
Staphylococcus aureus Decreases SUMOylation Host Response to Promote Intramacrophage Survival

 , , and
, , and 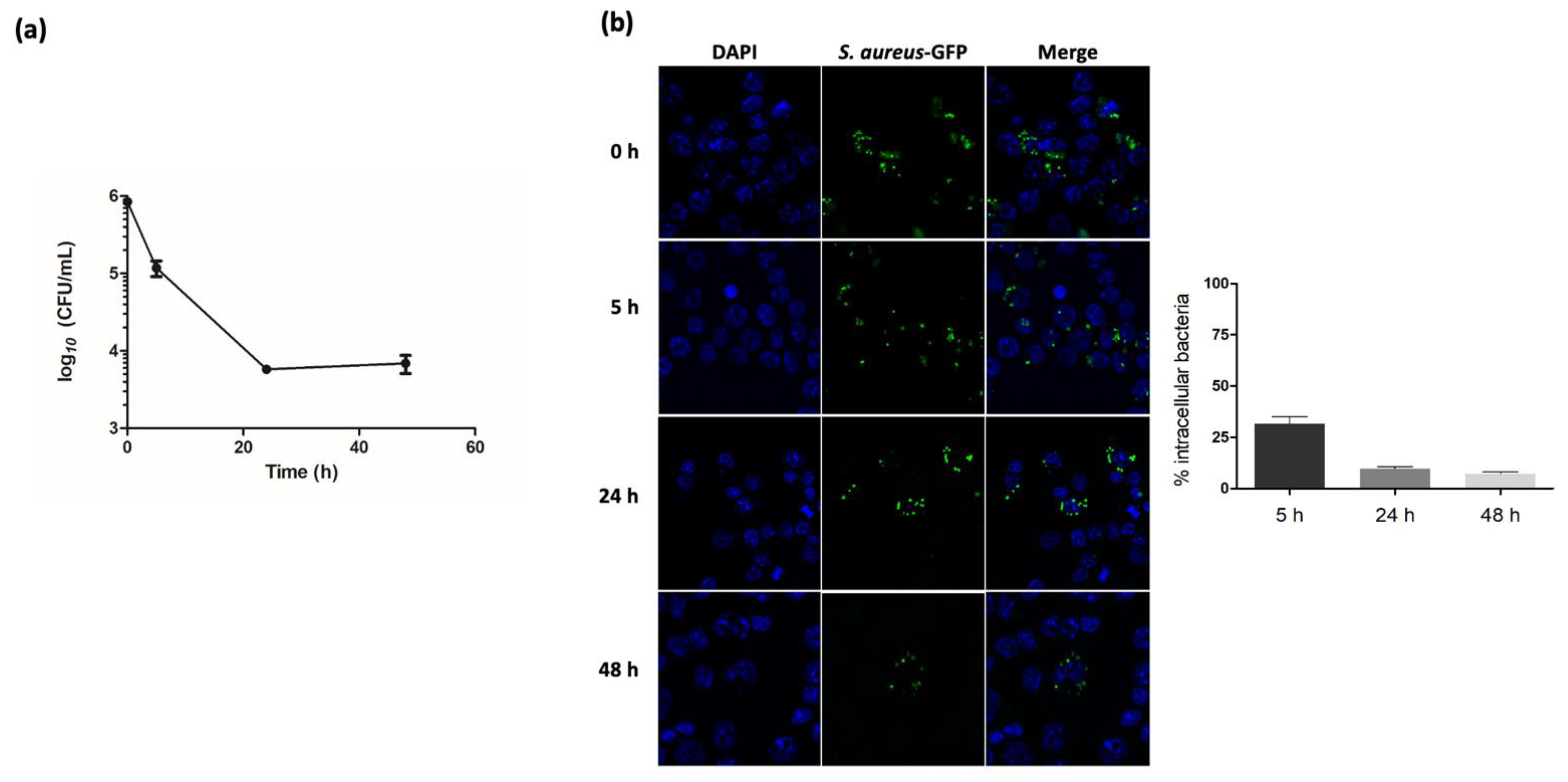{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
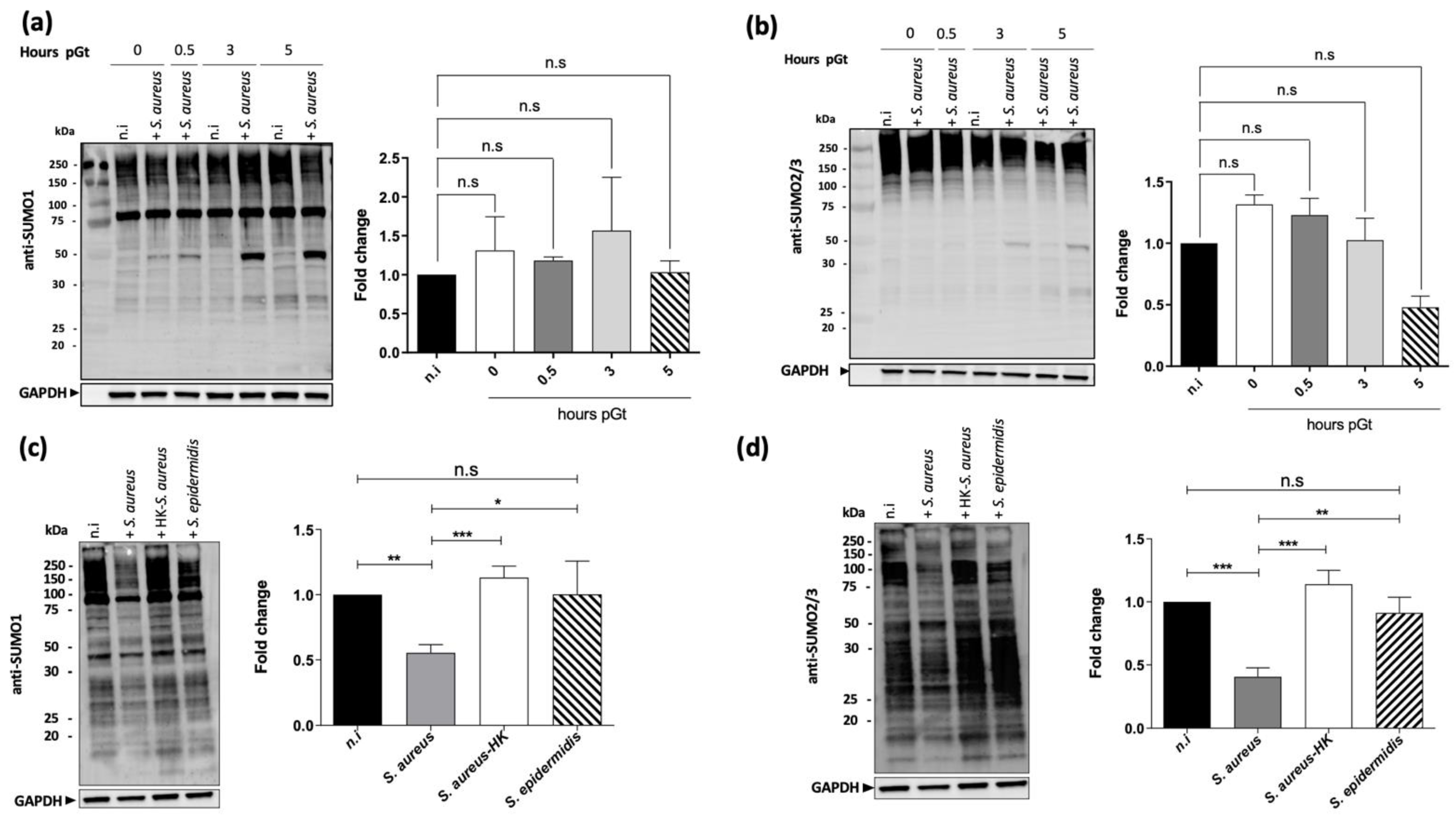
2.1. S. aureus Infection Decreases Macrophages SUMOylated Proteins
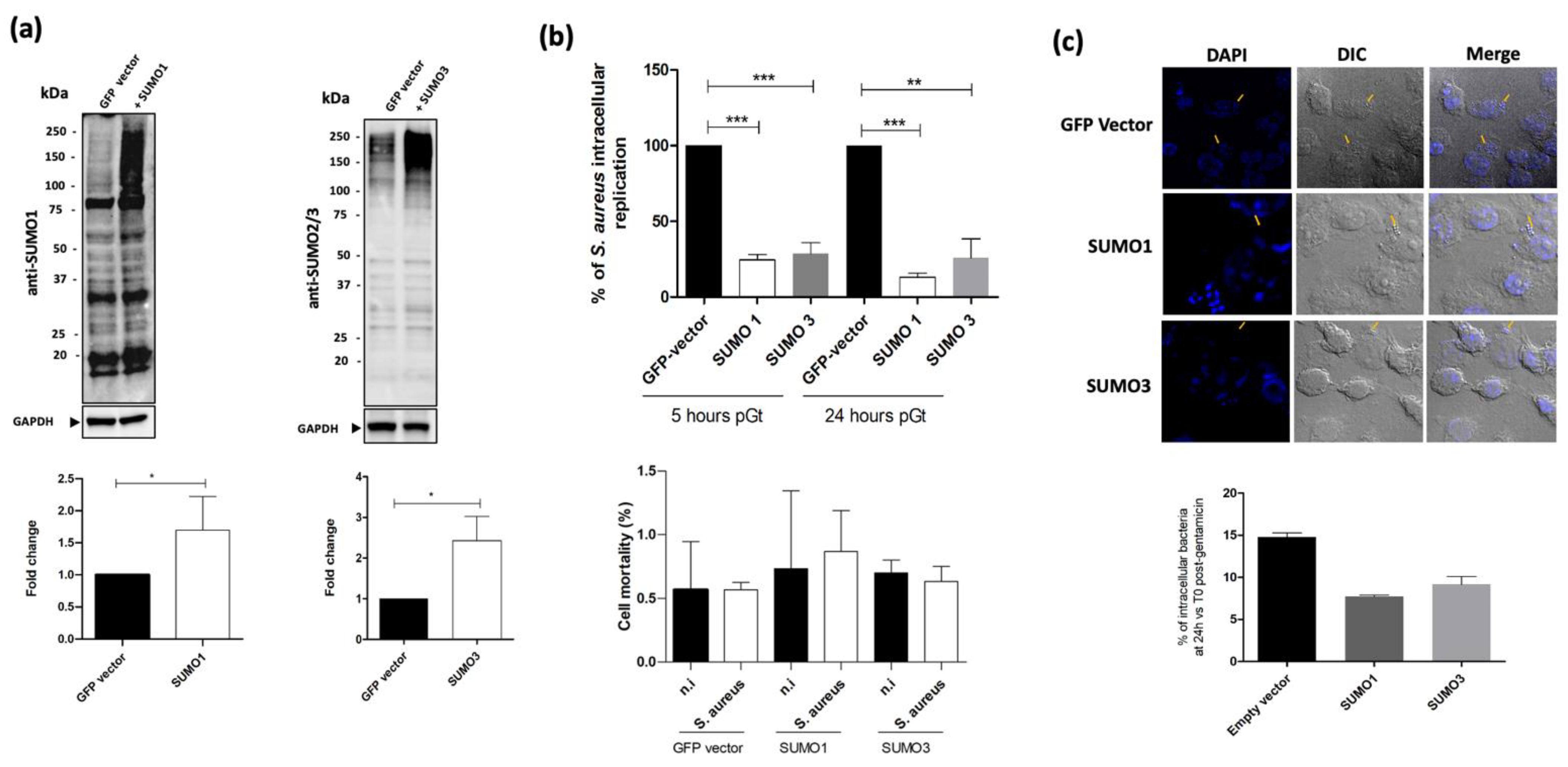
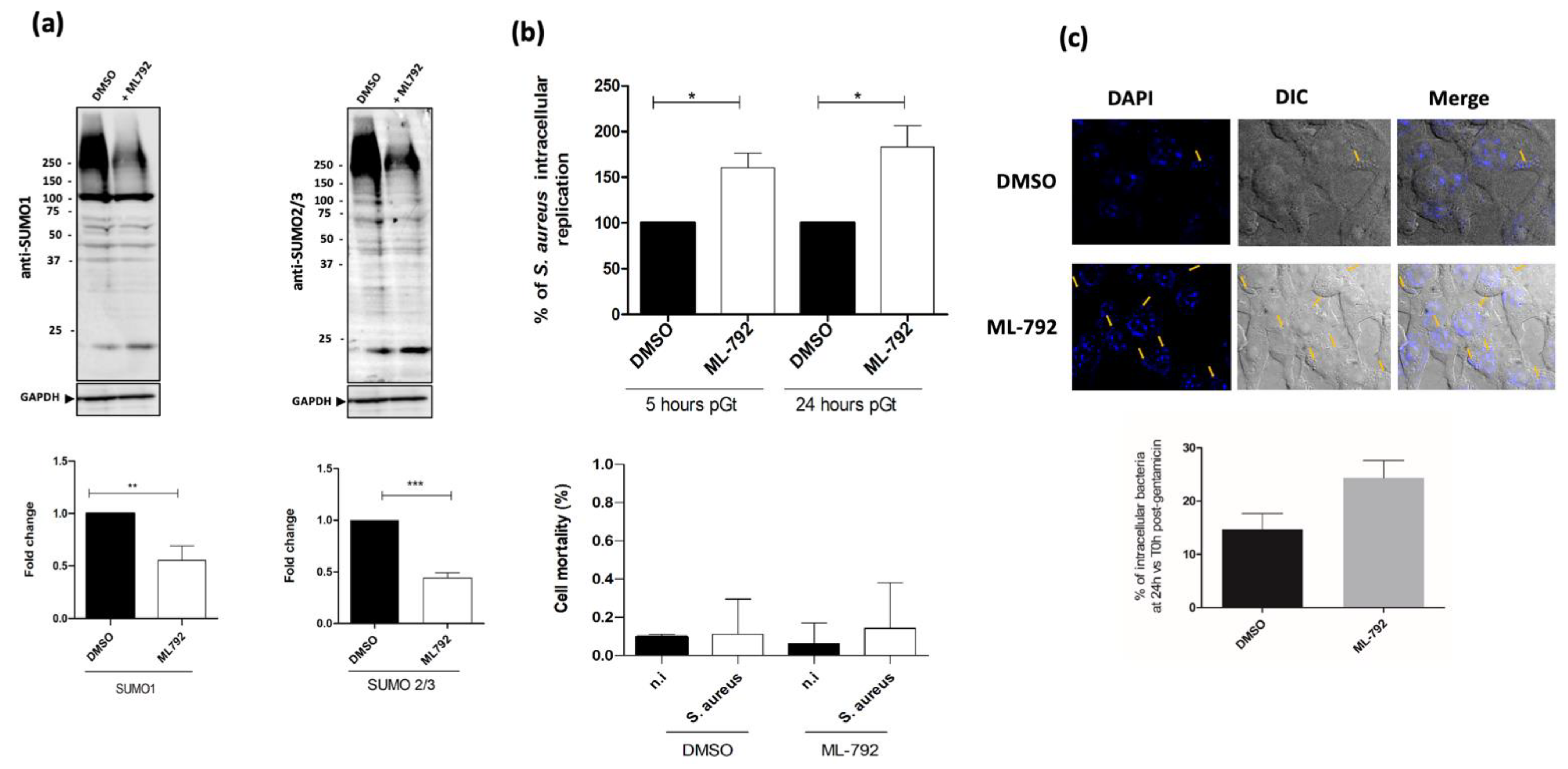
2.2. Host SUMOylation Diminishes S. aureus Intramacrophagic Replication
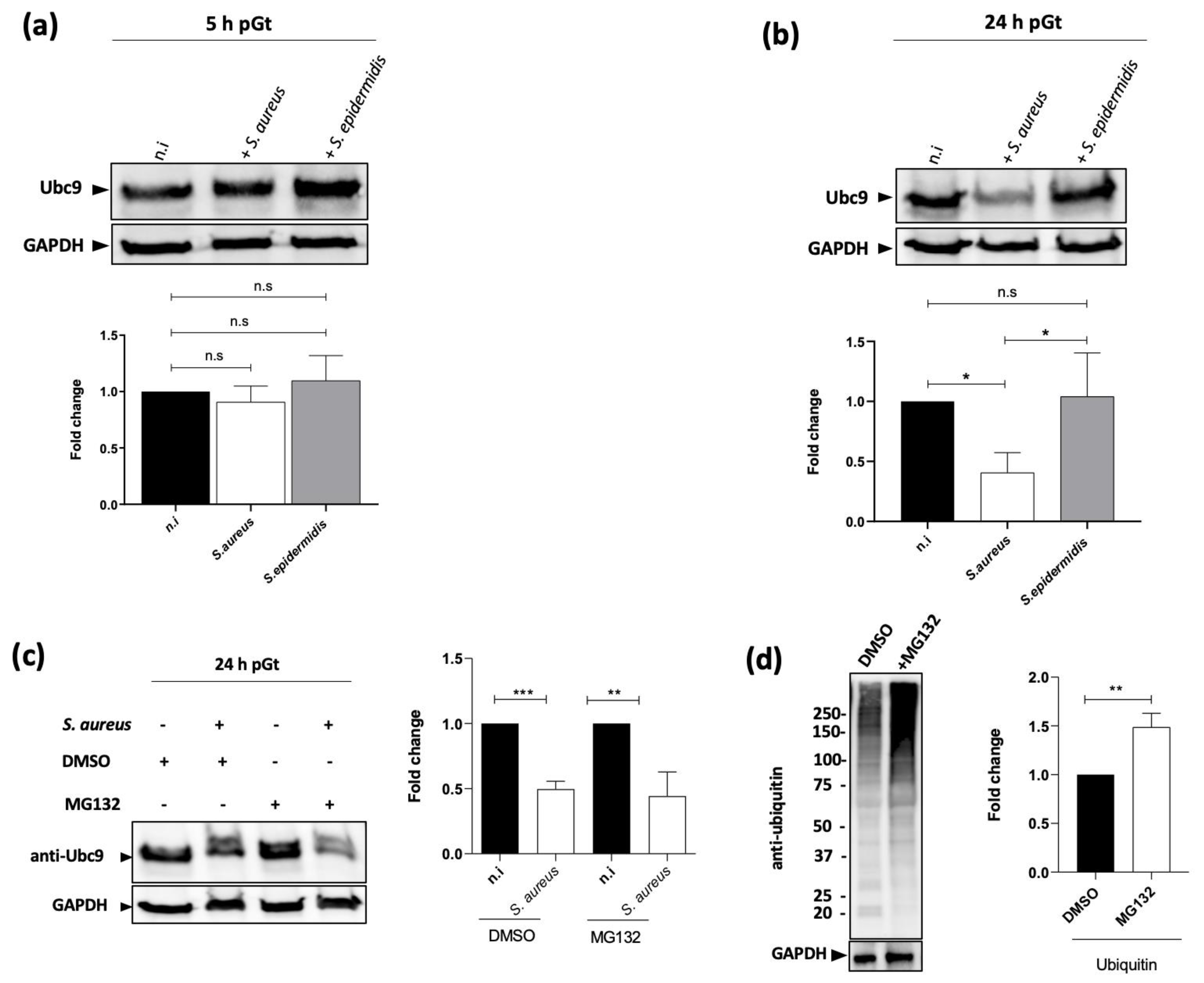
2.3. The Level of the SUMOylation Enzyme Ubc9 Is Reduced upon S. aureus Infection
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Macrophages Culture and Infection
4.3. Fluorescent Microscopy
4.4. Immunoblotting
4.5. Cell Viability
4.6. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Archer, G.L. Staphylococcus aureus: A Well–Armed Pathogen. Clin. Infect. Dis. 1998, 26, 1179–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowy, F.D. Staphylococcus aureus Infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef]
- Garzoni, C.; Kelley, W.L. Staphylococcus aureus: New Evidence for Intracellular Persistence. Trends Microbiol. 2009, 17, 59–65. [Google Scholar] [CrossRef]
- Kubica, M.; Guzik, K.; Koziel, J.; Zarebski, M.; Richter, W.; Gajkowska, B.; Golda, A.; Maciag-Gudowska, A.; Brix, K.; Shaw, L.; et al. A Potential New Pathway for Staphylococcus aureus Dissemination: The Silent Survival of S. aureus Phagocytosed by Human Monocyte-Derived Macrophages. PLoS ONE 2008, 3, e1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacoma, A.; Cano, V.; Moranta, D.; Regueiro, V.; Domínguez-Villanueva, D.; Laabei, M.; González-Nicolau, M.; Ausina, V.; Prat, C.; Bengoechea, J.A. Investigating Intracellular Persistence of Staphylococcus aureus within a Murine Alveolar Macrophage Cell Line. Virulence 2017, 8, 1761–1775. [Google Scholar] [CrossRef] [Green Version]
- Alto, N.M.; Orth, K. Subversion of Cell Signaling by Pathogens. Cold Spring Harb. Perspect. Biol. 2012, 4, a006114. [Google Scholar] [CrossRef] [PubMed]
- Hamon, M.A.; Cossart, P. Histone Modifications and Chromatin Remodeling during Bacterial Infections. Cell Host Microbe 2008, 4, 100–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribet, D.; Cossart, P. Ubiquitin, SUMO, and NEDD8: Key Targets of Bacterial Pathogens. Trends Cell Biol. 2018, 28, 926–940. [Google Scholar] [CrossRef] [PubMed]
- Ribet, D.; Cossart, P. Post-Translational Modifications in Host Cells during Bacterial Infection. FEBS Lett. 2010, 584, 2748–2758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribet, D.; Cossart, P. Pathogen-Mediated Posttranslational Modifications: A Re-Emerging Field. Cell 2010, 143, 694–702. [Google Scholar] [CrossRef] [Green Version]
- Rytkönen, A.; Holden, D.W. Bacterial Interference of Ubiquitination and Deubiquitination. Cell Host Microbe 2007, 1, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalmasso, G.; Nguyen, H.T.T.; Faïs, T.; Massier, S.; Barnich, N.; Delmas, J.; Bonnet, R. Crohn’s Disease-Associated Adherent-Invasive Escherichia coli Manipulate Host Autophagy by Impairing SUMOylation. Cells 2019, 8, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritah, S.; Lhocine, N.; Golebiowski, F.; Mounier, J.; Andrieux, A.; Jouvion, G.; Hay, R.T.; Sansonetti, P.; Dejean, A. Sumoylation Controls Host Anti-Bacterial Response to the Gut Invasive Pathogen Shigella flexneri. EMBO Rep. 2014, 15, 965–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribet, D.; Hamon, M.; Gouin, E.; Nahori, M.-A.; Impens, F.; Neyret-Kahn, H.; Gevaert, K.; Vandekerckhove, J.; Dejean, A.; Cossart, P. Listeria monocytogenes Impairs SUMOylation for Efficient Infection. Nature 2010, 464, 1192–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sá-Pessoa, J.; Przybyszewska, K.; Vasconcelos, F.N.; Dumigan, A.; Frank, C.G.; Hobley, L.; Bengoechea, J.A. Klebsiella Pneumoniae Reduces SUMOylation To Limit Host Defense Responses. mBio 2020, 11, e01733-20. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Mohapatra, G.; Ahmad, S.M.; Rana, S.; Jain, S.; Khalsa, J.K.; Srikanth, C.V. Salmonella Engages Host MicroRNAs To Modulate SUMOylation: A New Arsenal for Intracellular Survival. Mol. Cell. Biol. 2015, 35, 2932–2946. [Google Scholar] [CrossRef] [Green Version]
- Cubeñas-Potts, C.; Matunis, M.J. SUMO: A Multifaceted Modifier of Chromatin Structure and Function. Dev. Cell 2013, 24, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Flotho, A.; Melchior, F. Sumoylation: A Regulatory Protein Modification in Health and Disease. Annu. Rev. Biochem. 2013, 82, 357–385. [Google Scholar] [CrossRef]
- Boulanger, M.; Paolillo, R.; Piechaczyk, M.; Bossis, G. The SUMO Pathway in Hematomalignancies and Their Response to Therapies. Int. J. Mol. Sci. 2019, 20, 3895. [Google Scholar] [CrossRef] [Green Version]
- Celen, A.B.; Sahin, U. Sumoylation on Its 25th Anniversary: Mechanisms, Pathology, and Emerging Concepts. FEBS J. 2020, 287, 3110–3140. [Google Scholar] [CrossRef]
- Deplanche, M.; Mouhali, N.; Nguyen, M.-T.; Cauty, C.; Ezan, F.; Diot, A.; Raulin, L.; Dutertre, S.; Langouet, S.; Legembre, P.; et al. Staphylococcus aureus Induces DNA Damage in Host Cell. Sci. Rep. 2019, 9, 7694. [Google Scholar] [CrossRef] [PubMed]
- Gannoun-Zaki, L.; Pätzold, L.; Huc-Brandt, S.; Baronian, G.; Elhawy, M.I.; Gaupp, R.; Martin, M.; Blanc-Potard, A.-B.; Letourneur, F.; Bischoff, M.; et al. PtpA, a Secreted Tyrosine Phosphatase from Staphylococcus aureus, Contributes to Virulence and Interacts with Coronin-1A during Infection. J. Biol. Chem. 2018, 293, 15569–15580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, Y.; Bruns, S.A.; Rohde, M.; Prajsnar, T.K.; Foster, S.J.; Schmitz, I. Intracellular Staphylococcus aureus Eludes Selective Autophagy by Activating a Host Cell Kinase. Autophagy 2016, 12, 2069–2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonacci, T.; Audebert, S.; Camoin, L.; Baudelet, E.; Bidaut, G.; Garcia, M.; Witzel, I.-I.; Perkins, N.D.; Borg, J.-P.; Iovanna, J.-L.; et al. Identification of New Mechanisms of Cellular Response to Chemotherapy by Tracking Changes in Post-Translational Modifications by Ubiquitin and Ubiquitin-Like Proteins. J. Proteome Res. 2014, 13, 2478–2494. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Riceberg, J.; Soucy, T.; Koenig, E.; Minissale, J.; Gallery, M.; Bernard, H.; Yang, X.; Liao, H.; Rabino, C.; et al. Probing the Roles of SUMOylation in Cancer Cell Biology by Using a Selective SAE Inhibitor. Nat. Chem. Biol. 2017, 13, 1164–1171. [Google Scholar] [CrossRef] [PubMed]
- Everett, R.D.; Boutell, C.; Hale, B.G. Interplay between Viruses and Host Sumoylation Pathways. Nat. Rev. Microbiol. 2013, 11, 400–411. [Google Scholar] [CrossRef] [PubMed]
- Mattoscio, D.; Medda, A.; Chiocca, S. Recent Highlights: Onco Viral Exploitation of the SUMO System. Curr. Issues Mol. Biol. 2020, 35, 16. [Google Scholar] [CrossRef] [Green Version]
- Sidik, S.M.; Salsman, J.; Dellaire, G.; Rohde, J.R. Shigella Infection Interferes with SUMOylation and Increases PML-NB Number. PLoS ONE 2015, 10, e0122585. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-G.; Stork, W.; Mudgett, M.B. Xanthomonas Type III Effector XopD Desumoylates Tomato Transcription Factor SlERF4 to Suppress Ethylene Responses and Promote Pathogen Growth. Cell Host Microbe 2013, 13, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Orth, K. Disruption of Signaling by Yersinia Effector YopJ, a Ubiquitin-Like Protein Protease. Science 2000, 290, 1594–1597. [Google Scholar] [CrossRef]
- Messad, N.; Prajsnar, T.K.; Lina, G.; O’Callaghan, D.; Foster, S.J.; Renshaw, S.A.; Skaar, E.P.; Bes, M.; Dunyach-Remy, C.; Vandenesch, F.; et al. Existence of a Colonizing Staphylococcus aureus Strain Isolated in Diabetic Foot Ulcers. Diabetes 2015, 64, 2991–2995. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youssouf, N.; Recasens-Zorzo, C.; Molle, V.; Bossis, G.; Soubeyran, P.; Gannoun-Zaki, L. Staphylococcus aureus Decreases SUMOylation Host Response to Promote Intramacrophage Survival. Int. J. Mol. Sci. 2021, 22, 8108. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158108
Youssouf N, Recasens-Zorzo C, Molle V, Bossis G, Soubeyran P, Gannoun-Zaki L. Staphylococcus aureus Decreases SUMOylation Host Response to Promote Intramacrophage Survival. International Journal of Molecular Sciences. 2021; 22(15):8108. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158108
Chicago/Turabian StyleYoussouf, Nadhuma, Clara Recasens-Zorzo, Virginie Molle, Guillaume Bossis, Philippe Soubeyran, and Laila Gannoun-Zaki. 2021. "Staphylococcus aureus Decreases SUMOylation Host Response to Promote Intramacrophage Survival" International Journal of Molecular Sciences 22, no. 15: 8108. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158108