
Cytogenetically Elusive Sex Chromosomes in Scincoidean Lizards

 , , and
, , and
Abstract
:1. Introduction
2. Results
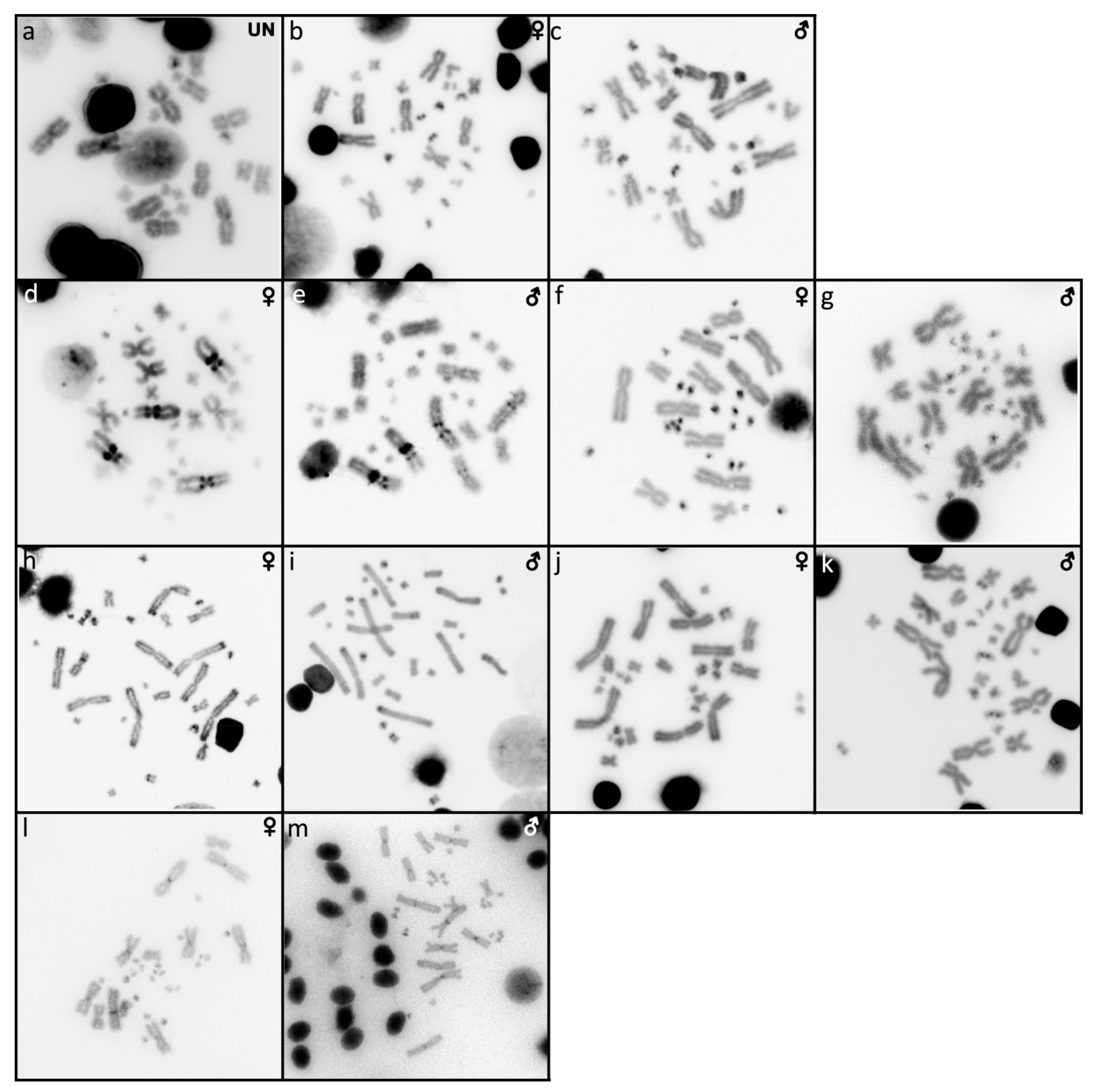
2.1. Acontias percivali
2.2. Cyclodomorphus gerrardii
2.3. Eutropis multifasciata
2.4. Platysaurus sp. 1
2.5. Platysaurus sp. 2
2.6. Scincopus fasciatus
2.7. Tiliqua gigas
2.8. Tiliqua scincoides
2.9. Tracheloptychus madagascariensis
3. Discussion
4. Materials and Methods
4.1. Samples and Species Verification
4.2. Chromosome Preparation and Staining
4.3. Fluorescence In Situ Hybridization with Probes for rDNA Loci (18S/28S) and Telomeric Motifs
4.4. Comparative Genomic Hybridization
4.5. Microscopy and Image Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uetz, P.; Freed, P.; Aguilar, R.; Hošek, J. The Reptile Database. Available online: http://www.reptile-database.org (accessed on 6 June 2021).
- Odierna, G.; Canapa, A.; Andreone, F.; Aprea, G.; Barucca, M.; Capriglione, T.; Olmo, E. A phylogenetic analysis of Cordyliformes (Reptilia: Squamata): Comparison of molecular and karyological data. Mol. Phylogenetics Evol. 2002, 23, 37–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olmo, E.; Signorino, G. Chromorep: A Reptile Chromosomes Database. Available online: http://chromorep.univpm.it (accessed on 6 June 2021).
- Giovannotti, M.; Caputo, V.; O’Brien, P.; Lovell, F.; Trifonov, V.; Cerioni, P.N.; Olmo, E.; Ferguson-Smith, M.; Rens, W. Skinks (Reptilia: Scincidae) have highly conserved karyotypes as revealed by chromosome painting. Cytogenet. Genome Res. 2009, 127, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Deweese, J.E.; Wright, J.W. A preliminary karyological analysis of scincid lizards. Mamm. Chromosome Newsl. 1970, 11, 95–96. [Google Scholar]
- Branch, W.R. Chromosome morphology of some reptiles from Oman and adjacent territories. J. Oman Stud. Rep. 1980, 2, 333–345. [Google Scholar]
- Caputo, V.; Odierna, G.; Aprea, G. A chromosomal study of Eumeces and Scincus, primitive members of the Scincidae (Reptilia, Squamata). Bolletino Zool. 1994, 61, 155–162. [Google Scholar] [CrossRef]
- King, M. Karyotypic studies of some Australian Scincidae (Reptilia). Aust. J. Zool. 1973, 21, 21–32. [Google Scholar] [CrossRef]
- Ota, H.; Hikida, T.; Matsui, M.; Hasegawa, M.; Labang, D.; Nabhitabhata, J. Chromosomal variation in the scincid genus Mabuya and its arboreal relatives (Reptilia: Squamata). Genetica 1996, 98, 87–94. [Google Scholar] [CrossRef]
- Gordon, D.H.; Haacke, W.D.; Jacobsen, N.H.G. Chromosomal studies of relationships in Gekkonidae, Chamaeleonidae and Scincidae in South Africa. J. Herpetol. Assoc. Afr. 1989, 36, 77. [Google Scholar] [CrossRef]
- Hass, C.A.; Hedges, S.B. Karyotype of the Cuban lizard Cricosaura typica and its implications for xantusiid phylogeny. Copeia 1992, 1992, 563. [Google Scholar] [CrossRef]
- Bezy, R.L. Karyotypic variation and evolution of the lizards in the family Xantusiidae. Los Angeles County Mus. Contr. Sci. 1972, 227, 1–29. [Google Scholar]
- Hardy, G.S. The karyotypes of two scincid lizards, and their bearing on relationships in genus Leiolopisma and its relatives (Scincidae: Lygosominae). N. Z. J. Zool. 1979, 6, 609–612. [Google Scholar] [CrossRef]
- Matsubara, K.; O’Meally, D.; Azad, B.; Georges, A.; Sarre, S.D.; Graves, J.; Matsuda, Y.; Ezaz, T. Amplification of microsatellite repeat motifs is associated with the evolutionary differentiation and heterochromatinization of sex chromosomes in Sauropsida. Chromosoma 2016, 125, 111–123. [Google Scholar] [CrossRef]
- Hill, P.; Shams, F.; Burridge, C.; Wapstra, E.; Ezaz, T. Differences in homomorphic sex chromosomes are associated with population divergence in sex determination in Carinascincus ocellatus (Scincidae: Lygosominae). Cells 2021, 10, 291. [Google Scholar] [CrossRef] [PubMed]
- Donnellan, S. Chromosomes of Australian lygosomine skinks (Lacertilia: Scincidae). Genetica 1991, 83, 207–222. [Google Scholar] [CrossRef]
- Hutchinson, M.N.; Donnellan, S. Taxonomy and genetic variation in the Australian lizards of the genus Pseudemoia (Scincidae: Lygosominae). J. Nat. Hist. 1992, 26, 215–264. [Google Scholar] [CrossRef]
- Shine, R.; Elphick, M.J.; Donnellan, S. Co-occurrence of multiple, supposedly incompatible modes of sex determination in a lizard population. Ecol. Lett. 2002, 5, 486–489. [Google Scholar] [CrossRef]
- Quinn, A.E.; Radder, R.S.; Sarre, S.D.; Georges, A.; Ezaz, T.; Shine, R. Isolation and development of a molecular sex marker for Bassiana duperreyi, a lizard with XX/XY sex chromosomes and temperature-induced sex reversal. Mol. Genet. Genom. 2009, 281, 665–672. [Google Scholar] [CrossRef]
- Wright, J.W. Evolution of the X1X2Y sex chromosome mechanism in the scincid lizard Scincella laterale (Say). Chromosoma 1973, 43, 101–108. [Google Scholar]
- Castiglia, R.; Bezerra, A.; Flores-Villela, O.; Annesi, F.; Muñoz, A.; Gornung, E. Comparative cytogenetics of two species of ground skinks: Scincella assata and S. cherriei (Squamata: Scincidae: Lygosominae) from Chiapas, Mexico. Acta Herpetol. 2013, 8, 69–73. [Google Scholar] [CrossRef]
- Hedin, M.C.; Sudman, P.D.; Greenbaum, I.F.; Sites, J.W. Synaptonemal complex analysis of sex chromosome pairing in the common ground skink, Scincella lateralis (Sauria, Scincidae). Copeia 1990, 1990, 1114. [Google Scholar] [CrossRef] [Green Version]
- Patawang, I.; Chuaynkern, Y.; Supanuam, P.; Maneechot, N.; Pinthong, K.; Tanomtong, A. Cytogenetics of the skinks (Reptilia, Scincidae) from Thailand; IV: Newly investigated karyotypic features of Lygosoma quadrupes and Scincella melanosticta. Caryologia 2018, 71, 29–34. [Google Scholar] [CrossRef]
- Kostmann, A.; Kratochvíl, L.; Rovatsos, M. Poorly differentiated XX/XY sex chromosomes are widely shared across skink radiation. Proc. R. Soc. B Biol. Sci. 2021, 288, 20202139. [Google Scholar] [CrossRef] [PubMed]
- Cornejo-Páramo, P.; Dissanayake, D.; Lira-Noriega, A.; Martínez-Pacheco, M.L.; Acosta, A.; Ramírez-Suástegui, C.; Méndez-De-La-Cruz, F.R.; Székely, T.; Urrutia, A.O.; Georges, A.; et al. Viviparous reptile regarded to have temperature-dependent sex determination has old XY chromosomes. Genome Biol. Evol. 2020, 12, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.V.; Pinto, B.J.; Guzmán-Méndez, I.A.; Gamble, T. First report of sex chromosomes in night lizards (Scincoidea: Xantusiidae). J. Hered. 2020, 111, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Kratochvíl, L.; Vukić, J.; Červenka, J.; Kubička, L.; Pokorná, M.J.; Kukačková, D.; Rovatsos, M.; Piálek, L. Mixed-sex offspring produced via cryptic parthenogenesis in a lizard. Mol. Ecol. 2020, 29, 4118–4127. [Google Scholar] [CrossRef]
- Kostmann, A.; Kratochvíl, L.; Rovatsos, M. First report of sex chromosomes in plated lizards (Squamata: Gerrhosauridae). Sex. Dev. 2020, 14, 1–6. [Google Scholar] [CrossRef]
- De Smet, W. Description of the orcein stained karyotypes of 36 lizard species (Lacertilia, Reptilia) belonging to the families Teiidae, Scincidae, Lacertidae, Cordylidae and Varanidae (Autarchoglossa). Acta. Zool. Pathol. Antverp. 1981, 76, 73–118. [Google Scholar]
- Clemente, L.; Mazzoleni, S.; Pensabene Bellavia, E.; Augstenová, B.; Auer, M.; Praschag, P.; Protiva, T.; Velenský, P.; Wagner, P.; Fritz, U.; et al. Interstitial telomeric repeats are rare in turtles. Genes 2020, 11, 657. [Google Scholar] [CrossRef]
- Mazzoleni, S.; Augstenová, B.; Clemente, L.; Auer, M.; Fritz, U.; Praschag, P.; Protiva, T.; Velenský, P.; Kratochvíl, L.; Rovatsos, M. Sex is determined by XX/XY sex chromosomes in Australasian side-necked turtles (Testudines: Chelidae). Sci. Rep. 2020, 10, 4276. [Google Scholar] [CrossRef] [Green Version]
- Badenhorst, D.; Stanyon, R.; Engstrom, T.; Valenzuela, N. A ZZ/ZW microchromosome system in the spiny softshell turtle, Apalone spinifera, reveals an intriguing sex chromosome conservation in Trionychidae. Chromosome Res. 2013, 21, 137–147. [Google Scholar] [CrossRef]
- Rovatsos, M.; Praschag, P.; Fritz, U.; Kratochvšl, L. Stable Cretaceous sex chromosomes enable molecular sexing in softshell turtles (Testudines: Trionychidae). Sci. Rep. 2017, 7, srep42150. [Google Scholar] [CrossRef]
- Porter, C.A.; Hamilton, M.J.; Sites, J.W., Jr.; Baker, R.J. Location of ribosomal DNA in chromosomes of squamate reptiles: Systematic and evolutionary implications. Herpetologica 1991, 47, 271–280. [Google Scholar]
- Stults, D.M.; Killen, M.W.; Pierce, H.H.; Pierce, A.J. Genomic architecture and inheritance of human ribosomal RNA gene clusters. Genome Res. 2007, 18, 13–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezaz, T.; Quinn, A.E.; Miura, I.; Sarre, S.D.; Georges, A.; Graves, J. The dragon lizard Pogona vitticeps has ZZ/ZW micro-sex chromosomes. Chromosome Res. 2005, 13, 763–776. [Google Scholar] [CrossRef]
- Matsubara, K.; Sarre, S.D.; Georges, A.; Matsuda, Y.; Graves, J.A.M.; Ezaz, T. Highly differentiated ZW sex microchromosomes in the Australian Varanus species evolved through rapid amplification of repetitive sequences. PLoS ONE 2014, 9, e95226. [Google Scholar] [CrossRef]
- Altmanová, M.; Rovatsos, M.; Kratochvíl, L.; Johnson Pokorná, M. Minute Y chromosomes and karyotype evolution in Madagascan iguanas (Squamata: Iguania: Opluridae). Biol. J. Linn. Soc. 2016, 118, 618–633. [Google Scholar] [CrossRef]
- Rovatsos, M.; Johnson Pokorná, M.; Altmanová, M.; Kratochvíl, L. Mixed-up sex chromosomes: Identification of sex chromosomes in the X1X1X2X2/X1X2Y system of the legless lizards of the genus Lialis (Squamata: Gekkota: Pygopodidae). Cytogenet. Genome Res. 2016, 149, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Altmanová, M.; Johnson Pokorná, M.; Augstenová, B.; Kratochvíl, L. Cytogenetics of the Javan file snake (Acrochordus javanicus) and the evolution of snake sex chromosomes. J. Zool. Syst. Evol. Res. 2017, 56, 117–125. [Google Scholar] [CrossRef]
- Rovatsos, M.; Altmanová, M.; Augstenová, B.; Mazzoleni, S.; Velenský, P.; Kratochvíl, L. ZZ/ZW sex determination with multiple neo-sex chromosomes is common in Madagascan chameleons of the genus Furcifer (Reptilia: Chamaeleonidae). Genes 2019, 10, 1020. [Google Scholar] [CrossRef] [Green Version]
- Rice, W.R. Evolution of the Y sex chromosome in animals. BioScience 1996, 46, 331–343. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, B.; Charlesworth, D. The degeneration of Y chromosomes. Philos. Trans. R. Soc. B: Biol. Sci. 2000, 355, 1563–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrin, N. Sex reversal: A fountain of youth for sex chromosomes? Evolution 2009, 63, 3043–3049. [Google Scholar] [CrossRef]
- Jeffries, D.L.; Lavanchy, G.; Sermier, R.; Sredl, M.J.; Miura, I.; Borzée, A.; Barrow, L.N.; Canestrelli, D.; Crochet, P.-A.; Dufresnes, C.; et al. A rapid rate of sex-chromosome turnover and non-random transitions in true frogs. Nat. Commun. 2018, 9, 4088. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Kratochvíl, L.; Altmanová, M.; Johnson Pokorná, M. Interstitial telomeric motifs in squamate reptiles: When the exceptions outnumber the rule. PLoS ONE 2015, 10, e0134985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolzán, A.D.; Bianchi, M.S. Telomeres, interstitial telomeric repeat sequences, and chromosomal aberrations. Mutat. Res. Mutat. Res. 2006, 612, 189–214. [Google Scholar] [CrossRef]
- Ruiz-Herrera, A.; Nergadze, S.G.; Santagostino, M.; Giulotto, E. Telomeric repeats far from the ends: Mechanisms of origin and role in evolution. Cytogenet. Genome Res. 2008, 122, 219–228. [Google Scholar] [CrossRef]
- Aksenova, A.Y.; Greenwell, P.W.; Dominska, M.; Shishkin, A.A.; Kim, J.C.; Petes, T.D.; Mirkin, S.M. Genome rearrangements caused by interstitial telomeric sequences in yeast. Proc. Natl. Acad. Sci. USA 2013, 110, 19866–19871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolzán, A.D. Interstitial telomeric sequences in vertebrate chromosomes: Origin, function, instability and evolution. Mutat. Res. Mutat. Res. 2017, 773, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Shubernetskaya, O.; Skvortsov, D.; Dontsova, O.; Kireev, I.; Evfratov, S.; Rubtsova, M.; Belova, E.; Strelkova, O.; Cherepaninets, V.; Zhironkina, O.; et al. Interstitial telomeric repeats-associated DNA breaks. Nucleus 2017, 8, 641–653. [Google Scholar] [CrossRef] [Green Version]
- Rovatsos, M.T.; Marchal, J.A.; Romero-Fernández, I.; Fernandez, F.J.; Giagia-Athanosopoulou, E.B.; Sánchez, A. Rapid, independent, and extensive amplification of telomeric repeats in pericentromeric regions in karyotypes of arvicoline rodents. Chromosome Res. 2011, 19, 869–882. [Google Scholar] [CrossRef]
- Augstenová, B.; Mazzoleni, S.; Kratochvíl, L.; Rovatsos, M. Evolutionary dynamics of the W chromosome in caenophidian snakes. Genes 2017, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Rovatsos, M.; Marchal, J.; Giagia-Athanasopoulou, E.; Sánchez, A. Molecular composition of heterochromatin and its contribution to chromosome variation in the Microtus thomasi/Microtus atticus species complex. Genes 2021, 12, 807. [Google Scholar] [CrossRef] [PubMed]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Endow, S. Polytenization of the ribosomal genes on the X and Y chromosomes of Drosophila melanogaster. Genetics 1982, 100, 375–385. [Google Scholar] [CrossRef]
- Ijdo, J.W.; Baldini, A.; Ward, D.C.; Reeders, S.T.; Wells, R.A. Origin of human chromosome 2: An ancestral telomere-telomere fusion. Proc. Natl. Acad. Sci. USA 1991, 88, 9051–9055. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | Subfamily | Sex | ||
|---|---|---|---|---|---|
| Male | Female | Unknown | |||
| Platysaurus sp. 1 | Cordylidae | Platysaurinae | - | 1 | - |
| Platysaurus sp. 2 | Cordylidae | Platysaurinae | 1 | - | - |
| Tracheloptychus madagascariensis | Gerrhosauridae | Zonosaurinae | 1 | 1 | - |
| Acontias percivali | Scincidae | Acontinae | - | - | 1 |
| Cyclodomorphus gerrardii | Scincidae | Lygosominae | 2 | 2 | - |
| Eutropis multifasciata | Scincidae | Lygosominae | 2 | 1 | - |
| Tiliqua gigas | Scincidae | Lygosominae | 5 | 3 | - |
| Tiliqua scincoides | Scincidae | Lygosominae | 1 | 3 | - |
| Scincopus fasciatus | Scincidae | Scincinae | 1 | 1 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostmann, A.; Augstenová, B.; Frynta, D.; Kratochvíl, L.; Rovatsos, M. Cytogenetically Elusive Sex Chromosomes in Scincoidean Lizards. Int. J. Mol. Sci. 2021, 22, 8670. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168670
Kostmann A, Augstenová B, Frynta D, Kratochvíl L, Rovatsos M. Cytogenetically Elusive Sex Chromosomes in Scincoidean Lizards. International Journal of Molecular Sciences. 2021; 22(16):8670. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168670
Chicago/Turabian StyleKostmann, Alexander, Barbora Augstenová, Daniel Frynta, Lukáš Kratochvíl, and Michail Rovatsos. 2021. "Cytogenetically Elusive Sex Chromosomes in Scincoidean Lizards" International Journal of Molecular Sciences 22, no. 16: 8670. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168670