
Genome-Wide Identification and Characterization of the Brassinazole-resistant (BZR) Gene Family and Its Expression in the Various Developmental Stage and Stress Conditions in Wheat (Triticum aestivum L.)

 ,
,  , ,
, ,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Identification of BZR Family Members in T. aestivum
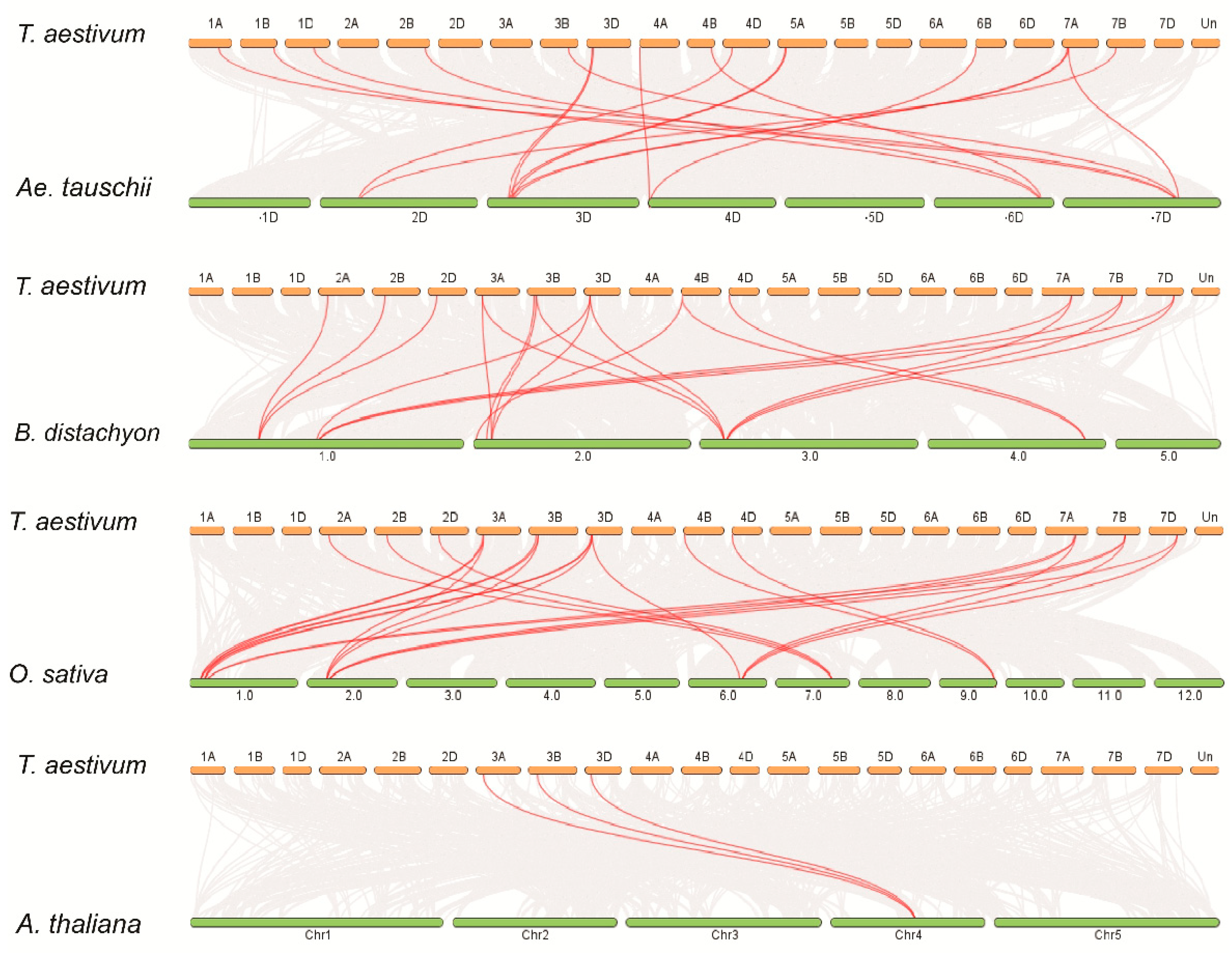
2.2. Chromosomal Distribution of TaBZR Genes
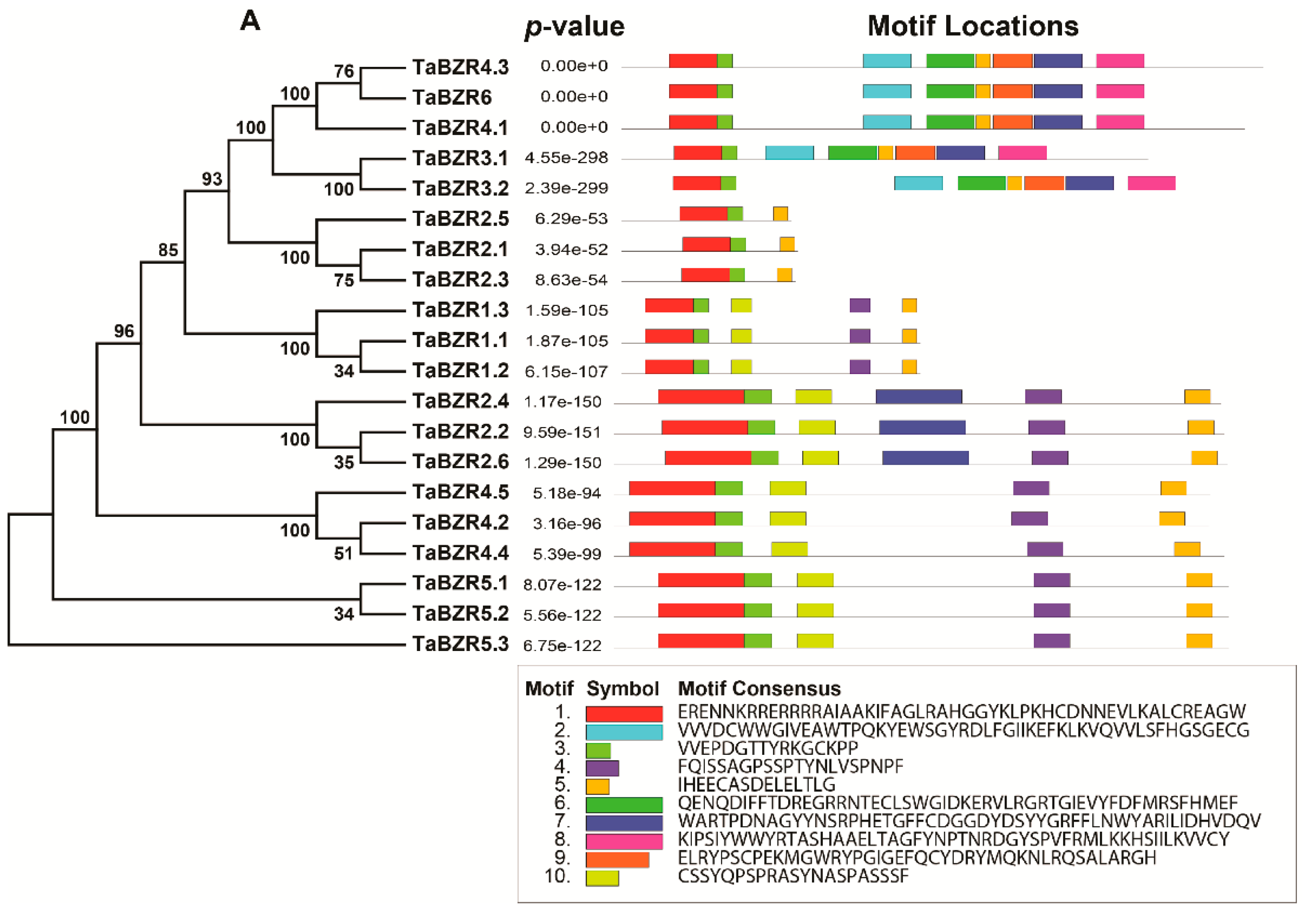
2.3. Gene Structure and Conserved Motif Analysis of TaBZR Genes
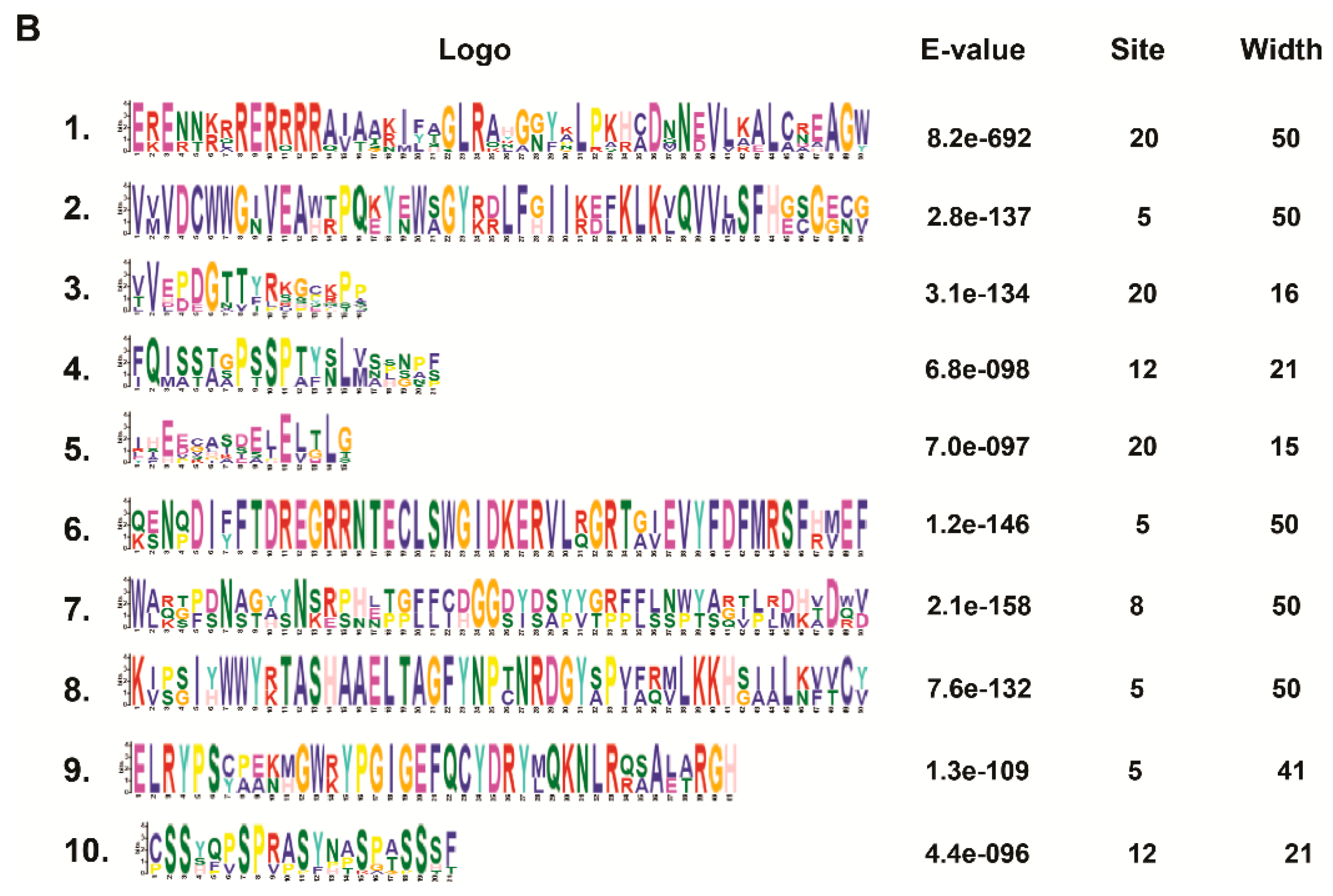
2.4. Conserved Motif Analysis
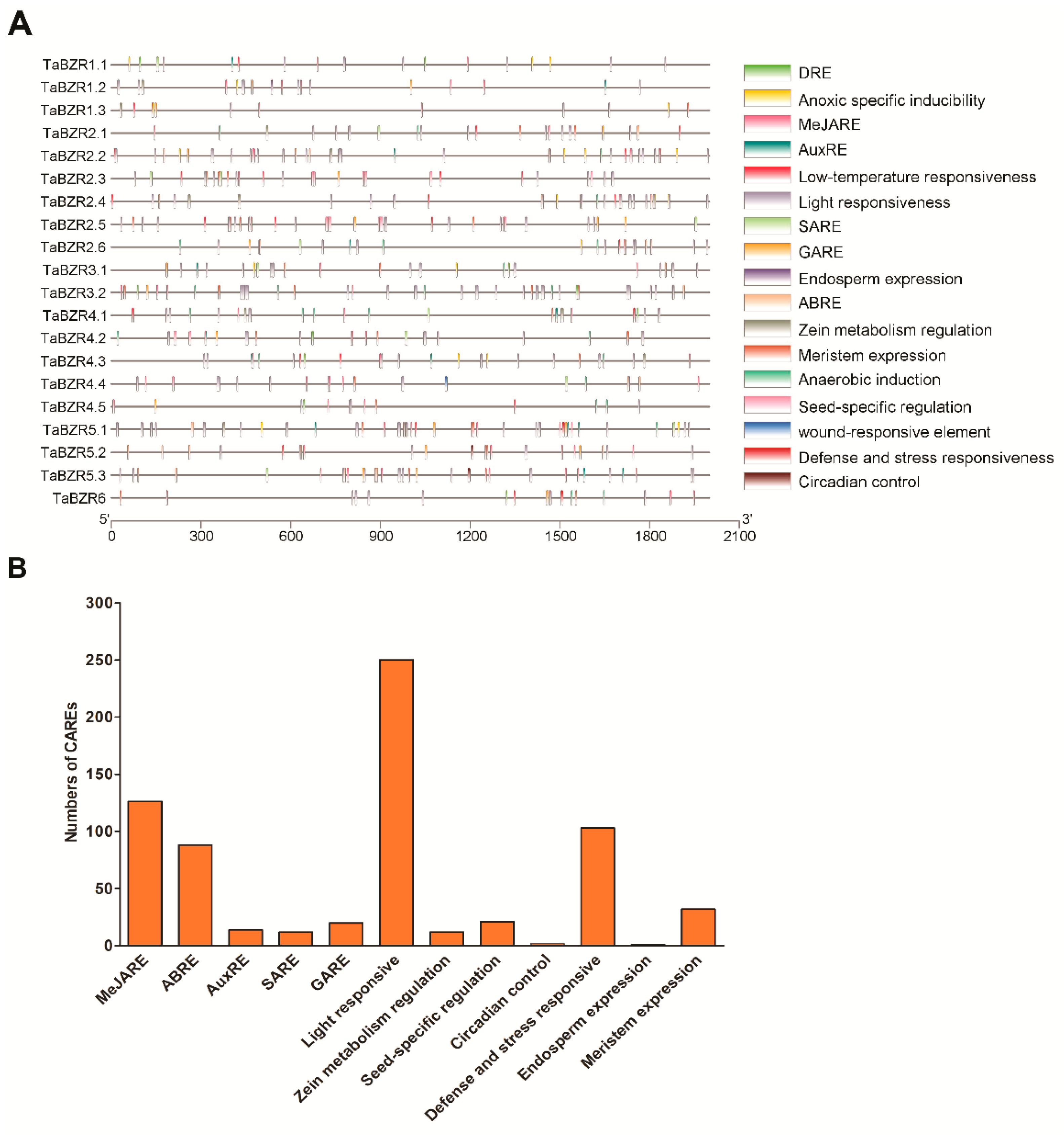
2.5. Cis-Acting Regulatory Elements (CAREs) Analysis of TaBZR Genes
2.6. Gene Ontology (GO) Enrichment of TaBZR Genes
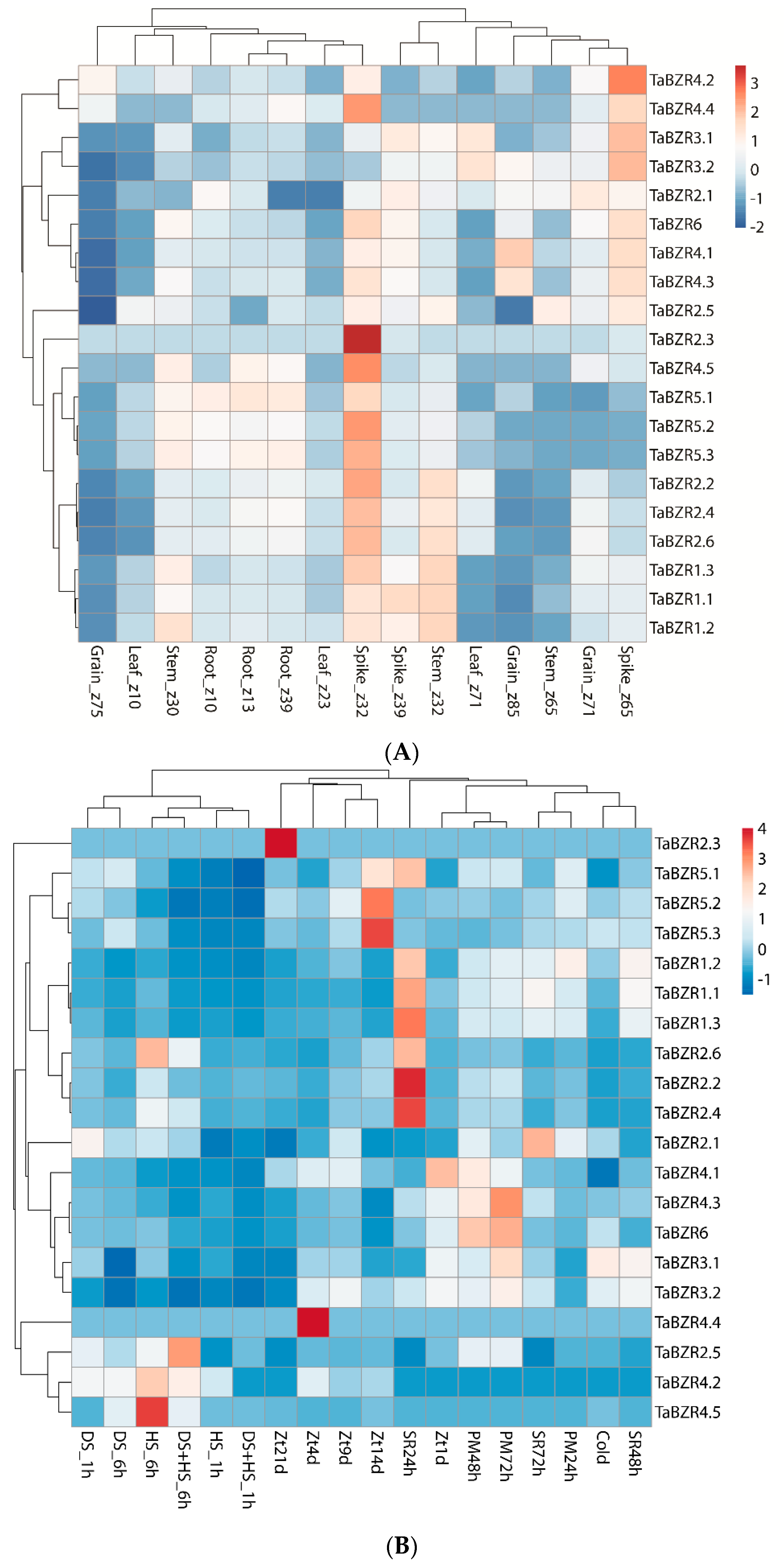
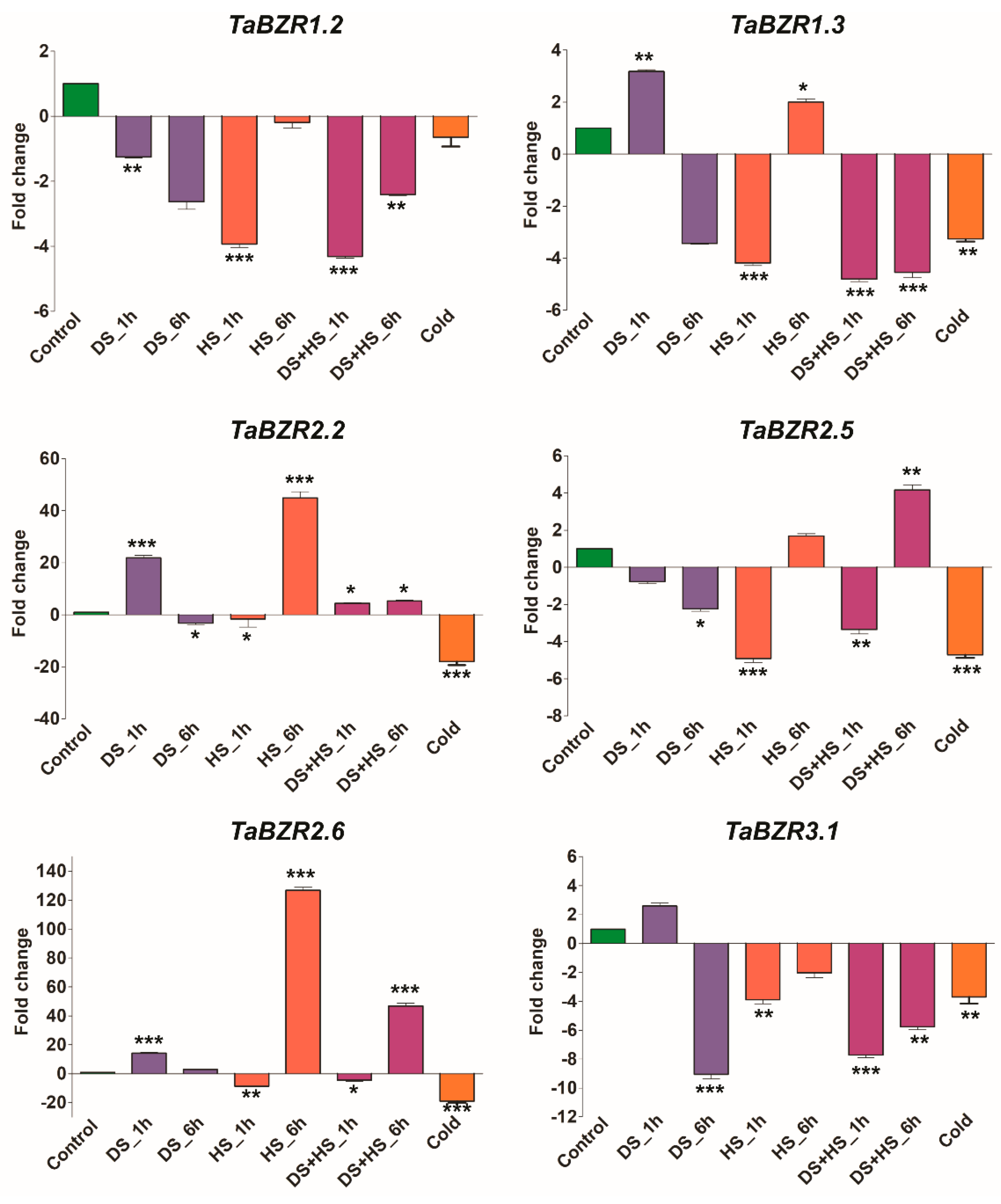
2.7. Expression Profiling of TaBZR Genes
2.8. Protein-Protein Networks Analysis of the TaBZR Family Genes
3. Discussion
4. Materials and Methods
4.1. Identification of BZR Genes in the Wheat Genome
4.2. Chromosome Localization and Gene Duplication
4.3. Physico-Chemical Characteristics, Subcellular Localization, and 3D Structure
4.4. Gene Structure, Gene Ontology, and Motif Analysis
4.5. Cis-Acting Regulatory Elements (CAREs) Analysis and Protein Interaction Network
4.6. Expression Profiling of TaBZR Genes
4.7. Plant Material, Growth Conditions, Drought, and Heat Treatment
4.8. RNA Isolation and Real-Time PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benjamins, R.; Scheres, B. Auxin: The Looping Star in Plant Development. Annu. Rev. Plant Biol. 2008, 59, 443–465. [Google Scholar] [CrossRef] [PubMed]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P. The Role of Gibberellin Signalling in Plant Responses to Abiotic Stress. J. Exp. Biol. 2014, 217, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Šimura, J.; Antoniadi, I.; Široká, J.; Tarkowská, D.; Strnad, M.; Ljung, K.; Novák, O. Plant Hormonomics: Multiple Phytohormone Profiling by Targeted Metabolomics. Plant Physiol. 2018, 177, 476–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Planas-Riverola, A.; Gupta, A.; Betegón-Putze, I.; Bosch, N.; Ibañes, M.; Caño-Delgado, A.I. Brassinosteroid Signaling in Plant Development and Adaptation to Stress. Development 2019, 146, dev151894. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Yang, J.; Li, X.; Zhang, Y. Salicylic Acid: Biosynthesis and Signaling. Annu. Rev. Plant Biol. 2021, 72, 761–791. [Google Scholar] [CrossRef]
- Cheng, M.-C.; Kathare, P.K.; Paik, I.; Huq, E. Phytochrome Signaling Networks. Annu. Rev. Plant Biol. 2021, 72, 217–244. [Google Scholar] [CrossRef]
- Pauwels, L.; Barbero, G.F.; Geerinck, J.; Tilleman, S.; Grunewald, W.; Pérez, A.C.; Chico, J.M.; Bossche, R.V.; Sewell, J.; Gil, E. NINJA Connects the Co-repressor TOPLESS to Jasmonate Signalling. Nature 2010, 464, 788–791. [Google Scholar] [CrossRef] [Green Version]
- Al-Babili, S.; Bouwmeester, H.J. Strigolactones, a Novel Carotenoid-derived Plant Hormone. Annu. Rev. Plant Biol. 2015, 66, 161–186. [Google Scholar] [CrossRef]
- Ku, Y.S.; Sintaha, M.; Cheung, M.Y.; Lam, H.M. Plant Hormone Signaling Crosstalks between Biotic and Abiotic Stress Responses. Int. J. Mol. Sci. 2018, 19, 3206. [Google Scholar] [CrossRef] [Green Version]
- Shimotohno, A.; Aki, S.S.; Takahashi, N.; Umeda, M. Regulation of the Plant Cell Cycle in Response to Hormones and the Environment. Annu. Rev. Plant Biol. 2021, 72, 273–296. [Google Scholar] [CrossRef] [PubMed]
- Clouse, S.D.; Langford, M.; McMorris, T.C. A Brassinosteroid-insensitive Mutant in Arabidopsis thaliana Exhibits Multiple Defects in Growth and Development. Plant Physiol. 1996, 111, 671–678. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Ye, H.; Guo, H.; Yin, Y. Arabidopsis IWS1 Interacts with Transcription Factor BES1 and is Involved in Plant Steroid Hormone Brassinosteroid Regulated Gene Expression. Proc. Natl. Acad. Sci. USA 2010, 107, 3918–3923. [Google Scholar] [CrossRef] [Green Version]
- Clouse, S.D. Brassinosteroid Signal Transduction: From Receptor Kinase Activation to Transcriptional Networks Regulating Plant Development. Plant Cell 2011, 23, 1219–1230. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.Y.; Sae-Seaw, J.; Wang, Z.Y. Brassinosteroid Signalling. Development 2013, 140, 1615–1620. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Li, J. Brassinosteroids Regulate Root Growth, Development, and Symbiosis. Mol. Plant 2016, 9, 86–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.J.; Russinova, E. Brassinosteroid Signalling. Curr. Biol. 2020, 30, R294–R298. [Google Scholar] [CrossRef] [PubMed]
- Oklestkova, J.; Rárová, L.; Kvasnica, M.; Strnad, M. Brassinosteroids: Synthesis and Biological Activities. Phytochem. Rev. 2015, 14, 1053–1072. [Google Scholar] [CrossRef]
- Vardhini, B.V.; Anjum, N.A. Brassinosteroids make Plant Life Easier under Abiotic Stresses mainly by Modulating Major Components of Antioxidant Defense System. Front. Environ. Sci. 2015, 2, 67. [Google Scholar] [CrossRef] [Green Version]
- Saini, S.; Sharma, I.; Pati, P.K. Versatile Roles of Brassinosteroid in Plants in the Context of its Homoeostasis, Signaling and Crosstalks. Front. Plant Sci. 2015, 6, 950. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.P.; Savaldi-Goldstein, S. Growth Control: Brassinosteroid Activity Gets Context. J. Exp. Biol. 2015, 66, 1123–1132. [Google Scholar] [CrossRef] [Green Version]
- Derevyanchuk, M.; Litvinovskaya, R.; Khripach, V.; Kravets, V. Brassinosteroid-induced De Novo Protein Synthesis in Zea mays under Salinity and Bioinformatic Approach for Identification of Heat Shock Proteins. Plant Growth Regul. 2016, 78, 297–305. [Google Scholar] [CrossRef]
- An, W.; Chang, D.; Zhang, F. Expression Characteristics of Transcription Factor BES1/BRI1 of Cotton Seedling in Response to Brassinosteroid under Drought Stress. Acta Bot. Boreali. Occident. Sin. 2015, 35, 1311–1316. [Google Scholar]
- Yin, Y.; Wang, Z.Y.; Mora-Garcia, S.; Li, J.; Yoshida, S.; Asami, T.; Chory, J. BES1 Accumulates in the Nucleus in Response to Brassinosteroids to Regulate Gene Expression and Promote Stem Elongation. Cell 2002, 109, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Vafeados, D.; Tao, Y.; Yoshida, S.; Asami, T.; Chory, J. A New Class of Transcription Factors Mediates Brassinosteroid-regulated Gene Expression in Arabidopsis. Cell 2005, 120, 249–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.Y.; Nakano, T.; Gendron, J.; He, J.; Chen, M.; Vafeados, D.; Yang, Y.; Fujioka, S.; Yoshida, S.; Asami, T. Nuclear-localized BZR1 Mediates Brassinosteroid-induced Growth and Feedback Suppression of Brassinosteroid Biosynthesis. Dev. Cell 2002, 2, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Peng, P.; Schmitz, R.J.; Decker, A.D.; Tax, F.E.; Li, J. Two Putative BIN2 Substrates are Nuclear Components of Brassinosteroid Signaling. Plant Physiol. 2002, 130, 1221–1229. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Liu, H.; Guo, S.; Wang, B.; Li, Z.; Chong, K.; Xu, Y. OsmiR396d Affects Gibberellin and Brassinosteroid Signaling to Regulate Plant Architecture in Rice. Plant Physiol. 2018, 176, 946–959. [Google Scholar] [CrossRef] [Green Version]
- Oh, E.; Zhu, J.Y.; Wang, Z.Y. Interaction between BZR1 and PIF4 integrates brassinosteroid and environmental responses. Nat. Cell Biol. 2012, 14, 802–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, H.; Li, L.; Guo, H.; Yin, Y. MYBL2 is a Substrate of GSK3-like Kinase BIN2 and Acts as a Corepressor of BES1 in Brassinosteroid Signaling Pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 20142–20147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, S.; Sun, S.; Wang, L.; Wu, Z.; Li, C.; Li, X.; Wang, T.; Leng, L.; Tian, W.; Lu, T. The RLA1/SMOS1 Transcription Factor Functions with OsBZR1 to Regulate Brassinosteroid Signaling and Rice Architecture. Plant Cell 2017, 29, 292–309. [Google Scholar] [CrossRef] [Green Version]
- Manoli, A.; Trevisan, S.; Quaggiotti, S.; Varotto, S. Identification and Characterization of the BZR Transcription Factor Family and its Expression in Response to Abiotic Stresses in Zea mays L. Plant Growth Regul. 2018, 84, 423–436. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Ma, X.; Li, C.; Hu, J.; Yang, Q.; Wang, T.; Wang, L.; Wang, J.; Guo, D.; Ge, W. Comprehensive Analyses of the BES1 Gene Family in Brassica napus and Examination of their Evolutionary Pattern in Representative Species. BMC Genom. 2018, 19, 346. [Google Scholar] [CrossRef] [PubMed]
- Su, D.; Xiang, W.; Wen, L.; Lu, W.; Shi, Y.; Liu, Y.; Li, Z. Genome-wide Identification, Characterization and Expression Analysis of BES1 Gene Family in Tomato. BMC Plant Biol. 2021, 21, 161. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Song, X.; Wang, Z.; Duan, W.; Hu, R.; Wang, W.; Li, Y.; Hou, X. Genome-wide Analysis Of The BES1 Transcription Factor Family In Chinese Cabbage (Brassica rapa ssp. pekinensis). Plant Growth Regul. 2016, 80, 291–301. [Google Scholar] [CrossRef]
- Yu, H.; Feng, W.; Sun, F.; Zhang, Y.; Qu, J.; Liu, B.; Lu, F.; Yang, L.; Fu, F.; Li, W. Cloning and Characterization of BES1/BZR1 Transcription Factor Genes in Maize. Plant Growth Regul. 2018, 86, 235–249. [Google Scholar] [CrossRef]
- Wang, W.; Sun, Y.Q.; Li, G.L.; Zhang, S.Y. Genome-wide Identification, Characterization, and Expression Patterns of the BZR Transcription Factor Family in Sugar Beet (Beta vulgaris L.). BMC Plant Biol. 2019, 19, 191. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-G.; Gao, Z.; Zhao, Z.; Liu, X.; Li, Y.; Zhang, Y.; Liu, X.; Sun, Y.; Tang, W. BZR1 Family Transcription Factors Function Redundantly and Indispensably in BR Signaling but Exhibit BRI1-independent Function in Regulating Anther Development in Arabidopsis. Mol. Plant 2019, 12, 1408–1415. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Lv, M.; Wang, Y.; Wang, P.-A.; Cui, Y.; Li, M.; Wang, R.; Gou, X.; Li, J. BES1 is activated by EMS1-TPD1-SERK1/2-mediated Signaling to Control Tapetum Development in Arabidopsis thaliana. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Sahni, S.; Prasad, B.D.; Liu, Q.; Grbic, V.; Sharpe, A.; Singh, S.P.; Krishna, P. Overexpression of The Brassinosteroid Biosynthetic Gene DWF4 in Brassica Napus Simultaneously Increases Seed Yield and Stress Tolerance. Sci. Rep. 2016, 6, 1–14. [Google Scholar]
- Li, A.; Zhou, M.; Wei, D.; Chen, H.; You, C.; Lin, J. Transcriptome Profiling Reveals the Negative Regulation of Multiple Plant Hormone Signaling Pathways Elicited by Overexpression of C-repeat Binding Factors. Front. Plant Sci. 2017, 8, 1647. [Google Scholar] [CrossRef] [Green Version]
- Ye, H.; Liu, S.; Tang, B.; Chen, J.; Xie, Z.; Nolan, T.M.; Jiang, H.; Guo, H.; Lin, H.Y.; Li, L. RD26 Mediates Crosstalk between Drought and Brassinosteroid Signalling Pathways. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhang, Y.J.; Yang, B.J.; Yu, X.X.; Wang, D.; Zu, S.H.; Xue, H.W.; Lin, W.H. Functional Characterization of GmBZL2 (AtBZR1 Like Gene) Reveals the Conserved BR Signaling Regulation in Glycine max. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, G.; Park, J.I.; Jung, H.J.; Ahmed, N.U.; Kayum, M.A.; Kang, J.G.; Nou, I.S. Molecular Characterization of BZR Transcription Factor Family and Abiotic Stress Induced Expression Profiling in Brassica rapa. Plant Physiol. Biochem. 2015, 92, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.Y.; Gao, Y.; Guo, J.; Yu, T.F.; Zheng, W.J.; Liu, Y.W.; Chen, J.; Xu, Z.S.; Ma, Y.Z. BES/BZR Transcription Factor TaBZR2 Positively Regulates Drought Responses by Activation of TaGST1. Plant Physiol. 2019, 180, 605–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, F.; Ding, L.; Feng, W.; Cao, Y.; Lu, F.; Yang, Q.; Li, W.; Lu, Y.; Shabek, N.; Fu, F. Maize Transcription Factor ZmBES1/BZR1-5 Positively Regulates Kernel Size. J. Exp. Biol. 2021, 72, 1714–1726. [Google Scholar]
- Bai, M.Y.; Zhang, L.Y.; Gampala, S.S.; Zhu, S.W.; Song, W.Y.; Chong, K.; Wang, Z.Y. Functions of OsBZR1 and 14-3-3 Proteins in Brassinosteroid Signaling in Rice. Proc. Natl. Acad. Sci. USA 2007, 104, 13839–13844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Q.; Wang, L.; Li, X. GmBEHL1, a BES1/BZR1 Family Protein, Negatively Regulates Soybean Nodulation. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Kesawat, M.S.; Das, B.K.; Bhaganagare, G.R. Genome-wide Identification, Evolutionary and Expression Analyses of Putative Fe–S Biogenesis Genes in Rice (Oryza sativa). Genome 2012, 55, 571–583. [Google Scholar] [CrossRef]
- Kumar, M.; Kherawat, B.S.; Dey, P.; Saha, D.; Singh, A.; Bhatia, S.K.; Ghodake, G.S.; Kadam, A.A.; Kim, H.-U.; Manorama; et al. Genome-Wide Identification and Characterization of PIN-FORMED (PIN) Gene Family Reveals Role in Developmental and Various Stress Conditions in Triticum aestivum L. Int. J. Mol. Sci. 2021, 22, 7396. [Google Scholar] [CrossRef] [PubMed]
- Gill, B.S.; Appels, R.; Botha-Oberholster, A.M.; Buell, C.R.; Bennetzen, J.L.; Chalhoub, B.; Chumley, F.; Dvorák, J.; Iwanaga, M.; Keller, B. A Workshop Report on Wheat Genome Sequencing: International Genome Research on Wheat Consortium. Genetics 2004, 168, 1087–1096. [Google Scholar] [CrossRef] [Green Version]
- Consortium, I.W.G.S. A Chromosome-based Draft Sequence of the Hexaploid Bread Wheat (Triticum aestivum) Genome. Science 2014, 345, 1251788. [Google Scholar]
- Kumar, P.; Yadava, R.; Gollen, B.; Kumar, S.; Verma, R.K.; Yadav, S. Nutritional Contents and Medicinal Properties of Wheat: A Review. Life Sci. Med. Res. 2011, 22, 1–10. [Google Scholar]
- Chen, X.; He, X.; Fu, X.; Huang, Q. In-vitro Digestion and Physicochemical Properties of Wheat Starch/flour Modified by Heat-moisture Treatment. J. Cereal Sci. 2015, 63, 109–115. [Google Scholar] [CrossRef]
- Duan, Y.; Tao, X.; Zhao, H.; Xiao, X.; Li, M.; Wang, J.; Zhou, M. Activity of Demethylation Inhibitor Fungicide Metconazole on Chinese Fusarium Graminearum Species Complex and its Application in Carbendazim-resistance Management of Fusarium Head Blight in Wheat. Plant Dis. 2019, 103, 929–937. [Google Scholar] [CrossRef]
- Appels, R.; Eversole, K.; Stein, N.; Feuillet, C.; Keller, B.; Rogers, J.; Pozniak, C.J.; Choulet, F.; Distelfeld, A.; Poland, J. Shifting the Limits in Wheat Research and Breeding using a Fully Annotated Reference Genome. Science 2018, 361, eaar7191. [Google Scholar]
- Kumar, R.; Masthigowda, M.H.; Kaur, A.; Bhusal, N.; Pandey, A.; Kumar, S.; Mishra, C.; Singh, G.; Singh, G.P. Identification and Characterization of Multiple Abiotic Stress Tolerance Genes in Wheat. Mol. Biol. Rep. 2020, 47, 8629–8643. [Google Scholar] [CrossRef]
- Afzal, F.; Chaudhari, S.K.; Gul, A.; Farooq, A.; Ali, H.; Nisar, S.; Sarfraz, B.; Shehzadi, K.J.; Mujeeb-Kazi, A. Bread Wheat (Triticum aestivum L.) Under Biotic and Abiotic Stresses: An Overview. In Crop Production and Global Environmental Issues; Hakeem, K., Ed.; Springer: Cham, Switzerland, 2015; pp. 293–317. [Google Scholar]
- Yang, Z.; Nielsen, R. Mutation-selection Models of Codon Substitution and their Use to Estimate Selective Strengths on Codon Usage. Mol. Biol. Evol. 2008, 25, 568–579. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Gigras, P.; Mohapatra, H.; Goswami, V.K.; Chauhan, B. Microbial α-amylases: A Biotechnological Perspective. Process. Biochem. 2003, 38, 1599–1616. [Google Scholar] [CrossRef]
- Sindhu, R.; Binod, P.; Pandey, A. α-Amylases. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2017; pp. 3–24. [Google Scholar]
- Rathore, R.; Garg, N.; Garg, S.; Kumar, A. Starch Phosphorylase: Role in Starch Metabolism and Biotechnological Applications. Crit. Rev. Biotechnol. 2009, 29, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Cuesta-Seijo, J.A.; Ruzanski, C.; Krucewicz, K.; Meier, S.; Hägglund, P.; Svensson, B.; Palcic, M.M. Functional and Structural Characterization of Plastidic Starch Phosphorylase during Barley Endosperm Development. PLoS ONE 2017, 12, e0175488. [Google Scholar] [CrossRef] [Green Version]
- Zhiponova, M.K.; Vanhoutte, I.; Boudolf, V.; Betti, C.; Dhondt, S.; Coppens, F.; Mylle, E.; Maes, S.; González-García, M.P.; Caño-Delgado, A.I. Brassinosteroid Production and Signaling Differentially Control Cell Division and Expansion in the Leaf. New Phytol. 2013, 197, 490–502. [Google Scholar] [CrossRef]
- Palovaara, J.; Hallberg, H.; Stasolla, C.; Luit, B.; Hakman, I. Expression of a Gymnosperm PIN Homologous Gene Correlates with Auxin Immunolocalization Pattern at Cotyledon Formation and in Demarcation of the Procambium during Picea Abies Somatic Embryo Development and in Seedling Tissues. Tree Physiol. 2010, 30, 479–489. [Google Scholar] [CrossRef] [Green Version]
- Lespinet, O.; Wolf, Y.I.; Koonin, E.V.; Aravind, L. The Role of Lineage-specific Gene Family Expansion in the Evolution of Eukaryotes. Genome Res. 2002, 12, 1048–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; He, L.; Li, J.; Chen, J.; Liu, C. Genome-wide Identification, Characterization, and Expression Profiling of the Legume BZR Transcription Factor Gene Family. Front. Plant Sci. 2018, 9, 1332. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.C.; Purugganan, M.D. The Early Stages of Duplicate Gene Evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [Green Version]
- Lawton-Rauh, A. Evolutionary Dynamics of Duplicated Genes in Plants. Mol. Phylogenet. Evol. 2003, 29, 396–409. [Google Scholar] [CrossRef]
- Wu, A.; Allu, A.D.; Garapati, P.; Siddiqui, H.; Dortay, H.; Zanor, M.I.; Asensi-Fabado, M.A.; Munné-Bosch, S.; Antonio, C.; Tohge, T. JUNGBRUNNEN1, a Reactive Oxygen Species–responsive NAC Transcription Factor, Regulates Longevity in Arabidopsis. Plant Cell 2012, 24, 482–506. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and Validation of Promoters and Cis-acting Regulatory Elements. Plant Sci. 2014, 217, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.L.; Sen, R.; Roeder, R.G. Enhancer–promoter Communication and Transcriptional Regulation of Igh. Trends Immunol. 2011, 32, 532–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, A.L.; Singer, D.S. Core Promoters in Transcription: Old Problem, New Insights. Trends Biochem. Sci. 2015, 40, 165–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wong, C.H.; Birnbaum, R.Y.; Li, G.; Favaro, R.; Ngan, C.Y.; Lim, J.; Tai, E.; Poh, H.M.; Wong, E. Chromatin Connectivity Maps Reveal Dynamic Promoter–enhancer Long-range Associations. Nature 2013, 504, 306–310. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Tang, Q.; Li, Y.; Zhang, Y.; Zhao, C.; Yan, J.; You, H. Folding/unfolding Kinetics of G-quadruplexes Upstream of the P1 Promoter of the Human BCL-2 Oncogene. J. Biol. Chem. 2019, 294, 5890–5895. [Google Scholar] [CrossRef]
- Banerjee, J.; Sahoo, D.K.; Dey, N.; Houtz, R.L.; Maiti, I.B. An intergenic region shared by At4g35985 and At4g35987 in Arabidopsis thaliana is a Tissue Specific and Stress Inducible Bidirectional Promoter Analyzed in Transgenic Arabidopsis and Tobacco Plants. PLoS ONE 2013, 8, e79622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallego-Bartolomé, J.; Minguet, E.G.; Grau-Enguix, F.; Abbas, M.; Locascio, A.; Thomas, S.G.; Alabadí, D.; Blázquez, M.A. Molecular Mechanism for the Interaction Between Gibberellin and Brassinosteroid Signaling Pathways in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 13446–13451. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.; Guo, G.; Yan, H.; Qiu, Z.; Liu, Q.; Zeng, B. Characterization of Brassinazole Resistant (BZR) Gene Family and Stress Induced Expression in Eucalyptus grandis. Physiol. Mol. Biol. Plants 2018, 24, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Khaliq, A.; Lu, S.; Xie, M.; Ma, Z.; Mao, J.; Chen, B. Genome-wide Identification and Characterization of the BES1 Gene Family in Apple (Malus domestica). Plant Biol. 2020, 22, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.H.; Sun, J.; Oh, D.H.; Zielinski, R.E.; Clouse, S.D.; Huber, S.C. Enhancing Arabidopsis Leaf Growth by Engineering the BRASSINOSTEROID INSENSITIVE1 Receptor Kinase. Plant Physiol. 2011, 157, 120–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fridman, Y.; Elkouby, L.; Holland, N.; Vragović, K.; Elbaum, R.; Savaldi-Goldstein, S. Root Growth is Modulated by Differential Hormonal Sensitivity in Neighboring Cells. Genes Dev. 2014, 28, 912–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyman, J.; Cools, T.; Vandenbussche, F.; Heyndrickx, K.S.; Van Leene, J.; Vercauteren, I.; Vanderauwera, S.; Vandepoele, K.; De Jaeger, G.; Van Der Straeten, D. ERF115 Controls Root Quiescent Center Cell Division and Stem Cell Replenishment. Science 2013, 342, 860–863. [Google Scholar] [CrossRef] [Green Version]
- Vilarrasa-Blasi, J.; González-García, M.P.; Frigola, D.; Fàbregas, N.; Alexiou, K.G.; López-Bigas, N.; Rivas, S.; Jauneau, A.; Lohmann, J.U.; Benfey, P.N. Regulation of Plant Stem Cell Quiescence by a Brassinosteroid Signaling Module. Dev. Cell 2014, 30, 36–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahnejat-Bushehri, S.; Tarkowska, D.; Sakuraba, Y.; Balazadeh, S. Arabidopsis NAC Transcription Factor JUB1 Regulates GA/BR Metabolism and Signalling. Nat. Plants 2016, 2, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Seung, D.; Thalmann, M.; Sparla, F.; Abou Hachem, M.; Lee, S.K.; Issakidis-Bourguet, E.; Svensson, B.; Zeeman, S.C.; Santelia, D. Arabidopsis thaliana AMY3 is a Unique Redox-regulated Chloroplastic α-amylase. J. Biol. Chem. 2013, 288, 33620–33633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhold, H.; Soyk, S.; Šimková, K.; Hostettler, C.; Marafino, J.; Mainiero, S.; Vaughan, C.K.; Monroe, J.D.; Zeeman, S.C. β-Amylase–like Proteins Function as Transcription Factors in Arabidopsis, Controlling Shoot Growth and Development. Plant Cell 2011, 23, 1391–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soyk, S.; Šimková, K.; Zürcher, E.; Luginbühl, L.; Brand, L.H.; Vaughan, C.K.; Wanke, D.; Zeeman, S.C. The Enzyme-like Domain of Arabidopsis Nuclear Β-amylases is Critical for DNA Sequence Recognition and Transcriptional Activation. Plant Cell 2014, 26, 1746–1763. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program. For. Windows 95/98/NT, Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; pp. 95–98. [Google Scholar]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H. MCScanX: A Toolkit for Detection and Evolutionary Analysis of Gene Synteny and Collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Kozlowski, L.P. IPC–isoelectric point calculator. Biol. Direct 2016, 11, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. AgriGO V2. 0: A GO Analysis Toolkit for the Agricultural Community, 2017 Update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef]
- Grigorova, B.; Vaseva, I.; Demirevska, K.; Feller, U. Combined Drought and Heat Stress in Wheat: Changes in Some Heat Shock Proteins. Biol. Plant 2011, 55, 105–111. [Google Scholar] [CrossRef]
- Kim, D.K.; Kesawat, M.S.; Hong, C.B. One Gene Member of the ADP-ribosylation Factor Family is Heat-inducible and Enhances Seed Germination in Nicotiana tabacum. Genes Genom. 2017, 39, 1353–1365. [Google Scholar] [CrossRef]
- Kesawat, M.S.; Kim, D.K.; Zeba, N.; Suh, M.C.; Xia, X.; Hong, C.B. Ectopic RING Zinc Finger Gene from Hot Pepper Induces Totally Different Genes in Lettuce and Tobacco. Mol. Breed. 2018, 38, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Novelli, S.; Gismondi, A.; Di Marco, G.; Canuti, L.; Nanni, V.; Canini, A. Plant Defense Factors Involved in Olea europaea Resistance Against Xylella fastidiosa Infection. J. Plant Res. 2019, 132, 439–455. [Google Scholar] [CrossRef]
- Narancio, R.; John, U.; Mason, J.; Spangenberg, G. Selection of Optimal Reference Genes for Quantitative RT-PCR Transcript Abundance Analysis in White Clover (Trifolium Repens L.). Funct. Plant Biol. 2018, 45, 737–744. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proposed Gene Name | Gene ID | Genomic Location | Orientation | CDS Length (bp) | Intron Number | Protein Length (aa) | Molecular Weight (KDa) | pI | GRAVY | Predicted Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|---|
| TaBZR1.1 | TraesCS2A02G187800 | 2A:150124286-150124643 | Reverse | 942 | 1 | 313 | 33.68 | 7.7 | −0.5881 | Nucleus |
| TaBZR1.2 | TraesCS2B02G219300 | 2B:209436846-209437190 | Forward | 942 | 1 | 313 | 33.61 | 7.7 | −0.583 | Nucleus |
| TaBZR1.3 | TraesCS2D02G199900 | 2D:151325825-151326183 | Forward | 942 | 1 | 313 | 33.62 | 7.7 | −0.5859 | Nucleus |
| TaBZR2.1 | TraesCS3A02G123500 | 3A:99588221-99588358 | Forward | 558 | 1 | 185 | 19.96 | 8.8 | −0.4816 | Nucleus |
| TaBZR2.2 | TraesCS3A02G139000 | 3A:116518978-116520359 | Forward | 1071 | 1 | 356 | 37.41 | 8 | −0.5098 | Nucleus |
| TaBZR2.3 | TraesCS3B02G142600 | 3B:130257621-130257758 | Forward | 549 | 1 | 182 | 19.75 | 8.8 | −0.6978 | Nucleus |
| TaBZR2.4 | TraesCS3B02G156600 | 3B:149453779-149455098 | Forward | 1065 | 1 | 354 | 37.28 | 8 | −0.5166 | Nucleus |
| TaBZR2.5 | TraesCS3D02G125100 | 3D:83703425-83703710 | Forward | 537 | 1 | 178 | 19.26 | 9.1 | −0.7511 | Nucleus |
| TaBZR2.6 | TraesCS3D02G139300 | 3D:98871950-98873193 | Forward | 1077 | 1 | 358 | 37.51 | 8 | −0.5153 | Nucleus |
| TaBZR3.1 | TraesCS4B02G009900 | 4B:6146066-6147444 | Forward | 1656 | 7 | 551 | 62.34 | 5.2 | −0.5284 | Nucleus |
| TaBZR3.2 | TraesCS4D02G006100 | 4D:3399752-3399889 | Reverse | 2061 | 8 | 686 | 75.47 | 5.2 | −0.3704 | Nucleus |
| TaBZR4.1 | TraesCS6A02G085800 | 6A:54119841-54119972 | Reverse | 1959 | 6 | 652 | 73.1 | 6 | −0.3699 | Nucleus |
| TaBZR4.2 | TraesCS6A02G338000 | 6A:571795527-571797497 | Forward | 1044 | 1 | 347 | 36.41 | 8 | −0.5642 | Nucleus |
| TaBZR4.3 | TraesCS6B02G116400 | 6B:101917320-101917475 | Forward | 2019 | 8 | 672 | 75.48 | 6.5 | −0.3483 | Nucleus |
| TaBZR4.4 | TraesCS6B02G368700 | 6B:642915742-642917800 | Forward | 1071 | 1 | 356 | 37.71 | 8.3 | −0.6092 | Nucleus |
| TaBZR4.5 | TraesCS6D02G318800 | 6D:427088235-427090524 | Forward | 1047 | 1 | 348 | 36.44 | 7.8 | −0.516 | Nucleus |
| TaBZR5.1 | TraesCS7A02G354800 | 7A:519131141-519132953 | Forward | 1080 | 1 | 359 | 37.84 | 7.5 | −0.6506 | Nucleus |
| TaBZR5.2 | TraesCS7B02G272900 | 7B:500731158-500732708 | Reverse | 1080 | 1 | 359 | 37.86 | 7.5 | −0.6225 | Nucleus |
| TaBZR5.3 | TraesCS7D02G368000 | 7D:476687645-476689882 | Reverse | 1080 | 1 | 359 | 37.8 | 7.5 | −0.6197 | Nucleus |
| TaBZR6 | TraesCSU02G078100 | Un:70283162-70283317 | Forward | 1959 | 8 | 652 | 72.95 | 6 | −0.3601 | Nucleus |
| Plant Species | Genome Size (Approx.) | Coding Genes | BZR Genes |
|---|---|---|---|
| Triticum aestivum (6n *) | 17 Gb | 107,891 | 20 |
| Arabidopsis thaliana (2n) | 135 MB | 27,655 | 6 |
| Zea mays (2n) | 2.4 Gb | 39,591 | 11 |
| Glycine max (2n) | 1.15 Gb | 55,897 | 16 |
| Cajanus cajan (2n) | 833 MB | 48,680 | 6 |
| Medicago sativa (2n) | 360 MB | 50,444 | 7 |
| Cicer arietinum (2n) | 738 MB | 25,680 | 6 |
| Solanum lycopersicum (2n) | 828 MB | 34,658 | 9 |
| Beta vulgaris (2n) | 758 MB | 26,521 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kesawat, M.S.; Kherawat, B.S.; Singh, A.; Dey, P.; Kabi, M.; Debnath, D.; Saha, D.; Khandual, A.; Rout, S.; Manorama; et al. Genome-Wide Identification and Characterization of the Brassinazole-resistant (BZR) Gene Family and Its Expression in the Various Developmental Stage and Stress Conditions in Wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2021, 22, 8743. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168743
Kesawat MS, Kherawat BS, Singh A, Dey P, Kabi M, Debnath D, Saha D, Khandual A, Rout S, Manorama, et al. Genome-Wide Identification and Characterization of the Brassinazole-resistant (BZR) Gene Family and Its Expression in the Various Developmental Stage and Stress Conditions in Wheat (Triticum aestivum L.). International Journal of Molecular Sciences. 2021; 22(16):8743. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168743
Chicago/Turabian StyleKesawat, Mahipal Singh, Bhagwat Singh Kherawat, Anupama Singh, Prajjal Dey, Mandakini Kabi, Debanjana Debnath, Debanjana Saha, Ansuman Khandual, Sandeep Rout, Manorama, and et al. 2021. "Genome-Wide Identification and Characterization of the Brassinazole-resistant (BZR) Gene Family and Its Expression in the Various Developmental Stage and Stress Conditions in Wheat (Triticum aestivum L.)" International Journal of Molecular Sciences 22, no. 16: 8743. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168743