
Chromophore of an Enhanced Green Fluorescent Protein Can Play a Photoprotective Role Due to Photobleaching

 , , , and
, , , and 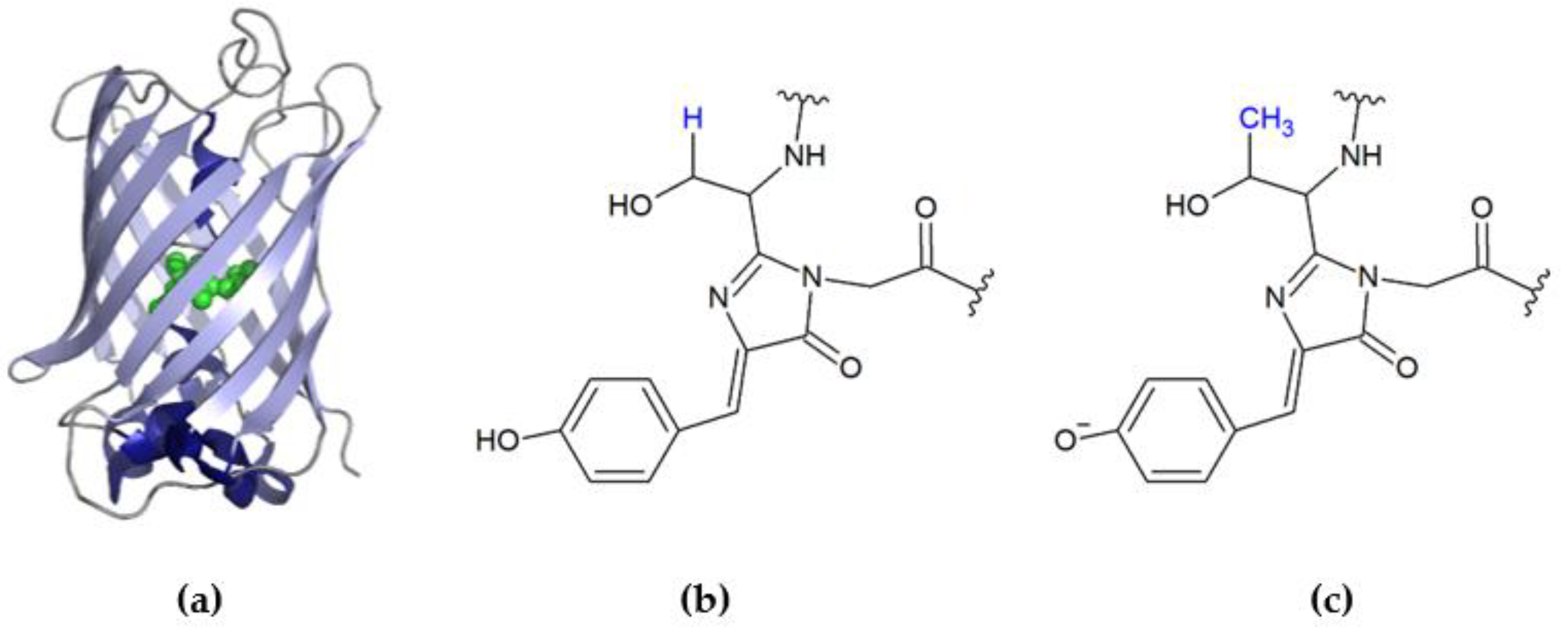{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
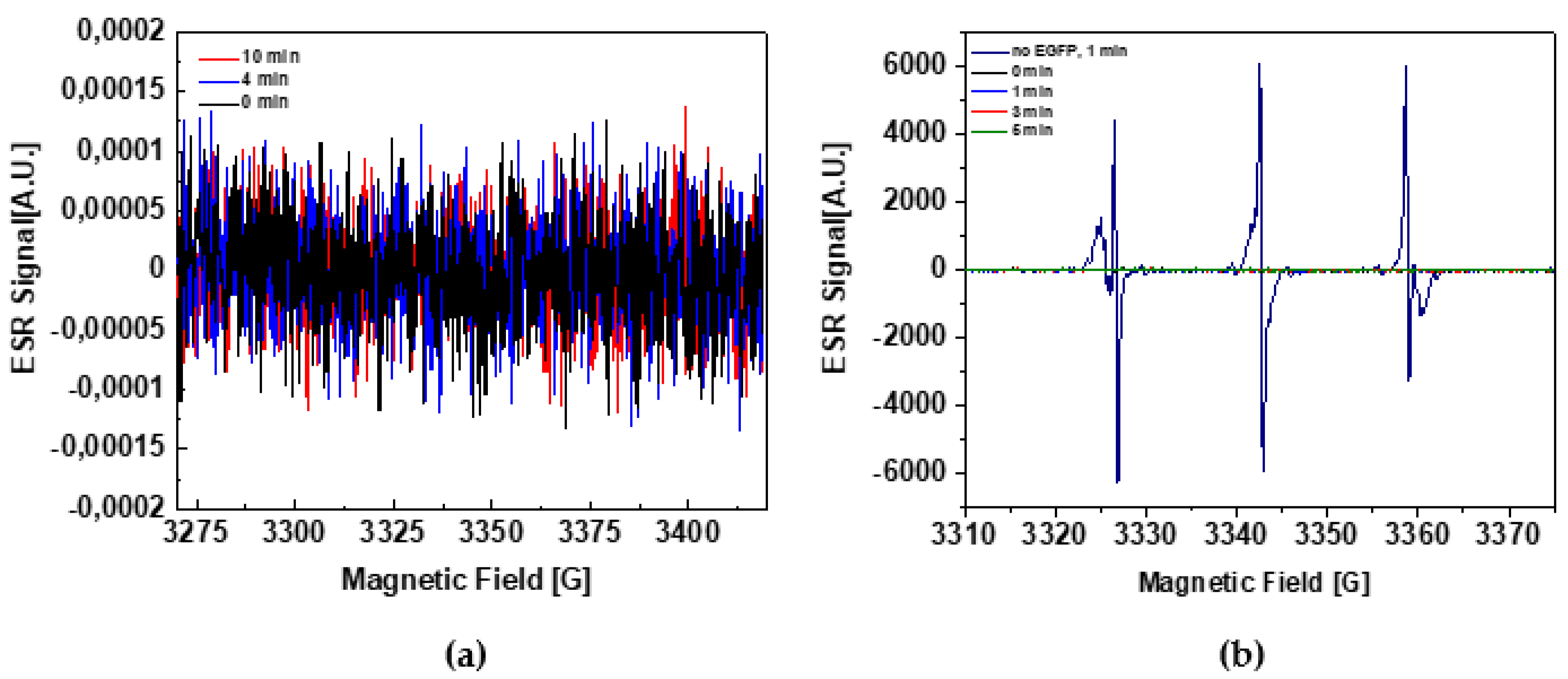
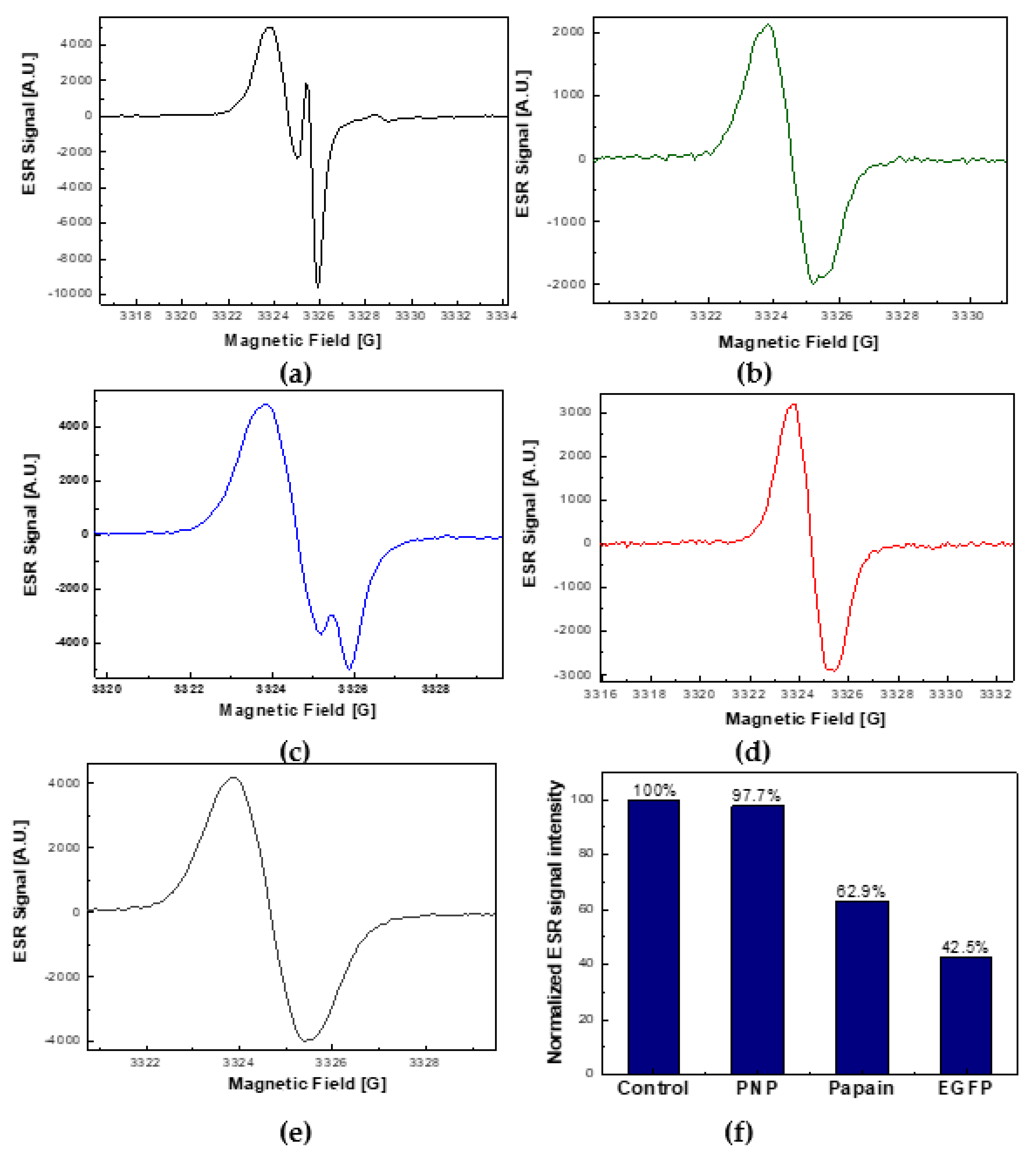
2.1. EGFP Is Not a ROS Generator
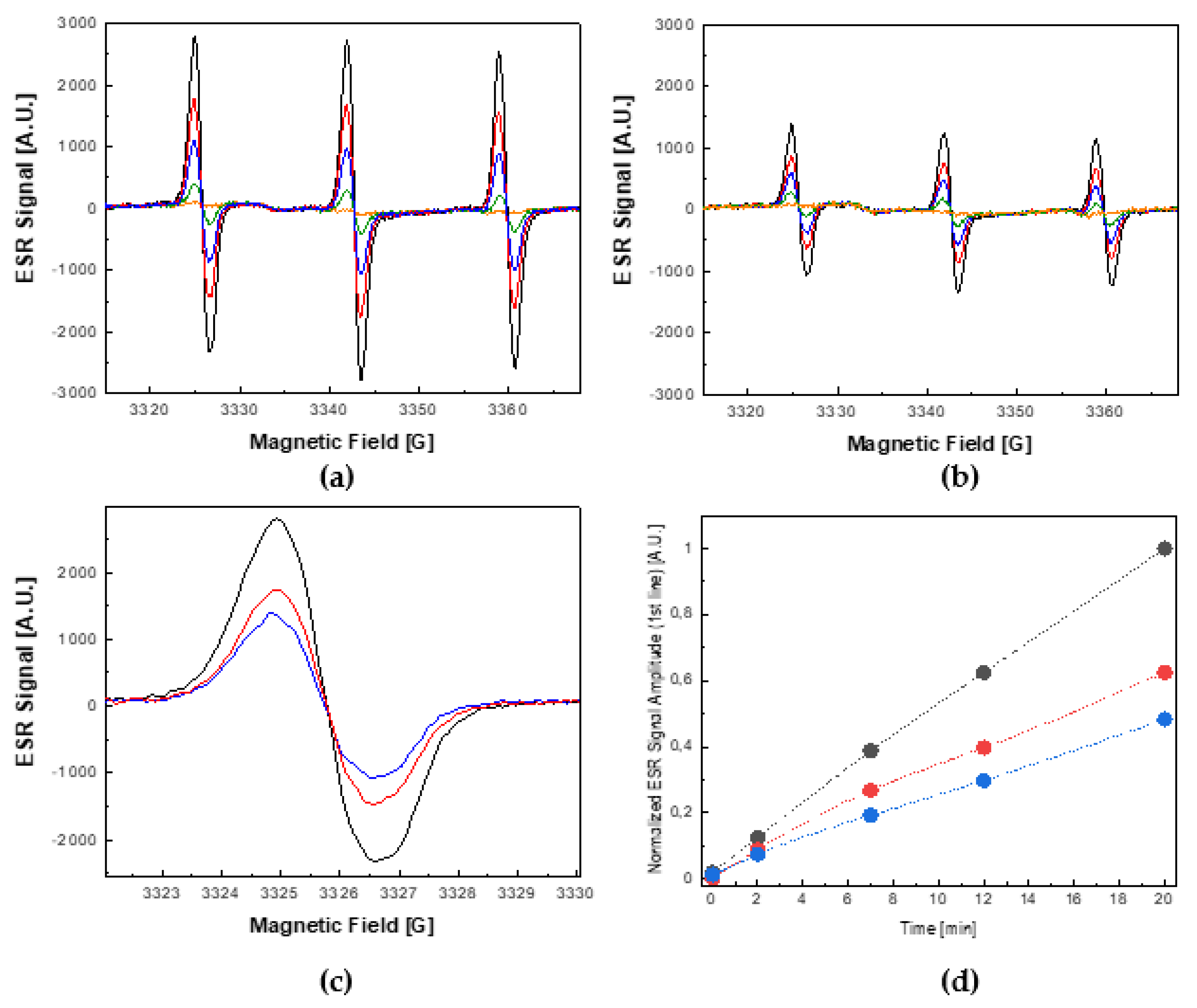
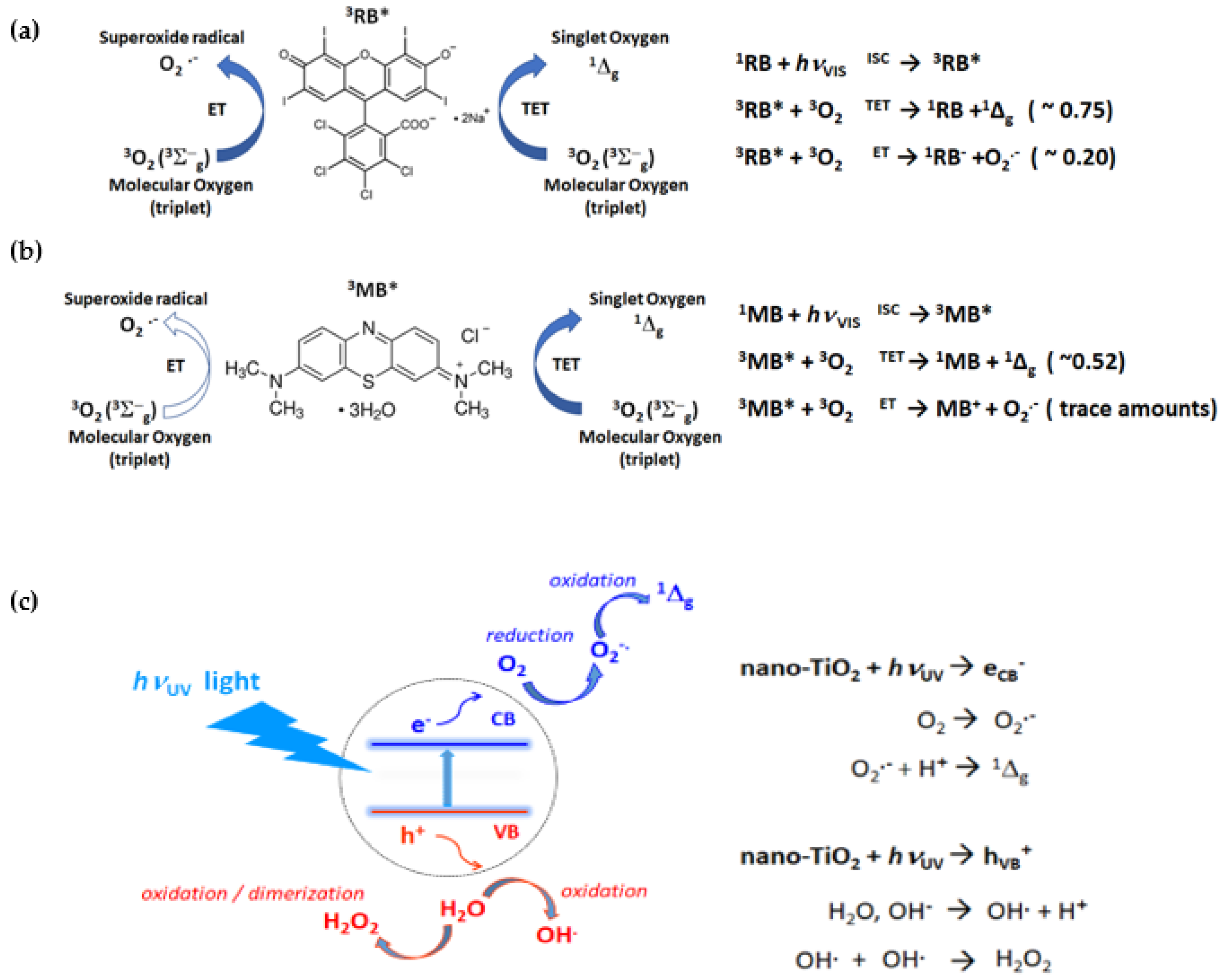
2.2. ROS Quenching by EGFP
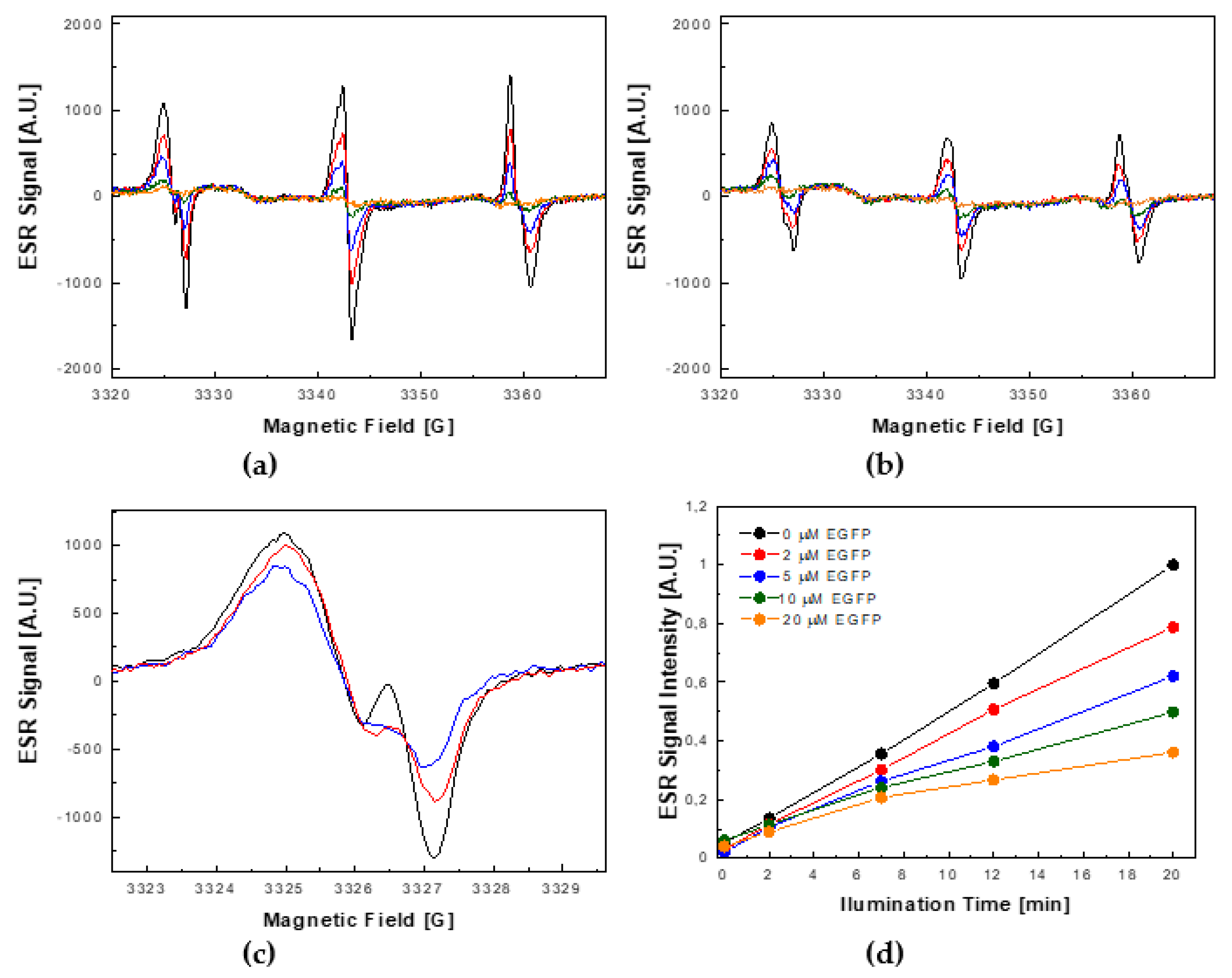
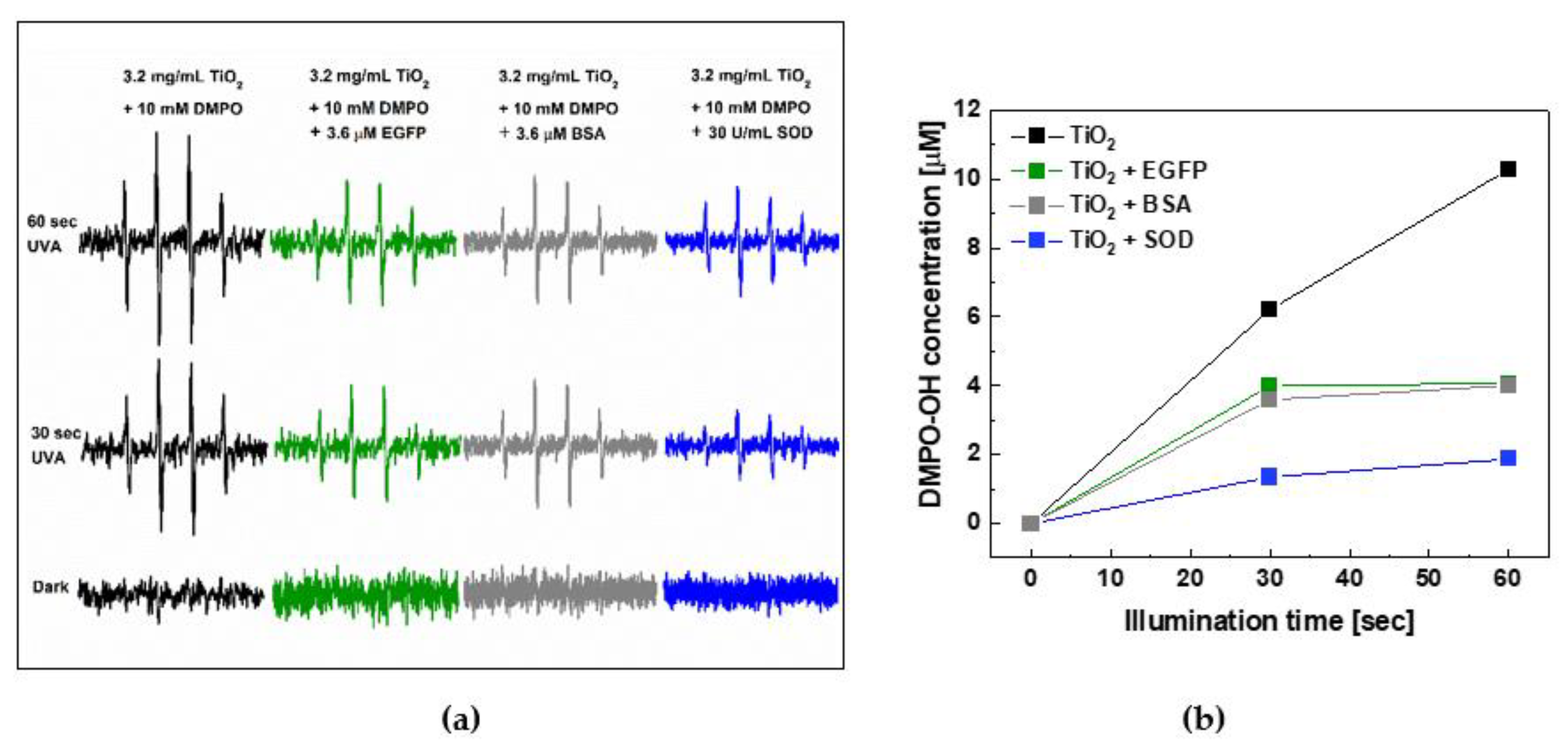
2.3. Photoprotective Role of EGFP in the Presence of Photocatalytically Generated ROS by Nano-TiO2
2.4. The Comparison of ROS Quenching Abilities of EGFP with PNP and Papain
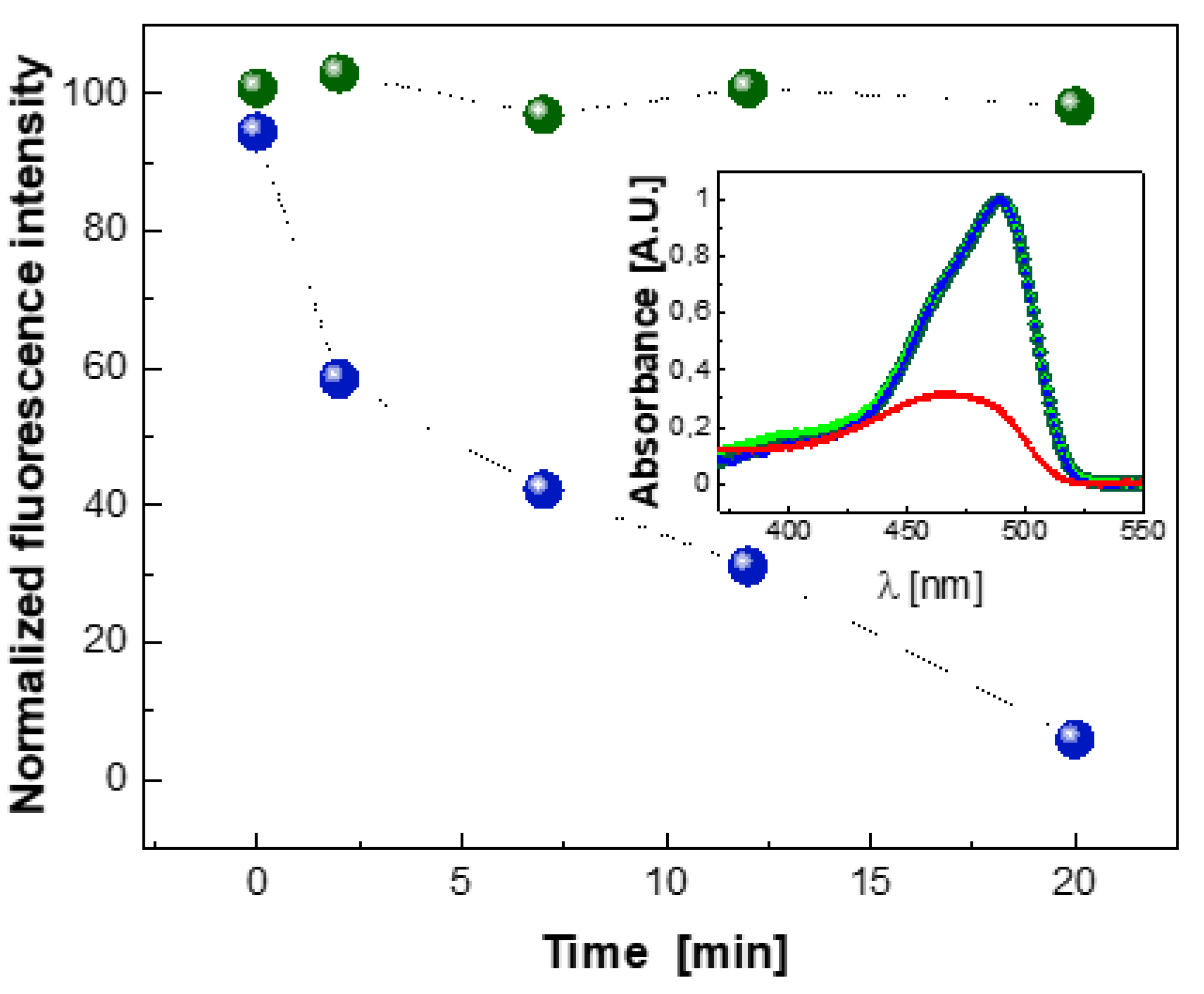
2.5. Photobleaching of EGFP Chromophore
3. Discussion
4. Materials and Methods
4.1. Chemicals and Protein
4.2. Electron Spin Resonance Spectroscopy
4.3. Photobleaching of EGFP
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Properties of the Employed ROS Photogenerators and Spin-Traps


References
- Chauhan, V.; Chauhan, A. Oxidative stress in Alzheimer’s disease. Pathophysiology 2006, 13, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Uttara, B.; Singh, A.V.; Zamboni, P.; Mahajan, R.T. Oxidative Stress and Neurodegenerative Diseases: A Review of Upstream and Downstream Antioxidant Therapeutic Options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Migliore, L.; Coppedè, F. Environmental-induced oxidative stress in neurodegenerative disorders and aging. Mutat. Res. Toxicol. Environ. Mutagen. 2009, 674, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Koppula, S.; Kumar, H.; More, S.V.; Kim, B.W.; Kim, I.S.; Choi, D.-K. Recent Advances on the Neuroprotective Potential of Antioxidants in Experimental Models of Parkinson’s Disease. Int. J. Mol. Sci. 2012, 13, 10608–10629. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Wagner, J.R. DNA Base Damage by Reactive Oxygen Species, Oxidizing Agents, and UV Radiation. Cold Spring Harb. Perspect. Biol. 2013, 5, a012559. [Google Scholar] [CrossRef]
- Aslani, B.A.; Ghobadi, S. Studies on oxidants and antioxidants with a brief glance at their relevance to the immune system. Life Sci. 2016, 146, 163–173. [Google Scholar] [CrossRef]
- Roth, M.S.; Latz, M.I.; Goericke, R.; Deheyn, D.D. Green fluorescent protein regulation in the coral Acropora yongei during photoacclimation. J. Exp. Biol. 2010, 213, 3644–3655. [Google Scholar] [CrossRef] [Green Version]
- Gittins, J.R.; D’Angelo, C.; Oswald, F.; Edwards, R.; Wiedenmann, J. Fluorescent protein-mediated colour polymorphism in reef corals: Multicopy genes extend the adaptation/acclimatization potential to variable light environments. Mol. Ecol. 2015, 24, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Palmer, C.V.; Modi, C.; Mydlarz, L.D. Coral Fluorescent Proteins as Antioxidants. PLoS ONE 2009, 4, e7298. [Google Scholar] [CrossRef] [Green Version]
- Leutenegger, A.; D’Angelo, C.; Matz, M.; Denzel, A.; Oswald, F.; Salih, A.; Nienhaus, G.U.; Wiedenmann, J. It’s cheap to be colorful Anthozoans show a slow turnover of GFP-like proteins. FEBS J. 2007, 274, 2496–2505. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, O.; Johnson, F.H.; Saiga, Y. Extraction, Purification and Properties of Aequorin, a Bioluminescent Protein from the Luminous Hydromedusan,Aequorea. J. Cell. Comp. Physiol. 1962, 59, 223–239. [Google Scholar] [CrossRef]
- Van Thor, J.J.; Georgiev, G.Y.; Towrie, M.; Sage, J.T. Ultrafast and Low Barrier Motions in the Photoreactions of the Green Fluorescent Protein. J. Biol. Chem. 2005, 280, 33652–33659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royant, A.; Noirclerc-Savoye, M. Stabilizing role of glutamic acid 222 in the structure of Enhanced Green Fluorescent Protein. J. Struct. Biol. 2011, 174, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Sun, Q.; Smith, S.C. The mechanism of oxidation in chromophore maturation of wild-type green fluorescent protein: A theoretical study. Phys. Chem. Chem. Phys. 2017, 19, 12942–12952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartkiewicz, M.; Kazazic, S.; Krasowska, J.; Clark, P.; Wielgus-Kutrowska, B.; Bzowska, A. Non–fluorescent mutant of green fluorescent protein sheds light on the mechanism of chromophore formation. FEBS Lett. 2018, 592, 1516–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsien, R.Y. The Green Fluorescent Protein. Annu. Rev. Biochem. 1998, 67, 509–544. [Google Scholar] [CrossRef] [PubMed]
- Reid, B.G.; Flynn, G.C. Chromophore Formation in Green Fluorescent Protein. Biochemistry 1997, 36, 6786–6791. [Google Scholar] [CrossRef]
- Cormack, B.P.; Valdivia, R.H.; Falkow, S. FACS-optimized mutants of the green fluorescent protein (GFP). Gene 1996, 173, 33–38. [Google Scholar] [CrossRef]
- Crone, D.E.; Huang, Y.-M.; Pitman, D.J.; Schenkelberg, C.; Fraser, K.; Macari, S.; Bystroff, C. GFP-Based Biosensors. In State of the Art in Biosensors—General Aspects; IntechOpen: London, UK, 2013. [Google Scholar]
- Dawidziak-Pakula, A.; Krasowska, J.; Wielgus-Kutrowska, B. Analytical ultracentrifugation as a tool in the studies of aggregation of the fluorescent marker, Enhanced Green Fluorescent Protein. Acta Biochim. Pol. 2020, 67, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Seifert, M.H.J.; Ksiazek, D.; Azim, M.K.; Smialowski, P.; Budisa, N.; Holak, T.A. Slow Exchange in the Chromophore of a Green Fluorescent Protein Variant. J. Am. Chem. Soc. 2002, 124, 7932–7942. [Google Scholar] [CrossRef] [PubMed]
- Patterson, G.; Knobel, S.; Sharif, W.; Kain, S.; Piston, D. Use of the green fluorescent protein and its mutants in quantitative fluorescence microscopy. Biophys. J. 1997, 73, 2782–2790. [Google Scholar] [CrossRef] [Green Version]
- Marshall, J.; Molloy, R.; Moss, G.W.; Howe, J.R.; Hughes, T. The jellyfish green fluorescent protein: A new tool for studying ion channel expression and function. Neuron 1995, 14, 211–215. [Google Scholar] [CrossRef] [Green Version]
- Bulina, M.E.; Chudakov, D.M.; Britanova, O.V.; Yanushevich, Y.G.; Staroverov, D.B.; Chepurnykh, T.V.; Merzlyak, E.M.; Shkrob, M.A.; Lukyanov, S.; Lukyanov, K.A. A genetically encoded photosensitizer. Nat. Biotechnol. 2006, 24, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Serebrovskaya, E.O.; Edelweiss, E.F.; Stremovskiy, O.A.; Lukyanov, K.; Chudakov, D.; Deyev, S.M. Targeting cancer cells by using an antireceptor antibody-photosensitizer fusion protein. Proc. Natl. Acad. Sci. USA 2009, 106, 9221–9225. [Google Scholar] [CrossRef] [Green Version]
- Vegh, R.B.; Solntsev, K.; Kuimova, M.K.; Cho, S.; Liang, Y.; Loo, B.L.W.; Tolbert, L.M.; Bommarius, A.S. Reactive oxygen species in photochemistry of the red fluorescent protein “Killer Red”. Chem. Commun. 2011, 47, 4887–4889. [Google Scholar] [CrossRef]
- Carpentier, P.; Violot, S.; Blanchoin, L.; Bourgeois, D. Structural basis for the phototoxicity of the fluorescent protein KillerRed. FEBS Lett. 2009, 583, 2839–2842. [Google Scholar] [CrossRef] [PubMed]
- Riuz-Gonzalez, R.; Rodriguez-Pulido, A. Photosensitization by GFP-Like Proteins. In Singet Oxygen. Applications in Biosciences and Nanosciences, Comprehensive Series in Photochemistry and Photobiology; Nonell, S., Flors, C., Eds.; The Royal Society of Chemistry: Cambridge, UK, 2013; Volume 13, pp. 274–284. [Google Scholar]
- Ragàs, X.; Cooper, L.P.; White, J.H.; Nonell, S.; Flors, C. Quantification of Photosensitized Singlet Oxygen Production by a Fluorescent Protein. Chem. Phys. Chem. 2010, 12, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, K.; Matsuda, T.; Sakai, N.; Fu, D.; Noda, M.; Uchiyama, S.; Kotera, I.; Arai, Y.; Horiuchi, M.; Fukui, K.; et al. SuperNova, a monomeric photosensitizing fluorescent protein for chromophore-assisted light inactivation. Sci. Rep. 2013, 3, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Mccord, J.M.; Fridovich, I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [CrossRef]
- Marshall, D.; Christian, M.L.; Gryn’Ova, G.; Coote, M.; Barker, P.; Blanksby, S. Oxidation of 4-substituted TEMPO derivatives reveals modifications at the 1- and 4-positions. Org. Biomol. Chem. 2011, 9, 4936–4947. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Branicky, R.; Noe, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Iglesias, J.; Abernethy, V.E.; Wang, Z.; Lieberthal, W.; Koh, J.S.; Levine, J.S. Albumin is a major serum survival factor for renal tubular cells and macrophages through scavenging of ROS. Am. J. Physiol. Physiol. 1999, 277, F711–F722. [Google Scholar] [CrossRef]
- Roche, M.; Rondeau, P.; Singh, N.R.; Tarnus, E.; Bourdon, E. The antioxidant properties of serum albumin. FEBS Lett. 2008, 582, 1783–1787. [Google Scholar] [CrossRef] [PubMed]
- Manosroi, A.; Chankhampan, C.; Pattamapun, K.; Manosroi, W.; Manosroi, J. Antioxidant and Gelatinolytic Activities of Papain from Papaya Latex and Bromelain from Pineapple Fruits. Chiang Mai. J. Sci. 2014, 41, 635–648. [Google Scholar]
- Baumann, P.; Balasubramanian, V.; Onaca-Fischer, O.; Sienkiewicz, A.; Palivan, C.G. Light-responsive polymer nanoreactors: A source of reactive oxygen species on demand. Nanoscale 2013, 5, 217–224. [Google Scholar] [CrossRef]
- Salih, A.; Larkum, A.W.D.; Cox, G.; Kühl, M.; Hoegh-Guldberg, O. Fluorescent pigments in corals are photoprotective. Nat. Cell Biol. 2000, 408, 850–853. [Google Scholar] [CrossRef]
- Bou-Abdallah, F.; Chasteen, N.D.; Lesser, M.P. Quenching of superoxide radicals by green fluorescent protein. Biochim. Biophys. Acta (BBA) Gen. Subj. 2006, 1760, 1690–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenbaum, L.; Rothmann, C.; Lavie, R.; Malik, Z. Green Fluorescent Protein Photobleaching: A Model for Protein Damage by Endogenous and Exogenous Singlet Oxygen. Biol. Chem. 2000, 381, 1251–1258. [Google Scholar] [CrossRef] [PubMed]
- Grigorenko, B.; Nemukhin, A.V.; Polyakov, I.; Khrenova, M.; Krylov, A.I. A Light-Induced Reaction with Oxygen Leads to Chromophore Decomposition and Irreversible Photobleaching in GFP-Type Proteins. J. Phys. Chem. B 2015, 119, 5444–5452. [Google Scholar] [CrossRef]
- Jiménez-Banzo, A.; Nonell, S.; Hofkens, J.; Flors, C. Singlet Oxygen Photosensitization by EGFP and its Chromophore HBDI. Biophys. J. 2008, 94, 168–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Banzo, A.; Ragàs, X.; Abbruzzetti, S.; Viappiani, C.; Campanini, B.; Flors, C.; Nonell, S. Singlet oxygen photosensitisation by GFP mutants: Oxygen accessibility to the chromophore. Photochem. Photobiol. Sci. 2010, 9, 1336–1341. [Google Scholar] [CrossRef]
- Ganini, D.; Leinisch, F.; Kumar, A.; Jiang, J.; Tokar, E.J.; Malone, C.C.; Petrovich, R.M.; Mason, R.P. Fluorescent proteins such as eGFP lead to catalytic oxidative stress in cells. Redox Biol. 2017, 12, 462–468. [Google Scholar] [CrossRef]
- Alnuami, A.A.; Zeedi, B.; Qadri, S.; Ashraf, S. Oxyradical-induced GFP damage and loss of fluorescence. Int. J. Biol. Macromol. 2008, 43, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Pey, A.; Zamoum, T.; Christen, R.; Merle, P.-L.; Furla, P. Characterization of glutathione peroxidase diversity in the symbiotic sea anemone Anemonia viridis. Biochimie 2017, 132, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Cass, A.E.G. Superoxide Dismutases. In Metalloproteins, Topics in Molecular and Structural Biology Book Series, 1st ed.; Palgrave Macmillan: London, UK, 1985; pp. 121–156. [Google Scholar]
- Peters, T. Serum Albumin. Adv. Protein Chem. 1985, 37, 161–245. [Google Scholar] [CrossRef] [PubMed]
- Mitchel, R.E.; Chaiken, I.M.; Smith, E.L. The Complete Amino Acid Sequence of Papain. J. Biol. Chem. 1970, 245, 3485–3492. [Google Scholar] [CrossRef]
- Lee, J.; Filosa, S.; Bonvin, J.; Guyon, S.; Aponte, R.A.; Turnbull, J.L. Expression, Purification, and Characterization of Recombinant Purine Nucleoside Phosphorylase from Escherichia coli. Protein Expr. Purif. 2001, 22, 180–188. [Google Scholar] [CrossRef]
- Mikleušević, G.; Štefanić, Z.; Narczyk, M.; Wielgus-Kutrowska, B.; Bzowska, A.; Luic, M. Validation of the catalytic mechanism of Escherichia coli purine nucleoside phosphorylase by structural and kinetic studies. Biochimie 2011, 93, 1610–1622. [Google Scholar] [CrossRef]
- Pace, C.N.; Vajdos, F.; Fee, L.; Grimsley, G.; Gray, T. How to measure and predict the molar absorption coefficient of a protein. Protein Sci. 1995, 4, 2411–2423. [Google Scholar] [CrossRef] [Green Version]
- Bzowska, A.; Kulikowska, E.; Shugar, D. Purine nucleoside phosphorylases: Properties, functions, and clinical aspects. Pharmacol. Ther. 2000, 88, 349–425. [Google Scholar] [CrossRef]
- Krasowska, J.; Olasek, M.; Bzowska, A.; Clark, P.L.; Wielgus-Kutrowska, B. The comparison of aggregation and folding of enhanced green fluorescent protein (EGFP) by spectroscopic studies. Spectroscopy 2010, 24, 343–348. [Google Scholar] [CrossRef]
- Buettner, G. The Spin Trapping of Superoxide and Hydroxyl Free Radicals with DMPO (5.5-Dimethylpyrroline-N-oxide): More About Iron. Free. Radic. Res. Commun. 1993, 19, s79–s87. [Google Scholar] [CrossRef] [Green Version]
- Ruales-Lonfat, C.; Barona, J.; Sienkiewicz, A.; Bensimon, M.; Colmenares, J.J.V.; Benítez, N.; Pulgarín, C. Iron oxides semiconductors are efficients for solar water disinfection: A comparison with photo-Fenton processes at neutral pH. Appl. Catal. B Environ. 2015, 166-167, 497–508. [Google Scholar] [CrossRef]
- Caputo, F.; Mameli, M.; Sienkiewicz, A.; Licoccia, S.; Stellacci, F.; Ghibelli, L.; Traversa, E. A novel synthetic approach of cerium oxide nanoparticles with improved biomedical activity. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lambert, C.; Sarna, T.; Truscott, T.G. Rose bengal radicals and their reactivity. J. Chem. Soc. Faraday Trans. 1990, 86, 3879–3882. [Google Scholar] [CrossRef]
- Hubenko, K.; Yefimova, S.; Tkacheva, T.; Maksimchuk, P.; Borovoy, I.; Klochkov, V.; Kavok, N.; Opolonin, O.; Malyukin, Y. Reactive oxygen species generation in aqueous solutions containing GdVO4:Eu3+ nanoparticles and their complexes with methylene blue. Nanoscale Res. Lett. 2018, 13, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lion, Y.; Delmelle, M.; Van De Vorst, A. New method of detecting singlet oxygen production. Nat. Cell Biol. 1976, 263, 442–443. [Google Scholar] [CrossRef] [PubMed]
- Pieper, G.M.; Felix, C.C.; Kalyanaraman, B.; Turk, M.; Roza, A.M. Detection by ESR of DMPO hydroxyl adduct formation from islets of langerhans. Free. Radic. Biol. Med. 1995, 19, 219–225. [Google Scholar] [CrossRef]
- Lloyd, R.V.; Mason, R.P. Evidence against transition metal-independent hydroxyl radical generation by xanthine oxidase. J. Biol. Chem. 1990, 265, 16733–16736. [Google Scholar] [CrossRef]
- Lu, H.; Zhen, J.; Wu, T.; Peng, A.; Ye, T.; Wang, T.; Yu, X.; Vaziri, N.D.; Mohan, C.; Zhou, X.J. Superoxide dismutase mimetic drug tempol aggravates anti-GBM antibody-induced glomerulonephritis in mice. Am. J. Physiol. Physiol. 2010, 299, F445–F452. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, C.S. Effects of tempol and redox-cycling nitroxides in models of oxidative stress. Pharmacol. Ther. 2010, 126, 119–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, K.; Takeshita, K.; Ueda, J.-I.; Ozawa, T. Two Reaction Sites of a Spin Label, TEMPOL (4-Hydroxy-2,2,6,6-tetramethylpiperidine-N-oxyl), with Hydroxyl Radical. J. Pharm. Sci. 2003, 92, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Kudo, W.; Yamato, M.; Yamada, K.-I.; Kinoshita, Y.; Shiba, T.; Watanabe, T.; Utsumi, H. Formation of TEMPOL-hydroxylamine during reaction between TEMPOL and hydroxyl radical: HPLC/ECD study. Free. Radic. Res. 2008, 42, 505–512. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krasowska, J.; Pierzchała, K.; Bzowska, A.; Forró, L.; Sienkiewicz, A.; Wielgus-Kutrowska, B. Chromophore of an Enhanced Green Fluorescent Protein Can Play a Photoprotective Role Due to Photobleaching. Int. J. Mol. Sci. 2021, 22, 8565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168565
Krasowska J, Pierzchała K, Bzowska A, Forró L, Sienkiewicz A, Wielgus-Kutrowska B. Chromophore of an Enhanced Green Fluorescent Protein Can Play a Photoprotective Role Due to Photobleaching. International Journal of Molecular Sciences. 2021; 22(16):8565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168565
Chicago/Turabian StyleKrasowska, Joanna, Katarzyna Pierzchała, Agnieszka Bzowska, László Forró, Andrzej Sienkiewicz, and Beata Wielgus-Kutrowska. 2021. "Chromophore of an Enhanced Green Fluorescent Protein Can Play a Photoprotective Role Due to Photobleaching" International Journal of Molecular Sciences 22, no. 16: 8565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168565