
Advanced Glycation End Products Are Retained in Decellularized Muscle Matrix Derived from Aged Skeletal Muscle

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Histological Assessment of Whole Muscle
2.2. Age-Associated Effects in Whole Muscle Proteins
2.3. Histological Assessment of Decellularized Muscle
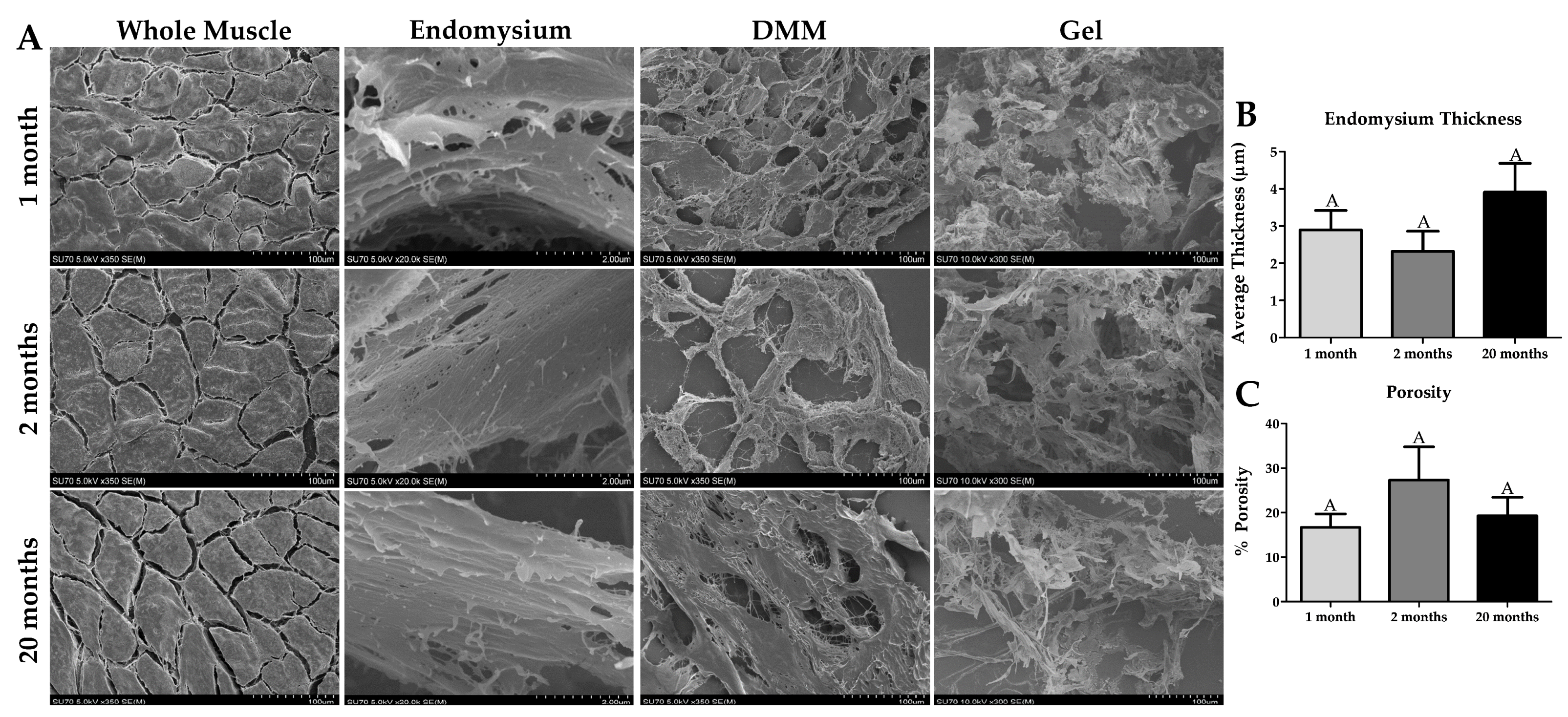
2.4. Ultrastructural Properties of Whole Muscle, DMM, and DMM Gels at Different Ages
2.5. Age-Dependent Effects in Collagen Are Preserved in Processed Muscle
2.6. Increased Stiffness in DMM from Old Muscle
2.7. Advanced Glycation End Products Are Retained in DMM
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hamerman, D. Aging and the Musculoskeletal System. Ann. Rheum. Dis. 1997, 56, 578–585. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.M.; Jang, H.C.; Lim, S. Differences among Skeletal Muscle Mass Indices Derived from Height-, Weight-, and Body Mass Index-Adjusted Models in Assessing Sarcopenia. Korean J. Intern. Med. 2016, 31, 643–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siparsky, P.N.; Kirkendall, D.T.; Garrett, W.E., Jr. Muscle Changes in Aging: Understanding Sarcopenia. Sports Health 2014, 6, 36. [Google Scholar] [CrossRef] [Green Version]
- Arthur, W. On the relationship between strength of grip and certain mental and sensory characters. Biometrika 1924, 16, 299–327. [Google Scholar] [CrossRef]
- Walston, J.D. Sarcopenia in Older Adults. Curr. Opin. Rheumatol. 2012, 24, 623–627. [Google Scholar] [CrossRef] [Green Version]
- Doherty, T.J. Invited Review: Aging and Sarcopenia. J. Appl. Physiol. 2003, 95, 1717–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, L.; Degens, H.; Li, M.; Salviati, L.; Lee, Y.I.; Thompson, W.; Kirkland, J.L.; Sandri, M. Sarcopenia: Aging-Related Loss of Muscle Mass and Function. Physiol. Rev. 2019, 99, 427–511. [Google Scholar] [CrossRef] [PubMed]
- Maillard, L.-C. Condensation Des Acides Amines En Presence de La Glycerine; Cycloglycylglycine et Polypeptides. C. R. Hebd. Seances Acad. Sci. 1911, 153, 1078–1080. [Google Scholar]
- Kawamura, S. Seventy Years of the Maillard Reaction. In The Maillard Reaction in Foods and Nutrition, ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1983; Volume 215, pp. 1–3. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, A.; Yabu, A.; Nakamura, H. Advanced Glycation End Products in Musculoskeletal System and Disorders. Methods 2020. [Google Scholar] [CrossRef] [PubMed]
- Svensson, R.B.; Smith, S.T.; Moyer, P.J.; Magnusson, S.P. Effects of Maturation and Advanced Glycation on Tensile Mechanics of Collagen Fibrils from Rat Tail and Achilles Tendons. Acta Biomater. 2018, 70, 270–280. [Google Scholar] [CrossRef]
- Gillies, A.R.; Lieber, R.L. Structure and Function of the Skeletal Muscle Extracellular Matrix. Muscle Nerve 2011, 44, 318–331. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite Cells and the Muscle Stem Cell Niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar] [CrossRef] [Green Version]
- Bentzinger, C.F.; Wang, Y.X.; von Maltzahn, J.; Soleimani, V.D.; Yin, H.; Rudnicki, M.A. Fibronectin Regulates Wnt7a Signaling and Satellite Cell Expansion. Cell Stem Cell 2013, 12, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Bentzinger, C.F.; von Maltzahn, J.; Rudnicki, M.A. Extrinsic Regulation of Satellite Cell Specification. Stem Cell Res. Ther. 2010, 1, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rayagiri, S.S.; Ranaldi, D.; Raven, A.; Mohamad Azhar, N.I.F.; Lefebvre, O.; Zammit, P.S.; Borycki, A.-G. Basal Lamina Remodeling at the Skeletal Muscle Stem Cell Niche Mediates Stem Cell Self-Renewal. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, W. On the Minute Structure and Movements of Voluntary Muscle. R. J. R. Taylor 1840, 130, 457–501. [Google Scholar]
- Seynnes, O.; Schiaffino, S.; Blottner, D.; Csapo, R.; Gumpenberger, M.; Wessner, B. Skeletal Muscle Extracellular Matrix—What Do We Know About Its Composition, Regulation, and Physiological Roles? A Narrative Review. Front. Physiol. 2020, 11, 253. [Google Scholar] [CrossRef] [Green Version]
- Stearns-Reider, K.M.; D’Amore, A.; Beezhold, K.; Rothrauff, B.; Cavalli, L.; Wagner, W.R.; Vorp, D.A.; Tsamis, A.; Shinde, S.; Zhang, C.; et al. Aging of the Skeletal Muscle Extracellular Matrix Drives a Stem Cell Fibrogenic Conversion. Aging Cell 2017, 16, 518–528. [Google Scholar] [CrossRef] [Green Version]
- Wood, L.K.; Kayupov, E.; Gumucio, J.P.; Mendias, C.L.; Claflin, D.R.; Brooks, S.V. Intrinsic Stiffness of Extracellular Matrix Increases with Age in Skeletal Muscles of Mice. J. Appl. Physiol. 2014, 117, 363–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauro, A. Satellite Cell of Skeletal Muscle FibersS. J. Biophys. Biochem. Cytol. 1961, 9, 493–495. [Google Scholar] [CrossRef]
- Bernet, J.D.; Doles, J.D.; Hall, J.K.; Kelly Tanaka, K.; Carter, T.A.; Olwin, B.B. P38 MAPK Signaling Underlies a Cell-Autonomous Loss of Stem Cell Self-Renewal in Skeletal Muscle of Aged Mice. Nat. Med. 2014, 20, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Urciuolo, A.; De Coppi, P. Decellularized Tissue for Muscle Regeneration. Int. J. Mol. Sci. 2018, 19, 2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilpin, A.; Yang, Y. Decellularization Strategies for Regenerative Medicine: From Processing Techniques to Applications. Biomed. Res. Int. 2017, 2017, 9831534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Hu, Z.Q.; Turner, N.J.; Teng, S.F.; Cheng, W.Y.; Zhou, H.Y.; Zhang, L.; Hu, H.W.; Wang, Q.; Badylak, S.F. Perfusion-Decellularized Skeletal Muscle as a Three-Dimensional Scaffold with a Vascular Network Template. Biomaterials 2016, 89, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Porzionato, A.; Sfriso, M.M.; Pontini, A.; Macchi, V.; Petrelli, L.; Pavan, P.G.; Natali, A.N.; Bassetto, F.; Vindigni, V.; De Caro, R. Decellularized Human Skeletal Muscle as Biologic Scaffold for Reconstructive Surgery. Int. J. Mol. Sci. 2015, 16, 14808–14831. [Google Scholar] [CrossRef] [Green Version]
- Kasukonis, B.; Kim, J.; Brown, L.; Jones, J.; Ahmadi, S.; Washington, T.; Wolchok, J. Codelivery of Infusion Decellularized Skeletal Muscle with Minced Muscle Autografts Improved Recovery from Volumetric Muscle Loss Injury in a Rat Model. Tissue Eng. Part. A 2016, 22, 1151–1163. [Google Scholar] [CrossRef] [Green Version]
- Wolf, M.T.; Dearth, C.L.; Sonnenberg, S.B.; Loboa, E.G.; Badylak, S.F. Naturally Derived and Synthetic Scaffolds for Skeletal Muscle Reconstruction. Adv. Drug Deliv. Rev. 2015, 84, 208–221. [Google Scholar] [CrossRef] [Green Version]
- Urciuolo, A.; Urbani, L.; Perin, S.; Maghsoudlou, P.; Scottoni, F.; Gjinovci, A.; Collins-Hooper, H.; Loukogeorgakis, S.; Tyraskis, A.; Torelli, S.; et al. Decellularised Skeletal Muscles Allow Functional Muscle Regeneration by Promoting Host Cell Migration. Sci. Rep. 2018, 8, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Perniconi, B.; Coletti, D.; Aulino, P.; Costa, A.; Aprile, P.; Santacroce, L.; Chiaravalloti, E.; Coquelin, L.; Chevallier, N.; Teodori, L.; et al. Muscle Acellular Scaffold as a Biomaterial: Effects on C2C12 Cell Differentiation and Interaction with the Murine Host Environment. Front. Physiol. 2014, 5, 354. [Google Scholar] [CrossRef] [Green Version]
- McClure, M.J.; Cohen, D.J.; Ramey, A.N.; Bivens, C.B.; Mallu, S.; Isaacs, J.E.; Imming, E.; Huang, Y.-C.; Sunwoo, M.; Schwartz, Z.; et al. Decellularized Muscle Supports New Muscle Fibers and Improves Function Following Volumetric Injury. Tissue Eng. Part. A 2018, 24, 1228–1241. [Google Scholar] [CrossRef]
- Chaturvedi, V.; Dye, D.E.; Kinnear, B.F.; Van Kuppevelt, T.H.; Grounds, M.D.; Coombe, D.R. Interactions between Skeletal Muscle Myoblasts and Their Extracellular Matrix Revealed by a Serum Free Culture System. PLoS ONE 2015, 10, e0127675. [Google Scholar] [CrossRef]
- Kim, J.T.; Kasukonis, B.M.; Brown, L.A.; Washington, T.A.; Wolchok, J.C. Recovery from Volumetric Muscle Loss Injury: A Comparison between Young and Aged Rats. Exp. Gerontol. 2016, 83, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.T.; Kasukonis, B.; Dunlap, G.; Perry, R.; Washington, T.; Wolchok, J.C. Regenerative Repair of Volumetric Muscle Loss Injury Is Sensitive to Age. Tissue Eng. Part. A 2020, 26, 3–14. [Google Scholar] [CrossRef]
- McClure, M.J.; Olson, L.C.; Cohen, D.J.; Huang, Y.C.; Zhang, S.; Nguyen, T.; Boyan, B.D.; Schwartz, Z. Rnu (Foxn1Rnu-Nude) Rats Demonstrate an Improved Ability to Regenerate Muscle in a Volumetric Muscle Injury Compared to Sprague Dawley Rats. Bioengineering 2021, 8, 12. [Google Scholar] [CrossRef]
- Giacomello, E.; Crea, E.; Torelli, L.; Bergamo, A.; Reggiani, C.; Sava, G.; Toniolo, L. Age Dependent Modification of the Metabolic Profile of the Tibialis Anterior Muscle Fibers in C57BL/6J Mice. Int. J. Mol. Sci. 2020, 21, 3923. [Google Scholar] [CrossRef] [PubMed]
- Agbulut, O.; Destombes, J.; Thiesson, D.; Butler-Browne, G. Age-Related Appearance of Tubular Aggregates in the Skeletal Muscle of Almost All Male Inbred Mice. Histochem. Cell Biol. 2000, 114, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Cundy, T.; Reid, I.R.; Grey, A. Metabolic Bone Disease. Clin. Biochem. Metab. Clin. Asp. Third Ed. 2014, 604–635. [Google Scholar] [CrossRef]
- Joanna, S.; Constance, C. Skeletal Muscle Fiber Types in the Adult Mouse. Acta Neurol. Scand. 1976, 54, 45–56. [Google Scholar] [CrossRef]
- Robert, S.S.; William, J.K.; Robert, S.H.; Andy, C.F.; Jerry, D.M.; Gerson, E.C. Fiber Type Composition of Four Hindlimb Muscles of Adult Fisher 344 Rats. Histochem. Cell Biol. 1999, 111, 117–123. [Google Scholar] [CrossRef]
- Berg, B.N. Muscular Dystrophy in Aging Rats. J. Gerontol. 1956, 11, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Real-Martinez, A.; Brull, A.; Huerta, J.; Tarrasó, G.; Lucia, A.; Martin, M.A.; Arenas, J.; Andreu, A.L.; Nogales-Gadea, G.; Vissing, J.; et al. Low Survival Rate and Muscle Fiber-Dependent Aging Effects in the McArdle Disease Mouse Model. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Schaub, M.C. The Ageing of Collagen in the Striated Muscle. Gerontology 1963, 8, 16–35. [Google Scholar] [CrossRef]
- Carmichael, D.J.; Lawrie, R.A. Bovine Collagen. I. Changes in Collagen Solubility with Animal Age. Int. J. Food Sci. Technol. 1967, 2, 299–311. [Google Scholar] [CrossRef]
- Haseeb, M.A.; Patnaik, B.K. Age-Related Changes in Collagenous and Noncollagenous Proteins of Skeletal Muscle of a Short-Lived Species of Reptile. Gerontology 1978, 24, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Verzijl, N.; DeGroot, J.; Thorpe, S.R.; Bank, R.A.; Shaw, J.N.; Lyons, T.J.; Bijlsma, J.W.J.; Lafeber, F.P.J.G.; Baynes, J.W.; TeKoppele, J.M. Effect of Collagen Turnover on the Accumulation of Advanced Glycation End Products. J. Biol. Chem. 2000, 275, 39027–39031. [Google Scholar] [CrossRef] [Green Version]
- Haus, J.M.; Carrithers, J.A.; Trappe, S.W.; Trappe, T.A. Collagen, Cross-Linking, and Advanced Glycation End Products in Aging Human Skeletal Muscle. J. Appl. Physiol. 2007, 103, 2068–2076. [Google Scholar] [CrossRef] [PubMed]
- Graber, T.G.; Kim, J.H.; Grange, R.W.; Mcloon, L.K.; Thompson, L.V. C57BL/6 Life Span Study: Age-Related Declines in Muscle Power Production and Contractile Velocity. Age 2015, 37, 1–16. [Google Scholar] [CrossRef]
- Hill, C.; James, R.S.; Cox, V.M.; Seebacher, F.; Tallis, J. Age-Related Changes in Isolated Mouse Skeletal Muscle Function Are Dependent on Sex, Muscle, and Contractility Mode. Am. J. Physiol. Integr. Comp. Physiol. 2020, 319, R296–R314. [Google Scholar] [CrossRef]
- Uchitomi, R.; Hatazawa, Y.; Senoo, N.; Yoshioka, K.; Fujita, M.; Shimizu, T.; Miura, S.; Ono, Y.; Kamei, Y. Metabolomic Analysis of Skeletal Muscle in Aged Mice. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Saulnier, P.-J.; Wheelock, K.M.; Howell, S.; Weil, E.J.; Tanamas, S.K.; Knowler, W.C.; Lemley, K.V.; Mauer, M.; Yee, B.; Nelson, R.G.; et al. Advanced Glycation End Products Predict Loss of Renal Function and Correlate With Lesions of Diabetic Kidney Disease in American Indians With Type 2 Diabetes. Diabetes 2016, 65, 3744–3753. [Google Scholar] [CrossRef] [Green Version]
- Flurkey, K.; Currer, J.M.; Harrison, D.E. Mouse Models in Aging Research. Mouse Biomed. Res. 2007, 3, 637–672. [Google Scholar] [CrossRef]
- Yuan, R.; Peters, L.L.; Paigen, B. Mice as a Mammalian Model for Research on the Genetics of Aging. ILAR J. 2011, 52, 4. [Google Scholar] [CrossRef] [Green Version]
- Valentine, M.S.; Link, P.A.; Herbert, J.A.; Gninzeko, F.J.K.; Schneck, M.B.; Shankar, K.; Nkwocha, J.; Reynolds, A.M.; Heise, R.L. Inflammation and Monocyte Recruitment Due to Aging and Mechanical Stretch in Alveolar Epithelium Are Inhibited by the Molecular Chaperone 4-Phenylbutyrate. Cell. Mol. Bioeng. 2018, 11, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Cousin, W.; Ho, M.L.; Desai, R.; Tham, A.; Chen, R.Y.; Kung, S.; Elabd, C.; Conboy, I.M. Regenerative Capacity of Old Muscle Stem Cells Declines without Significant Accumulation of DNA Damage. PLoS ONE 2013, 8, e63528. [Google Scholar] [CrossRef]
- Virginia, L.F.; Reed, A.A.; Ted, A.B.; Steven, J.S. Bone Development and Age-Related Bone Loss in Male C57BL/6J Mice. Bone 2003, 33, 387–398. [Google Scholar] [CrossRef]
- Linn, S.C.; Mustonen, A.M.; Silva, K.A.; Kennedy, V.E.; Sundberg, B.A.; Bechtold, L.S.; Alghamdi, S.; Hoehndorf, R.; Schofield, P.N.; Sundberg, D.J.P. Nail Abnormalities Identified in an Aging Study of 30 Inbred Mouse Strains. Exp. Dermatol. 2019, 28, 383–390. [Google Scholar] [CrossRef]
- Sugiyama, Y.; Naito, K.; Goto, K.; Kojima, Y.; Furuhata, A.; Igarashi, M.; Nagaoka, I.; Kaneko, K. Effect of Aging on the Tendon Structure and Tendon-associated Gene Expression in Mouse Foot Flexor Tendon. Biomed. Rep. 2019, 10, 238–244. [Google Scholar] [CrossRef] [Green Version]
- Tarrant, J.C.; Savickas, P.; Omodho, L.; Spinazzi, M.; Radaelli, E. Spontaneous Incidental Brain Lesions in C57BL/6J Mice. Vet. Pathol. 2019, 57, 172–182. [Google Scholar] [CrossRef]
- Gehwolf, R.; Wagner, A.; Lehner, C.; Bradshaw, A.D.; Scharler, C.; Niestrawska, J.A.; Holzapfel, G.A.; Bauer, H.-C.; Tempfer, H.; Traweger, A. Pleiotropic Roles of the Matricellular Protein Sparc in Tendon Maturation and Ageing. Sci. Rep. 2016, 6, 1–15. [Google Scholar] [CrossRef]
- Huang, H.; Skelly, J.D.; Ayers, D.C.; Song, J. Age-Dependent Changes in the Articular Cartilage and Subchondral Bone of C57BL/6 Mice after Surgical Destabilization of Medial Meniscus. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menees, K.B.; Earls, R.H.; Chung, J.; Jernigan, J.; Filipov, N.M.; Carpenter, J.M.; Lee, J.-K. Sex- and Age-dependent Alterations of Splenic Immune Cell Profile and NK Cell Phenotypes and Function in C57BL/6J Mice. Immun. Ageing 2021, 18, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-A.; Jeong, J.-J.; Yoo, S.-Y.; Kim, D.-H. Gut Microbiota Lipopolysaccharide Accelerates Inflamm-Aging in Mice. BMC Microbiol. 2016, 16, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mobbs, C.V.; Gee, D.M.; Finch, C.E. Reproductive Senescence in Female C57BL/6J Mice: Ovarian Impairments and Neuroendocrine Impairments That Are Partially Reversible and Delayable by Ovariectomy. Endocrinology 1984, 115, 1653–1662. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, X.; Han, J.; Fan, Z.; Sadia, S.; Zhang, R.; Guo, Y.; Jiang, Y.; Wu, Y. AKT and Its Related Molecular Feature in Aged Mice Skin. PLoS ONE 2017, 12, e0178969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovanen, V.; Suominen, H.; Peltonen, L. Effects of Aging and Life-Long Physical Training on Collagen in Slow and Fast Skeletal Muscle in Rats—A Morphometric and Immuno-Histochemical Study. Cell Tissue Res. 1987, 248, 247–255. [Google Scholar] [CrossRef]
- Petrany, M.J.; Swoboda, C.O.; Sun, C.; Chetal, K.; Chen, X.; Weirauch, M.T.; Salomonis, N.; Millay, D.P. Single-Nucleus RNA-Seq Identifies Transcriptional Heterogeneity in Multinucleated Skeletal Myofibers. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Herchenhan, A.; Uhlenbrock, F.; Eliasson, P.; Weis, M.; Eyre, D.; Kadler, K.E.; Magnusson, S.P.; Kjaer, M. Lysyl Oxidase Activity Is Required for Ordered Collagen Fibrillogenesis by Tendon Cells. J. Biol. Chem. 2015, 290, 16440–16450. [Google Scholar] [CrossRef] [Green Version]
- Chakkalakal, J.V.; Jones, K.M.; Albert Basson, M.; Brack, A.S. The Aged Niche Disrupts Muscle Stem Cell Quiescence. Nature 2012, 490, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Donalies, M.; Cramer, M.; Ringwald, M.; Starzinski-Powitz, A. Expression of M-Cadherin, a Member of the Cadherin Multigene Family, Correlates with Differentiation of Skeletal Muscle Cells. Proc. Natl. Acad. Sci. USA 1991, 88, 8024–8028. [Google Scholar] [CrossRef] [Green Version]
- Marti, M.; Montserrat, N.; Pardo, C.; Mulero, L.; Miquel-Serra, L.; Cavaco Rodrigues, A.M.; Andrés Vaquero, J.; Kuebler, B.; Morera, C.; Barrero, M.J.; et al. M-Cadherin-Mediated Intercellular Interactions Activate Satellite Cell Division. J. Cell Sci. 2013, 126, 5116–5131. [Google Scholar] [CrossRef] [Green Version]
- Sajko, P.; Kubínová, L.; Cvetko, E.; Kreft, M.; Wernig, A.; Er, I. Frequency of M-Cadherin-Stained Satellite Cells Declines in Human Muscles During Aging. J. Histochem. Cytochem. 2004, 52, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, R.; Walsh, F.S. The Cell Adhesion Molecule M-Cadherin Is Specifically Expressed in Developing and Regenerating, but Not Denervated Skeletal Muscle. Development 1993, 117, 1409–1420. [Google Scholar] [CrossRef] [PubMed]
- Carlson, B.M. Denervation and the Aging of Skeletal Muscle. Basic Appl. Myol. 2004, 14, 135–139. [Google Scholar]
- Dedkov, E.I.; Kostrominova, T.Y.; Borisov, A.B.; Carlson, B.M. MyoD and Myogenin Protein Expression in Skeletal Muscles of Senile Rats. Cell Tissue Res. 2003, 311, 401–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musarò, A.; De Cusella Angelis, M.G.; Germani, A.; Ciccarelli, C.; Molinaro, M.; Zani, B.M. Enhanced Expression of Myogenic Regulatory Genes in Aging Skeletal Muscle. Exp. Cell Res. 1995, 221, 241–248. [Google Scholar] [CrossRef]
- Riuzzi, F.; Sorci, G.; Sagheddu, R.; Chiappalupi, S.; Salvadori, L.; Donato, R. RAGE in the Pathophysiology of Skeletal Muscle. J. Cachexia Sarcopenia Muscle 2018, 9, 1213–1234. [Google Scholar] [CrossRef] [Green Version]
- Sorci, G.; Riuzzi, F.; Arcuri, C.; Giambanco, I.; Donato, R. Amphoterin Stimulates Myogenesis and Counteracts the Antimyogenic Factors Basic Fibroblast Growth Factor and S100B via RAGE Binding Downloaded From. Mol. Cell. Biol. 2004, 24, 4880–4894. [Google Scholar] [CrossRef] [Green Version]
- Riuzzi, F.; Sorci, G.; Sagheddu, R.; Donato, R. HMGB1-RAGE Regulates Muscle Satellite Cell Homeostasis through P38-MAPK- and Myogenindependent Repression of Pax7 Transcription. J. Cell Sci. 2012, 125, 1440–1454. [Google Scholar] [CrossRef] [Green Version]
- Beccafico, S.; Riuzzi, F.; Puglielli, C.; Mancinelli, R.; Fulle, S.; Sorci, G.; Donato, R. Human Muscle Satellite Cells Show Age-Related Differential Expression of S100B Protein and RAGE. Age 2011, 33, 523–541. [Google Scholar] [CrossRef] [Green Version]
- Boppart, M.D.; Mahmassani, Z.S. Integrin Signaling: Linking Mechanical Stimulation to Skeletal Muscle Hypertrophy. Am. J. Physiol. Physiol. 2019, 317, C629–C641. [Google Scholar] [CrossRef] [PubMed]
- Sarathy, A.; Wuebbles, R.D.; Fontelonga, T.M.; Tarchione, A.R.; Mathews Griner, L.A.; Heredia, D.J.; Nunes, A.M.; Duan, S.; Brewer, P.D.; Van Ry, T.; et al. SU9516 Increases A7β1 Integrin and Ameliorates Disease Progression in the Mdx Mouse Model of Duchenne Muscular Dystrophy. Mol. Ther. 2017, 25, 1395–1407. [Google Scholar] [CrossRef] [Green Version]
- Rozo, M.; Li, L.; Fan, C.M. Targeting Β1-Integrin Signaling Enhances Regeneration in Aged and Dystrophic Muscle in Mice. Nat. Med. 2016, 22, 889–896. [Google Scholar] [CrossRef] [Green Version]
- Garg, K.; Mahmassani, Z.S.; Dvoretskiy, S.; Valero, M.C.; Huntsman, H.D.; Lapp, S.; Wu, Y.-F.; Hauschka, S.D.; Burkin, D.J.; Boppart, M.D. Laminin-111 Improves the Anabolic Response to Mechanical Load in Aged Skeletal Muscle. J. Gerontol. Ser. A 2021, 76, 586–590. [Google Scholar] [CrossRef] [PubMed]
- Frontera, W.R.; Miljkovic, N.; Lim, J.-Y.; Miljkovic, I. Aging of Skeletal Muscle Fibers. Ann. Rehabil. Med. Rev. Artic. Ann. Rehabil Med. 2015, 39, 155–162. [Google Scholar] [CrossRef]
- Elliott, B.; Youl Moon, H.; Levinger, I.; Reggiani, C.; Naro, F.; Venturelli, M.; Monaco, L.; Toniolo, L.; Muti, E.; Milanese, C.; et al. Skeletal Muscle Fiber Size and Gene Expression in the Oldest-Old With Differing Degrees of Mobility. Front. Physiol. 2019, 10, 313. [Google Scholar] [CrossRef] [Green Version]
- Snow, L.M.; McLoon, L.K.; Thompson, L.V. Adult and Developmental Myosin Heavy Chain Isoforms in Soleus Muscle of Aging Fischer Brown Norway Rat. Anat. Rec. Part. A Discov. Mol. Cell. Evol. Biol. 2005, 286, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Ballak, S.B.; Degens, H.; de Haan, A.; Jaspers, R.T. Aging Related Changes in Determinants of Muscle Force Generating Capacity: A Comparison of Muscle Aging in Men and Male Rodents. Ageing Res. Rev. 2014, 14, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Haus, J.M.; Carrithers, J.A.; Miller, B.I.; Trappe, T.A. Intramuscular Collagen Cross-Linking in Aging Men and Women. FASEB J. 2007, 21, LB118. [Google Scholar] [CrossRef]
- Rahman, N.; O’neill, E.; Irnaten, M.; Wallace, D.; O’brien, C. Corneal Stiffness and Collagen Cross-Linking Proteins in Glaucoma: Potential for Novel Therapeutic Strategy. J. Ocul. Pharmacol. Ther. 2020, 36, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Pavan, P.; Monti, E.; Bondí, M.; Fan, C.; Stecco, C.; Narici, M.; Reggiani, C.; Marcucci, L. Alterations of Extracellular Matrix Mechanical Properties Contribute to Age-Related Functional Impairment of Human Skeletal Muscles. Int. J. Mol. Sci. Artic 2020, 21, 3992. [Google Scholar] [CrossRef] [PubMed]
- Olivares-Navarrete, R.; Lee, E.M.; Smith, K.; Hyzy, S.L.; Doroudi, M.; Williams, J.K. Substrate Stiffness Controls Osteoblastic and Chondrocytic Differentiation of Mesenchymal Stem Cells without Exogenous Stimuli. PLoS ONE 2017, 12, 170312. [Google Scholar] [CrossRef] [PubMed]
- Rowlands, A.S.; George, P.A.; Cooper-White, J.J. Directing Osteogenic and Myogenic Differentiation of MSCs: Interplay of Stiffness and Adhesive Ligand Presentation. Am. J. Physiol. Cell Physiol. 2008, 295, 1037–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imayama, S.; Braverman, I.M. A Hypothetical Explanation for the Aging of Skin Chronologic Alteration of the Three-Dimensional Arrangement of Collagen and Elastic Fibers in Connective Tissue. Am. J. Pathol. 1989, 134, 1019. [Google Scholar]
- Kragstrup, T.W.; Kjaer, M.; Mackey, A.L.; Wenzel Kragstrup, T. Structural, Biochemical, Cellular, and Functional Changes in Skeletal Muscle Extracellular Matrix with Aging. Scand. J. Med. Sci. Sports 2011, 21, 749–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabbani, N.; Thornalley, P.J. Hidden Complexities in the Measurement of Fructosyl-Lysine and Advanced Glycation End Products for Risk Prediction of Vascular Complications of Diabetes. Diabetes 2015, 64, 9–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhar, A.; Udumula, M.P.; Medapi, B.; Bhat, A.; Dhar, I.; Malapati, P.; Babu, M.S.; Kalra, J.; Sriram, D.; Desai, K.M. Pharmacological Evaluation of Novel Alagebrium Analogs as Methylglyoxal Scavengers in Vitro in Cardiac Myocytes and in Vivo in SD Rats. Int. J. Cardiol. 2016, 223, 581–589. [Google Scholar] [CrossRef]
- Johnson, T.D.; DeQuach, J.A.; Gaetani, R.; Ungerleider, J.; Elhag, D.; Nigam, V.; Behfar, A.; Christman, K.L. Human versus Porcine Tissue Sourcing for an Injectable Myocardial Matrix Hydrogel. Biomater. Sci. 2014, 2014, 60283D. [Google Scholar] [CrossRef] [Green Version]
- Wedgwood, J.; Freemont, A.J.; Tirelli, N. Rheological and Turbidity Study of Fibrin Hydrogels. Macromol. Symp. 2013, 334, 117–125. [Google Scholar] [CrossRef]
- Briguet, A.; Courdier-Fruh, I.; Foster, M.; Meier, T.; Magyar, J.P. Histological Parameters for the Quantitative Assessment of Muscular Dystrophy in the Mdx-Mouse. Neuromuscul. Disord. 2004, 14, 675–682. [Google Scholar] [CrossRef]
- Vigelsø, A.; Dybboe, R.; Hansen, C.N.; Dela, F.; Helge, J.W.; Grau, A.G. GAPDH and β-Actin Protein Decreases with Aging, Making Stain-Free Technology a Superior Loading Control in Western Blotting of Human Skeletal Muscle. J. Appl. Physiol. 2015, 118, 386–394. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olson, L.C.; Nguyen, T.M.; Heise, R.L.; Boyan, B.D.; Schwartz, Z.; McClure, M.J. Advanced Glycation End Products Are Retained in Decellularized Muscle Matrix Derived from Aged Skeletal Muscle. Int. J. Mol. Sci. 2021, 22, 8832. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168832
Olson LC, Nguyen TM, Heise RL, Boyan BD, Schwartz Z, McClure MJ. Advanced Glycation End Products Are Retained in Decellularized Muscle Matrix Derived from Aged Skeletal Muscle. International Journal of Molecular Sciences. 2021; 22(16):8832. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168832
Chicago/Turabian StyleOlson, Lucas C., Tri M. Nguyen, Rebecca L. Heise, Barbara D. Boyan, Zvi Schwartz, and Michael J. McClure. 2021. "Advanced Glycation End Products Are Retained in Decellularized Muscle Matrix Derived from Aged Skeletal Muscle" International Journal of Molecular Sciences 22, no. 16: 8832. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168832