
Methylprednisolone Induces Extracellular Trap Formation and Enhances Bactericidal Effect of Canine Neutrophils


Abstract
:1. Introduction
2. Results and Discussion
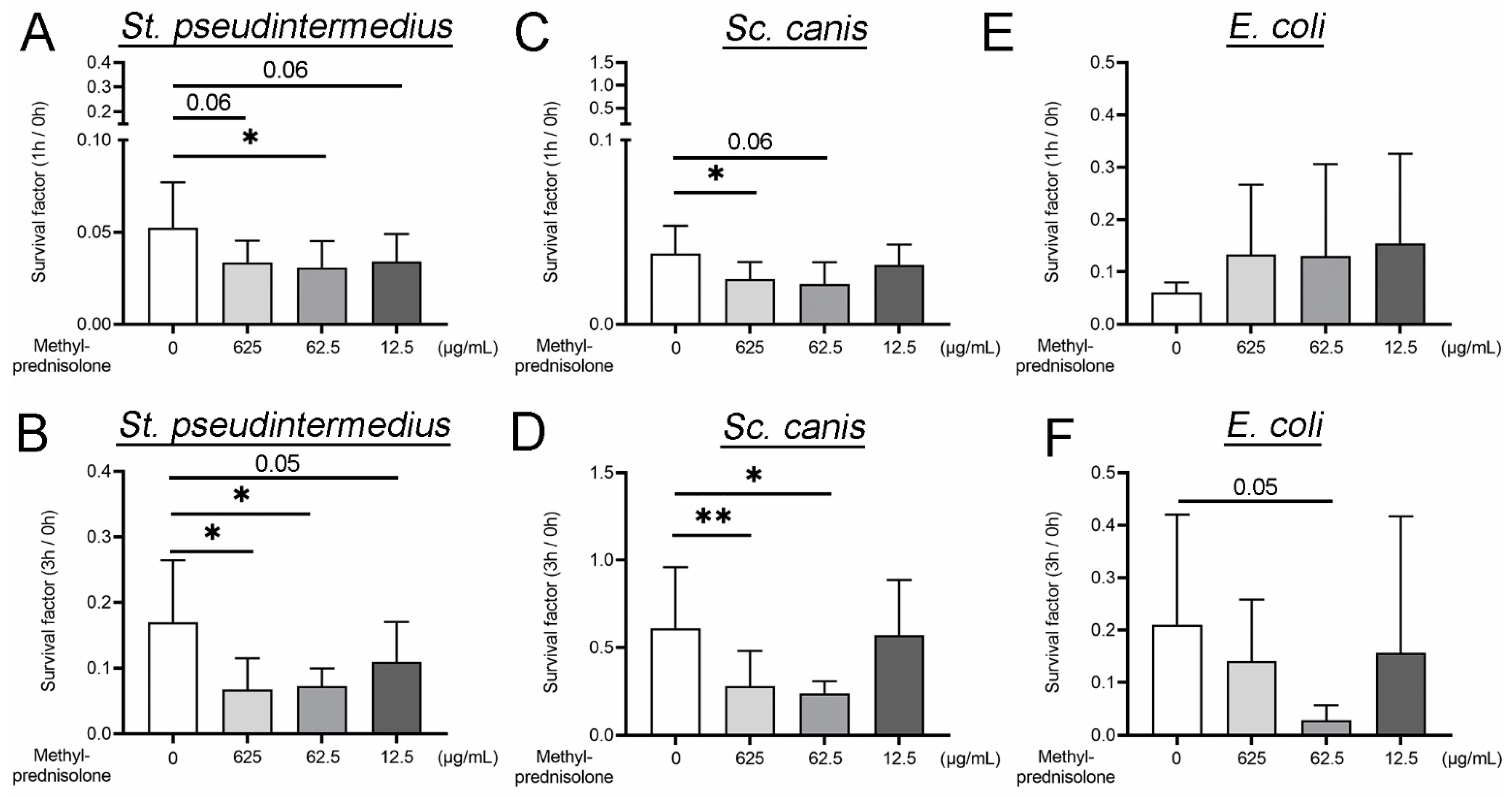
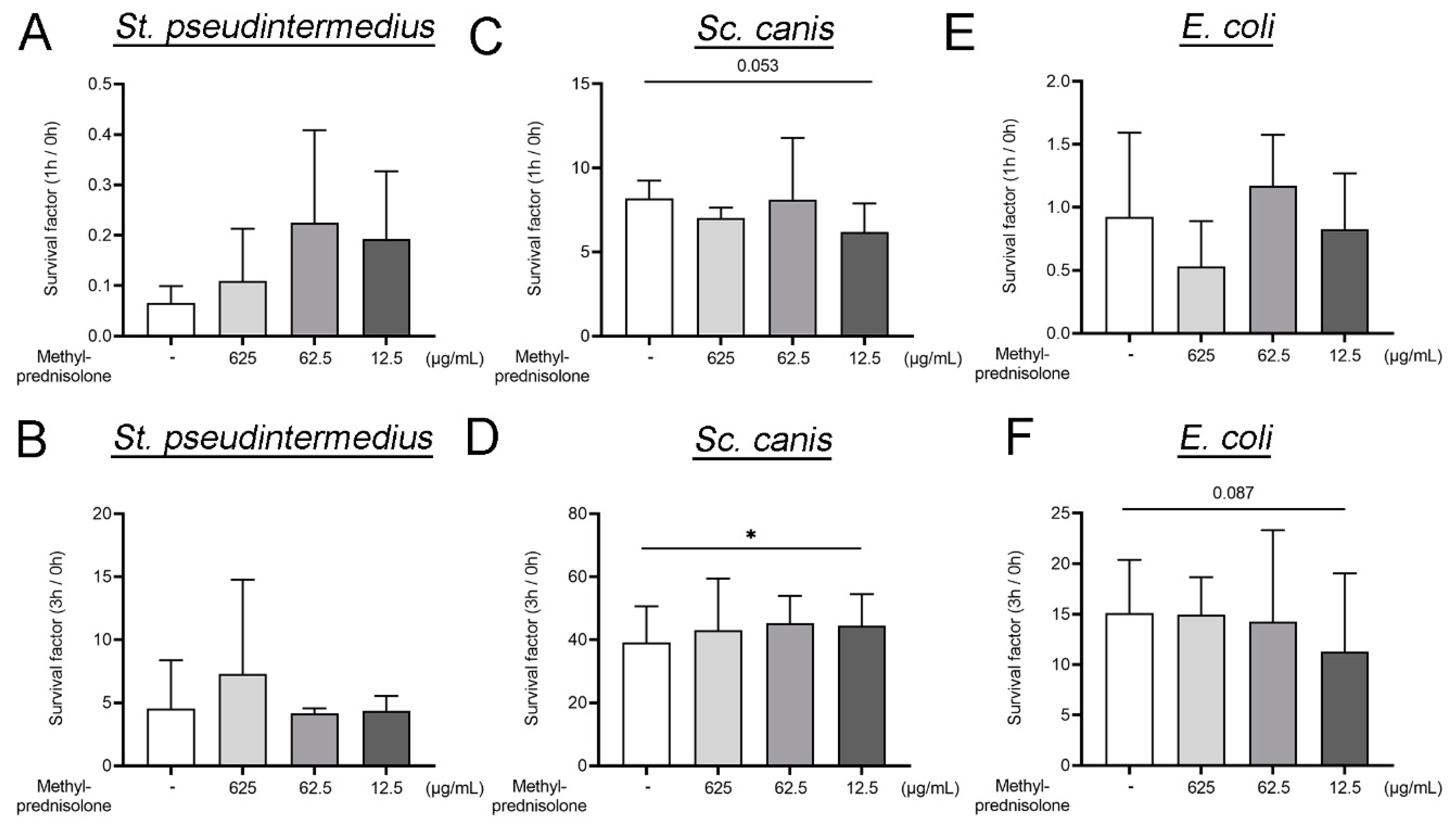
2.1. Methylprednisolone Enhances Neutrophil Killing of Gram Positve Bacteria, but Not of Gram Negative Bacteria
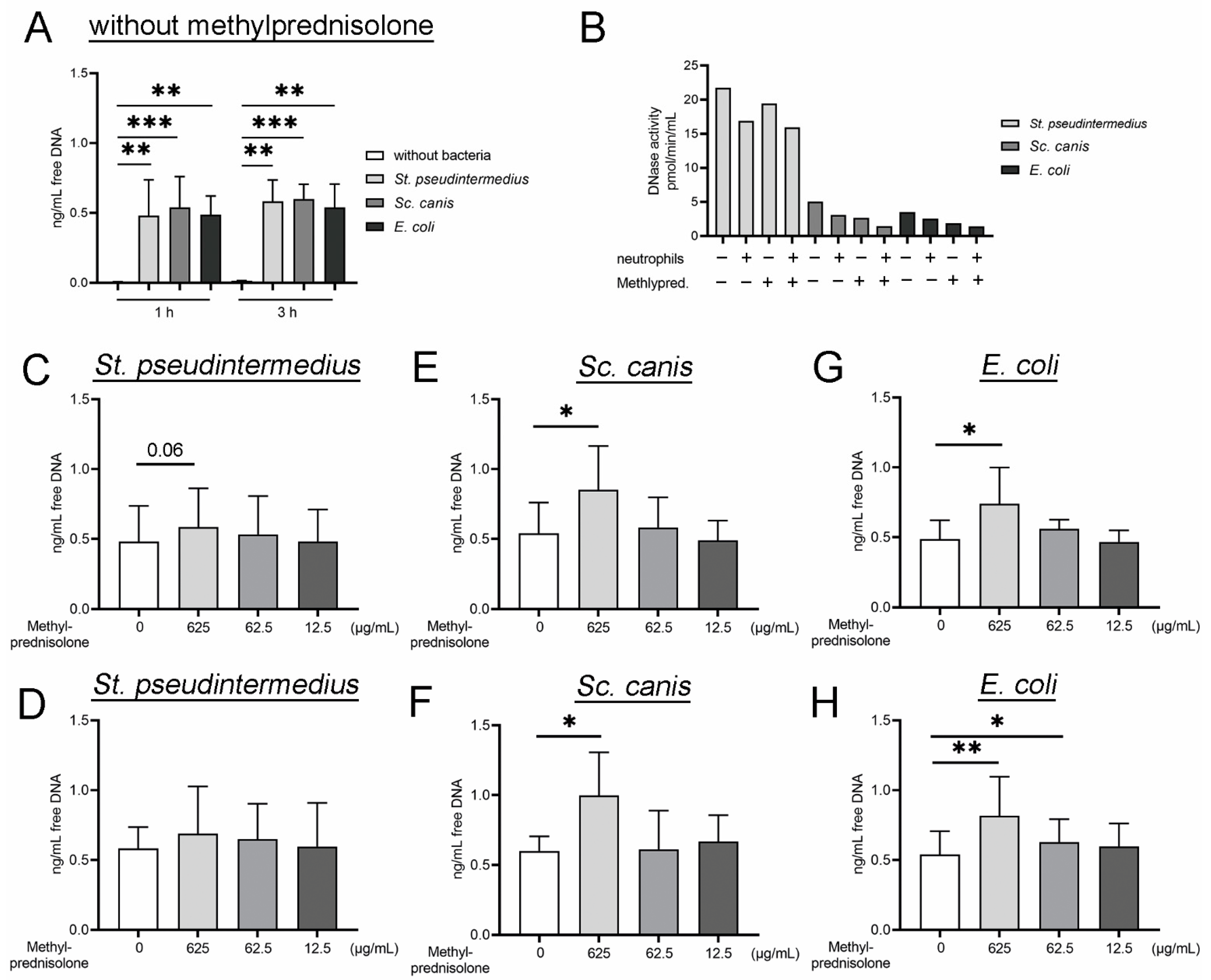
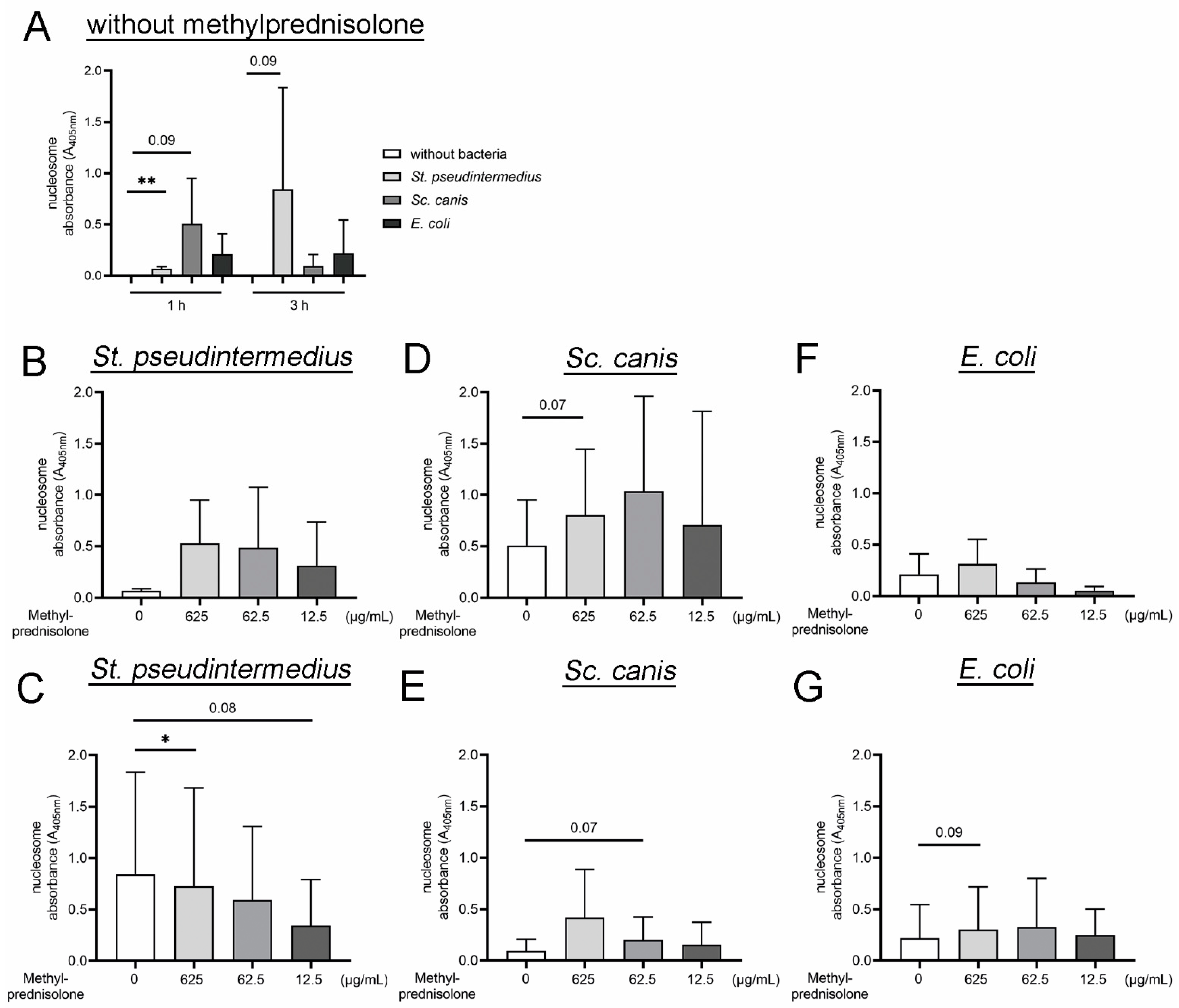
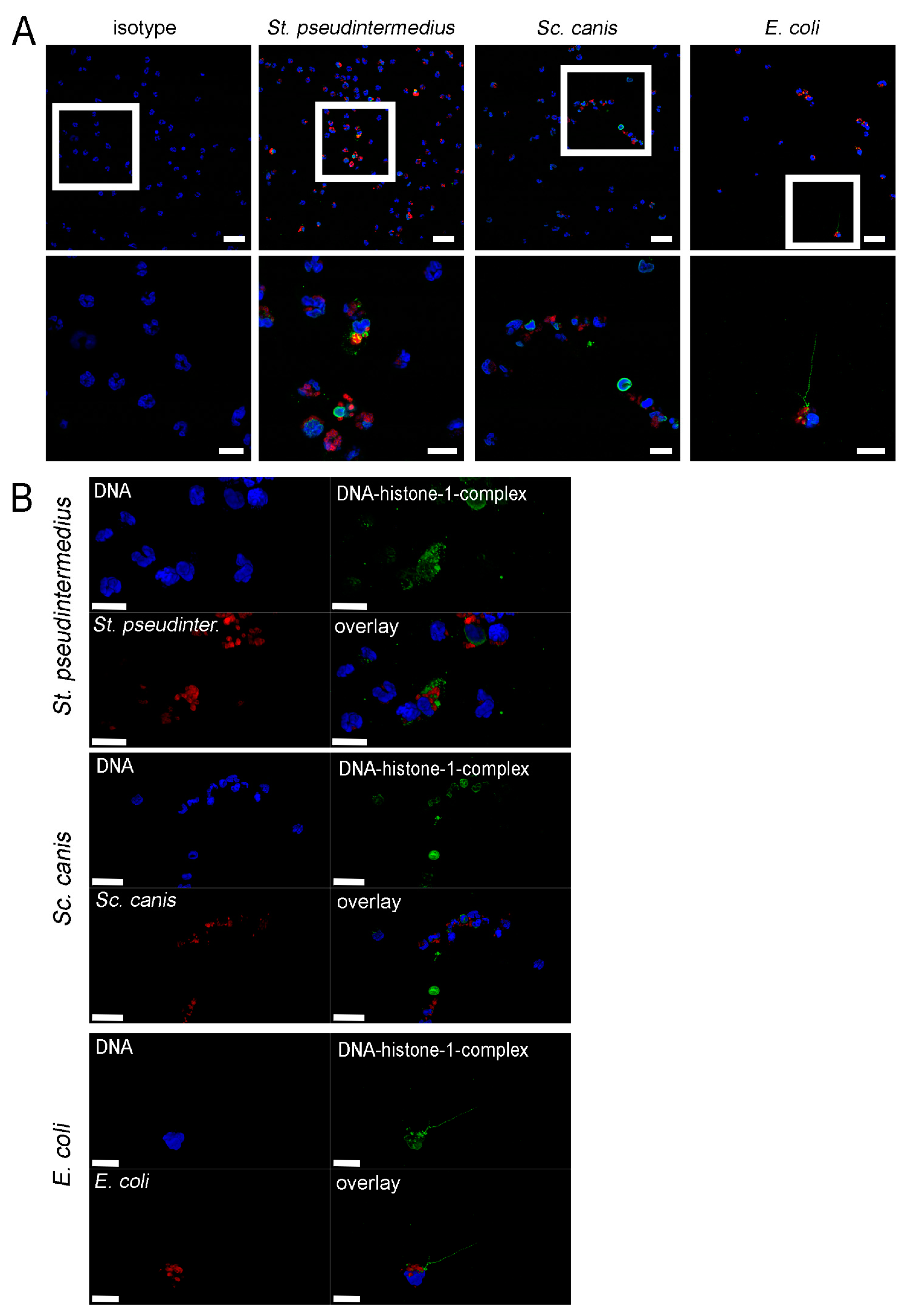
2.2. Methylprednislone Triggers the Release of Free DNA and Nucleosomes by Neutrophils during Bacterial Infection
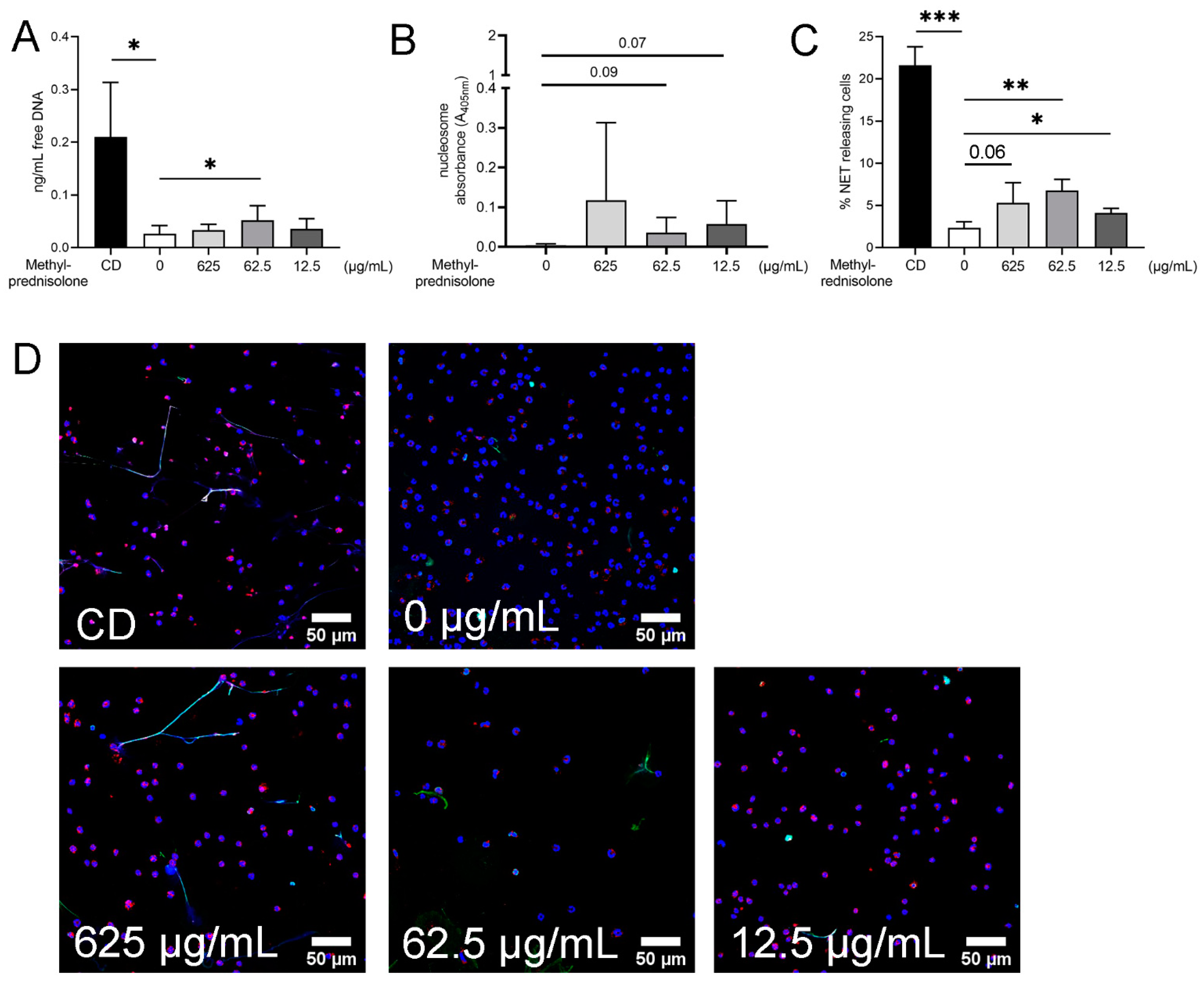
2.3. Detection of NETs Induced by Methylprednisolone and Release of Free DNA and Nucleosomes in Absence of Plasma and Bacteria
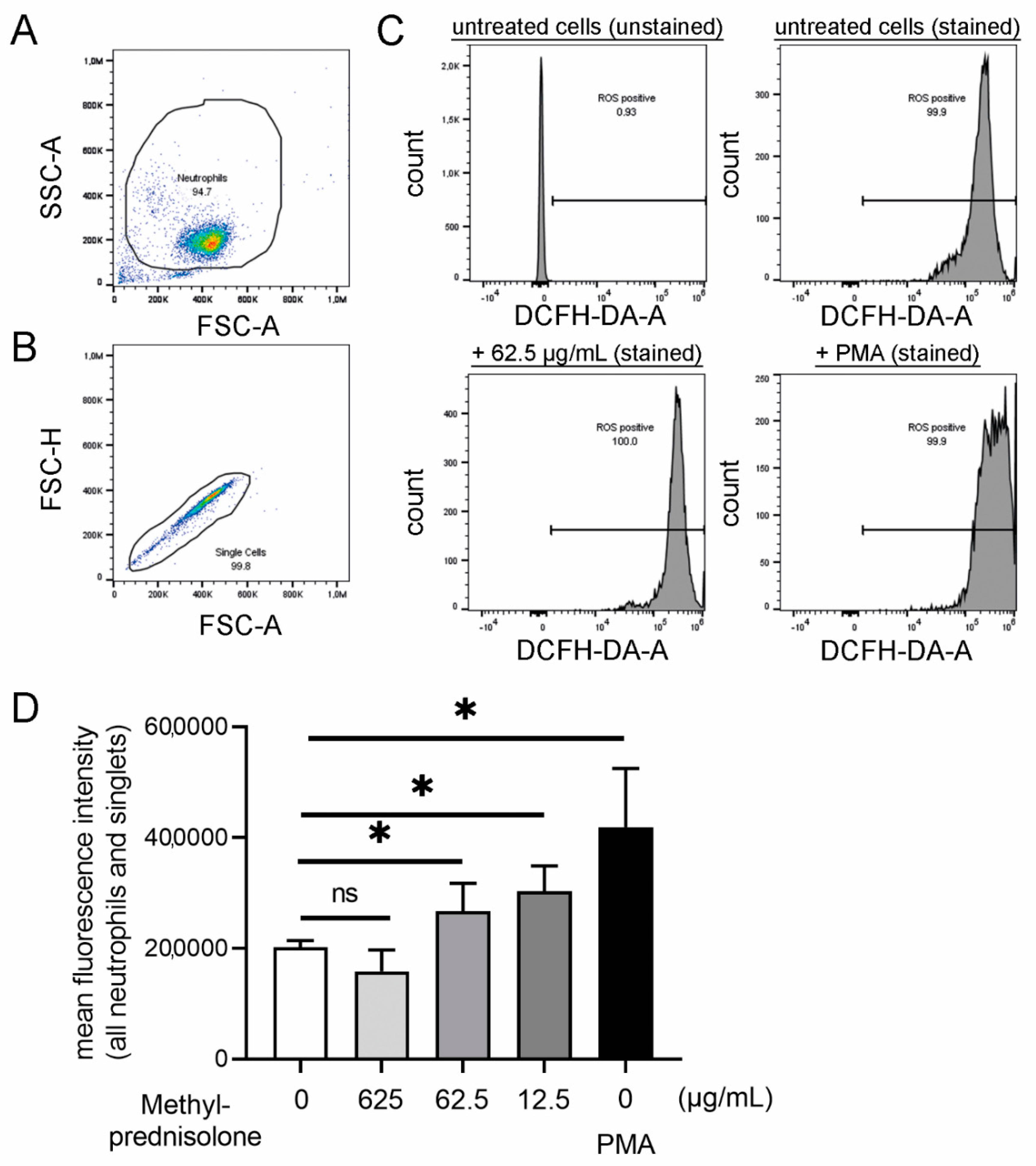
2.4. Other Neutrophil Mechanisms Influenced by Methlyprednisolone: Oxidative Burst, Phagocytosis, Chemotaxis and Cytokine Release
3. Materials and Methods
3.1. Sample Collection
3.2. Isolation of Canine Granulocytes
3.3. Bacterial Strains and Growth Conditions
3.4. Neutrophil Killing Assay
3.5. Survival of Bacteria in Presence of Methylprednisolone and Canine Plasma
3.6. NET Induction with Isolated Neutrophils
3.7. Immunofluorescence Staining of NETs
3.8. Co-Staining of Bacteria and NETs
3.9. Immunofluorescence Microscopy and NET Quantification
3.10. Pico Green Assay to Detect Free DNA
3.11. DNase Activity Assay
3.12. Measurement of Nucloesome Fragments by ELISA
3.13. ROS Analysis
3.14. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CCL17 | Thymus and activation-regulated chemokine |
| CFU | Colony forming unit |
| DNA | Deoxyribonucleic acid |
| DPI | Diphenyleneiodonium |
| E. coli | Escherichia coli |
| FACS | Fluorescence-activating cell sorting |
| h | Hours |
| IL-1 | Interleukin 1 |
| IL-6 | Interleukin 6 |
| IL-8 | Interleukin 8 |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MOI | Multiplicity of infection |
| NET | Neutrophil extracellular trap |
| PMN | Polymorphonuclear granulocyte |
| ROS | Reactive oxygen species |
| RPMI | Roswell Park Memorial Institute |
| Sc. | Streptococcus |
| SD | Standard deviation |
| SF | Survival factor |
| St. | Staphylococcus |
| THB | Todd Hewitt Broth |
| TNF-α | Tumor necrosis factor alpha |
| TSB | Tryptic soy broth |
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dog | WBC ×103/µL (6–12) | PMNs ×103/µL (3–10) | RBC ×106/µL (6–9) | PLT ×103/µL (150–500) | Blood Sample mL | Isolated PMNs ×106/mL |
|---|---|---|---|---|---|---|
| 1 | 7.91 | 5.02 | 6.23 | 226 | 10 | 19.6 |
| 2 | 10.97 | 7.26 | 6.66 | 221 | 13 | 24.6 |
| 3 | 6.27 | 3.81 | 6.16 | 232 | 10 | 14.9 |
| 4 | 10.63 | 6.73 | 7.4 | 204 | 10 | 19.9 |
| 5 | 6.38 | 3.23 | 7.23 | 199 | 10 | 17.1 |
| 6 | 8.11 | 5.62 | 7.43 | 358 | 12 | 26.1 |
| 7 | 6.92 | 4.45 | 6.19 | 169 | 11 | 13 |
| 8 | 5.43 | 2.81 | 6.68 | 206 | 12 | 15.6 |
| 9 | 8.87 | 5.59 | 7.08 | 242 | 10 | 16.6 |
| 10 | 7.24 | 4.43 | 6.9 | 122 | 13 | 26.8 |
References
- Whitley, N.T.; Day, M.J. Immunomodulatory drugs and their application to the management of canine immune-mediated disease. J. Small Anim. Pract. 2011, 52, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Viviano, K.R. Update on Immununosuppressive Therapies for Dogs and Cats. Vet. Clin. N. Am. Small Anim. Pract. 2013, 43, 1149–1170. [Google Scholar] [CrossRef] [PubMed]
- Cohn, L.A. Glucocorticosteroids as immunosuppressive agents. Semin. Vet. Med. Surg. Small Anim. 1997, 12, 150–156. [Google Scholar] [CrossRef]
- Swann, J.W.; Garden, O.A.; Fellman, C.L.; Glanemann, B.; Goggs, R.; LeVine, D.N.; Mackin, A.J.; Whitley, N.T. ACVIM consensus statement on the treatment of immune-mediated hemolytic anemia in dogs. J. Vet. Intern. Med. 2019, 33, 1141–1172. [Google Scholar] [CrossRef] [Green Version]
- Steffan, J.; Alexander, D.; Brovedani, F.; Fisch, R.D. Comparison of cyclosporine A with methylprednisolone for treatment of canine atopic dermatitis: A parallel, blinded, randomized controlled trial. Vet. Dermatol. 2003, 14, 11–22. [Google Scholar] [CrossRef]
- Cornelis, I.; Van Ham, L.; Gielen, I.; De Decker, S.; Bhatti, S.F.M. Clinical presentation, diagnostic findings, prognostic factors, treatment and outcome in dogs with meningoencephalomyelitis of unknown origin: A review. Vet. J. 2019, 244, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Craven, M.D.; Washabau, R.J. Comparative pathophysiology and management of protein-losing enteropathy. J. Vet. Intern. Med. 2019, 33, 383–402. [Google Scholar] [CrossRef]
- Johnson, K.C.; Mackin, A. Canine immune-mediated polyarthritis: PART 2: Diagnosis and treatment. J. Am. Anim. Hosp. Assoc. 2012, 48, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Cerami, A. Inflammatory cytokines. Clin. Immunol. Immunopathol. 1992, 62, S3–S10. [Google Scholar] [CrossRef]
- Rossi, A.G.; Haslett, C. Inflammation, Cell Injury, and Apoptosis. In Proinflammatory and Antiinflammatory Peptides; CRC Press: New York, NY, USA, 2020; pp. 9–24. [Google Scholar]
- Segal, A.W. How neutrophils kill microbes. Annu. Rev. Immunol. 2005, 23, 197–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayadas, T.N.; Cullere, X.; Lowell, C.A. The Multifaceted Functions of Neutrophils. Annu. Rev. Pathol. Mech. Dis. 2014, 9, 181–218. [Google Scholar] [CrossRef] [Green Version]
- Malech, H.L.; DeLeo, F.R.; Quinn, M.T. The Role of Neutrophils in the Immune System: An Overview. Methods Mol. Biol. 2020, 2087, 3–10. [Google Scholar]
- Teng, T.S.; Ji, A.L.; Ji, X.Y.; Li, Y.Z. Neutrophils and immunity: From bactericidal action to being conquered. J. Immunol. Res. 2017, 2017, 1–14. [Google Scholar] [CrossRef]
- Ermert, D.; Zychlinsky, A.; Urban, C. Fungal and bacterial killing by neutrophils. Methods Mol. Biol. 2008, 470, 293–312. [Google Scholar]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Brinkmann, V. Neutrophil Extracellular Traps in the Second Decade. J. Innate Immun. 2018, 10, 414–421. [Google Scholar] [CrossRef]
- Yang, H.; Biermann, M.H.; Brauner, J.M.; Liu, Y.; Zhao, Y.; Herrmann, M. New Insights into Neutrophil Extracellular Traps: Mechanisms of Formation and Role in Inflammation. Front. Immunol. 2016, 7, 302. [Google Scholar] [CrossRef] [Green Version]
- Goggs, R.; Jeffery, U.; LeVine, D.N.; Li, R.H.L. Neutrophil-Extracellular Traps, Cell-Free DNA, and Immunothrombosis in Companion Animals: A Review. Vet. Pathol. 2020, 57, 6–23. [Google Scholar] [CrossRef]
- Pieterse, E.; van der Vlag, J. Breaking Immunological Tolerance in Systemic Lupus Erythematosus. Front. Immunol. 2014, 5, 164. [Google Scholar] [CrossRef]
- Hakkim, A.; Furnrohr, B.G.; Amann, K.; Laube, B.; Abed, U.A.; Brinkmann, V.; Herrmann, M.; Voll, R.E.; Zychlinsky, A. Impairment of neutrophil extracellular trap degradation is associated with lupus nephritis. Proc. Natl. Acad. Sci. USA 2010, 107, 9813–9818. [Google Scholar] [CrossRef] [Green Version]
- Knight, J.S.; Carmona-Rivera, C.; Kaplan, M.J. Proteins derived from neutrophil extracellular traps may serve as self-antigens and mediate organ damage in autoimmune diseases. Front. Immunol. 2012, 3, 380. [Google Scholar] [CrossRef] [Green Version]
- Knight, J.S.; Kaplan, M.J. Lupus neutrophils: “NET” gain in understanding lupus pathogenesis. Curr. Opin. Rheumatol. 2012, 24, 441–450. [Google Scholar] [CrossRef]
- Jeffery, U.; LeVine, D.N. Canine Neutrophil Extracellular Traps Enhance Clot Formation and Delay Lysis. Vet. Pathol. 2018, 55, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Jeffery, U.; Kimura, K.; Gray, R.; Lueth, P.; Bellaire, B.; LeVine, D. Dogs cast NETs too: Canine neutrophil extracellular traps in health and immune-mediated hemolytic anemia. Vet. Immunol. Immunopathol. 2015, 168, 262–268. [Google Scholar] [CrossRef]
- Lawson, C.; Smith, S.A.; O’Brien, M.; McMichael, M. Neutrophil Extracellular Traps in Plasma from Dogs with Immune-mediated Hemolytic Anemia. J. Vet. Intern. Med. 2018, 32, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Caro, T.; Conejeros, I.; Zhou, E.; Pikhovych, A.; Gärtner, U.; Hermosilla, C.; Kulke, D.; Taubert, A. Dirofilaria immitis microfilariae and third-stage larvae induce canine NETosis resulting in different types of neutrophil extracellular traps. Front. Immunol. 2018, 9, 968. [Google Scholar] [CrossRef]
- de Buhr, N.; Bonilla, M.C.; Jimenez-Soto, M.; von Köckritz-Blickwede, M.; Dolz, G. Extracellular Trap Formation in Response to Trypanosoma cruzi Infection in Granulocytes Isolated from Dogs and Common Opossums, Natural Reservoir Hosts. Front. Microbiol. 2018, 9, 966. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Wang, Z.; Liu, X.; Wang, C.; Han, Z.; Wu, D.; Zhang, Y.; Zhang, X.; Yang, Z.; Liu, Q. Toxoplasma gondii Triggers Neutrophil Extracellular Traps Release in Dogs. Front. Cell. Infect. Microbiol. 2020, 10, 429. [Google Scholar] [CrossRef]
- Li, R.H.L.; Johnson, L.R.; Kohen, C.; Tablin, F. A novel approach to identifying and quantifying neutrophil extracellular trap formation in septic dogs using immunofluorescence microscopy. BMC Vet. Res. 2018, 14, 210. [Google Scholar] [CrossRef] [Green Version]
- Rebordão, M.R.; Alexandre-Pires, G.; Carreira, M.; Adriano, L.; Carneiro, C.; Nunes, T.; Mateus, L.; Ferreira-Dias, G. Bacteria causing pyometra in bitch and queen induce neutrophil extracellular traps. Vet. Immunol. Immunopathol. 2017, 192, 8–12. [Google Scholar] [CrossRef]
- Baien, S.H.; Seele, J.; Henneck, T.; Freibrodt, C.; Szura, G.; Moubasher, H.; Nau, R.; Brogden, G.; Mörgelin, M.; Singh, M.; et al. Antimicrobial and Immunomodulatory Effect of Gum Arabic on Human and Bovine Granulocytes Against Staphylococcus aureus and Escherichia coli. Front. Immunol. 2020, 10, 3119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corriden, R.; Hollands, A.; Olson, J.; Derieux, J.; Lopez, J.; Chang, J.T.; Gonzalez, D.J.; Nizet, V. Tamoxifen augments the innate immune function of neutrophils through modulation of intracellular ceramide. Nat. Commun. 2015, 6, 8369. [Google Scholar] [CrossRef] [Green Version]
- Jerjomiceva, N.; Seri, H.; Völlger, L.; Wang, Y.; Zeitouni, N.; Naim, H.Y.; von Köckritz-Blickwede, M. Enrofloxacin Enhances the Formation of Neutrophil Extracellular Traps in Bovine Granulocytes. J. Innate Immun. 2014, 6, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Munguia, J.; Nizet, V. Pharmacological Targeting of the Host–Pathogen Interaction: Alternatives to Classical Antibiotics to Combat Drug-Resistant Superbugs. Trends Pharmacol. Sci. 2017, 38, 473–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, S.H.; Kang, M.H.; Sur, J.H.; Park, H.M. Staphylococcus pseudintermedius infection associated with nodular skin lesions and systemic inflammatory response syndrome in a dog. Can. Vet. J. 2014, 55, 480. [Google Scholar]
- Bannoehr, J.; Guardabassi, L. Staphylococcus pseudintermedius in the dog: Taxonomy, diagnostics, ecology, epidemiology and pathogenicity. Vet. Dermatol. 2012, 23, 253-e52. [Google Scholar] [CrossRef]
- Hillier, A.; Lloyd, D.H.; Weese, J.S.; Blondeau, J.M.; Boothe, D.; Breitschwerdt, E.; Guardabassi, L.; Papich, M.G.; Rankin, S.; Turnidge, J.D.; et al. Guidelines for the diagnosis and antimicrobial therapy of canine superficial bacterial folliculitis (Antimicrobial Guidelines Working Group of the International Society for Companion Animal Infectious Diseases). Vet. Dermatol. 2014, 25, 163-e43. [Google Scholar] [CrossRef]
- Rubin, J.E.; Gaunt, M.C. Urinary tract infection caused by methicillin-resistant Staphylococcus pseudintermedius in a dog. Can. Vet. J. 2011, 52, 162. [Google Scholar]
- Weese, J.S.; Blondeau, J.; Boothe, D.; Guardabassi, L.G.; Gumley, N.; Papich, M.; Jessen, L.R.; Lappin, M.; Rankin, S.; Westropp, J.L.; et al. International Society for Companion Animal Infectious Diseases (ISCAID) guidelines for the diagnosis and management of bacterial urinary tract infections in dogs and cats. Vet. J. 2019, 247, 8–25. [Google Scholar] [CrossRef]
- Lappin, M.R.; Blondeau, J.; Boothe, D.; Breitschwerdt, E.B.; Guardabassi, L.; Lloyd, D.H.; Papich, M.G.; Rankin, S.C.; Sykes, J.E.; Turnidge, J.; et al. Antimicrobial use Guidelines for Treatment of Respiratory Tract Disease in Dogs and Cats: Antimicrobial Guidelines Working Group of the International Society for Companion Animal Infectious Diseases. J. Vet. Intern. Med. 2017, 31, 279–294. [Google Scholar] [CrossRef]
- Greene, C.E.; Prescott, J.F. Streptococcal and other gram-positive bacterial infections. In Infectious Diseases of the Dog and Cat; Elsevier Saunders: St. Louis, MO, USA, 2006. [Google Scholar]
- Murase, T.; Morita, T.; Sunagawa, Y.; Sawada, M.; Shimada, A.; Sato, K.; Hikasa, Y. Isolation of Streptococcus canis from a Japanese raccoon dog with fibrinous pleuropneumonia. Vet. Rec. 2003, 153, 471–472. [Google Scholar] [CrossRef] [PubMed]
- White, S. Further evidence of streptococcal toxic shock syndrome in pets. J. Am. Vet. Med. Assoc. 1996, 209, 1994–1995. [Google Scholar]
- Sharma, B.; Srivastava, M.K.; Srivastava, A.; Singh, R. Canine streptococcal toxic shock syndrome associated with necrotizing fasciitis: An overview. Vet. World 2012, 5, 311. [Google Scholar] [CrossRef]
- DeWinter, L.M.; Prescott, J.F. Relatedness of Streptococcus canis from canine streptococcal toxic shock syndrome and necrotizing fasciitis. Can. J. Vet. Res. 1999, 63, 90–95. [Google Scholar] [PubMed]
- Miller, C.W.; Prescott, J.F.; Mathews, K.A.; Betschel, S.D.; Yager, J.A.; Guru, V.; DeWinter, L.; Low, D.E. Streptococcal toxic shock syndrome in dogs. J. Am. Vet. Med. Assoc. 1996, 209, 1421–1426. [Google Scholar] [PubMed]
- Harrer, J.; Dorsch, R. Bacterial urinary tract infection and subclinical bacteriuria in the dog: A current review. Tierarztl. Prax. Ausgabe K Kleintiere Heimtiere 2020, 48, 270–284. [Google Scholar]
- Beutin, L. Escherichia coli as a pathogen in dogs and cats. Vet. Res. 1999, 30, 285–298. [Google Scholar]
- Marks, S.L.; Rankin, S.C.; Byrne, B.A.; Weese, J.S. Enteropathogenic Bacteria in Dogs and Cats: Diagnosis, Epidemiology, Treatment, and Control. J. Vet. Intern. Med. 2011, 25, 1195–1208. [Google Scholar] [CrossRef]
- Pilla, R.; Suchodolski, J.S. The Role of the Canine Gut Microbiome and Metabolome in Health and Gastrointestinal Disease. Front. Vet. Sci. 2020, 6, 498. [Google Scholar] [CrossRef] [Green Version]
- Minamoto, Y.; Minamoto, T.; Isaiah, A.; Sattasathuchana, P.; Buono, A.; Rangachari, V.R.; McNeely, I.H.; Lidbury, J.; Steiner, J.M.; Suchodolski, J.S. Fecal short-chain fatty acid concentrations and dysbiosis in dogs with chronic enteropathy. J. Vet. Intern. Med. 2019, 33, 1608–1618. [Google Scholar] [CrossRef] [Green Version]
- Toutain, P.L.; Autefage, A.; Oukessou, M.; Alvinerie, M. Pharmacokinetics of methylprednisolone succinate, methylprednisolone, and lidocaine in the normal dog and during hemorrhagic shock. J. Pharm. Sci. 1987, 76, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Toutain, P.L.; Koritz, G.D.; Fayolle, P.M.; Alvinerie, M. Pharmacokinetics of methylprednisolone, methylprednisolone sodium succinate, and methylprednisolone acetate in dogs. J. Pharm. Sci. 1986, 75, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Aharon, M.A.; Prittie, J.E.; Buriko, K. A review of associated controversies surrounding glucocorticoid use in veterinary emergency and critical care. J. Vet. Emerg. Crit. Care 2017, 27, 267–277. [Google Scholar] [CrossRef]
- Dowling, S.R.; Webb, J.; Foster, J.D.; Ginn, J.; Foy, D.S.; Trepanier, L.A. Opportunistic fungal infections in dogs treated with ciclosporin and glucocorticoids: Eight cases. J. Small Anim. Pract. 2016, 57, 105–109. [Google Scholar] [CrossRef]
- Peterson, A.L.; Torres, S.M.F.; Rendahl, A.; Koch, S.N. Frequency of urinary tract infection in dogs with inflammatory skin disorders treated with ciclosporin alone or in combination with glucocorticoid therapy: A retrospective study. Vet. Dermatol. 2012, 23, 201-e43. [Google Scholar] [CrossRef]
- Torres, S.M.F.; Diaz, S.F.; Nogueira, S.A.; Jessen, C.; Polzin, D.J.; Gilbert, S.M.; Horne, K.L. Frequency of urinary tract infection among dogs with pruritic disorders receiving long-term glucocorticoid treatment. J. Am. Vet. Med. Assoc. 2005, 227, 239–243. [Google Scholar] [CrossRef]
- Meduri, G.U.; Kanangat, S.; Bronze, M.; Patterson, D.R.; Meduri, C.U.; Pak, C.; Tolley, E.A.; Schaberg, D.R. Effects of methylprednisolone on intracellular bacterial growth. Clin. Diagn. Lab. Immunol. 2001, 8, 1156–1163. [Google Scholar] [CrossRef] [Green Version]
- Cao, K.; Huang, L.; Liu, J.; An, H.; Shu, Y.; Han, Z. Inhibitory effects of high-dose methylprednisolone on bacterial translocation from gut and endotoxin release following acute spinal cord injury-induced paraplegia in rats. Neural Regen. Res. 2010, 5, 456–460. [Google Scholar]
- Arvidsson, S.; Falt, K.; Haglund, U. Feline E. Coli bacteremia—Effects of misoprostol/scavengers or methylprednisolone on hemodynamic reactions and gastrointestinal mucosal injury. Acta Chir. Scand. 1990, 156, 215–221. [Google Scholar]
- Flynn, P.M.; Shenep, J.L.; Stokes, D.C.; Hildner, W.K.; Mackert, P.W.; Snellgrove, R.L.; Rehg, J.E. Effect of Methylprednisolone on Bacterial Clearance and Endotoxin Liberation during Experimental Sepsis Induced by Gram-Negative Bacteria. Infect. Immun. 1986, 52, 26–30. [Google Scholar]
- Robinson, J.M. Reactive oxygen species in phagocytic leukocytes. Histochem. Cell Biol. 2008, 130, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Margraf, S.; Lögters, T.; Reipen, J.; Altrichter, J.; Scholz, M.; Windolf, J. Neutrophil-derived circulating free DNA (cf-DNA/NETs): A potential prognostic marker for posttraumatic development of inflammatory second hit and sepsis. Shock 2008, 30, 352–358. [Google Scholar] [CrossRef]
- Altrichter, J.; Zedler, S.; Kraft, R.; Faist, E.; Mitzner, S.R.; Sauer, M.; Windolf, J.; Scholz, M.; Lögters, T. Neutrophil-derived circulating free DNA (cf-DNA/NETs), a potential prognostic marker for mortality in patients with severe burn injury. Eur. J. Trauma Emerg. Surg. 2010, 36, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Herzog, S.; Dach, F.; de Buhr, N.; Niemann, S.; Schlagowski, J.; Chaves-Moreno, D.; Neumann, C.; Goretzko, J.; Schwierzeck, V.; Mellmann, A.; et al. High Nuclease Activity of Long Persisting Staphylococcus aureus Isolates Within the Airways of Cystic Fibrosis Patients Protects Against NET-Mediated Killing. Front. Immunol. 2019, 10, 2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Buhr, N.; Neumann, A.; Jerjomiceva, N.; von Köckritz-Blickwede, M.; Baums, C.G. Streptococcus suis DNase SsnA contributes to degradation of neutrophil extracellular traps (NETs) and evasion of NET-mediated antimicrobial activity. Microbiology 2014, 160, 385–395. [Google Scholar] [CrossRef] [Green Version]
- de Buhr, N.; Stehr, M.; Neumann, A.; Naim, H.Y.; Valentin-Weigand, P.; von Köckritz-Blickwede, M.; Baums, C.G. Identification of a novel DNase of Streptococcus suis (EndAsuis) important for neutrophil extracellular trap degradation during exponential growth. Microbiology 2015, 161, 838–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storisteanu, D.M.L.; Pocock, J.M.; Cowburn, A.S.; Juss, J.K.; Nadesalingam, A.; Nizet, V.; Chilvers, E.R. Evasion of neutrophil extracellular traps by respiratory pathogens. Am. J. Respir. Cell Mol. Biol. 2017, 56, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, T.; Fisher, J.; Bakochi, A.; Neumann, A.; Cardoso, J.F.P.; Karlsson, C.A.Q.; Pavan, C.; Lundgaard, I.; Nilson, B.; Reinstrup, P.; et al. Neutrophil extracellular traps in the central nervous system hinder bacterial clearance during pneumococcal meningitis. Nat. Commun. 2019, 10, 1667. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, U.; Ruterbories, L.; Hanel, R.; LeVine, D.N. Cell-Free DNA and DNase Activity in Dogs with Immune-Mediated Hemolytic Anemia. J. Vet. Intern. Med. 2017, 31, 1441–1450. [Google Scholar] [CrossRef] [PubMed]
- Bang, O.Y.; Chung, J.-W.; Cho, Y.H.; Oh, M.J.; Seo, W.-K.; Kim, G.-M.; Ahn, M.-J. Circulating DNAs, a Marker of Neutrophil Extracellular Traposis and Cancer-Related Stroke. Stroke 2019, 50, 2944–2947. [Google Scholar] [CrossRef]
- Aucamp, J.; Bronkhorst, A.J.; Badenhorst, C.P.S.; Pretorius, P.J. The diverse origins of circulating cell-free DNA in the human body: A critical re-evaluation of the literature. Biol. Rev. 2018, 93, 1649–1683. [Google Scholar] [CrossRef]
- Peng, L.; Li, L.; He, X.L.; Yu, J.Y.; Zeng, Z.J.; Yang, W.J.; Zhang, B.; Zhang, T.S.; Cao, H.; Huang, S.H.; et al. Memantine Displays Antimicrobial Activity by Enhancing Escherichia coli Pathogen-Induced Formation of Neutrophil Extracellular Traps. Front. Cell. Infect. Microbiol. 2020, 10, 47. [Google Scholar] [CrossRef]
- Hoppenbrouwers, T.; Autar, A.S.A.; Sultan, A.R.; Abraham, T.E.; van Cappellen, W.A.; Houtsmuller, A.B.; van Wamel, W.J.B.; van Beusekom, H.M.M.; van Neck, J.W.; de Maat, M.P.M. In vitro induction of NETosis: Comprehensive live imaging comparison and systematic review. PLoS ONE 2017, 12, e0176472. [Google Scholar] [CrossRef] [Green Version]
- Croxen, M.A.; Law, R.J.; Scholz, R.; Keeney, K.M.; Wlodarska, M.; Finlay, B.B. Recent advances in understanding enteric pathogenic Escherichia coli. Clin. Microbiol. Rev. 2013, 26, 822–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Buhr, N.; von Köckritz-Blickwede, M. How Neutrophil Extracellular Traps Become Visible. J. Immunol. Res. 2016, 2016, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmann, V.; Laube, B.; Abed, U.A.; Goosmann, C.; Zychlinsky, A. Neutrophil extracellular traps: How to generate and visualize them. J. Vis. Exp. 2010, 36, 1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gál, Z.; Gézsi, A.; Pállinger, É.; Visnovitz, T.; Nagy, A.; Kiss, A.; Sultész, M.; Csoma, Z.; Tamási, L.; Gálffy, G.; et al. Plasma neutrophil extracellular trap level is modified by disease severity and inhaled corticosteroids in chronic inflammatory lung diseases. Sci. Rep. 2020, 10, 4320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, A.; Boivin, R.; Cano, P.; Murcia, Y.; Bazin, I.; Lavoie, J.P. Neutrophil extracellular traps are downregulated by glucocorticosteroids in lungs in an equine model of asthma. Respir. Res. 2017, 18, 207. [Google Scholar] [CrossRef]
- Fan, F.; Huang, X.; Yuan, K.; Zhu, B.; Zhao, Y.; Hu, R.; Wan, T.; Zhu, L.; Jin, X. Glucocorticoids May Exacerbate Fungal Keratitis by Increasing Fungal Aggressivity and Inhibiting the Formation of Neutrophil Extracellular Traps. Curr. Eye Res. 2020, 45, 124–133. [Google Scholar] [CrossRef]
- Lapponi, M.J.; Carestia, A.; Landoni, V.I.; Rivadeneyra, L.; Etulain, J.; Negrotto, S.; Pozner, R.G.; Schattner, M. Regulation of neutrophil extracellular trap formation by anti-inflammatory drugs. J. Pharmacol. Exp. Ther. 2013, 345, 430–437. [Google Scholar] [CrossRef]
- Bystrzycka, W.; Moskalik, A.; Sieczkowska, S.; Manda-Handzlik, A.; Demkow, U.; Ciepiela, O. The effect of clindamycin and amoxicillin on neutrophil extracellular trap (NET) release. Cent. Eur. J. Immunol. 2016, 41, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Konstantinidis, T.; Kambas, K.; Mitsios, A.; Panopoulou, M.; Tsironidou, V.; Dellaporta, E.; Kouklakis, G.; Arampatzioglou, A.; Angelidou, I.; Mitroulis, I.; et al. Immunomodulatory role of clarithromycin in Acinetobacter baumannii infection via formation of neutrophil extracellular traps. Antimicrob. Agents Chemother. 2016, 60, 1040–1048. [Google Scholar] [CrossRef] [Green Version]
- Manda-Handzlik, A.; Bystrzycka, W.; Sieczkowska, S.; Demkow, U.; Ciepiela, O. Antibiotics modulate the ability of neutrophils to release neutrophil extracellular traps. Adv. Exp. Med. Biol. 2017, 944, 47–52. [Google Scholar]
- Bystrzycka, W.; Manda-Handzlik, A.; Sieczkowska, S.; Moskalik, A.; Demkow, U.; Ciepiela, O. Azithromycin and chloramphenicol diminish neutrophil extracellular traps (NETs) release. Int. J. Mol. Sci. 2017, 18, 2666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fingerhut, L.; Dolz, G.; de Buhr, N. What Is the Evolutionary Fingerprint in Neutrophil Granulocytes? Int. J. Mol. Sci. 2020, 21, 4523. [Google Scholar] [CrossRef] [PubMed]
- Capucetti, A.; Albano, F.; Bonecchi, R. Multiple Roles for Chemokines in Neutrophil Biology. Front. Immunol. 2020, 11, 1259. [Google Scholar] [CrossRef] [PubMed]
- Rosales, C. Neutrophils at the crossroads of innate and adaptive immunity. J. Leukoc. Biol. 2020, 108, 377–396. [Google Scholar] [CrossRef] [PubMed]
- Stoiber, W.; Obermayer, A.; Steinbacher, P.; Krautgartner, W. The Role of Reactive Oxygen Species (ROS) in the Formation of Extracellular Traps (ETs) in Humans. Biomolecules 2015, 5, 702–723. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel cell death program leads to neutrophil extracellular traps. J. Cell Biol. 2007, 176, 231–241. [Google Scholar] [CrossRef]
- Nunes-Silva, A.; Bernardes, P.T.T.; Rezende, B.M.; Lopes, F.; Gomes, E.C.; Marques, P.E.; Lima, P.M.A.; Coimbra, C.C.; Menezes, G.B.; Teixeira, M.M.; et al. Treadmill Exercise Induces Neutrophil Recruitment into Muscle Tissue in a Reactive Oxygen Species-Dependent Manner. An Intravital Microscopy Study. PLoS ONE 2014, 9, e96464. [Google Scholar] [CrossRef]
- Jorch, S.K.; Kubes, P. An emerging role for neutrophil extracellular traps in noninfectious disease. Nat. Med. 2017, 23, 279–287. [Google Scholar] [CrossRef]
- Yipp, B.G.; Kubes, P. NETosis: How vital is it? Blood 2013, 122, 2784–2794. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.R.; Ma, A.C.; Tavener, S.A.; McDonald, B.; Goodarzi, Z.; Kelly, M.M.; Patel, K.D.; Chakrabarti, S.; McAvoy, E.; Sinclair, G.D.; et al. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nat. Med. 2007, 13, 463–469. [Google Scholar] [CrossRef]
- Pilsczek, F.H.; Salina, D.; Poon, K.K.H.; Fahey, C.; Yipp, B.G.; Sibley, C.D.; Robbins, S.M.; Green, F.H.Y.; Surette, M.G.; Sugai, M.; et al. A Novel Mechanism of Rapid Nuclear Neutrophil Extracellular Trap Formation in Response to Staphylococcus aureus. J. Immunol. 2010, 185, 7413–7425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yipp, B.G.; Petri, B.; Salina, D.; Jenne, C.N.; Scott, B.N.V.; Zbytnuik, L.D.; Pittman, K.; Asaduzzaman, M.; Wu, K.; Meijndert, H.C.; et al. Infection-induced NETosis is a dynamic process involving neutrophil multitasking in vivo. Nat. Med. 2012, 18, 1386–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trowald-Wigh, G.; Håkansson, L.; Johannisson, A.; Edqvist, L.E. The effect of prednisolone on canine neutrophil function: In vivo and in vitro studies. Acta Vet. Scand. 1998, 39, 201–213. [Google Scholar] [CrossRef]
- Shimamura, S.; Kanayama, K.; Shimada, T.; Maeda, K.; Nakao, R.; Kobayashi, S.; Sato, R.; Okano, S. Evaluation of the function of polymorphonuclear neutrophilic leukocytes in healthy dogs given a high dose of methylprednisolone sodium succinate. Am. J. Vet. Res. 2010, 71, 541–546. [Google Scholar] [CrossRef]
- Dandona, P.; Suri, M.; Hamouda, W.; Aljada, A.; Kumbkarni, Y.; Thusu, K. Hydrocortisone-induced inhibition of reactive oxygen species by polymorphonuclear neutrophils. Crit. Care Med. 1999, 27, 2442–2444. [Google Scholar] [CrossRef]
- Sanner, B.M.; Meder, U.; Zidek, W.; Tepel, M. Effects of glucocorticoids on generation of reactive oxygen species in platelets. Steroids 2002, 67, 715–719. [Google Scholar] [CrossRef]
- Wang, T.; Han, C.; Tian, P.; Li, P.-F.; Ma, X.-L. Role of Teriparatide in Glucocorticoid-induced Osteoporosis through Regulating Cellular Reactive Oxygen Species. Orthop. Surg. 2018, 10, 152–159. [Google Scholar] [CrossRef] [Green Version]
- de Kruif, M.D.; Lemaire, L.C.; Giebelen, I.A.; van Zoelen, M.A.D.; Pater, J.M.; van den Pangaart, P.S.; Groot, A.P.; de Vos, A.F.; Elliott, P.J.; Meijers, J.C.M.; et al. Prednisolone dose-dependently influences inflammation and coagulation during human endotoxemia. J. Immunol. 2007, 178, 1845–1851. [Google Scholar] [CrossRef]
- Dandrieux, J.R.S.; Narayanan, L.; Firestone, S.; Archer, T.M.; Mansfield, C.S. Effect of immunosuppressive drugs on cytokine production in canine whole blood stimulated with lipopolysaccharide or a combination of ionomycin and phorbol 12-myristate 13-acetate. Vet. Med. Sci. 2019, 5, 199–205. [Google Scholar] [CrossRef] [PubMed]
- De Vries, F.; Leuschner, J.; Jilma, B.; Derhaschnig, U. Establishment of a low dose canine endotoxemia model to test anti-inflammatory drugs: Effects of prednisolone. Int. J. Immunopathol. Pharmacol. 2013, 26, 861–869. [Google Scholar] [CrossRef]
- Asahina, R.; Ueda, K.; Oshima, Y.; Kanei, T.; Kato, M.; Furue, M.; Tsukui, T.; Nagata, M.; Maeda, S. Serum canine thymus and activation-regulated chemokine (TARC/CCL17) concentrations correlate with disease severity and therapeutic responses in dogs with atopic dermatitis. Vet. Dermatol. 2020, 31, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, M.C.; Fingerhut, L.; Alfonso-Castro, A.; Mergani, A.; Schwennen, C.; von Köckritz-Blickwede, M.; de Buhr, N. How Long Does a Neutrophil Live?—The Effect of 24 h Whole Blood Storage on Neutrophil Functions in Pigs. Biomedicines 2020, 8, 278. [Google Scholar] [CrossRef] [PubMed]
- de Buhr, N.; Bonilla, M.C.; Pfeiffer, J.; Akhdar, S.; Schwennen, C.; Kahl, B.C.; Waldmann, K.; Valentin-Weigand, P.; Hennig-Pauka, I.; von Köckritz-Blickwede, M. Degraded neutrophil extracellular traps promote the growth of Actinobacillus pleuropneumoniae. Cell Death Dis. 2019, 10, 657. [Google Scholar] [CrossRef]
- de Buhr, N.; Reuner, F.; Neumann, A.; Stump-Guthier, C.; Tenenbaum, T.; Schroten, H.; Ishikawa, H.; Müller, K.; Beineke, A.; Hennig-Pauka, I.; et al. Neutrophil extracellular trap formation in the Streptococcus suis-infected cerebrospinal fluid compartment. Cell. Microbiol. 2017, 19, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beineke, A.; Bennecke, K.; Neis, C.; Schröder, C.; Waldmann, K.-H.; Baumgärtner, W.; Valentin-Weigand, P.; Baums, C.G. Comparative evaluation of virulence and pathology of Streptococcus suis serotypes 2 and 9 in experimentally infected growers. Vet. Microbiol. 2008, 128, 423–430. [Google Scholar] [CrossRef]
- Recovery Collaborative Group; Horby, P.; Lim, W.S.; Emberson, J.R.; Mafham, M.; Bell, J.L.; Linsell, L.; Staplin, N.; Brightling, C.; Ustianowski, A.; et al. Dexamethasone in Hospitalized Patients with Covid-19. N. Engl. J. Med. 2021, 384, 693–704. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steffensen, N.; Imker, R.; Lassnig, S.; Fulde, M.; Rieder, J.C.; de Buhr, N. Methylprednisolone Induces Extracellular Trap Formation and Enhances Bactericidal Effect of Canine Neutrophils. Int. J. Mol. Sci. 2021, 22, 7734. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147734
Steffensen N, Imker R, Lassnig S, Fulde M, Rieder JC, de Buhr N. Methylprednisolone Induces Extracellular Trap Formation and Enhances Bactericidal Effect of Canine Neutrophils. International Journal of Molecular Sciences. 2021; 22(14):7734. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147734
Chicago/Turabian StyleSteffensen, Nicole, Rabea Imker, Simon Lassnig, Marcus Fulde, Johanna C. Rieder, and Nicole de Buhr. 2021. "Methylprednisolone Induces Extracellular Trap Formation and Enhances Bactericidal Effect of Canine Neutrophils" International Journal of Molecular Sciences 22, no. 14: 7734. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147734