
The Pharmacological Profile of Second Generation Pyrovalerone Cathinones and Related Cathinone Derivative


Abstract
:1. Introduction
2. Results
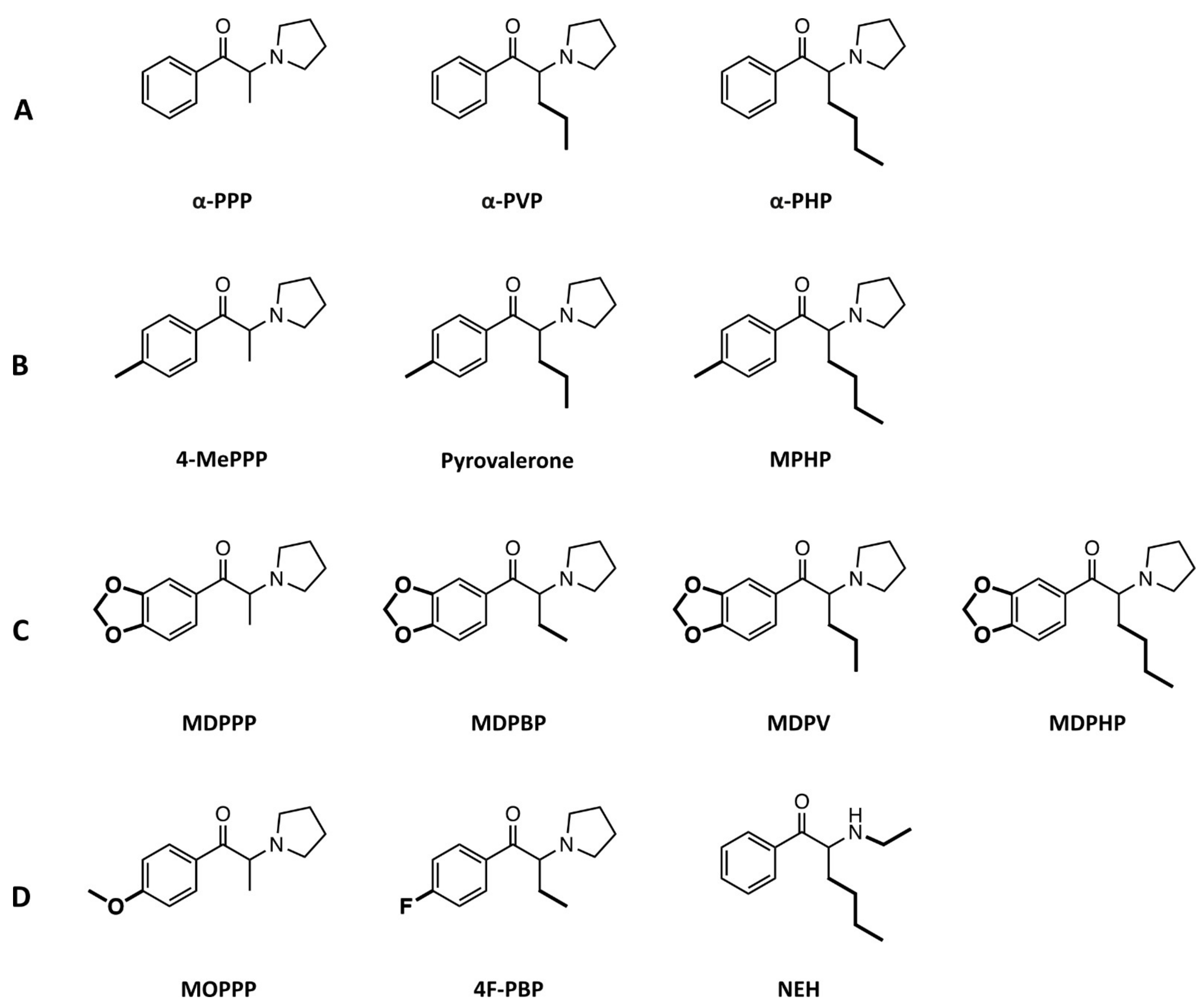
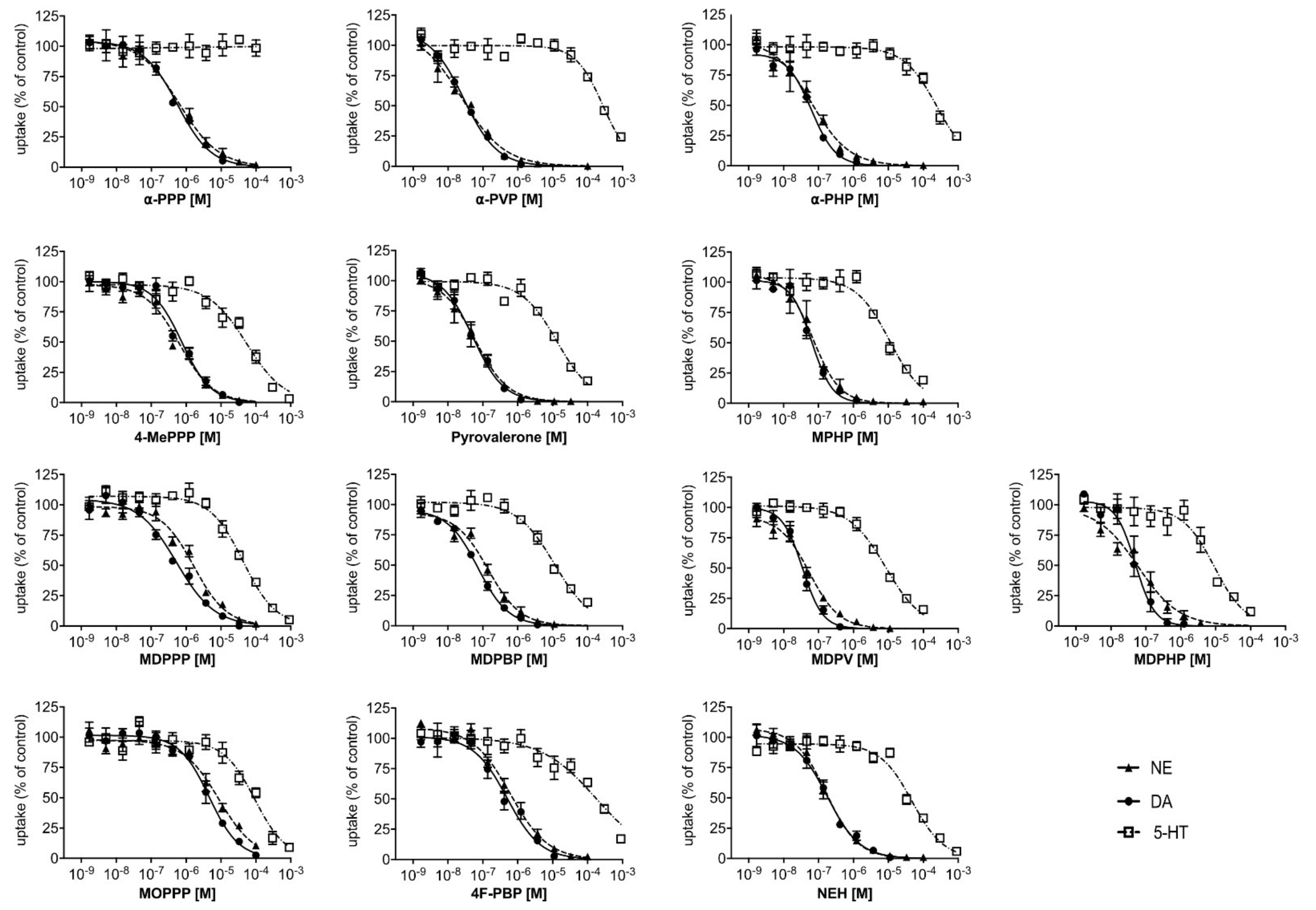
2.1. Monoamine Uptake Transporter Inhibition
2.2. Serotonergic Receptor Interactions: Binding Affinity and Activation Potency
2.3. Monoamine Transporter and Non-Serotonergic Receptor Binding Interactions
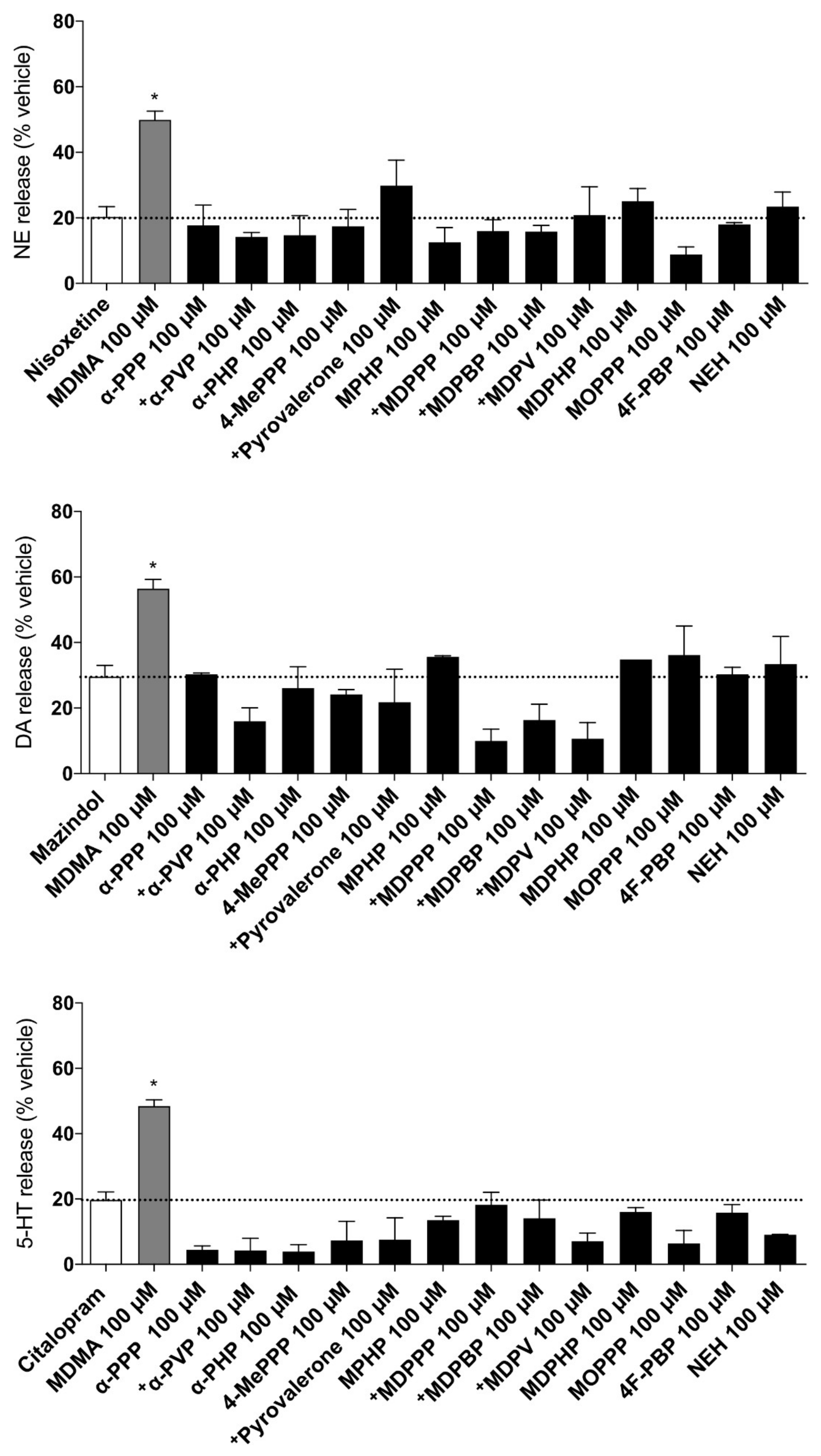
2.4. Transporter-Mediated Monoamine Efflux
3. Discussion
3.1. Monoamine Transporter Inhibition and Transporter-Mediated Efflux
3.2. Serotonergic Receptor Binding and Activation Interactions
3.3. Monoamine Transporter and Non-Serotonergic Receptor Binding Interactions
4. Materials and Methods
4.1. Compounds
4.2. Monoamine Uptake Transporter Inhibition
4.3. Monoamine Efflux Mediated by the Transporters
4.4. Radioligand Binding, Activation Potency, and Efficacy at the 5-HT Receptors
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zawilska, J.B.; Wojcieszak, J. Designer cathinones-an emerging class of novel recreational drugs. Forensic Sci. Int. 2013, 231, 42–53. [Google Scholar] [CrossRef]
- German, C.L.; Fleckenstein, A.E.; Hanson, G.R. Bath salts and synthetic cathinones: An emerging designer drug phenomenon. Life Sci. 2014, 97, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Baumann, M.H.; Solis, E., Jr.; Watterson, L.R.; Marusich, J.A.; Fantegrossi, W.E.; Wiley, J.L. Baths salts, spice, and related designer drugs: The science behind the headlines. J. Neurosci. 2014, 34, 15150–15158. [Google Scholar] [CrossRef] [Green Version]
- Winstock, A.R.; Mitcheson, L.R.; Deluca, P.; Davey, Z.; Corazza, O.; Schifano, F. Mephedrone, new kid for the chop? Addiction 2011, 106, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Brandt, S.D.; King, L.A.; Evans-Brown, M. The new drug phenomenon. Drug Test. Anal. 2014, 6, 587–597. [Google Scholar] [CrossRef]
- Spiller, H.A.; Ryan, M.L.; Weston, R.G.; Jansen, J. Clinical experience with and analytical confirmation of “bath salts” and “legal highs” (synthetic cathinones) in the United States. Clin. Toxicol. 2011, 49, 499–505. [Google Scholar] [CrossRef] [PubMed]
- James, D.; Adams, R.D.; Spears, R.; Cooper, G.; Lupton, D.J.; Thompson, J.P.; Thomas, S.H. Clinical characteristics of mephedrone toxicity reported to the U.K. National Poisons Information Service. Emerg. Med. J. 2011, 28, 686–689. [Google Scholar] [CrossRef]
- Ross, E.A.; Watson, M.; Goldberger, B. “Bath salts” intoxication. N. Engl. J. Med. 2011, 365, 967–968. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.P. Cathinone derivatives: A review of their chemistry, pharmacology and toxicology. Drug Test. Anal. 2011, 3, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Baumann, M.H.; Walters, H.M.; Niello, M.; Sitte, H.H. Neuropharmacology of synthetic cathinones. In New Psychoactive Substances; Springer: Cham, Switzerland, 2018; pp. 113–142. [Google Scholar]
- Eshleman, A.J.; Wolfrum, K.M.; Reed, J.F.; Kim, S.O.; Swanson, T.; Johnson, R.A.; Janowsky, A. Structure-activity relationships of substituted cathinones, with transporter binding, uptake, and release. J. Pharmacol. Exp. Ther. 2017, 360, 33–47. [Google Scholar] [CrossRef]
- Marusich, J.A.; Antonazzo, K.R.; Wiley, J.L.; Blough, B.E.; Partilla, J.S.; Baumann, M.H. Pharmacology of novel synthetic stimulants structurally related to the “bath salts” constituent 3,4-methylenedioxypyrovalerone (MDPV). Neuropharmacology 2014, 87, 206–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rickli, A.; Hoener, M.C.; Liechti, M.E. Monoamine transporter and receptor interaction profiles of novel psychoactive substances: Para-halogenated amphetamines and pyrovalerone cathinones. Eur. Neuropsychopharmacol. 2015, 25, 365–376. [Google Scholar] [CrossRef] [Green Version]
- EMCDDA. Report of the Risk Assessment of MDPV in the Framework of the Council Decision on New Psychoactive Substances; EMCDDA: Lisbon, Portugal, 2014. [Google Scholar]
- Helander, A.; Bäckberg, M.; Hultén, P.; Al-Saffar, Y.; Beck, O. Detection of new psychoactive substance use among emergency room patients: Results from the Swedish STRIDA project. Forensic Sci. Int. 2014, 243, 23–29. [Google Scholar] [CrossRef]
- Zuba, D.; Byrska, B. Prevalence and co-existence of active components of ‘legal highs’. Drug Test. Anal. 2013, 5, 420–429. [Google Scholar] [CrossRef]
- Baumann, M.H.; Partilla, J.S.; Lehner, K.R.; Thorndike, E.B.; Hoffman, A.F.; Holy, M.; Rothman, R.B.; Goldberg, S.R.; Lupica, C.R.; Sitte, H.H.; et al. Powerful cocaine-like actions of 3,4-methylenedioxypyrovalerone (MDPV), a principal constituent of psychoactive ‘bath salts’ products. Neuropsychopharmacology 2013, 38, 552–562. [Google Scholar] [CrossRef] [Green Version]
- Simmler, L.D.; Buser, T.A.; Donzelli, M.; Schramm, Y.; Dieu, L.H.; Huwyler, J.; Chaboz, S.; Hoener, M.C.; Liechti, M.E. Pharmacological characterization of designer cathinones in vitro. Br. J. Pharmacol. 2013, 168, 458–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, S.D.; Freeman, S.; Sumnall, H.R.; Measham, F.; Cole, J. Analysis of NRG ‘legal highs’ in the UK: Identification and formation of novel cathinones. Drug Test. Anal. 2011, 3, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Elliott, S.; Evans, J. A 3-year review of new psychoactive substances in casework. Forensic Sci. Int. 2014, 243, 55–60. [Google Scholar] [CrossRef]
- Marinetti, L.J.; Antonides, H.M. Analysis of synthetic cathinones commonly found in bath salts in human performance and postmortem toxicology: Method development, drug distribution and interpretation of results. J. Anal. Toxicol. 2013, 37, 135–146. [Google Scholar] [CrossRef]
- Shanks, K.G.; Dahn, T.; Behonick, G.; Terrell, A. Analysis of first and second generation legal highs for synthetic cannabinoids and synthetic stimulants by ultra-performance liquid chromatography and time of flight mass spectrometry. J. Anal. Toxicol. 2012, 36, 360–371. [Google Scholar] [CrossRef] [Green Version]
- Westphal, F.; Junge, T.; Rösner, P.; Fritschi, G.; Klein, B.; Girreser, U. Mass spectral and NMR spectral data of two new designer drugs with an alpha-aminophenone structure: 4′-methyl-alpha-pyrrolidinohexanophenone and 4′-methyl-alpha-pyrrolidinobutyrophenone. Forensic Sci. Int. 2007, 169, 32–42. [Google Scholar] [CrossRef]
- Eiden, C.; Mathieu, O.; Cathala, P.; Debruyne, D.; Baccino, E.; Petit, P.; Peyriere, H. Toxicity and death following recreational use of 2-pyrrolidino valerophenone. Clin. Toxicol. 2013, 51, 899–903. [Google Scholar] [CrossRef] [PubMed]
- Luethi, D.; Liechti, M.E. Monoamine transporter and receptor interaction profiles in vitro predict reported human doses of novel psychoactive stimulants and psychedelics. Int. J. Neuropsychopharmacol. 2018, 21, 926–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudin, D.; Liechti, M.E.; Luethi, D. Molecular and clinical aspects of potential neurotoxicity induced by new psychoactive stimulants and psychedelics. Exp. Neurol. 2021, 343, 113778. [Google Scholar] [CrossRef] [PubMed]
- Luethi, D.; Liechti, M.E. Designer drugs: Mechanism of action and adverse effects. Arch. Toxicol. 2020, 94, 1085–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Jia, W.; Li, T.; Hua, Z.; Qian, Z. Identification and analytical characterization of nine synthetic cathinone derivatives N-ethylhexedrone, 4-Cl-pentedrone, 4-Cl-α-EAPP, propylone, N-ethylnorpentylone, 6-MeO-bk-MDMA, α-PiHP, 4-Cl-α-PHP, and 4-F-α-PHP. Drug Test. Anal. 2017, 9, 1162–1171. [Google Scholar] [CrossRef]
- Simmler, L.D.; Buchy, D.; Chaboz, S.; Hoener, M.C.; Liechti, M.E. In vitro characterization of psychoactive substances at rat, mouse, and human trace amine-associated receptor 1. J. Pharmacol. Exp. Ther. 2016, 357, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Luethi, D.; Kolaczynska, K.E.; Walter, M.; Suzuki, M.; Rice, K.C.; Blough, B.E.; Hoener, M.C.; Baumann, M.H.; Liechti, M.E. Metabolites of the ring-substituted stimulants MDMA, methylone and MDPV differentially affect human monoaminergic systems. J. Psychopharmacol. 2019, 33, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Rothman, R.B.; Baumann, M.H.; Dersch, C.M.; Romero, D.V.; Rice, K.C.; Carroll, F.I.; Partilla, J.S. Amphetamine-type central nervous system stimulants release norepinephrine more potently than they release dopamine and serotonin. Synapse 2001, 39, 32–41. [Google Scholar] [CrossRef]
- Baumann, M.H.; Ayestas, M.A.; Partilla, J.S.; Sink, J.R.; Shulgin, A.T.; Daley, P.F.; Brandt, S.D.; Rothman, R.B.; Ruoho, A.E.; Cozzi, N.V. The designer methcathinone analogs, mephedrone and methylone, are substrates for monoamine transporters in brain tissue. Neuropsychopharmacology 2012, 37, 1192–1203. [Google Scholar] [CrossRef] [PubMed]
- Gatch, M.B.; Dolan, S.B.; Forster, M.J. Locomotor activity and discriminative stimulus effects of a novel series of synthetic cathinone analogs in mice and rats. Psychopharmacology 2017, 234, 1237–1245. [Google Scholar] [CrossRef] [Green Version]
- Eshleman, A.J.; Nagarajan, S.; Wolfrum, K.M.; Reed, J.F.; Swanson, T.L.; Nilsen, A.; Janowsky, A. Structure-activity relationships of bath salt components: Substituted cathinones and benzofurans at biogenic amine transporters. Psychopharmacology 2019, 236, 939–952. [Google Scholar] [CrossRef]
- Saha, K.; Partilla, J.S.; Lehner, K.R.; Seddik, A.; Stockner, T.; Holy, M.; Sandtner, W.; Ecker, G.F.; Sitte, H.H.; Baumann, M.H. ‘Second-generation’ mephedrone analogs, 4-MEC and 4-MePPP, differentially affect monoamine transporter function. Neuropsychopharmacology 2015, 40, 1321–1331. [Google Scholar] [CrossRef] [Green Version]
- Baumann, M.H.; Clark, R.D.; Woolverton, W.L.; Wee, S.; Blough, B.E.; Rothman, R.B. In vivo effects of amphetamine analogs reveal evidence for serotonergic inhibition of mesolimbic dopamine transmission in the rat. J. Pharmacol. Exp. Ther. 2011, 337, 218–225. [Google Scholar] [CrossRef]
- Gatch, M.B.; Dolan, S.B.; Forster, M.J. Comparative behavioral pharmacology of three pyrrolidine-containing synthetic cathinone derivatives. J. Pharmacol. Exp. Ther. 2015, 354, 103–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roesner, P.; Junge, T.; Fritschi, G.; Klein, B.; Thielert, K.; Kozlowski, M. Neue synthetische drogen: Piperazin-, Procyclidin-und alpha-Aminopropiophenonderivate. Toxichem. Krimtech. 1999, 66, 81–90. [Google Scholar]
- Springer, D.; Fritschi, G.; Maurer, H.H. Metabolism and toxicological detection of the new designer drug 4′-methoxy-alpha-pyrrolidinopropiophenone studied in rat urine using gas chromatography-mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2003, 793, 331–342. [Google Scholar] [CrossRef]
- Springer, D.; Staack, R.F.; Paul, L.D.; Kraemer, T.; Maurer, H.H. Identification of cytochrome P450 enzymes involved in the metabolism of 4′-methoxy-α-pyrrolidinopropiophenone (MOPPP), a designer drug, in human liver microsomes. Xenobiotica 2003, 33, 989–998. [Google Scholar] [CrossRef]
- Gaspar, H.; Bronze, S.; Ciríaco, S.; Queirós, C.R.; Matias, A.; Rodrigues, J.; Oliveira, C.; Cordeiro, C.; Santos, S. 4F-PBP (4′-fluoro-α-pyrrolidinobutyrophenone), a new substance of abuse: Structural characterization and purity NMR profiling. Forensic Sci. Int. 2015, 252, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Kovács, K.; Kereszty, É.; Berkecz, R.; Tiszlavicz, L.; Sija, É.; Körmöczi, T.; Jenei, N.; Révész-Schmehl, H.; Institóris, L. Fatal intoxication of a regular drug user following N-ethyl-hexedrone and ADB-FUBINACA consumption. J. Forensic Leg. Med. 2019, 65, 92–100. [Google Scholar] [CrossRef]
- Mikołajczyk, A.; Adamowicz, P.; Tokarczyk, B.; Sekuła, K.; Gieroń, J.; Wrzesień, W. Determination of N-ethylhexedrone, a new cathinone derivative, in blood collected from drivers–analysis of three cases. Probl. Forensic Sci. 2017, 109, 53–63. [Google Scholar]
- WHO. WHO Expert Committee on Drug Dependence: Forty-Second Report; WHO: Geneva, Switzerland, 2020. [Google Scholar]
- Madsen, M.K.; Fisher, P.M.; Stenbæk, D.S.; Kristiansen, S.; Burmester, D.; Lehel, S.; Páleníček, T.; Kuchař, M.; Svarer, C.; Ozenne, B.; et al. A single psilocybin dose is associated with long–term increased mindfulness, preceded by a proportional change in neocortical 5-HT2A receptor binding. Eur. Neuropsychopharmacol. 2020, 33, 71–80. [Google Scholar] [CrossRef]
- Nichols, D.E. Psychedelics. Pharmacol. Rev. 2016, 68, 264–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preller, K.H.; Herdener, M.; Pokorny, T.; Planzer, A.; Kraehenmann, R.; Stämpfli, P.; Liechti, M.E.; Seifritz, E.; Vollenweider, F.X. The Fabric of Meaning and Subjective Effects in LSD-Induced States Depend on Serotonin 2A Receptor Activation. Curr. Biol. 2017, 27, 451–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holze, F.; Vizeli, P.; Ley, L.; Muller, F.; Dolder, P.; Stocker, M.; Duthaler, U.; Varghese, N.; Eckert, A.; Borgwardt, S.; et al. Acute dose-dependent effects of lysergic acid diethylamide in a double-blind placebo-controlled study in healthy subjects. Neuropsychopharmacology 2021, 46, 537–544. [Google Scholar] [CrossRef]
- Hysek, C.M.; Schmid, Y.; Simmler, L.D.; Domes, G.; Heinrichs, M.; Eisenegger, C.; Preller, K.H.; Quednow, B.B.; Liechti, M.E. MDMA enhances emotional empathy and prosocial behavior. Soc. Cogn. Affect. Neurosci. 2014, 9, 1645–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holze, F.; Vizeli, P.; Müller, F.; Ley, L.; Duerig, R.; Varghese, N.; Eckert, A.; Borgwardt, S.; Liechti, M.E. Distinct acute effects of LSD, MDMA, and D-amphetamine in healthy subjects. Neuropsychopharmacology 2020, 45, 462–471. [Google Scholar] [CrossRef] [Green Version]
- Rothman, R.B.; Baumann, M.H.; Savage, J.E.; Rauser, L.; McBride, A.; Hufeisen, S.J.; Roth, B.L. Evidence for possible involvement of 5-HT(2B) receptors in the cardiac valvulopathy associated with fenfluramine and other serotonergic medications. Circulation 2000, 102, 2836–2841. [Google Scholar] [CrossRef] [Green Version]
- Doly, S.; Valjent, E.; Setola, V.; Callebert, J.; Hervé, D.; Launay, J.-M.; Maroteaux, L. Seretonim 5-Hit2B receptors are required for 3,4-methylenedioxymethamphetamine-induced hyperlocomtion and 5-HT release In Vivo and In Vitro. J. Neurosci. 2008, 28, 2933–2940. [Google Scholar] [CrossRef] [Green Version]
- Hysek, C.; Schmid, Y.; Rickli, A.; Simmler, L.D.; Donzelli, M.; Grouzmann, E.; Liechti, M.E. Carvedilol inhibits the cardiostimulant and thermogenic effects of MDMA in humans. Br. J. Pharmacol. 2012, 166, 2277–2288. [Google Scholar] [CrossRef] [Green Version]
- Hysek, C.M.; Brugger, R.; Simmler, L.D.; Bruggisser, M.; Donzelli, M.; Grouzmann, E.; Hoener, M.C.; Liechti, M.E. Effects of the α2-adrenergic agonist clonidine on the pharmacodynamics and pharmacokinetics of 3,4-methylenedioxymethamphetamine in healthy volunteers. J. Pharmacol. Exp. Ther. 2012, 340, 286–294. [Google Scholar] [CrossRef] [Green Version]
- Di Cara, B.; Maggio, R.; Aloisi, G.; Rivet, J.-M.; Lundius, E.G.; Yoshitake, T.; Svenningsson, P.; Brocco, M.; Gobert, A.; De Groote, L.; et al. Genetic deletion of trace amine 1 receptors reveals their role in auto-inhibiting the actions of ecstasy (MDMA). J. Neurosci. 2011, 31, 16928–16940. [Google Scholar] [CrossRef]
- Hysek, C.M.; Simmler, L.D.; Nicola, V.G.; Vischer, N.; Donzelli, M.; Krähenbühl, S.; Grouzmann, E.; Huwyler, J.; Hoener, M.C.; Liechti, M.E. Duloxetine Inhibits Effects of MDMA (“Ecstasy”) In Vitro and in Humans in a Randomized Placebo-Controlled Laboratory Study. PLoS ONE 2012, 7, e36476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatsumi, M.; Groshan, K.; Blakely, R.D.; Richelson, E. Pharmacological profile of antidepressants and related compounds at human monoamine transporters. Eur. J. Pharmacol. 1997, 340, 249–258. [Google Scholar] [CrossRef]
- Scholze, P.; Zwach, J.; Kattinger, A.; Pifl, C.; Singer, E.A.; Sitte, H.H. Transporter-mediated release: A superfusion study on human embryonic kidney cells stably expressing the human serotonin transporter. J. Pharmacol. Exp. Ther. 2000, 293, 870–878. [Google Scholar] [PubMed]
- Luethi, D.; Trachsel, D.; Hoener, M.C.; Liechti, M.E. Monoamine receptor interaction profiles of 4-thio-substituted phenethylamines (2C-T drugs). Neuropharmacology 2018, 134, 141–148. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| NET | DAT | SERT | DAT/SERT Ratio | |
|---|---|---|---|---|
| IC50 [μM] (95% CI) | IC50 [μM] (95% CI) | IC50 [μM] (95% CI) | Ratio (95% CI) | |
| Pyrovalerone cathinones | ||||
| α-PPP | 0.64 (0.41–0.99) | 0.56 (0.40–0.76) | >1000 | >1000 |
| α-PVP | 0.02 (0.01–0.05) | 0.03 (0.02–0.04) | 279 (209–372) | >1000 |
| α-PHP | 0.06 (0.03–0.12) | 0.06 (0.05–0.08) | 245 (173–348) | >1000 |
| 4-MePPP | 0.64 (0.43–0.95) | 0.75 (0.58–0.97) | 55 (38–78) | 72 (39–134) |
| Pyrovalerone | 0.06 (0.04–0.09) | 0.05 (0.04–0.08) | 13 (10–18) | 256 (125–450) |
| MPHP | 0.07 (0.05–0.10) | 0.06 (0.05–0.08) | 11 (8.0–15) | 169 (100–300) |
| MDPPP | 1.7 (1.3–2.2) | 0.54 (0.37–0.79) | 43 (34–55) | 80 (43–149) |
| MDPBP | 0.14 (0.10–0.19) | 0.07 (0.06–0.09) | 11 (8.6–15) | 155 (96–250) |
| MDPV | 0.05 (0.04–0.08) | 0.03 (0.03–0.05) | 8.4 (6.6–11) | 241 (132–367) |
| MDPHP | 0.06 (0.03–0.13) | 0.05 (0.04–0.07) | 9 (6.0–14) | 184 (86–350) |
| MOPPP | 8.7 (6.3–12) | 4.6 (3.4–6.3) | 94 (65–135) | 20 (4.0–40) |
| 4F-PBP | 0.61 (0.43–0.87) | 0.50 (0.36–0.69) | 177 (97–325) | 356 (141–903) |
| Other | ||||
| NEH | 0.17 (0.12–0.24) | 0.18 (0.14–0.24) | 47 (38–60) | 264 (158–429) |
| Reference substances | ||||
| MDMA | 0.41 (0.33–0.52) | 13 (11–16) | 1.6 (1.2–2.2) | 0.12 (0.08–0.20) |
| Amphetamine | 0.07 (0.05–0.1) a | 1.3 (0.8–2.0) a | 45 (24–85) a | 35 (12–106) a |
| h5-HT1A | h5-HT2A | h5-HT2B | h5-HT2C | ||
|---|---|---|---|---|---|
| Receptor Binding | Receptor Binding | Activation Potency | Activation Potency | Receptor Binding | |
| Ki ± SD [μM] | Ki ± SD [μM] | EC50 ± SD [μM] | EC50 ± SD [μM] | Ki ± SD [μM] | |
| [3H]-8-OH-DPAT | [3H]-Ketanserin | [3H]-Mesulgerine | |||
| Pyrovalerone cathinones | |||||
| α-PPP | 0.7 ± 0.3 | 1.1 ± 0.3 | >20 | >20 | >15 |
| α-PVP | 6.1 ± 0.7 | >13 | >20 | >20 | >15 |
| α-PHP | 11 ± 5.3 | >13 | >20 | >20 | >15 |
| 4-MePPP | 12 ± 2.2 | 1.3 ± 0.4 | >10 | >10 | >5.1 |
| Pyrovalerone | 13 ± 1.4 | >13 | >20 | >20 | >15 |
| MPHP | 13 ± 1.9 | 7.3 ± 1.2 | >20 | >20 | >15 |
| MDPPP | 1.9 ± 0.8 | 8.0 ± 1.2 | >20 | >20 | >15 |
| MDPBP | 9.0 ± 1.5 | >13 | >20 | >20 | >15 |
| MDPV | 7.7 ± 0.6 | >13 | >20 | >20 | >15 |
| MDPHP | 13 ± 1.9 | >13 | >20 | >20 | >15 |
| MOPPP | >17 | >13 | >20 | >20 | >15 |
| 4F-PBP | 6.0 ± 3.4 | >13 | >20 | >20 | >15 |
| Other | |||||
| NEH | >17 | >13 | >20 | >20 | >15 |
| Reference substances | |||||
| MDMA | 11 ± 2.0 a | 6.3 ± 2.4 b | 6.1 ± 0.3 b | >20 b | 4.4 ± 0.8 a |
| Amphetamine | 6.7 ± 1.4 c | >13 c | NA | 9.4 ± 1.6 b | >13 c |
| hNET | hDAT | hSERT | D2 | α1A | α2A | Human TAAR1 | Rat TAAR1 | Mouse TAAR1 | |
|---|---|---|---|---|---|---|---|---|---|
| Receptor Binding | Activation Potency | Receptor Binding | |||||||
| Ki ± SD [μM] | EC50 ± SD [μM] | Ki ± SD [μM] | |||||||
| N-Methyl-[3H]-nisoxetine | [3H]-WIN-35,428 | [3H]-Citalopram | [3H]-Spiperone | [3H]-Prazosin | [3H]-Rauwolscine | [3H]-RO5166017 | |||
| Pyrovalerone cathinones | |||||||||
| α-PPP | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| α-PVP | 0.06 ± 0.02 d | 0.007 ± 0.002 d | >30 d | >10 d | >15 d | >20 d | NA | 16 ± 6.4 d | >20 d |
| α-PHP | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| 4-MePPP | 2.5 ± 0.7 | 0.3 ± 0.02 | >7.4 | >13 | 2.2 ± 0.1 | >4.7 | NA | NA | NA |
| Pyrovalerone | 0.06 ± 0.01 c | 0.03 ± 0.01 c | 5.0 ± 0.3 c | >30 c | >6 c | >20 c | NA | >12.5 c | >10 c |
| MPHP | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| MDPPP | 3.5 ± 1.0 d | 0.18 ± 0.05 d | 12 ± 1.0 d | >10 d | >15 d | 14 ± 0.9 d | NA | 16 ± 6.7 d | >20 d |
| MDPBP | 1.1 ± 0.1 d | 0.02 ± 0.002 d | 4.1 ± 1.2 d | >20 d | >4.9 d | 9.4 ± 1.6 d | NA | >20 d | >20 d |
| MDPV | 0.08 ± 0.02 c | 0.01 ± 0.002 c | 2.9 ± 0.1 c | >30 c | >6 c | >20 c | >30 b | 7.2 ± 1.1 c | >10 c |
| MDPHP | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| MOPPP | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| 4F-PBP | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| Other | |||||||||
| NEH | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| Reference substances | |||||||||
| MDMA | >8.7 a | >8.5 a | >7.5 a | >13 a | 6.9 ± 1.2 a | 4.6 ± 1.1 a | 35 ± 21 b | 0.25 ± 0.01 a | 3.1 ± 0.7 a |
| Amphetamine | 1.0 ± 0.6 c | 5.7 ± 3.8 c | >25 c | >30 c | >6.0 c | 2.8 ± 0.8 c | 2.8 ± 0.8 b | 0.23 ± 0.18 c | 0.09 ± 0.06 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolaczynska, K.E.; Thomann, J.; Hoener, M.C.; Liechti, M.E. The Pharmacological Profile of Second Generation Pyrovalerone Cathinones and Related Cathinone Derivative. Int. J. Mol. Sci. 2021, 22, 8277. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158277
Kolaczynska KE, Thomann J, Hoener MC, Liechti ME. The Pharmacological Profile of Second Generation Pyrovalerone Cathinones and Related Cathinone Derivative. International Journal of Molecular Sciences. 2021; 22(15):8277. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158277
Chicago/Turabian StyleKolaczynska, Karolina E., Jan Thomann, Marius C. Hoener, and Matthias E. Liechti. 2021. "The Pharmacological Profile of Second Generation Pyrovalerone Cathinones and Related Cathinone Derivative" International Journal of Molecular Sciences 22, no. 15: 8277. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158277