
Der f 38 Is a Novel TLR4-Binding Allergen Related to Allergy Pathogenesis from Dermatophagoides farinae

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Identification of Der f 38 in DF
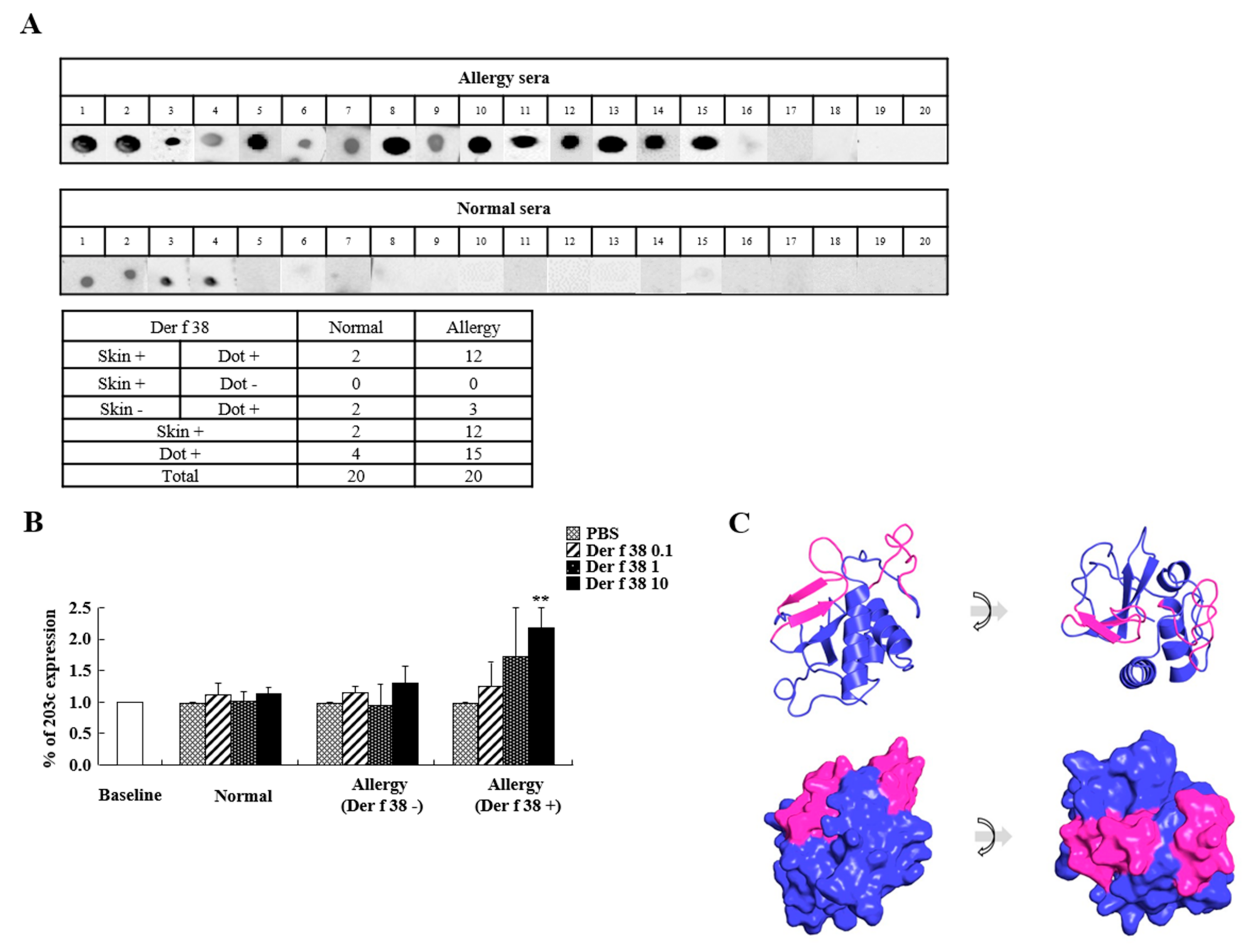
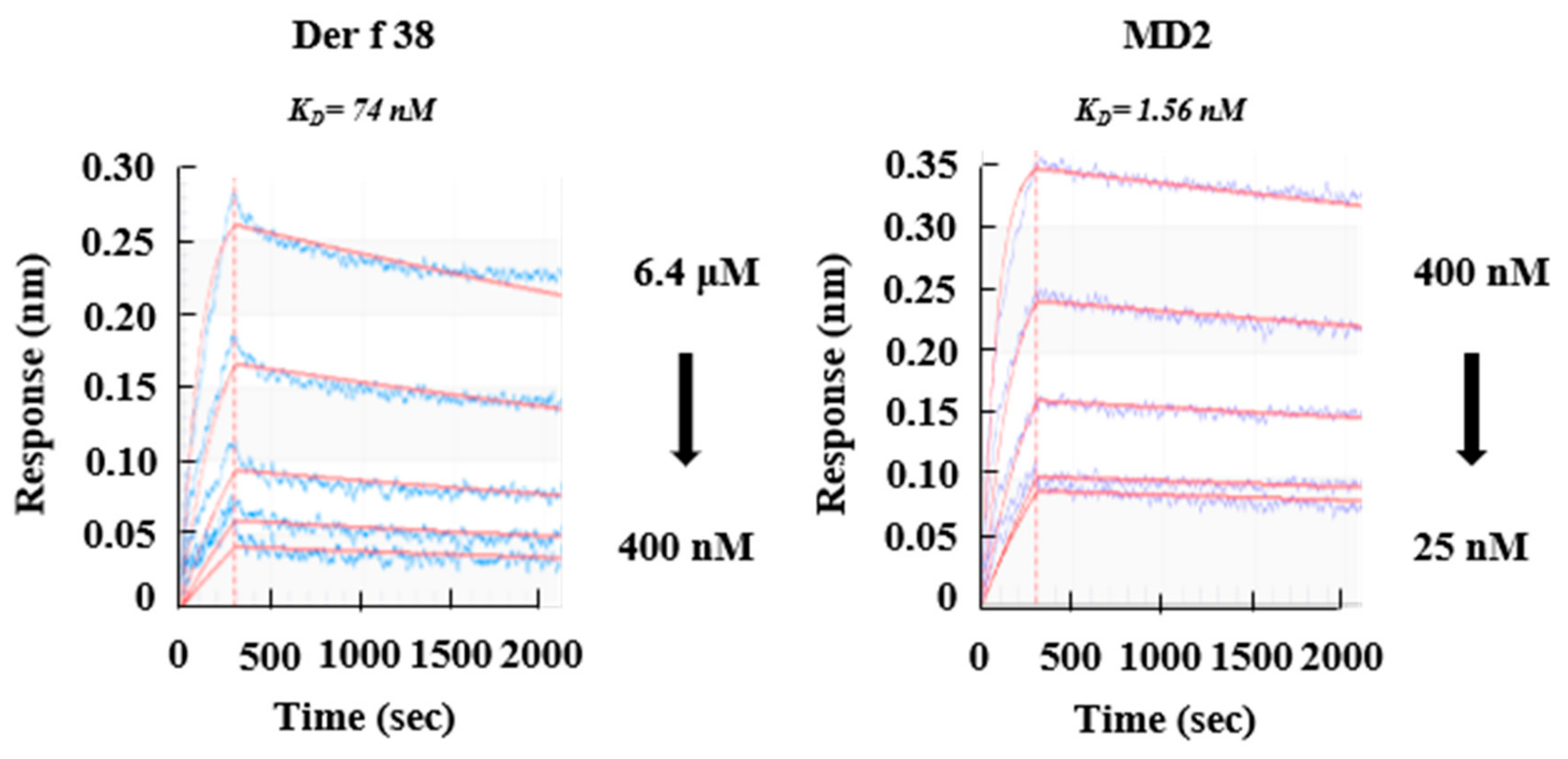
2.2. Allergenicity of Der f 38
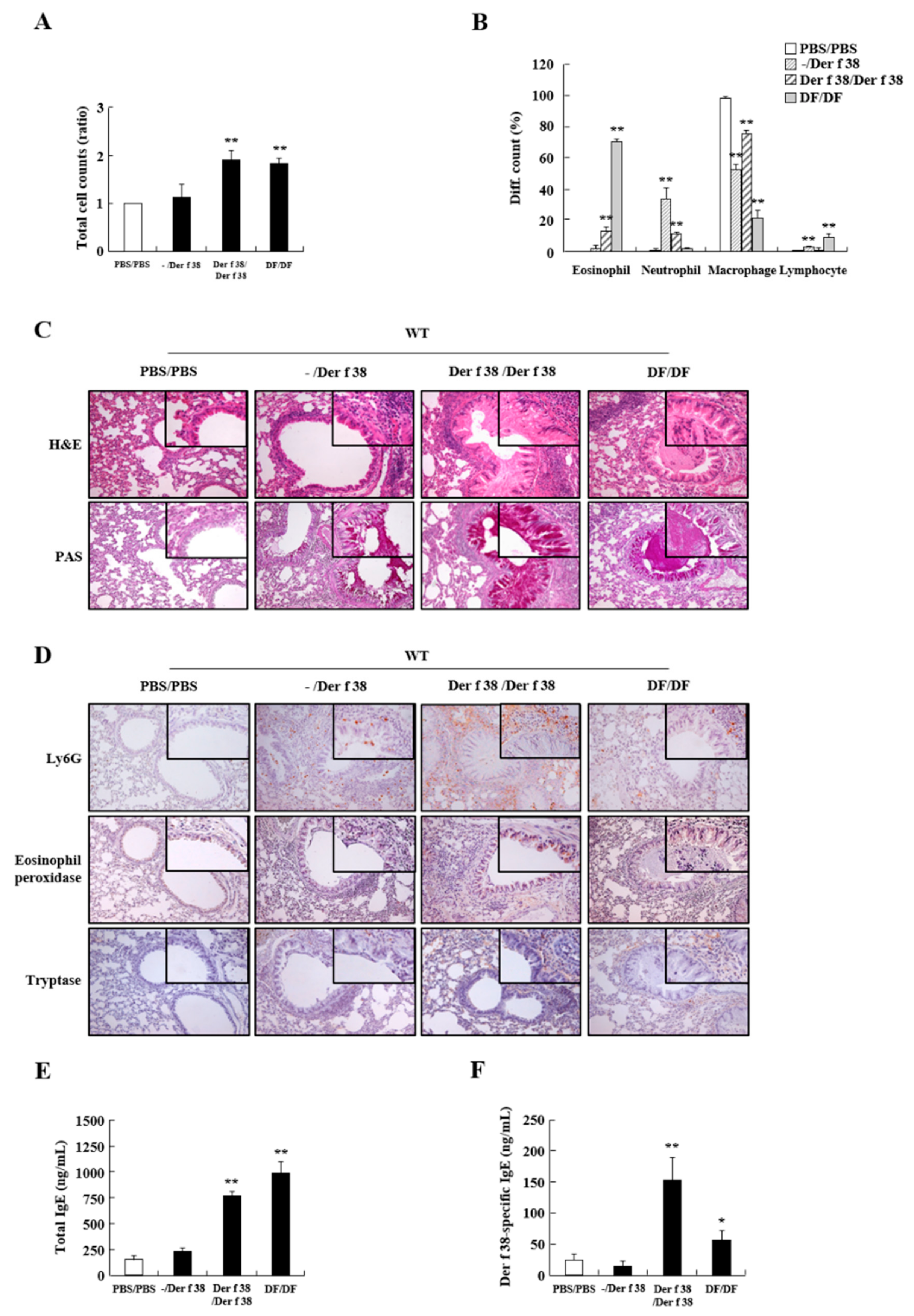
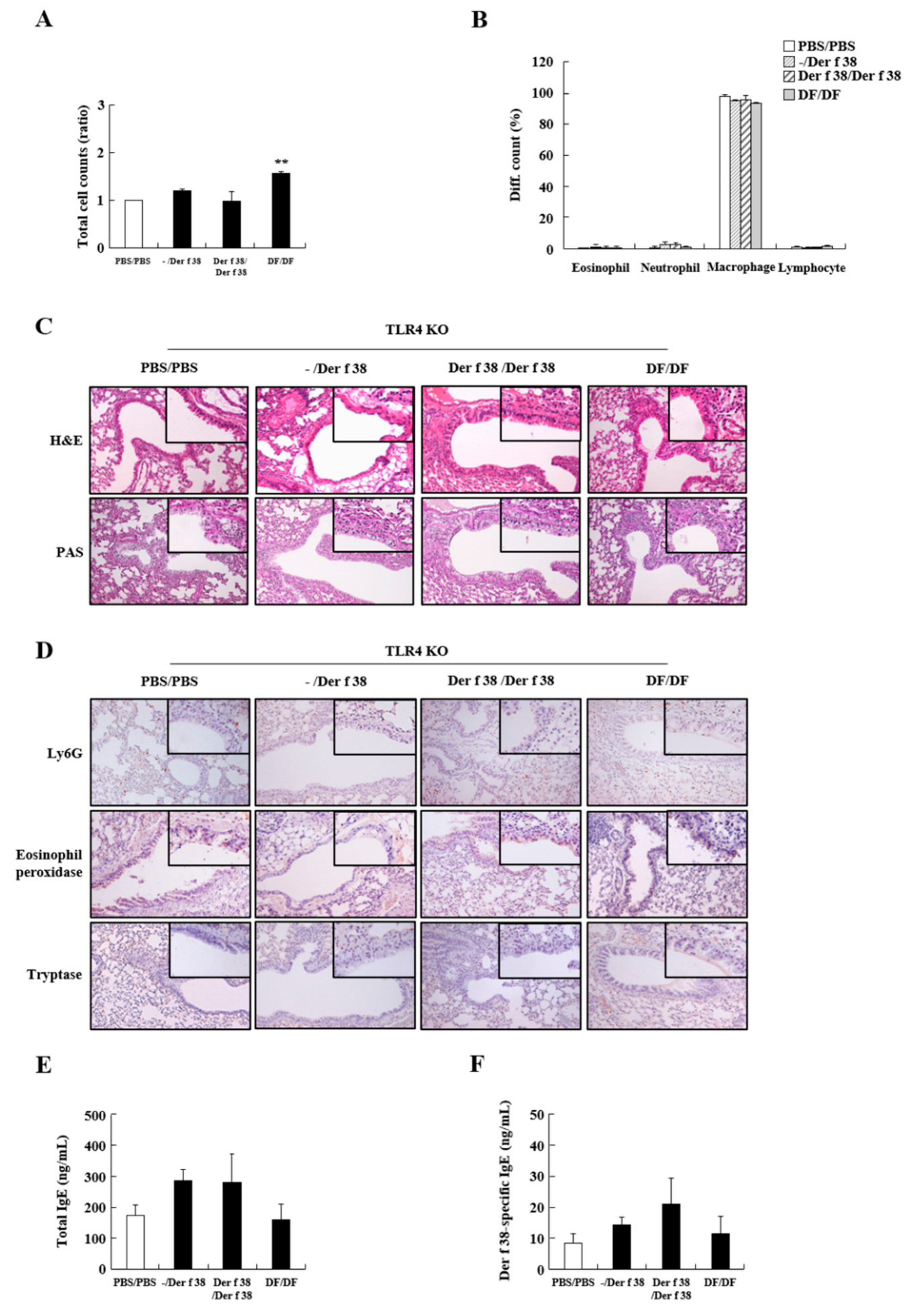
2.3. Der f 38 Induces Airway Inflammation and Increases Total and Der f 38-Specific IgE via TLR4 in Mice
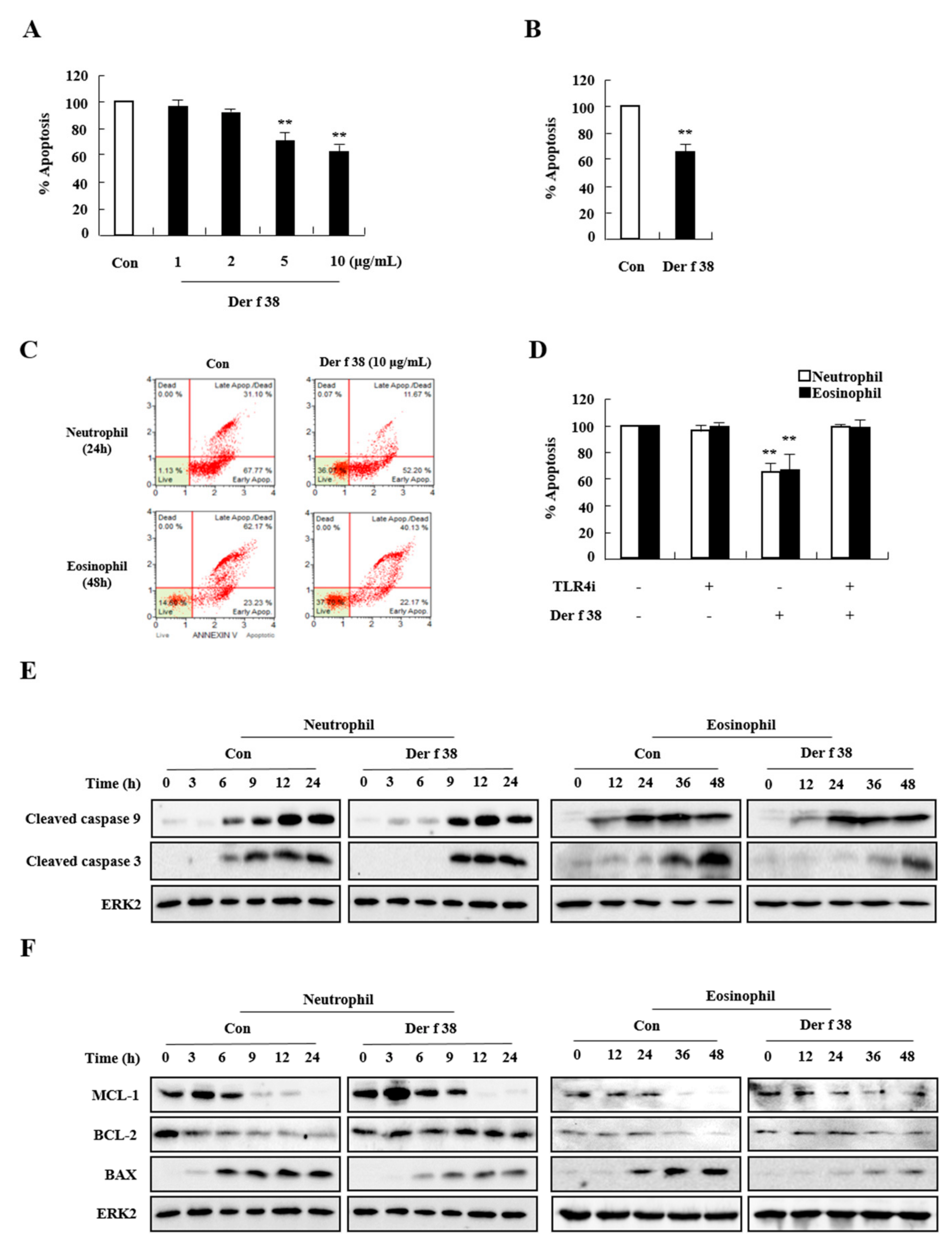
2.4. Der f 38 Suppresses Constitutive Apoptosis of Neutrophils and Eosinophils
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Production of Recombinant Der f 38
4.3. Detection of Native Der f 38 in DF Extract
4.4. Block Preparation for Transmission Electron Microscopy (TEM) and Immuno-Gold Staining for Der f 38 Labeling
4.5. Normal and Allergic Subjects
4.6. Basophil Activation Assay
4.7. Skin Prick Test
4.8. Dot Blot Assay
4.9. Modeling of the Tertiary Structure of Der f 38
4.10. Asthma Induction by Der f 38 Administration in Mice
4.11. Collection of Bronchoalveolar Lavage Fluid (BALF) and Serum
4.12. Detection of Total IgE and Der f 38-Spexific IgE in the Sera
4.13. Histological Analysis
4.14. Biolayer Inteferometry
4.15. Isolation of Neutrophils and Eosinophils and Western Blotting
4.16. Detection of Apoptosis
4.17. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BALF | Bronchoalveolar lavage fluid |
| BLI | Biolayer interferometry |
| DF | Dermatophagoides farinae |
| DP | Dermatophagoides pteronyssinus |
| FITC | fluorescein isothiocyanate |
| HDM | House dust mite |
| H&E | Hematoxylin and eosin |
| IN | Intranasal |
| IP | Intraperitoneal |
| KO | Knockout |
| NOR | NOD-like receptor |
| PAR | Protease-activated receptor |
| PAS | Periodic acid Schiff |
| SDS-PAGE | Sodium dodecyl sulfide-polyacrylamide gel electrophoresis |
| SPF | Specific pathogen-free |
| TEM | transmission electron microscopy |
| Th | T helper |
| TLR | Toll-like receptor |
| WT | wild type |
References
- Jacquet, A. The role of innate immunity activation in house dust mite allergy. Trends Mol. 2011, 17, 604–611. [Google Scholar] [CrossRef]
- Jo, K.M.; Lim, H.K.; Sull, J.W.; Choi, E.; Lee, J.-S.; Cheong, M.A.; Kim, Y.; Kim, I.S. Thymus and activation-regulated chemokine (TARC)/CCL17 and IgE are associated with elderly asthmatics. Immun. Ageing 2018, 15, 13. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.D. The Role of Dust Mites in Allergy. Clin. Rev. Allergy Immunol. 2019, 57, 312–329. [Google Scholar]
- Jacquet, A. The role of the house dust mite-induced innate immunity in development of allergic response. Int. Arch. Allergy Immunol. 2011, 155, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.L.; Liao, E.C.; Yu, S.J. House dust mite allergy: Its innate immune response and immunotherapy. Immunobiology 2018, 223, 300–302. [Google Scholar] [CrossRef]
- Batard, T.; Baron-Bodo, V.; Martelet, A.; Le Mignon, M.; Lemoine, P.; Jain, K.; Mariano, S.; Horiot, S.; Chabre, H.; Harwanegg, C.; et al. Patterns of IgE sensitization in house dust mite-allergic patients: Implications for allergen immunotherapy. Allergy 2016, 71, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Gregory, L.G.; Lloyd, C.M. Orchestrating house dust mite-associated allergy in the lung. Trends Immunol. 2011, 32, 402–411. [Google Scholar] [CrossRef] [Green Version]
- Khweek, A.A.; Kim, E.; Joldrichsen, M.R.; Amer, A.O.; Boyaka, P.N. Insights into mucosal innate immune responses in house dust mite-mediated allergic asthma. Front. Immunol. 2020, 11, 534501. [Google Scholar] [CrossRef] [PubMed]
- Kirtland, M.E.; Tsitoura, D.C.; Durham, S.R.; Shamji, M.H. Toll-Like Receptor Agonists as Adjuvants for Allergen Immunotherapy. Front. Immunol. 2020, 11, 599083. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, S.; Takai, T.; Yashiki, T.; Takahashi, S.; Okumura, K.; Ogawa, H.; Kohda, D.; Hatanaka, H. Lipopolysaccharide binding of the mite allergen Der f 2. Genes Cells 2009, 14, 1055–1065. [Google Scholar] [CrossRef]
- Trompette, A.; Divanovic, S.; Visintin, A.; Blanchard, C.; Hegde, R.S.; Madan, R.; Thorne, P.S.; Wills-Karp, M.; Gioannini, T.L.; Weiss, J.P.; et al. Allergenicity resulting from functional mimicry of a Toll-like receptor complex protein. Nature 2009, 457, 585–588. [Google Scholar] [CrossRef]
- Stremnitzer, C.; Manzano-Szalai, K.; Starkl, P.; Willensdorfer, A.; Schrom, S.; Singer, J.; Reichart, U.; Jarolim, E.J. Epicutaneously applied Der p 2 induces a strong TH 2-biased antibody response in C57BL/6 mice, independent of functional TLR4. Allergy 2014, 69, 741–751. [Google Scholar] [CrossRef]
- Fujimura, T.; Aki, T.; Isobe, T.; Matsuoka, A.; Hayashi, T.; Ono, K.; Kawamoto, S. Der f 35: An MD-2-like house dust mite allergen that cross-reacts with Der f 2 and Pso o 2 Allergen. Allergy 2017, 72, 1728–1736. [Google Scholar] [CrossRef]
- Wang, H.; Lin, J.; Liu, X.; Liang, Z.; Yang, P.; Ran, P.; Liu, Z. Identification of α-tubulin, Der f 33, as a novel allergen from Dermatophagoides farinae. Immunobiology 2016, 221, 911–917. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Sun, X.; Liu, Z.G. Morphology and three-dimensional reconstruction of the digestive system of Dermatophagoides farinae. Int. Arch. Allergy Immunol. 2008, 146, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Jeong, K.Y.; Lee, I.Y.; Ree, H.I.; Hong, C.S.; Yong, T.S. Localization of Der f 2 in the gut and fecal pellets of Dermatophagoides farinae. Allergy 2002, 57, 729–731. [Google Scholar] [CrossRef]
- Chan, T.F.; Ji, K.M.; Yim, A.K.; Liu, X.Y.; Zhou, J.W.; Li, R.Q.; Yang, K.Y.; Li, J.; Li, M.; Law, P.T.; et al. The draft genome, transcriptome, and microbiome of Dermatophagoides farinae reveal a broad spectrum of dust mite allergens. J. Allergy Clin. Immunol. 2015, 135, 539–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.Y.; Yang, K.Y.; Wang, M.Q.; Kwok, J.S.; Zeng, X.; Yang, A.; Xiao, X.J.; Lau, C.P.; Huang, Z.M.; Ba, J.G.; et al. High-quality assembly of Dermatophagoides pteronyssinus genome and transcriptome reveals a wide range of novel allergens. J. Allergy Clin. Immunol. 2018, 141, 2268–2271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, S.; Chen, L.; Long, C.; Liu, X.; Xu, X.; Lu, X.; Rong, M.; Liu, Z.; Lai, R. Dermatophagoides farinae allergens diversity identification by proteomics. Mol. Cell Proteom. 2013, 12, 1818–1828. [Google Scholar] [CrossRef] [Green Version]
- An, S.; Shen, C.; Liu, X.; Chen, L.; Xu, X.; Rong, M.; Liu, Z.; Lai, R. Alpha-actinin is a new type of house dust mite allergen. PLoS ONE 2013, 8, e81377. [Google Scholar]
- Pang, S.L.; Ho, K.L.; Waterman, J.; Rambo, R.P.; The, A.H.; Mathavan, I.; Harris, G.; Beis, K.; Say, Y.H.; Anusha, M.S. Crystal structure and epitope analysis of house dust mite allergen Der f 21. Sci. Rep. 2019, 9, 4933. [Google Scholar] [CrossRef]
- Glesner, J.; Kapingidza, A.B.; Godzwon, M.; Offermann, L.R.; Mueller, G.A.; DeRose, E.F.; Wright, P.; Richardson, C.M.; Woodfolk, J.A.; Vailes, L.D.; et al. A Human IgE Antibody Binding Site on Der p 2 for the Design of a Recombinant Allergen for Immunotherapy. J. Immunol. 2019, 203, 2545–2556. [Google Scholar] [CrossRef]
- Wu, Y.; Jiang, C.; Li, M.; Yu, H.; Xiao, X.; Fan, X.; Lin, J.; Liu, X.; Zhang, M.; Yang, P.; et al. Der f 21, a novel allergen from Dermatophagoides farinae. Am. J. Transl. Res. 2016, 8, 49–59. [Google Scholar]
- Ansotegui, I.J.; Melioli, G.; Canonica, G.W.; Caraballo, L.; Villa, E.; Ebisawa, M.; Passalacqua, G.; Savi, E.; Ebo, D.; Gomez, R.M.; et al. IgE allergy diagnostics and other relevant tests in allergy. World Allergy Organ. J. 2020, 13, 100080. [Google Scholar] [CrossRef] [PubMed]
- Resch, Y.; Blatt, K.; Malkus, U.; Fercher, C.; Swoboda, I.; Focke-Tejkl, M.; Chen, K.W.; Seiberler, S.; Mittermann, I.; Lupinek, C.; et al. Molecular, Structural and Immunological Characterization of Der p 18, a Chitinase-Like House Dust Mite Allergen. PLoS ONE 2016, 11, e0160641. [Google Scholar]
- Bouley, J.; Groeme, R.; Le Mignon, M.; Jain, K.; Chabre, H.; Bordas-Le, F.; Couret, M.N.; Bussières, L.; Lautrette, A.; Naveau, M.; et al. Identification of the cysteine protease Amb a 11 as a novel major allergen from short ragweed. J. Allergy Clin. Immunol. 2015, 136, 1055–1064. [Google Scholar] [CrossRef] [Green Version]
- ElRamlawy, K.G.; Fujimura, T.; Aki, T.; Okada, A.; Suzuki, T.; Abe, T.; Hayashi, T.; Epton, M.J.; Thomas, W.R.; Rafeet, I.H.; et al. Prominent IgE-binding and cytokine-inducing capacities of a newly cloned N-terminal region of Der f 14, an apolipophorin-like house dust mite allergen. J. Biochem. 2018, 163, 51–60. [Google Scholar] [CrossRef]
- Kianmeher, M.; Ghorani, V.; Boskabady, M.H. Animal Model of Asthma, Various Methods and Measured Parameters: A Methodological Review. Iran. J. Allergy Asthma Immunol. 2016, 15, 445–465. [Google Scholar] [PubMed]
- Yasuda, Y.; Nagano, T.; Kobayashi, K.; Nishimura, Y. Group 2 Innate Lymphoid Cells and the House Dust Mite-Induced Asthma Mouse Model. Cells 2020, 9, 1178. [Google Scholar] [CrossRef]
- Lee, N.R.; Park, B.S.; Kim, S.Y.; Gu, A.; Kim, D.H.; Lee, J.S.; Kim, I.S. Cytokine secreted by S100A9 via TLR4 in monocytes delays neutrophil apoptosis by inhibition of caspase 9/3 pathway. Cytokine 2016, 86, 53–63. [Google Scholar] [CrossRef] [PubMed]
- El Kebir, D.; Filep, J.G. Modulation of Neutrophil Apoptosis and the Resolution of Inflammation through β2 Integrins. Front. Immunol. 2013, 4, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, I.S.; Kim, M.J.; Kim, D.H.; Choi, E.; Lee, J.S. Different anti-apoptotic effects of normal and asthmatic serum on normal eosinophil apoptosis depending on house dust mite-specific IgE. Mol. Biol. Rep. 2013, 40, 5875–5881. [Google Scholar] [CrossRef]
- Barry, J.; Loh, Z.; Collison, A.; Mazzone, S.; Lalwani, A.; Zhang, V.; Dacidson, S.; Wybacz, E.; Gralanda, C.; Mantovani, A.; et al. Absence of Toll-IL-1 receptor 8/single immunoglobulin IL-1 receptor-related molecule reduces house dust mite-induced allergic airway inflammation in mice. Am. J. Respir. Cell. Mol. Biol. 2013, 49, 481–490. [Google Scholar] [CrossRef]
- Ullah, M.A.; Loh, Z.; Gan, W.J.; Zhang, V.; Yang, H.; Li, J.H.; Yamamoto, Y.; Schmidt, A.M.; Armour, C.L.; Hughes, J.M.; et al. Receptor for advanced glycation end products and its ligand high-mobility group box-1 mediate allergic airway sensitization and airway inflammation. J. Allergy Clin. Immunol. 2014, 134, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.; Nagler, C. TLR4 signaling regulates an inflammatory response to food allergens. J. Immunol. 2019, 202 (Suppl. 1), 26895–26906. [Google Scholar]
- Lucía, C.J.; María, A.D.P.; María, L.R.; Joaquín, S.; Pablo, M.; Ignacio, M.F.; Selene, B.; Blanca, C. Prioritizing molecular biomarkers in asthma and respiratory allergy using systems biology. Front. Immuol. 2021, 12, 640791. [Google Scholar]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Saradna, A.; Do, D.C.; Kumar, S.; Fu, Q.L.; Gao, P. Macrophage polarization and allergic asthma. Transl. Res. 2018, 191, 1–14. [Google Scholar] [CrossRef]
- Panda, S.K.; Colonna, M. Innate lymphoid cells in mucosal immunity. Front. Immunol. 2019, 10, 861. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Zhang, J.; Toki, S.; Goleniewska, K.; Norlander, A.E.; Newcomb, D.C.; Wu, P.; Boyd, K.L.; Kita, H.; Strokes Peebles Jr, R. COX inhibition increases Alternaria-Induced pulmonary group 2 innate lymphoid cell responses and IL-33 release in mice. J. Immunol. 2020, 205, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Mueller, G.A.; Glesner, J.; Daniel, J.L.; Zhang, J.; Hyduke, N.; Richardson, C.M.; De Rose, E.F.; Chapman, M.D.; Peebles, R.S., Jr.; Smith, S.A.; et al. Mapping Human Monoclonal IgE Epitopes on the Major Dust Mite Allergen Der p 2. J. Immunol. 2020, 205, 1999–2007. [Google Scholar] [CrossRef] [PubMed]
- Shamji, M.H.; Durham, S.R. Mechanisms of allergen immunotherapy for inhaled allergens and predictive biomarkers. J. Allergy Clin. Immunol. 2017, 140, 1485–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stranzl, T.; Ipsen, H.; Christensen, L.H.; Eiwegger, T.; Johansen, N.; Lund, K.; Andersen, P.S. Limited impact of Der p 23 IgE on treatment outcomes in tablet allergy immunotherapy phase III study. Allergy 2020, 76, 1235–1238. [Google Scholar] [CrossRef] [Green Version]
- Satitsuksanoa, P.; Głobińska, A.; Jansen, K.; van de Veen, W.; Akdis, M. Modified Allergens for Immunotherapy. Curr. Allergy Asthma Rep. 2018, 18, 9. [Google Scholar] [CrossRef]
- Kim, D.H.; Gu, A.; Lee, J.S.; Yang, E.J.; Kashif, A.; Hong, M.H.; Kim, G.Y.; Park, B.S.; Lee, S.J.; Kim, I.S. Suppressive effects of S100A8 and S100A9 on neutrophil apoptosis by cytokine release of human bronchial epithelial cells in asthma. Int. J. Med. Sci. 2020, 17, 498–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.S.; Lee, N.R.; Kahif, A.; Yang, S.J.; Nam, A.R.; Song, I.C.; Gong, S.J.; Hong, M.H.; Kim, G.Y.; Seok, P.R.; et al. S100A8 and S100A9 promote apoptosis of chronic eosinophilic leukemia cells. Front. Immuol. 2020, 11, 1258. [Google Scholar] [CrossRef] [PubMed]
- Hosaka, Y.; Araya, J.; Fujita, Y.; Kadota, T.; Tsubouchi, K.; Yoshida, M.; Minagawa, S.; Hara, H.; Kawamoto, H.; Watanabe, N.; et al. Chaperone-Mediated Autophagy Suppresses Apoptosis via Regulation of the Unfolded Protein Response during Chronic Obstructive Pulmonary Disease Pathogenesis. J. Immunol. 2020, 205, 1256–1267. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal (n = 20) | Allergy (n = 20) | |
|---|---|---|
| DP+, DF+ | 6 (30%) | 20 (100%) |
| DP+, DF− | 0 (0%) | 0 (0%) |
| DP−, DF+ | 2 (10%) | 0 (0%) |
| DP−, DF− | 12 (60%) | 0 (0%) |
| Der f 38 | 2 (10%) | 12 (60%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, G.; Hong, M.; Kashif, A.; Hong, Y.; Park, B.-S.; Mun, J.-Y.; Choi, H.; Lee, J.-S.; Yang, E.-J.; Woo, R.-S.; et al. Der f 38 Is a Novel TLR4-Binding Allergen Related to Allergy Pathogenesis from Dermatophagoides farinae. Int. J. Mol. Sci. 2021, 22, 8440. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168440
Kim G, Hong M, Kashif A, Hong Y, Park B-S, Mun J-Y, Choi H, Lee J-S, Yang E-J, Woo R-S, et al. Der f 38 Is a Novel TLR4-Binding Allergen Related to Allergy Pathogenesis from Dermatophagoides farinae. International Journal of Molecular Sciences. 2021; 22(16):8440. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168440
Chicago/Turabian StyleKim, Geunyeong, Minhwa Hong, Ayesha Kashif, Yujin Hong, Beom-Seok Park, Ji-Young Mun, Hyosun Choi, Ji-Sook Lee, Eun-Ju Yang, Ran-Sook Woo, and et al. 2021. "Der f 38 Is a Novel TLR4-Binding Allergen Related to Allergy Pathogenesis from Dermatophagoides farinae" International Journal of Molecular Sciences 22, no. 16: 8440. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168440