
Gran1: A Granulysin-Derived Peptide with Potent Activity against Intracellular Mycobacterium tuberculosis

 and
and
Abstract
:1. Introduction
2. Results
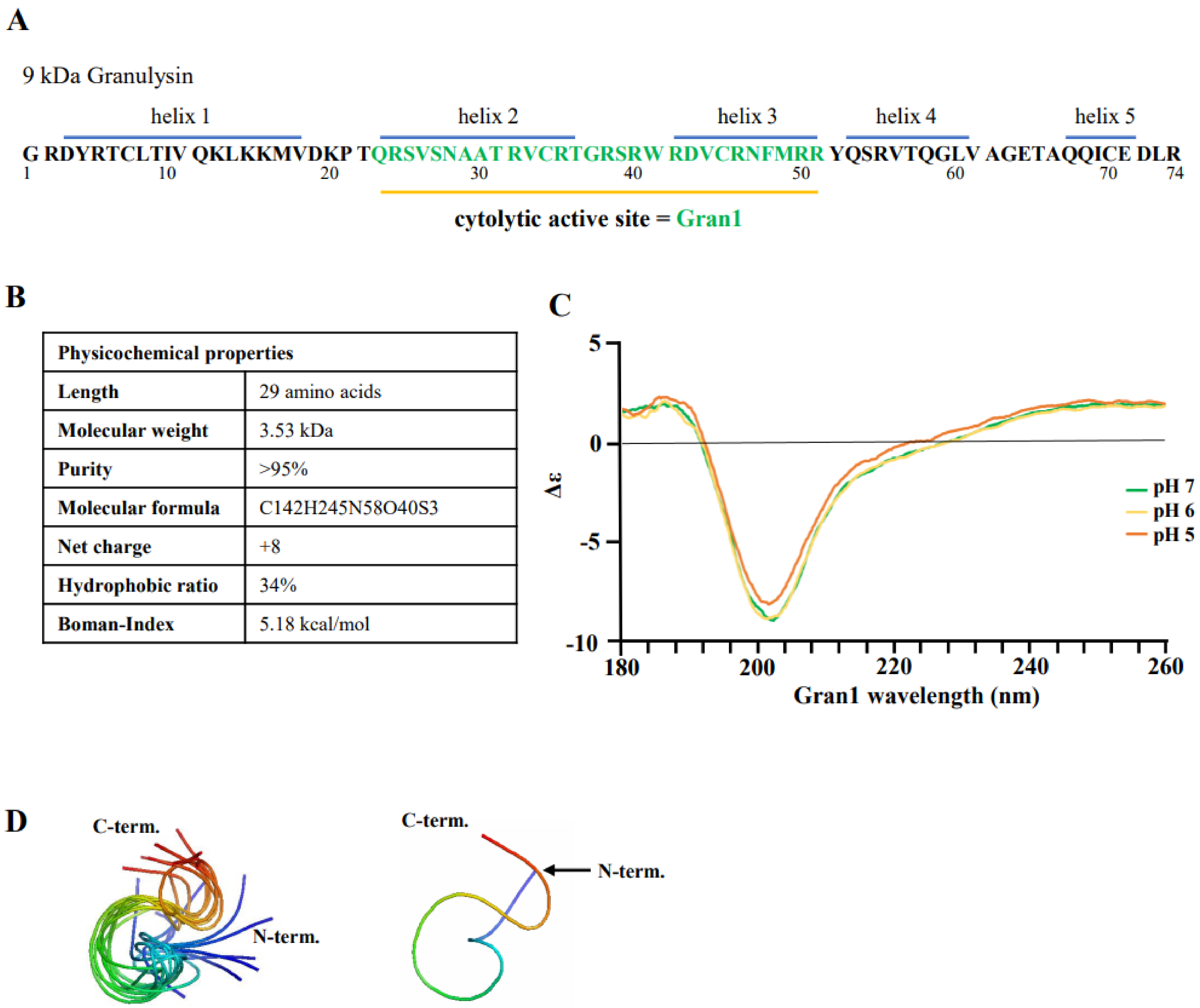
2.1. Structure and Physicochemical Characteristics of Gran1
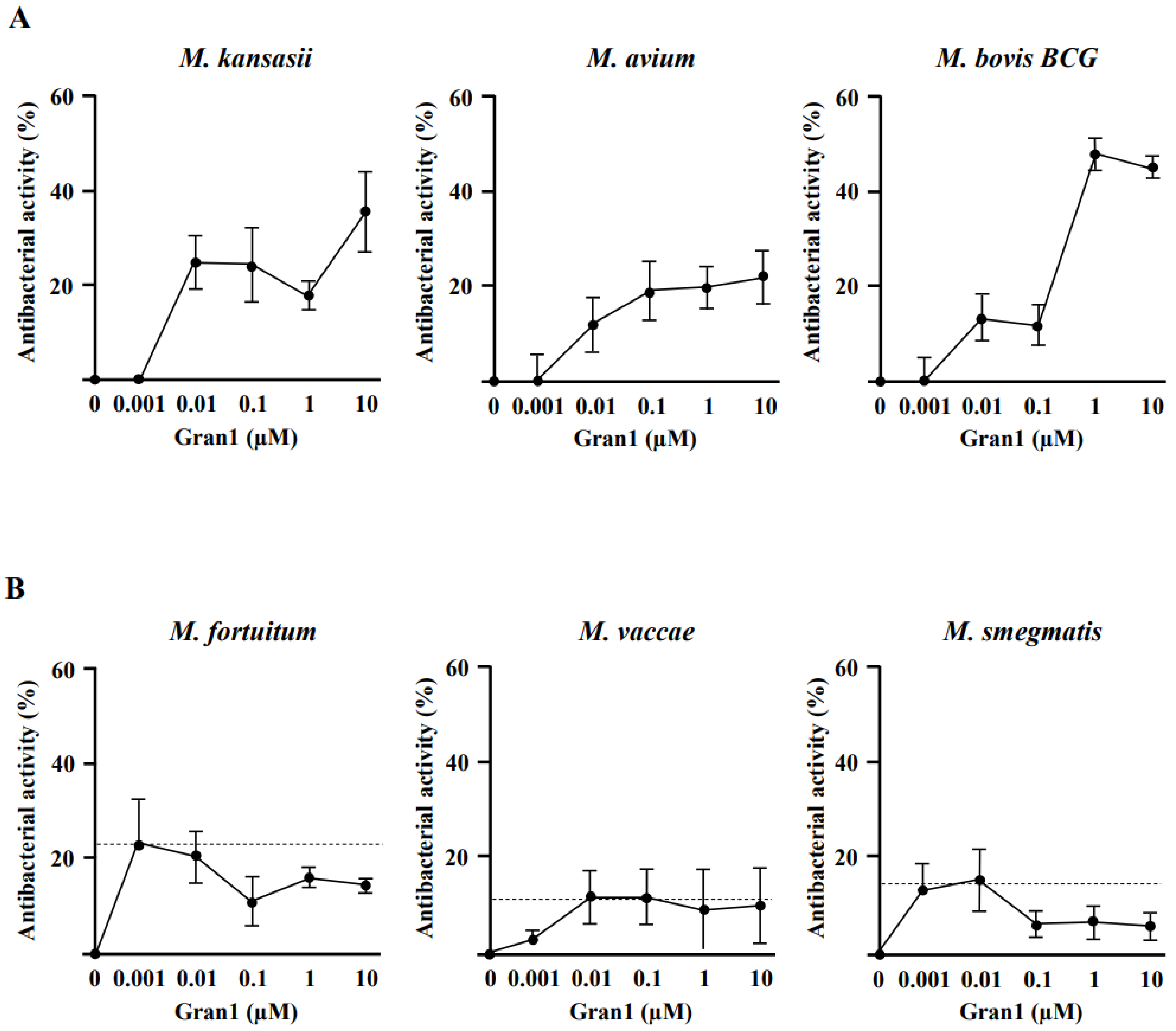
2.2. Antibacterial Activity of Gran1 against Extracellular Mycobacteria
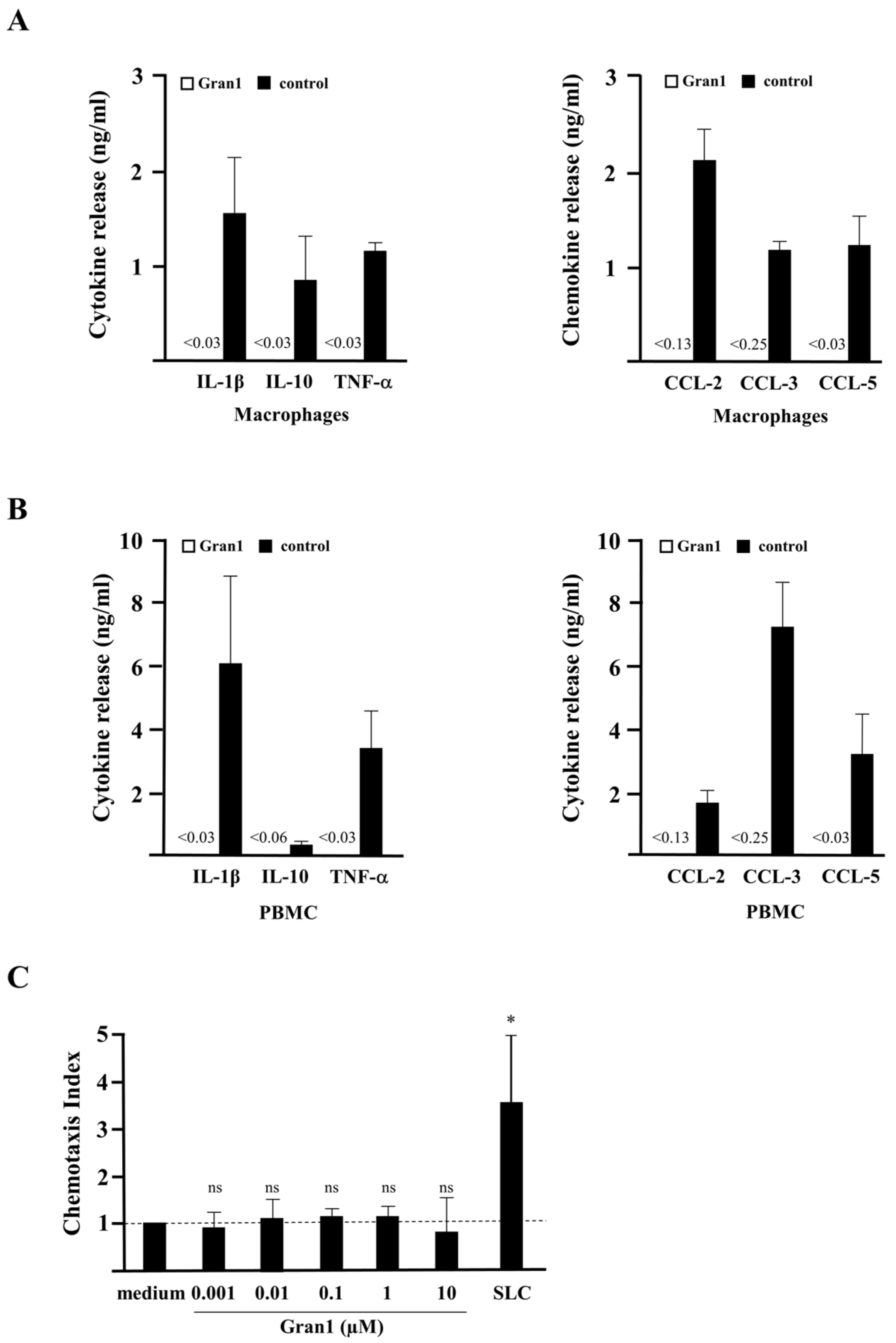
2.3. Off-Target Effects and Toxicity of Gran1
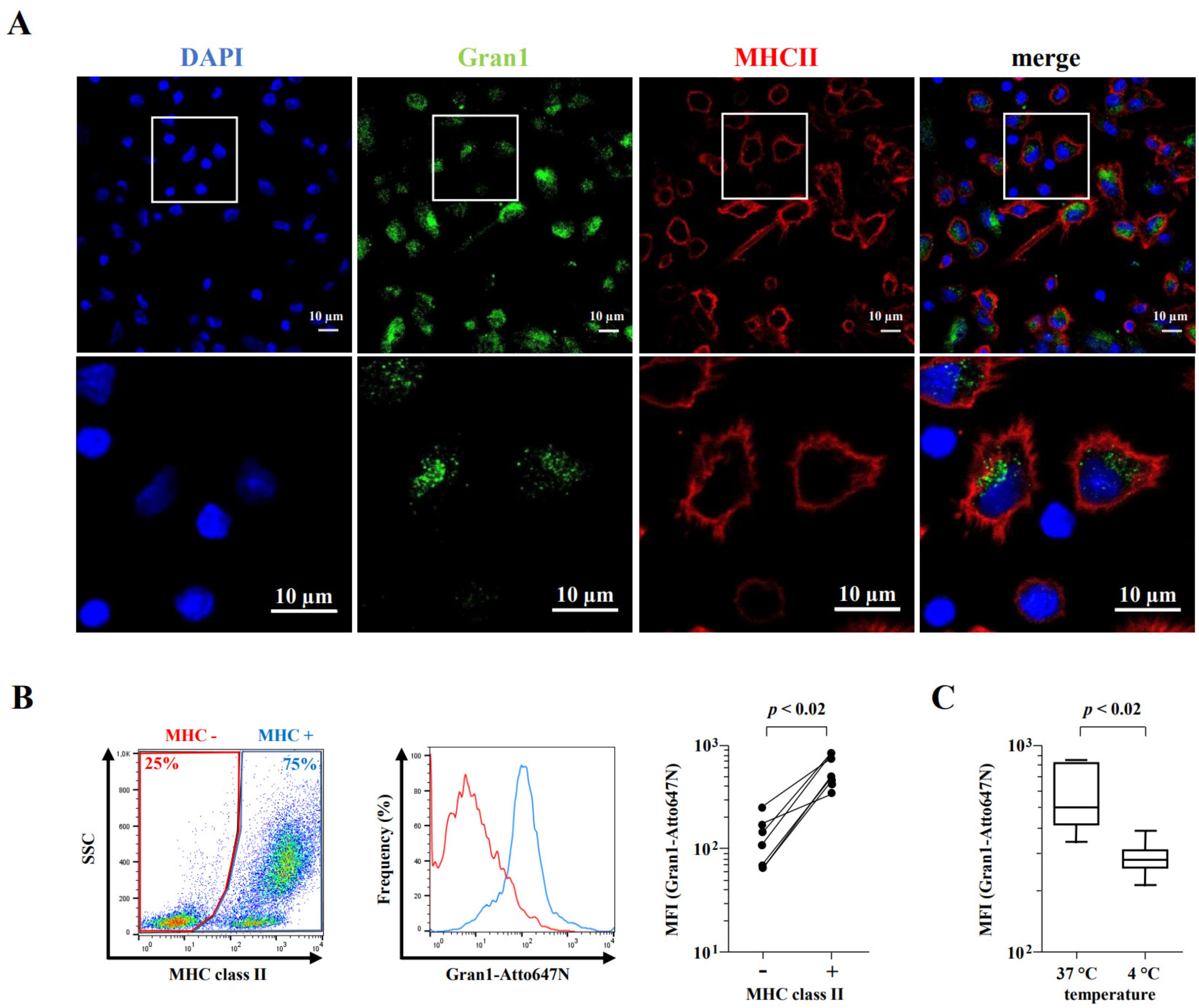
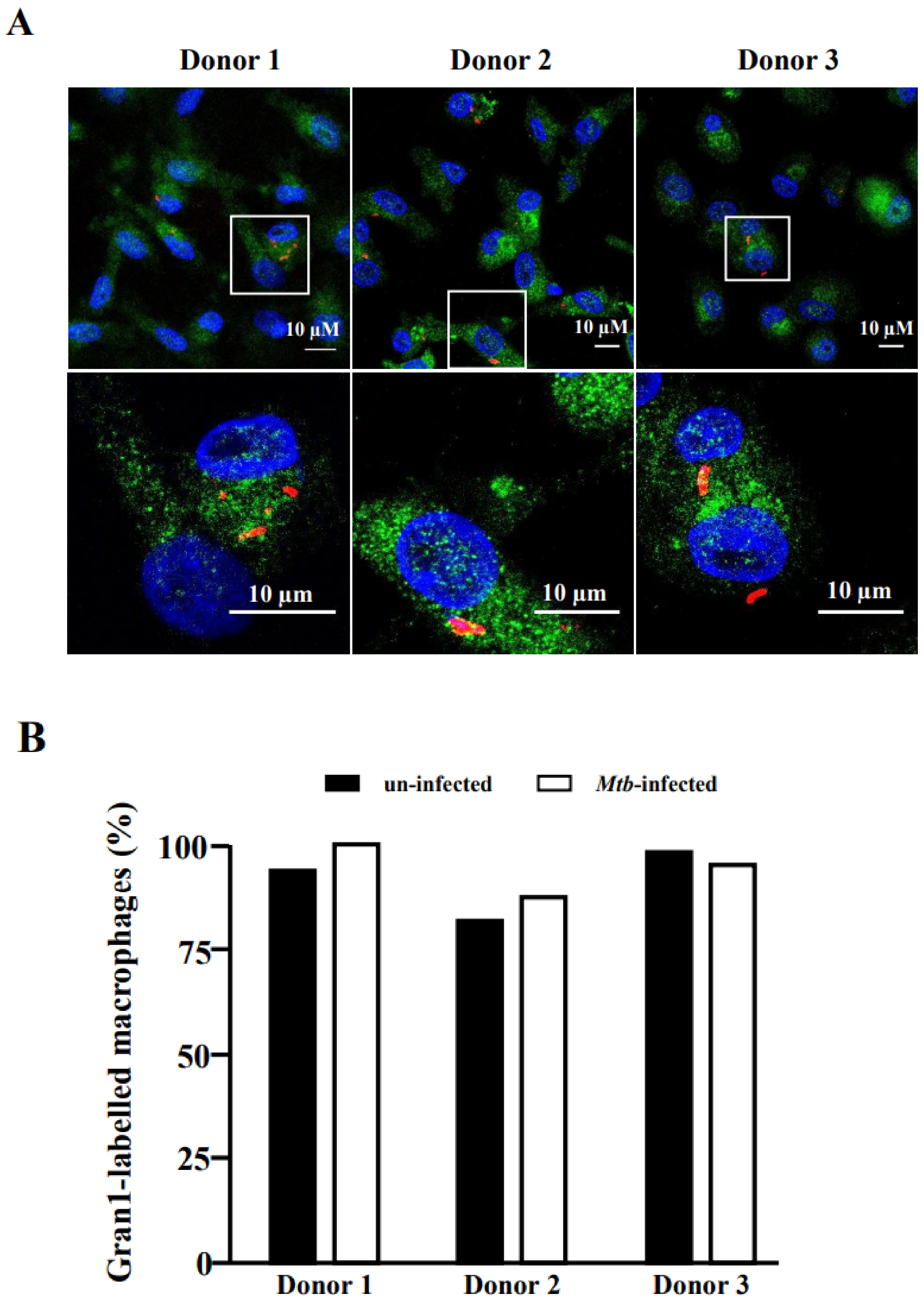
2.4. Internalization of Gran1 by Macrophages
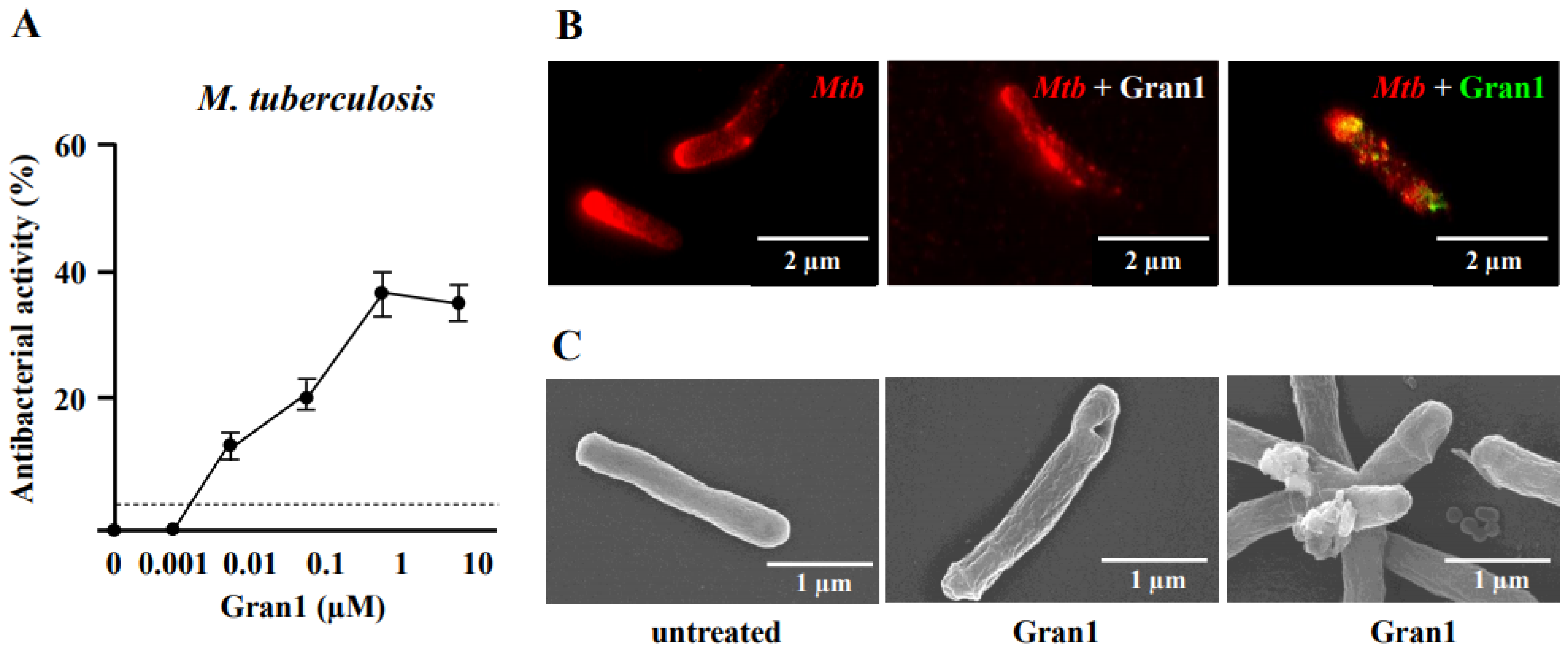
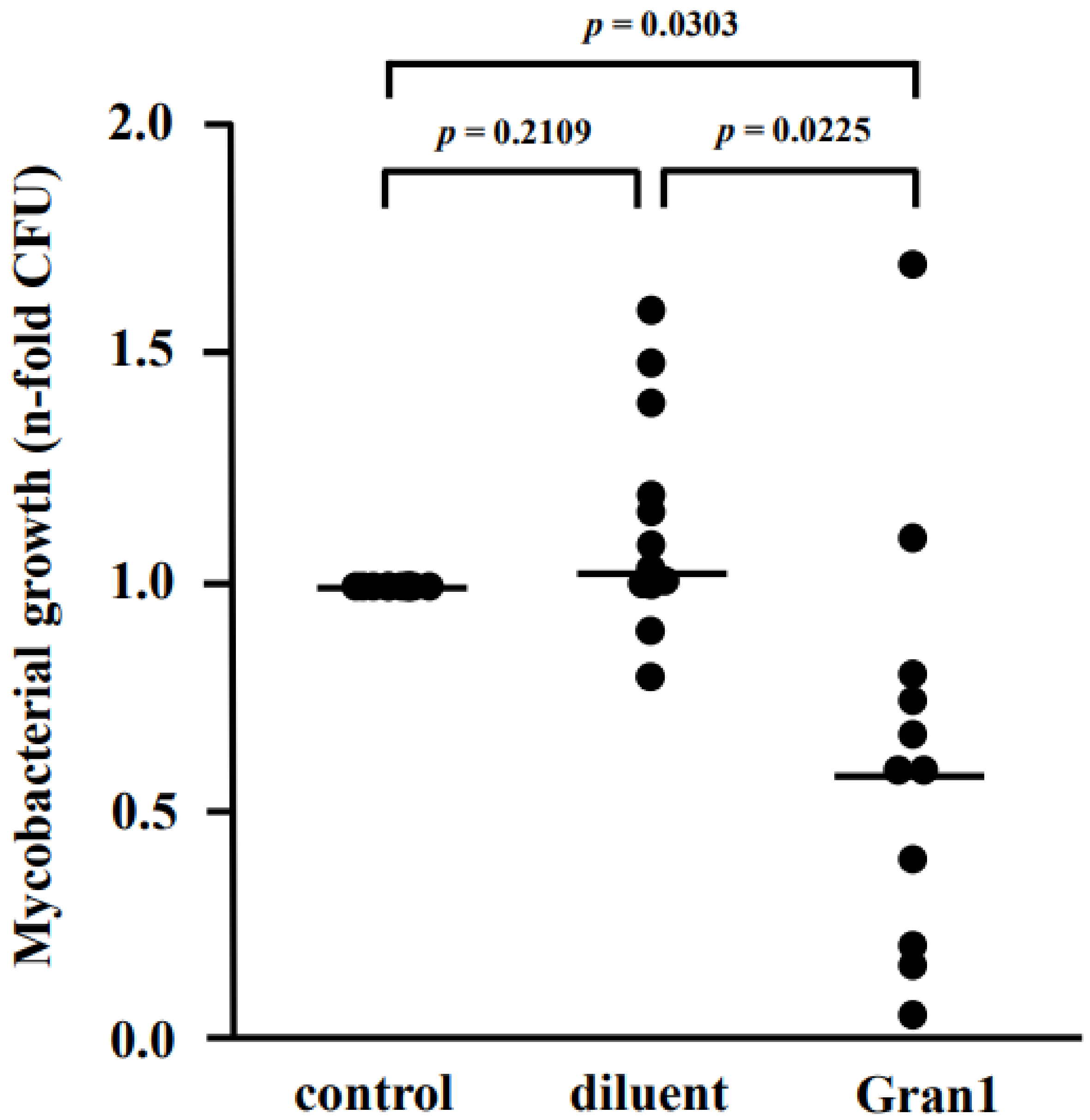
2.5. Antibacterial Activity of Gran1 against Intracellular Mtb
3. Discussion
4. Materials and Methods
4.1. Peptide Synthesis
4.2. Structural Analysis of Gran1
4.2.1. Circular Dichroism Spectrometry
4.2.2. Nuclear Magnetic Resonance
4.3. Source and Culture of Mycobacteria
4.4. Antibodies and Reagents
4.5. Chemotaxis Assay
4.6. Cytokine and Chemokine Release
4.7. Growth of Extracellular Mycobacteria: 3H-Uracil Proliferation Assay
4.8. Stimulated Emission Depletion (STED) Microscopy
4.9. Scanning Electron Microscopy
4.10. Confocal Laser Scanning Microscopy
4.11. Toxicity of Gran1 against Macrophages and Zebrafish
4.12. Intracellular Detection of Gran1 by Flow Cytometry
4.13. Quantification of Intracellular Mycobacterial Growth
4.14. Statistical Analysis
4.15. Ethical Statement
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| aa | Amino acids |
| APD3 | Antimicrobial Peptide Database 3 |
| AMP | antimicrobial peptide |
| ATCC | American Type Culture Collection |
| BSA | bovine serum albumin |
| CCL | Chemokine (C-C motif) ligand |
| CFU | colony forming units |
| dpf | days post fertilization |
| hpf | hours post fertilization |
| FACS | fluorescence activated cell sorting |
| MHCII | major histocompatibility complex class 2 |
| MOI | multiplicity of infection |
| Mtb | Mycobacterium tuberculosis |
| NTM | Non-tuberculous mycobateria |
| PBMC | peripheral blood mononuclear cell |
| PBS | phosphate buffered saline |
| PHA | Phytohaemagglutinin |
| PFA | Paraformaldehyde |
| SD | Standard deviation |
| SEM | Standard error of the mean |
| SFM | Serum-free medium |
| SLC | secondary lymphoid-tissue chemokine |
| STED | Stimulated emission depletion |
References
- Peña, S.V.; Hanson, D.A.; Carr, B.A.; Goralski, T.J.; Krensky, A.M. Processing, Subcellular Localization, and Function of 519 (Granulysin), a Human Late T Cell Activation Molecule with Homology to Small, Lytic, Granule Proteins. J. Immunol. 1997, 158, 2680–2688. [Google Scholar]
- Stenger, S.; Hanson, D.A.; Teitelbaum, R.; Dewan, P.; Niazi, K.R.; Froelich, C.J.; Ganz, T.; Thoma-Uszynski, S.; Melián, A.; Bogdan, C.; et al. An Antimicrobial Activity of Cytolytic T Cells Mediated by Granulysin. Science 1998, 282. [Google Scholar] [CrossRef] [Green Version]
- Pardo, J.; Pérez-Galán, P.; Gamen, S.; Marzo, I.; Monleón, I.; Kaspar, A.A.; Susín, S.A.; Kroemer, G.; Krensky, A.M.; Naval, J.; et al. A Role of the Mitochondrial Apoptosis-Inducing Factor in Granulysin-Induced Apoptosis. J. Immunol. 2001, 167, 1222–1229. [Google Scholar] [CrossRef] [Green Version]
- Kaspar, A.A.; Okada, S.; Kumar, J.; Poulain, F.R.; Drouvalakis, K.A.; Kelekar, A.; Hanson, D.A.; Kluck, R.M.; Hitoshi, Y.; Johnson, D.E.; et al. A Distinct Pathway of Cell-Mediated Apoptosis Initiated by Granulysin. J. Immunol. 2001, 167, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Gamen, S.; Hanson, D.A.; Kaspar, A.; Naval, J.; Krensky, A.M.; Anel, A. Granulysin-Induced Apoptosis. I. Involvement of at Least Two Distinct Pathways. J. Immunol. 1998, 161, 1758–1764. [Google Scholar]
- Deng, A.; Chen, S.; Li, Q.; Lyu, S.; Clayberger, C.; Krensky, A.M. Granulysin, a Cytolytic Molecule, Is Also a Chemoattractant and Proinflammatory Activator. J. Immunol. 2005, 174, 5243–5248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dotiwala, F.; Lieberman, J. Granulysin: Killer Lymphocyte Safeguard against Microbes. Curr. Opin. Immunol. 2019, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Krensky, A.M.; Clayberger, C. Biology and Clinical Relevance of Granulysin: REVIEW ARTICLE. Tissue Antigens. 2009, 193–198. [Google Scholar] [CrossRef]
- Dotiwala, F.; Mulik, S.; Polidoro, R.B.; Ansara, J.A.; Burleigh, B.A.; Walch, M.; Gazzinelli, R.T.; Lieberman, J. Killer Lymphocytes Use Granulysin, Perforin and Granzymes to Kill Intracellular Parasites. Nat. Med. 2016, 22, 210–216. [Google Scholar] [CrossRef] [PubMed]
- McInturff, J.E.; Wang, S.J.; Machleidt, T.; Lin, T.R.; Oren, A.; Hertz, C.J.; Krutzik, S.R.; Hart, S.; Zeh, K.; Anderson, D.H.; et al. Granulysin-Derived Peptides Demonstrate Antimicrobial and Anti-Inflammatory Effects against Propionibacterium Acnes. J. Investig. Dermatol. 2005, 125, 256–263. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.S.; Chun, S.M.; Soung, M.G.; Kim, J.; Kim, S.J. Antimicrobial Efficacy of Granulysin-Derived Synthetic Peptides in Acne Vulgaris. Int. J. Dermatol. 2015, 54, 853–862. [Google Scholar] [CrossRef]
- Da Silva, A.P.G.; Unks, D.; Lyu, S.C.; Ma, J.; Zbozien-Pacamaj, R.; Chen, X.; Krensky, A.M.; Clayberger, C. In Vitro and In Vivo Antimicrobial Activity of Granulysin-Derived Peptides against Vibrio Cholerae. J. Antimicrob. Chemother. 2008, 61, 1103–1109. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Choice, E.; Kaspar, A.; Hanson, D.; Okada, S.; Lyu, S.-C.; Krensky, A.M.; Clayberger, C. Bactericidal and Tumoricidal Activities of Synthetic Peptides Derived from Granulysin. J. Immunol. 2000, 165, 1486–1490. [Google Scholar] [CrossRef] [Green Version]
- Siano, A.; Tonarelli, G.; Imaz, M.; Perin, J.; Ruggeri, N.; Lopez, M.; Santi, M.; Zerbini, E. Bactericidal and Hemolytic Activities of Synthetic Peptides Derived from Granulysin. Protein Pept. Lett. 2010, 17, 517–521. [Google Scholar] [CrossRef]
- Andreu, D.; Carreño, C.; Linde, C.; Boman, H.G.; Andersson, M. Identification of an Anti-Mycobacterial Domain in NK-Lysin and Granulysin. Biochem. J. 1999, 344. [Google Scholar] [CrossRef]
- Global Tuberculosis Report 2020; World Health Organization: Geneva, Switzerland, 2020; Licence: CC BY-NC-SA 3.0 IGO.
- Rieping, W.; Bardiaux, B.; Bernard, A.; Malliavin, T.E.; Nilges, M. ARIA2: Automated NOE Assignment and Data Integration in NMR Structure Calculation. Bioinformatics 2007, 23, 381–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Li, X.; Wang, Z. APD3: The Antimicrobial Peptide Database as a Tool for Research and Education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [Green Version]
- Gutsmann, T. Interaction between Antimicrobial Peptides and Mycobacteria. Biochim. Biophys. Acta-Biomembr. 2016, 1858, 1034–1043. [Google Scholar] [CrossRef]
- Morash, M.G.; Douglas, S.E.; Robotham, A.; Ridley, C.M.; Gallant, J.W.; Soanes, K.H. The Zebrafish Embryo as a Tool for Screening and Characterizing Pleurocidin Host-Defense Peptides as Anti-Cancer Agents. DMM Dis. Model. Mech. 2011, 4, 622–633. [Google Scholar] [CrossRef] [Green Version]
- Beitzinger, B.; Gerbl, F.; Vomhof, T.; Schmid, R.; Noschka, R.; Rodriguez, A.; Wiese, S.; Weidinger, G.; Ständker, L.; Walther, P.; et al. Delivery by Dendritic Mesoporous Silica Nanoparticles Enhances the Antimicrobial Activity of a Napsin-Derived Peptide Against Intracellular Mycobacterium Tuberculosis. Adv. Healthc. Mater. 2021, 2100453. [Google Scholar] [CrossRef]
- Ernst, W.A.; Thoma-Uszynski, S.; Teitelbaum, R.; Ko, C.; Hanson, D.A.; Clayberger, C.; Krensky, A.M.; Leippe, M.; Bloom, B.R.; Ganz, T.; et al. Granulysin, a T Cell Product, Kills Bacteria by Altering Membrane Permeability. J. Immunol. 2000, 165. [Google Scholar] [CrossRef] [Green Version]
- Linde, C.M.A.; Grundström, S.; Nordling, E.; Refai, E.; Brennan, P.J.; Andersson, M. Conserved Structure and Function in the Granulysin and NK-Lysin Peptide Family. Infect. Immun. 2005, 73, 6332–6339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boman, H.G. Antibacterial Peptides: Basic Facts and Emerging Concepts. J. Intern. Med. 2003, 254. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, H.; Sun, Z. Susceptibility of Mycobacterium Tuberculosis to Weak Acids. J. Antimicrob. Chemother. 2003, 52, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Post-Martens, K.; Denkin, S. New Drug Candidates and Therapeutic Targets for Tuberculosis Therapy. Drug Discov. Today 2006, 11, 21–27. [Google Scholar] [CrossRef]
- Deshpande, D.; Grieshober, M.; Wondany, F.; Gerbl, F.; Noschka, R.; Michaelis, J.; Stenger, S. Super-Resolution Microscopy Reveals a Direct Interaction of Intracellular Mycobacterium Tuberculosis with the Antimicrobial Peptide LL-37. Int. J. Mol. Sci. 2020, 21, 6741. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Lu, X.; Deng, Z.; Xiao, S.; Yuan, B.; Yang, K. How Melittin Inserts into Cell Membrane: And Disturbance on the Membrane. Molecules 2019, 24, 1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Dong, C.; Deng, A.; Katsumata, M.; Nakadai, A.; Kawada, T.; Okada, S.; Clayberger, C.; Krensky, A.M. Hemolysis of Erythrocytes by Granulysin-Derived Peptides but Not by Granulysin. Antimicrob. Agents Chemother. 2005, 49, 388–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jobin, K.; Müller, D.N.; Jantsch, J.; Kurts, C. Sodium and Its Manifold Impact on Our Immune System. Trends Immunol. 2021, 42, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Di, A.; Xiong, S.; Ye, Z.; Malireddi, R.K.S.; Kometani, S.; Zhong, M.; Mittal, M.; Hong, Z.; Kanneganti, T.-D.; Rehman, J.; et al. The TWIK2 Potassium Efflux Channel in Macrophages Mediates NLRP3 Inflammasome-Induced Inflammation. Immunity 2018, 49. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.; Silwal, P.; Kim, I.; Modlin, R.L.; Jo, E.K. Vitamin D-Cathelicidin Axis: At the Crossroads between Protective Immunity and Pathological Inflammation during Infection. Immune Netw. 2020, 20, 1–26. [Google Scholar] [CrossRef]
- Liu, P.T.; Stenger, S.; Li, H.; Wenzel, L.; Tan, B.H.; Krutzik, S.R.; Ochoa, M.T.; Schauber, J.; Wu, K.; Meinken, C.; et al. Toll-like Receptor Triggering of a Vitamin D-Mediated Human Antimicrobial Response. Science 2006, 311, 1770–1773. [Google Scholar] [CrossRef]
- Gutierrez, M.G.; Mishra, B.B.; Jordao, L.; Elliott, E.; Anes, E.; Griffiths, G. NF-ΚB Activation Controls Phagolysosome Fusion-Mediated Killing of Mycobacteria by Macrophages. J. Immunol. 2008, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, M.G.; Master, S.S.; Singh, S.B.; Taylor, G.A.; Colombo, M.I.; Deretic, V. Autophagy Is a Defense Mechanism Inhibiting BCG and Mycobacterium Tuberculosis Survival in Infected Macrophages. Cell 2004, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Harten, R.M.; van Woudenbergh, E.; van Dijk, A.; Haagsman, H.P. Cathelicidins: Immunomodulatory Antimicrobials. Vaccines 2018, 6, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, J.P.; Appelberg, R.; Gama, F.M. Antimicrobial Peptides as Novel Anti-Tuberculosis Therapeutics. Biotechnol. Adv. 2016, 34, 924–940. [Google Scholar] [CrossRef] [Green Version]
- Sonawane, A.; Santos, J.C.; Mishra, B.B.; Jena, P.; Progida, C.; Sorensen, O.E.; Gallo, R.; Appelberg, R.; Griffiths, G. Cathelicidin Is Involved in the Intracellular Killing of Mycobacteria in Macrophages. Cell. Microbiol. 2011, 13, 1601–1617. [Google Scholar] [CrossRef]
- Stek, C.; Allwood, B.; Walker, N.F.; Wilkinson, R.J.; Lynen, L.; Meintjes, G. The Immune Mechanisms of Lung Parenchymal Damage in Tuberculosis and the Role of Host-Directed Therapy. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ständker, L.; Wobst, P.; Mark, S.; Forssmann, W.G. Isolation and Characterization of Circulating 13-KDa C-Terminal Fragments of Human Insulin-like Growth Factor Binding Protein-5. FEBS Lett. 1998, 441, 281–286. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Mao, X.A.; Ye, C.; Huang, H.; Nicholson, J.K.; Lindon, J.C. Improved Watergate Pulse Sequences for Solvent Suppression in NMR Spectroscopy. J. Magn. Reson. 1998, 132, 125–129. [Google Scholar] [CrossRef]
- Bax, A.D.; Donald, G.D. MLEV-17-Based Two-Dimensional Homonuclear Magnetization Transfer Spectroscopy. J. Magn. Reson. 1969, 65, 355–360. [Google Scholar] [CrossRef]
- Piotto, M.; Saudek, V.; Sklenář, V. Gradient-Tailored Excitation for Single-Quantum NMR Spectroscopy of Aqueous Solutions. J. Biomol. NMR 1992, 2, 661–665. [Google Scholar] [CrossRef]
- Sklenáŕ, V.; Piotto, M.; Leppik, R.; Saudek, V. Gradient-Tailored Water Suppression for 1H-15N HSQC Experiments Optimized to Retain Full Sensitivity. J. Magn. Reson. Ser. A. 1993, 241–245. [Google Scholar] [CrossRef]
- Lee, W.; Tonelli, M.; Markley, J.L. NMRFAM-SPARKY: Enhanced Software for Biomolecular NMR Spectroscopy. Bioinformatics 2015, 31, 1325–1327. [Google Scholar] [CrossRef] [Green Version]
- Bruns, H.; Stegelmann, F.; Fabri, M.; Döhner, K.; van Zandbergen, G.; Wagner, M.; Skinner, M.; Modlin, R.L.; Stenger, S. Abelson Tyrosine Kinase Controls Phagosomal Acidification Required for Killing of Mycobacterium Tuberculosis in Human Macrophages. J. Immunol. 2012, 189, 4069–4078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stegelmann, F.; Bastian, M.; Swoboda, K.; Bhat, R.; Kiessler, V.; Krensky, A.M.; Roellinghoff, M.; Modlin, R.L.; Stenger, S. Coordinate Expression of CC Chemokine Ligand 5, Granulysin, and Perforin in CD8 + T Cells Provides a Host Defense Mechanism against Mycobacterium Tuberculosis. J. Immunol. 2005, 175, 7474–7483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noschka, R.; Gerbl, F.; Löffler, F.; Kubis, J.; Rodríguez, A.A.; Mayer, D.; Grieshober, M.; Holch, A.; Raasholm, M.; Forssmann, W.G.; et al. Unbiased Identification of Angiogenin as an Endogenous Antimicrobial Protein With Activity Against Virulent Mycobacterium Tuberculosis. Front. Microbiol. 2021, 11. [Google Scholar] [CrossRef]
- Osseforth, C.; Moffitt, J.R.; Schermelleh, L.; Michaelis, J. Simultaneous Dual-Color 3D STED Microscopy. Opt. Express 2014, 22, 7028. [Google Scholar] [CrossRef] [PubMed]
- Walther, P.; Wang, L.; Ließem, S.; Frascaroli, G. Viral Infection of Cells in Culture. Approaches for Electron Microscopy; Academic Press: Cambridge, MA, USA, 2010; Volume 96. [Google Scholar] [CrossRef]
- Raftery, T.D.; Isales, G.M.; Yozzo, K.L.; Volz, D.C. High-Content Screening Assay for Identification of Chemicals Impacting Spontaneous Activity in Zebrafish Embryos. Environ. Sci. Technol. 2014, 48, 804–810. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Source |
|---|---|
| Mycobacterium tuberculosis | ATCC 27294 a |
| Mycobacterium avium | ATCC 25291 a |
| Mycobacterium kansasii | ATCC 12478 a |
| Mycobacterium bovis BCG | BCG medac, PZN: 02736484 |
| Mycobacterium smegmatis | ATCC 19420 a |
| Mycobacterium fortuitum | ATCC 6841 a |
| Mycobacterium vaccae | ATCC 15483 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noschka, R.; Wondany, F.; Kizilsavas, G.; Weil, T.; Weidinger, G.; Walther, P.; Michaelis, J.; Stenger, S. Gran1: A Granulysin-Derived Peptide with Potent Activity against Intracellular Mycobacterium tuberculosis. Int. J. Mol. Sci. 2021, 22, 8392. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168392
Noschka R, Wondany F, Kizilsavas G, Weil T, Weidinger G, Walther P, Michaelis J, Stenger S. Gran1: A Granulysin-Derived Peptide with Potent Activity against Intracellular Mycobacterium tuberculosis. International Journal of Molecular Sciences. 2021; 22(16):8392. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168392
Chicago/Turabian StyleNoschka, Reiner, Fanny Wondany, Gönül Kizilsavas, Tanja Weil, Gilbert Weidinger, Paul Walther, Jens Michaelis, and Steffen Stenger. 2021. "Gran1: A Granulysin-Derived Peptide with Potent Activity against Intracellular Mycobacterium tuberculosis" International Journal of Molecular Sciences 22, no. 16: 8392. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168392