 | 735 | Nucleus | Yes | Yes |
| MsGRAS21 | MsG0480021020.01.T01 | 5.5 | 68,897.38 | 601 | Nucleus | Yes | Yes |
| MsGRAS22 | MsG0480021642.01.T01 | 5.23 | 61,704.47 | 542 | Nucleus | Yes | Yes |
| MsGRAS23 | MsG0480021764.01.T01 | 5.3 | 50,123.6 | 452 | Nucleus | Yes | Yes |
| MsGRAS24 | MsG0480021771.01.T01 | 5.91 | 11,053.79 | 98 | Nucleus | No | Yes |
| MsGRAS25 | MsG0480021850.01.T01 | 5.2 | 36,549.71 | 329 | Chloroplast | Yes | Yes |
| MsGRAS26 | MsG0480023446.01.T01 | 5.58 | 43,284.82 | 377 | Cytoplasmic | No | No |
| MsGRAS27 | MsG0580024328.01.T01 | 5.52 | 54,028.86 | 481 | Cytoplasmic | Yes | Yes |
| MsGRAS28 | MsG0580024757.01.T01 | 6.1 | 47,251.93 | 417 | Cytoplasmic | No | No |
| MsGRAS29 | MsG0580027451.01.T05 | 6.61 | 30,294.32 | 271 | Chloroplast | No | No |
| MsGRAS30 | MsG0580027451.01.T04 | 6.52 | 34,146.74 | 304 | Chloroplast | No | No |
| MsGRAS31 | MsG0580027451.01.T01 | 5.03 | 52,231.82 | 467 | Nuclear | No | No |
| MsGRAS32 | MsG0580027451.01.T03 | 4.86 | 55,748.52 | 498 | Cytoplasmic | No | No |
| MsGRAS33 | MsG0580027451.01.T02 | 4.84 | 56,619.51 | 505 | Cytoplasmic | No | No |
| MsGRAS34 | MsG0580029906.01.T01 | 5.13 | 59,194.08 | 524 | Cytoplasmic | Yes | Yes |
| MsGRAS35 | MsG0580030094.01.T01 | 6 | 40,152.43 | 362 | Nucleus | Yes | Yes |
| MsGRAS36 | MsG0680032758.01.T01 | 6.41 | 74,615.66 | 671 | Chloroplast | Yes | Yes |
| MsGRAS37 | MsG0680033030.01.T01 | 6.62 | 74,073.15 | 666 | Chloroplast | Yes | Yes |
| MsGRAS38 | MsG0780037085.01.T01 | 5.61 | 75,040.53 | 676 | Nucleus | No | No |
| MsGRAS39 | MsG0780037984.01.T01 | 5.6 | 81,306.47 | 730 | Nucleus | Yes | Yes |
| MsGRAS40 | MsG0780037985.01.T01 | 5.4 | 81,430.55 | 731 | Nucleus | Yes | Yes |
| MsGRAS41 | MsG0780038783.01.T01 | 5.93 | 69,852.63 | 623 | Nucleus | Yes | Yes |
| MsGRAS42 | MsG0780039578.01.T01 | 6.19 | 73,091.07 | 658 | Nucleus | Yes | Yes |
| MsGRAS43 | MsG0780041365.01.T01 | 5.58 | 64,634.64 | 556 | Nucleus | No | No |
| MsGRAS44 | MsG0880042740.01.T01 | 5.72 | 61,482.27 | 552 | Nucleus | Yes | No |
| MsGRAS45 | MsG0880044171.01.T01 | 4.77 | 61,196.24 | 541 | Cytoplasmic | Yes | No |
| MsGRAS46 | MsG0880045957.01.T01 | 5.66 | 43,074.96 | 383 | Cytoplasmic | Yes | No |
| MsGRAS47 | MsG0880045975.01.T02 | 5.42 | 39,808.08 | 353 | Cytoplasmic | No | No |
| MsGRAS48 | MsG0880045975.01.T01 | 5.5 | 37,882 | 335 | Cytoplasmic | No | No |
| MsGRAS49 | MsG0880045989.01.T01 | 4.98 | 23,447.59 | 204 | Nuclear | No | No |
| MsGRAS50 | MsG0880046208.01.T01 | 5.51 | 75,345.09 | 678 | Chloroplast | Yes | No |
| MsGRAS51 | MsG0880047717.01.T01 | 5.81 | 60,581.39 | 540 | Nucleus | Yes | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Liu, X.; Wang, X.; Sun, M.; Song, R.; Mao, P.; Jia, S. Genome-Wide Identification of GRAS Gene Family and Their Responses to Abiotic Stress in Medicago sativa. Int. J. Mol. Sci. 2021, 22, 7729. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147729
Zhang H, Liu X, Wang X, Sun M, Song R, Mao P, Jia S. Genome-Wide Identification of GRAS Gene Family and Their Responses to Abiotic Stress in Medicago sativa. International Journal of Molecular Sciences. 2021; 22(14):7729. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147729
Chicago/Turabian StyleZhang, Han, Xiqiang Liu, Xuemeng Wang, Ming Sun, Rui Song, Peisheng Mao, and Shangang Jia. 2021. "Genome-Wide Identification of GRAS Gene Family and Their Responses to Abiotic Stress in Medicago sativa" International Journal of Molecular Sciences 22, no. 14: 7729. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147729