
Applicability of Scrape Loading-Dye Transfer Assay for Non-Genotoxic Carcinogen Testing


Abstract
:1. Introduction
“With respect to cancer causation, integration of the analyses suggest that the inhibition of gap junctional intercellular communication is involved in non-genotoxic cancer induction or in the non-genotoxic phase of the carcinogenic process (such as inflammation, cell toxicity, cell proliferation, inhibition of cell differentiation, and apoptosis)” [1].
“Here, we review the literature surrounding connexins in cancer cells in terms of specific connexin functions and propose that connexins function up stream of most, if not all, of the hallmarks of cancer” [2].
2. GJIC as the Key Mechanism in Tissue Homeostasis
3. Cell Lines and Methods for In Vitro GJIC Assessment

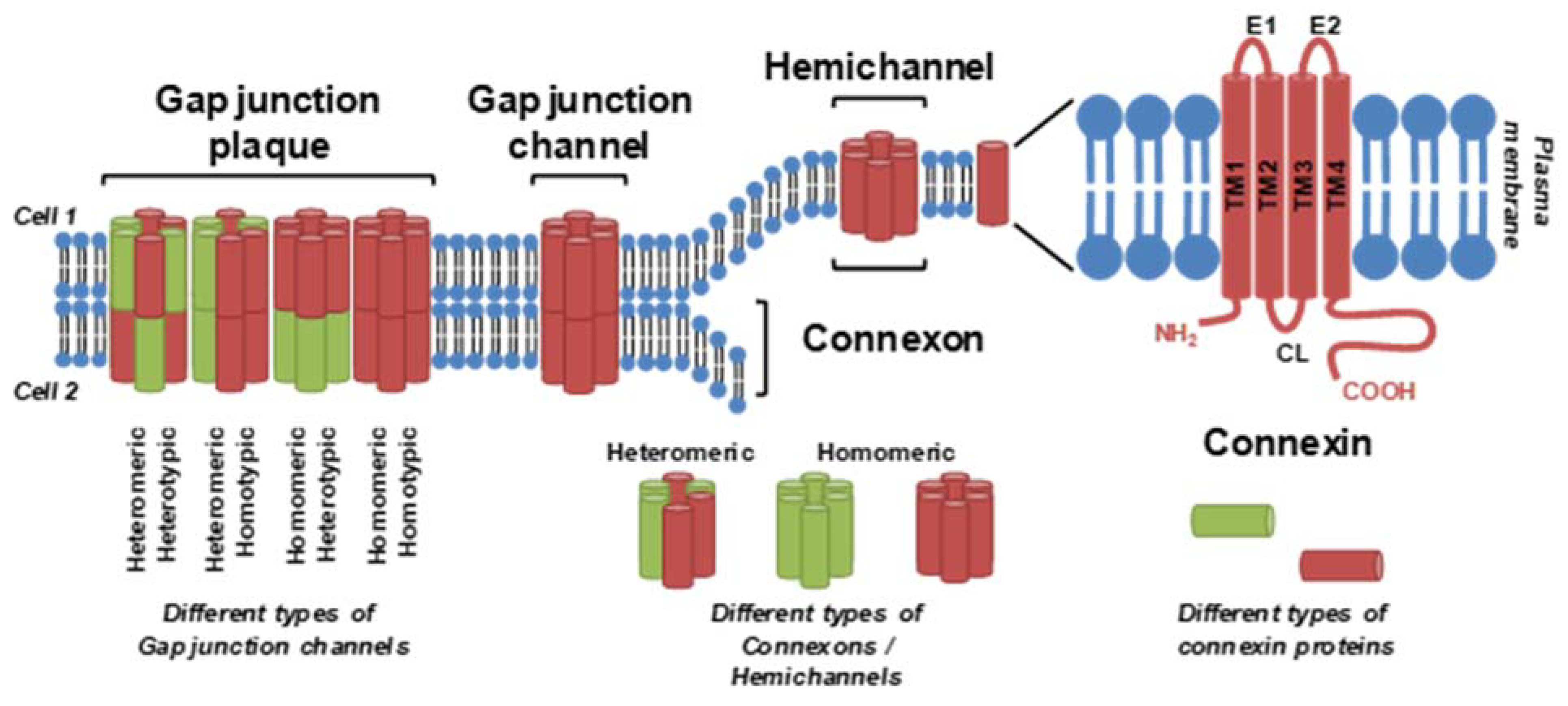{kind=link}
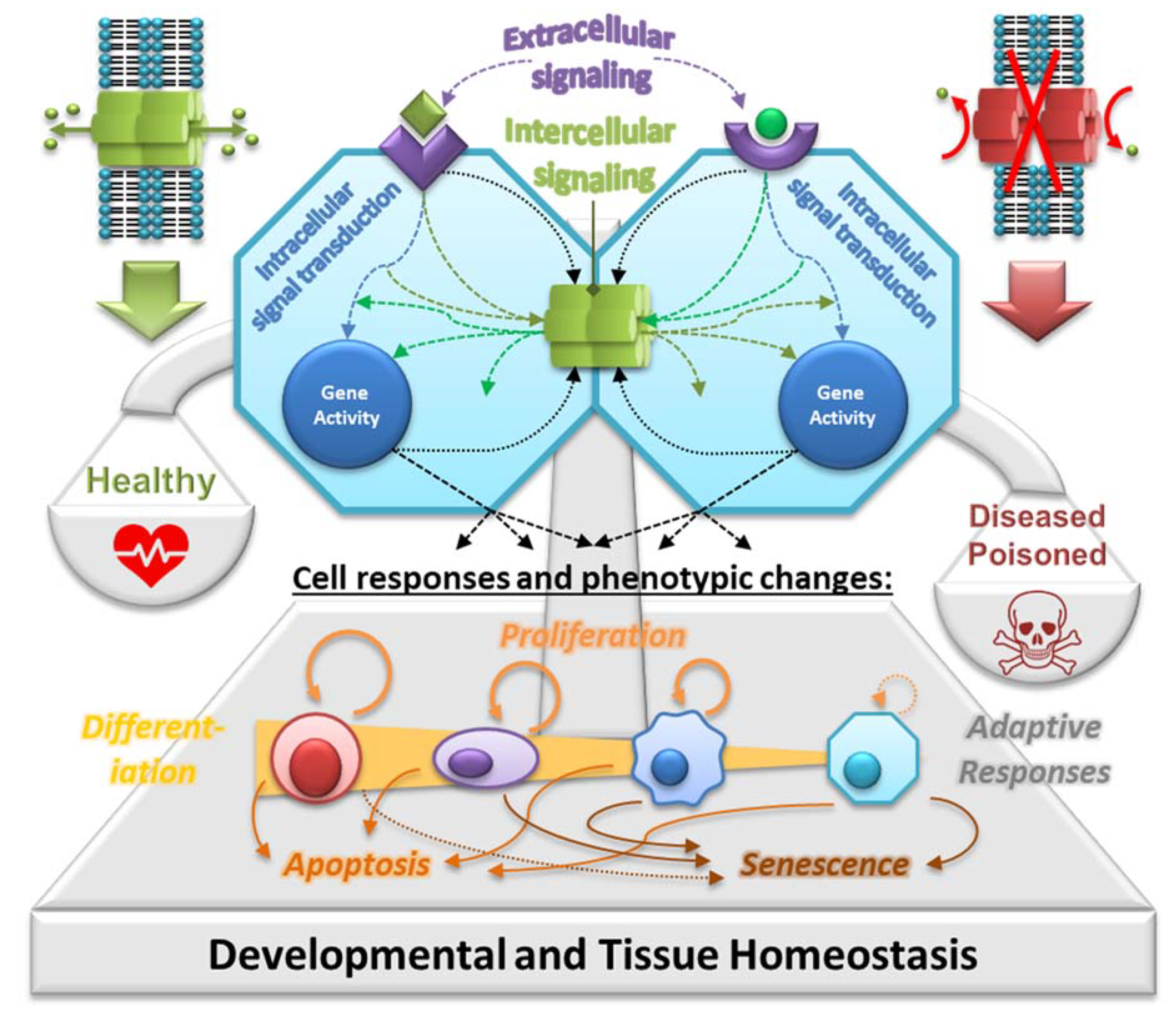{kind=link}
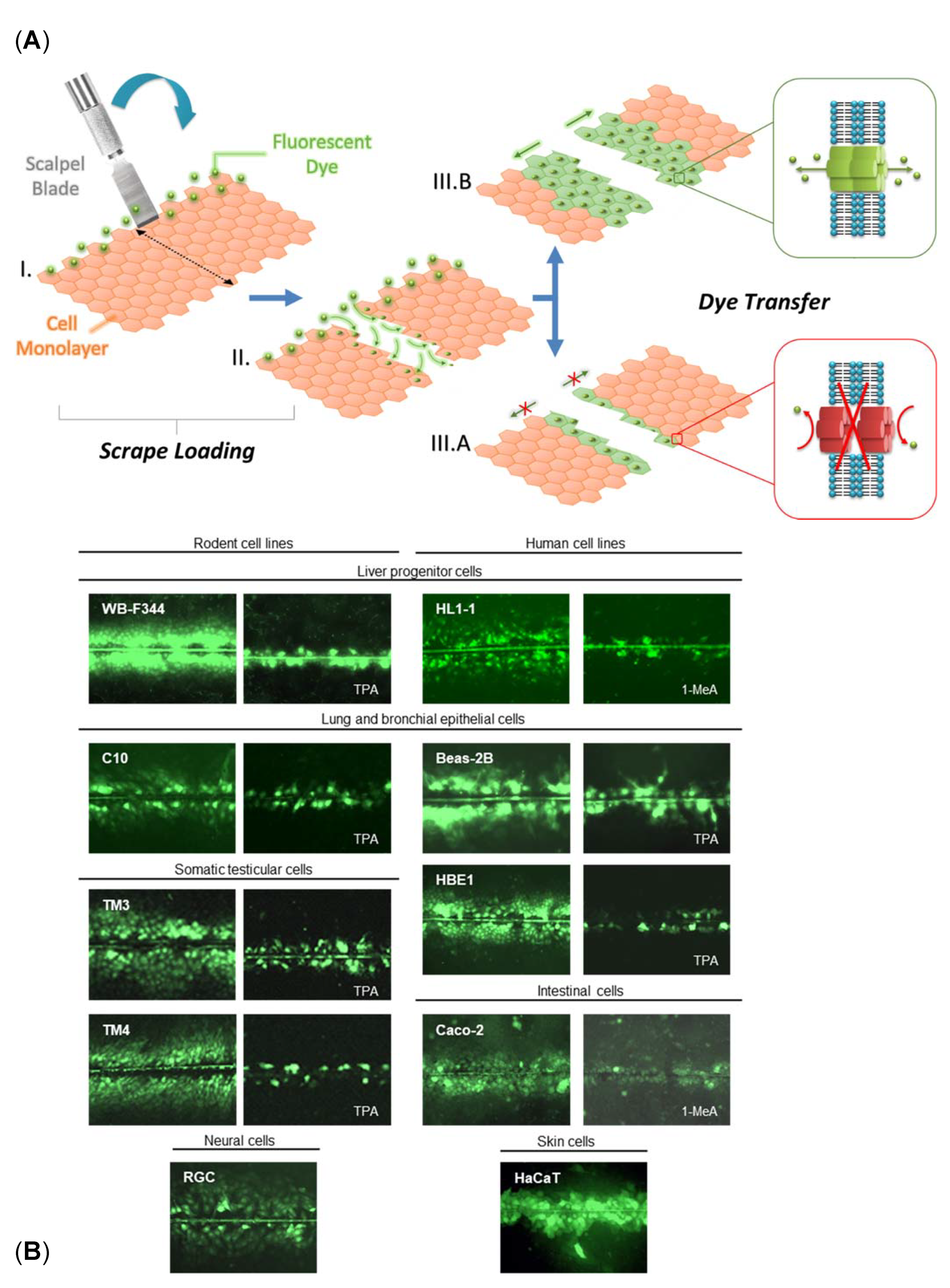{kind=link}
| Organ/Cell Line | Species | Major Connexins (Method) | GJIC (Method) | Ref. |
|---|---|---|---|---|
| Brain: | ||||
| BT5C1 | R | Y (MI) | [100] | |
| RG2 | R | Cx43 (WB) | Y (SL) | [101] |
| RGC | R | Y (SL) | [27] | |
| Ear: | ||||
| HEI-OC1 | M | Cx26, Cx30, Cx31, Cx43 (WB, IF, RT-qPCR) | Y (SL) | [102] |
| Eye: | ||||
| RGC-5 | R/M | Y (SL) | [103] | |
| Intestine: | ||||
| Caco-2 | H | Cx43, Cx26 (RT-PCR, WB, IF) | Y (SL) | [27,104,105,106] |
| IEC-6 | R | Cx43 (RT-PCR, WB) | Y (MI) | [107] |
| Kidney: | ||||
| BHK 21/13 | GH | Y (MC, MI) | [108] | |
| G401.2/6TG.1 | H | Y | [109] | |
| MDCK | D | Cx43 (WB) | Y (SL) | [110] |
| Liver: | ||||
| ARL-18 | R | Cx43 (WB, IF) | Y (FR) | [111] |
| BRL | R | Y (MI, MC) | [100,108] | |
| BRL 3A | R | Cx43, Cx32 (RT-(q)PCR, IF, WB) | Y (SL, Par) | [112,113,114,115,116,117,118,119,120] |
| Chang Liver | H | Y (SL) | [121] | |
| Clone 9 | R | Cx43, Cx26 (WB, IF, RT-qPCR) | Y (SL, MI, FR) | [122,123,124,125,126,127,128,129,130,131] |
| G27 | R | Y (MI) | [132,133,134] | |
| HepG2 | H | Cx43, Cx32 (RT-PCR) | Y (MI, Par, SL) | [121,135,136,137] |
| HL1-1 | H | Y (SL) | [27] | |
| HLEC-04 | H | Y (MI) | [138] | |
| Huh-7 | H | Y (SL) | [121] | |
| IAR-20 | R | Cx43 (RT-PCR, WB, IF) | Y (SL, MI, Par, MC) | [139,140,141,142,143,144,145,146,147,148,149,150,151,152] |
| IAR-203 | R | Cx43 (WB, IF, RT-PCR) | Y (MI) | [153,154] |
| IAR-6.1 | R | Cx43 (WB) | Y (SL) | [81,141,143,144] |
| N1S1-67 | R | Cx43 (WB, NB) | Y (Pre) | [155] |
| REL | R | Cx43 (WB, IF) | Y (MI) | [156,157,158,159] |
| T51B | R | Cx43 (WB, NB) | Y (MI) | [160,161,162,163,164] |
| WB-F344 | R | Cx43 (WB, IF, RT-(q)PCR) | Y (SL, MI, Par, DWCPC, MC, FR, Pre) | [39,51,75,78,79,89,90,103,165,166,167,168,169,170,171,172,173,174,175,176,177,178,179,180,181,182,183,184,185,186,187,188,189,190,191,192,193,194,195,196,197,198,199,200,201,202,203,204,205,206,207,208,209,210,211,212,213,214,215,216,217,218,219,220,221,222,223,224,225,226,227,228,229,230,231,232,233,234,235,236,237,238,239,240,241,242,243,244,245,246,247,248,249,250,251,252,253,254,255,256] |
| Lung: | ||||
| 16HBE14o- | H | Cx43 (IF) | [257] | |
| A549 | H | Cx43 (IF) | [258] | |
| Beas-2B | H | Y (SL) | [27,259] | |
| C10 | M | Cx43 (IF, WB) | Y (MI) | [27,260] |
| HBE1 | H | Cx43 (WB) | Y (SL) | [259,261] |
| V79 | CH | Y (MC, MI) | [108,236,262,263,264,265,266,267] | |
| Mammary gland: | ||||
| BICR/M1Rk | R | Cx43 (WB, RT-PCR) | Y (MI, SL) | [100,268] |
| Pancreas: | ||||
| H6c7 | H | Cx43 (WB, IF, RTPCR) | Y (SL) | [269] |
| Placenta: | ||||
| FL | H | Y (MI) | [100] | |
| Prostate: | ||||
| RWPE-1 | H | Cx43, Cx32 (WB) | Y (SL, FR) | [270,271] |
| Skin: | ||||
| 3PC | M | Y (MI) | [136] | |
| 3T3 | M | Y (MI, MC) | [108,272] | |
| CA3/7 | M | Y (MI) | [136] | |
| HaCaT | H | Cx43, Cx26 (RT-PCR, WB, IF) | Y (FR, MI, SL) | [273,274,275] |
| HEL37 | M | Y (MI) | [100] | |
| Testes: | ||||
| 42GPA9 | R | Cx43 (WB, RT-PCR, IF) | Y (FR) | [276] |
| LC540 | R | Y (FR, SL) | [236] | |
| SerW3 | R | Cx43 (RT-qPCR, IF) | Y (MI) | [277] |
| TM3 | M | Cx43, Cx45 (WB, RT-PCR, IF) | Y (SL) | [77,278] |
| TM4 | M | Cx43, Cx45 (WB, RT-PCR) | Y (SL, MI) | [77,279] |
4. Scrape Loading-Dye Transfer Using the WB-F344 Cell Line
5. Chemical Effects on GJIC in WB-F344 Cells Evaluated by SL-DT Assay—A Systematic Search
5.1. Summary of the Results
5.2. Reproducibility of the Assay
5.3. Comparison of SL-DT Results with Data on Genotoxicity and Carcinogenicity
5.3.1. Genotoxicity Data
5.3.2. IARC Carcinogenicity
5.3.3. ComTox/ToxRefDB Data
5.3.4. OncoLogic System
5.3.5. Other Assays for In Vitro GJIC Assessment
6. Conclusions and Future Perspective
7. Literature Search and Statistical Measures
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rosenkranz, H.S.; Pollack, N.; Cunningham, A.R. Exploring the relationship between the inhibition of gap junctional intercellular communication and other biological phenomena. Carcinogenesis 2000, 21, 1007–1011. [Google Scholar] [CrossRef] [PubMed]
- Mulkearns-Hubert, E.E.; Reizes, O.; Lathia, J.D. Connexins in cancer: Jekyll or Hyde? Biomolecules 2020, 10, 1654. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, M.N.; Colacci, A.; Corvi, R.; Vaccari, M.; Aguila, M.C.; Corvaro, M.; Delrue, N.; Desaulniers, D.; Ertych, N.; Jacobs, A.; et al. Chemical carcinogen safety testing: OECD expert group international consensus on the development of an integrated approach for the testing and assessment of chemical non-genotoxic carcinogens. Arch. Toxicol. 2020, 1, 3. [Google Scholar] [CrossRef] [PubMed]
- WHO. Cancer. Available online: https://www.who.int/health-topics/cancer#tab=tab_1 (accessed on 10 April 2021).
- Madia, F.; Worth, A.; Whelan, M.; Corvi, R. Carcinogenicity assessment: Addressing the challenges of cancer and chemicals in the environment. Environ. Int. 2019, 128, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Luijten, M.; Corvi, R.; Mehta, J.; Corvaro, M.; Delrue, N.; Felter, S.; Haas, B.; Hewitt, N.J.; Hilton, G.; Holmes, T.; et al. A comprehensive view on mechanistic approaches for cancer risk assessment of non-genotoxic agrochemicals. Regul. Toxicol. Pharmacol. 2020, 118, 104789. [Google Scholar] [CrossRef]
- Jacobs, M.N.; Colacci, A.; Louekari, K.; Luijten, M.; Hakkert, B.C.; Paparella, M.; Vasseur, P. International regulatory needs for development of an IATA for non-genotoxic carcinogenic chemical substances. ALTEX 2016, 33, 359–392. [Google Scholar] [CrossRef] [Green Version]
- Abel, E.L.; DiGiovanni, J.; Mendelsohn, J.; Howley, P.; Israel, M.; Gray, J.; Thompson, C. Environmental carcinogenesis. In The Molecular Basis of Cancer, 4th ed.; Mendelsohn, J., Howley, P., Israel, M., Gray, J., Thompson, C., Eds.; W.B. Saunders: Philadelphia, PA, USA, 2015; pp. 103–128.e2. ISBN 978-1-4557-4066-6. [Google Scholar]
- Basu, A.K. DNA damage, mutagenesis and cancer. Int. J. Mol. Sci. 2018, 19, 970. [Google Scholar] [CrossRef] [Green Version]
- Luijten, M.; Olthof, E.D.; Hakkert, B.C.; Rorije, E.; van der Laan, J.-W.; Woutersen, R.A.; van Benthem, J. An integrative test strategy for cancer hazard identification. Crit. Rev. Toxicol. 2016, 46, 615–639. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Fouad, Y.A.; Aanei, C. Revisiting the hallmarks of cancer. Am. J. Cancer Res. 2017, 7, 1016–1036. [Google Scholar]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Trosko, J.E.; Chang, C.-C.; Upham, B.L.; Tai, M.-H. Ignored hallmarks of carcinogenesis: Stem cells and cell-cell communication. Ann. N. Y. Acad. Sci. 2004, 1028, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.J.; Perfetti, T.A.; Hayes, A.W.; Berry, S.C.; Trosko, J.E.; King, J.A.; Goodman, J.I.; Begley, C.G.; Dayan, A. Categorizing the characteristics of human carcinogens: A need for specificity. Arch. Toxicol. 2021, 95. [Google Scholar] [CrossRef] [PubMed]
- Baba, A.; Câtoi, C. Carcinogenesis. In Comparative Oncology; The Publishing House of the Romanian Academy: Bucharest, Romania, 2007. [Google Scholar]
- Bevan, R.J.; Harrison, P.T.C. Threshold and non-threshold chemical carcinogens: A survey of the present regulatory landscape. Regul. Toxicol. Pharmacol. 2017, 88, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.J.; Perfetti, T.A.; King, J.A. Rodent 2-year cancer bioassays and in vitro and in vivo genotoxicity tests insufficiently predict risk or model development of human carcinomas. Toxicol. Res. Appl. 2019, 3, 2397847319849648. [Google Scholar] [CrossRef] [Green Version]
- Goodman, J.I. Goodbye to the bioassay. Toxicol. Res. 2018, 7, 558–564. [Google Scholar] [CrossRef] [Green Version]
- Cohen, S.M. Evaluation of possible carcinogenic risk to humans based on liver tumors in rodent assays: The two-year bioassay is no longer necessary. Toxicol. Pathol. 2010, 38, 487–501. [Google Scholar] [CrossRef]
- Paparella, M.; Colacci, A.; Jacobs, M.N. Uncertainties of testing methods: What do we (want to) know about carcinogenicity? ALTEX 2017, 34, 235–252. [Google Scholar] [CrossRef] [Green Version]
- OECD. Overview of the Set of OECD Genetic Toxicology Test Guidelines and Updates Performed in 2014–2015. 2017. Available online: https://www.oecd.org/officialdocuments/publicdisplaydocumentpdf/?cote=ENV-JM-MONO(2016)33/rev1&doclanguage=en (accessed on 16 June 2021).
- Nielsen, M.S.; Nygaard Axelsen, L.; Sorgen, P.L.; Verma, V.; Delmar, M.; Holstein-Rathlou, N.-H. Gap junctions. Compr. Physiol. 2012, 2, 1981–2035. [Google Scholar]
- Zefferino, R.; Piccoli, C.; Di Gioia, S.; Capitanio, N.; Conese, M. Gap junction intercellular communication in the carcinogenesis hallmarks: Is this a phenomenon or epiphenomenon? Cells 2019, 8, 896. [Google Scholar] [CrossRef] [Green Version]
- Aasen, T.; Mesnil, M.; Naus, C.C.; Lampe, P.D.; Laird, D.W. Gap junctions and cancer: Communicating for 50 years. Nat. Rev. Cancer 2016, 16, 775–788. [Google Scholar] [CrossRef] [Green Version]
- Axelsen, L.N.; Calloe, K.; Holstein-Rathlou, N.-H.; Nielsen, M.S. Managing the complexity of communication: Regulation of gap junctions by post-translational modification. Front. Pharmacol. 2013, 4, 130. [Google Scholar] [CrossRef] [Green Version]
- Babica, P. Gap Junctional Intercellular Communication: In Vitro Assessment of Hazardous and Beneficial Effects of Chemical. Docent Habilitation Thesis, Masaryk University, Brno, Czech Republic, 2017. Available online: https://is.muni.cz/do/rect/habilitace/1431/12437/habilitacni_prace/Babica_2017_FINAL_TEXT_.pdf (accessed on 16 June 2021).
- Trosko, J.E. The gap junction as a “Biological Rosetta Stone”: Implications of evolution, stem cells to homeostatic regulation of health and disease in the Barker hypothesis. J. Cell Commun. Signal. 2011, 5, 53–66. [Google Scholar] [CrossRef] [Green Version]
- Trosko, J.E. Evolution of microbial quorum sensing to human global quorum sensing: An insight into how gap junctional intercellular communication might be linked to the global metabolic disease crisis. Biology 2016, 5, 29. [Google Scholar] [CrossRef]
- Trosko, J.E. Gap junctional intercellular communication as a biological “Rosetta stone” in understanding, in a systems biological manner, stem cell behavior, mechanisms of epigenetic toxicology, chemoprevention and chemotherapy. J. Membr. Biol. 2007, 218, 93–100. [Google Scholar] [CrossRef]
- Upham, B.L.; Trosko, J.E. Oxidative-dependent integration of signal transduction with intercellular gap junctional communication in the control of gene expression. Antioxid. Redox Signal. 2009, 11, 297–307. [Google Scholar] [CrossRef] [Green Version]
- Upham, B.L. Role of integrative signaling through gap junctions in toxicology. Curr. Protoc. Toxicol. 2011, 2, 2.18. [Google Scholar] [CrossRef]
- Upham, B.L.; Sovadinova, I.; Babica, P. Gap junctional intercellular communication: A functional biomarker to assess adverse effects of toxicants and toxins, and health benefits of natural products. J. Vis. Exp. 2016, 2016, e54281. [Google Scholar] [CrossRef]
- Mesnil, M.; Defamie, N.; Naus, C.; Sarrouilhe, D. Brain disorders and chemical pollutants: A gap junction link? Biomolecules 2020, 11, 51. [Google Scholar] [CrossRef]
- Aasen, T.; Leithe, E.; Graham, S.V.; Kameritsch, P.; Mayán, M.D.; Mesnil, M.; Pogoda, K.; Tabernero, A. Connexins in cancer: Bridging the gap to the clinic. Oncogene 2019, 38, 4429–4451. [Google Scholar] [CrossRef] [Green Version]
- Trosko, J.E. Reflections on the use of 10 IARC carcinogenic characteristics for an objective approach to identifying and organizing results from certain mechanistic studies. Toxicol. Res. Appl. 2017, 1, 2397847317710837. [Google Scholar] [CrossRef]
- Czyż, J.; Piwowarczyk, K.; Paw, M.; Luty, M.; Wróbel, T.; Catapano, J.; Madeja, Z.; Ryszawy, D. Connexin-dependent intercellular stress signaling in tissue homeostasis and tumor development. Acta Biochim. Pol. 2017, 64, 377–389. [Google Scholar] [CrossRef]
- Jiang, J.X.; Penuela, S. Connexin and pannexin channels in cancer. BMC Cell Biol. 2016, 17 (Suppl. 1), 12. [Google Scholar] [CrossRef] [Green Version]
- Loch-Caruso, R.; Galvez, M.M.; Brant, K.; Chung, D. Cell and toxicant specific phosphorylation of conexin43: Effects of lindane and TPA on rat myometrial and WB-F344 liver cell gap junctions. Cell Biol. Toxicol. 2004, 20, 147–169. [Google Scholar] [CrossRef]
- Wong, P.; Laxton, V.; Srivastava, S.; Fiona Chan, Y.W.; Tse, G. The role of gap junctions in inflammatory and neoplastic disorders (Review). Int. J. Mol. Med. 2017, 39, 498–506. [Google Scholar] [CrossRef] [Green Version]
- Nalewajska, M.; Marchelek-Myśliwiec, M.; Opara-Bajerowicz, M.; Dziedziejko, V.; Pawlik, A. Connexins-therapeutic targets in cancers. Int. J. Mol. Sci. 2020, 21, 9119. [Google Scholar] [CrossRef]
- Trosko, J.E. On the potential origin and characteristics of cancer stem cells. Carcinogenesis 2021, 42, 905–912. [Google Scholar] [CrossRef]
- Leone, A.; Longo, C.; Trosko, J.E. The chemopreventive role of dietary phytochemicals through gap junctional intercellular communication. Phytochem. Rev. 2012, 11, 285–307. [Google Scholar] [CrossRef]
- Mesnil, M.; Crespin, S.; Avanzo, J.L.; Zaidan-Dagli, M.L. Defective gap junctional intercellular communication in the carcinogenic process. Biochim. Biophys. Acta-Biomembr. 2005, 1719, 125–145. [Google Scholar] [CrossRef] [Green Version]
- Vinken, M.; Henkens, T.; Vanhaecke, T.; Papeleu, P.; Geerts, A.; Van Rossen, E.; Chipman, J.K.; Meda, P.; Rogiers, V. Trichostatin a enhances gap junctional intercellular communication in primary cultures of adult rat hepatocytes. Toxicol. Sci. 2006, 91, 484–492. [Google Scholar] [CrossRef] [Green Version]
- Mantz, J.; Cordier, J.; Giaume, C. Effects of general anesthetics on intercellular communications mediated by gap junctions between astrocytes in primary culture. Anesthesiology 1993, 78, 892–901. [Google Scholar] [CrossRef]
- Vera, B.; Sánchez-Abarca, L.I.; Bolaños, J.P.; Medina, J.M. Inhibition of astrocyte gap junctional communication by ATP depletion is reversed by calcium sequestration. FEBS Lett. 1996, 392, 225–228. [Google Scholar] [CrossRef] [Green Version]
- Blomstrand, F.; Aberg, N.D.; Eriksson, P.S.; Hansson, E.; Rönnbäck, L. Extent of intercellular calcium wave propagation is related to gap junction permeability and level of connexin-43 expression in astrocytes in primary cultures from four brain regions. Neuroscience 1999, 92, 255–265. [Google Scholar] [CrossRef]
- Köster-Patzlaff, C.; Hosseini, S.M.; Reuss, B. Persistent Borna disease virus infection changes expression and function of astroglial gap junctions in vivo and in vitro. Brain Res. 2007, 1184, 316–332. [Google Scholar] [CrossRef]
- Morioka, N.; Suekama, K.; Zhang, F.F.; Kajitani, N.; Hisaoka-Nakashima, K.; Takebayashi, M.; Nakata, Y. Amitriptyline up-regulates connexin43-gap junction in rat cultured cortical astrocytes via activation of the p38 and c-Fos/AP-1 signalling pathway. Br. J. Pharmacol. 2014, 171, 2854–2867. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, S.; Seidelin, M.; Bisgaard, H.C.; Vang, O. Indolo[3,2-b]carbazole inhibits gap junctional intercellular communication in rat primary hepatocytes and acts as a potential tumor promoter. Carcinogenesis 2002, 23, 1861–1868. [Google Scholar] [CrossRef] [Green Version]
- Leaphart, C.L.; Qureshi, F.; Cetin, S.; Li, J.; Dubowski, T.; Batey, C.; Beer–Stolz, D.; Guo, F.; Murray, S.A.; Hackam, D.J. Interferon-γ inhibits intestinal restitution by preventing gap junction communication between enterocytes. Gastroenterology 2007, 132, 2395–2411. [Google Scholar] [CrossRef]
- Fukumoto, M.; Kujiraoka, T.; Hara, M.; Shibasaki, T.; Hosoya, T.; Yoshida, M. Effect of cadmium on gap junctional intercellular communication in primary cultures of rat renal proximal tubular cells. Life Sci. 2001, 69, 247–254. [Google Scholar] [CrossRef]
- Yoshida, M.; Kujiraoka, T.; Hara, M.; Nakazawa, H.; Sumi, Y. Methylmercury inhibits gap junctional intercellular communication in primary cultures of rat proximal tubular cells. Arch. Toxicol. 1998, 72, 192–196. [Google Scholar] [CrossRef]
- Hills, C.E.; Price, G.W.; Wall, M.J.; Kaufmann, T.J.; Tang, C.-W.; Yiu, W.H.; Squires, P.E. Transforming growth factor beta 1 drives a switch in connexin mediated cell-to-cell communication in tubular cells of the diabetic kidney. Cell. Physiol. Biochem. 2018, 45, 2369–2388. [Google Scholar] [CrossRef] [Green Version]
- Yaoita, E.; Yao, J.; Yoshida, Y.; Morioka, T.; Nameta, M.; Takata, T.; Kamiie, J.; Fujinaka, H.; Oite, T.; Yamamoto, T. Up-regulation of connexin43 in glomerular podocytes in response to injury. Am. J. Pathol. 2002, 161, 1597–1606. [Google Scholar] [CrossRef] [Green Version]
- Avanzo, J.L.; Mennecier, G.; Mesnil, M.; Hernandez-Blazquez, F.J.; Fukumasu, H.; da Silva, T.C.; Rao, K.V.K.; Dagli, M.L.Z. Deletion of a single allele of Cx43 is associated with a reduction in the gap junctional intercellular communication and increased cell proliferation of mouse lung pneumocytes type II. Cell Prolif. 2007, 40, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Trovato-Salinaro, A.; Trovato-Salinaro, E.; Failla, M.; Mastruzzo, C.; Tomaselli, V.; Gili, E.; Crimi, N.; Condorelli, F.D.; Vancheri, C. Altered intercellular communication in lung fibroblast cultures from patients with idiopathic pulmonary fibrosis. Respir. Res. 2006, 7, 122. [Google Scholar] [CrossRef] [Green Version]
- Mensink, A.; de Haan, L.H.; Lakemond, C.M.; Koelman, C.A.; Koeman, J.H. Inhibition of gap junctional intercellular communication between primary human smooth muscle cells by tumor necrosis factor alpha. Carcinogenesis 1995, 16, 2063–2067. [Google Scholar] [CrossRef]
- Dookwah, H.D.; Barhoumi, R.; Narasimhan, T.R.; Safe, S.H.; Burghardt, R.C. Gap junctions in myometrial cell cultures: Evidence for modulation by cyclic adenosine 3′:5′-monophosphate. Biol. Reprod. 1992, 47, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Loch-Caruso, R.; Upham, B.L.; Harris, C.; Trosko, J.E. Divergent roles for glutathione in lindane-induced acute and delayed-onset inhibition of rat myometrial gap junctions. Toxicol. Sci. 2005, 85, 694–702. [Google Scholar] [CrossRef] [Green Version]
- Changchien, C.-Y.; Sung, M.-H.; Chang, H.-H.; Tsai, W.-C.; Peng, Y.-S.; Chen, Y. Uremic toxin indoxyl sulfate suppresses myocardial Cx43 assembly and expression via JNK activation. Chem. Biol. Interact. 2020, 319, 108979. [Google Scholar] [CrossRef]
- Hervé, J.C.; Pluciennik, F.; Verrecchia, F.; Bastide, B.; Delage, B.; Joffre, M.; Délèze, J. Influence of the molecular structure of steroids on their ability to interrupt gap junctional communication. J. Membr. Biol. 1996, 149, 179–187. [Google Scholar] [CrossRef]
- Gingrich, J.; Pu, Y.; Upham, B.L.; Hulse, M.; Pearl, S.; Martin, D.; Avery, A.; Veiga-Lopez, A. Bisphenol S enhances gap junction intercellular communication in ovarian theca cells. Chemosphere 2021, 263, 128304. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Chang, H.-M.; Cheng, J.-C.; Tsai, H.-D.; Wu, C.-H.; Leung, P.C.K. Transforming growth factor-β1 up-regulates connexin43 expression in human granulosa cells. Hum. Reprod. 2015, 30, 2190–2201. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.-C.; Wang, K.-H.; Chuang, K.-H.; Kao, A.-P.; Kuo, T.-C. Downregulation of gap junctional intercellular communication and connexin 43 expression by bisphenol A in human granulosa cells. Biotechnol. Appl. Biochem. 2020, 68, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Durlej, M.; Kopera, I.; Knapczyk-Stwora, K.; Hejmej, A.; Duda, M.; Koziorowski, M.; Slomczynska, M.; Bilinska, B. Connexin 43 gene expression in male and female gonads of porcine offspring following in utero exposure to an anti-androgen, flutamide. Acta Histochem. 2011, 113, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Cillo, F.; de Eguileor, M.; Gandolfi, F.; Brevini, T.A.L. Aroclor-1254 affects mRNA polyadenylation, translational activation, cell morphology, and DNA integrity of rat primary prostate cells. Endocr. Relat. Cancer 2007, 14, 257–266. [Google Scholar] [CrossRef]
- Herve, J.C.; Pluciennik, F.; Bastide, B.; Cronier, L.; Verrecchia, F.; Malassine, A.; Joffre, M.; Deleze, J. Contraceptive gossypol blocks cell-to-cell communication in human and rat cells. Eur. J. Pharmacol. 1996, 313, 243–255. [Google Scholar] [CrossRef]
- You, S.; Li, W.; Lin, T. Expression and regulation of connexin43 in rat Leydig cells. J. Endocrinol. 2000, 166, 447–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.-L.; Wang, H.; Wang, Z.-R.; Zhang, Y.-F.; Chen, Y.-Q.; Zhu, F.-H.; Zhang, Y.-Q.; Ma, J.; Li, Z. TGF-β1 regulation of estrogen production in mature rat Leydig cells. PLoS ONE 2013, 8, e60197. [Google Scholar] [CrossRef] [Green Version]
- Pamies, D.; Hartung, T. 21st century cell culture for 21st century toxicology. Chem. Res. Toxicol. 2017, 30, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Bai, X.; Liu, Y.; Wang, K.; Shen, B.; Sun, X. Current concepts and perspectives on connexin43: A mini review. Curr. Protein Pept. Sci. 2018, 19, 1049–1057. [Google Scholar] [CrossRef]
- Maes, M.; Crespo Yanguas, S.; Willebrords, J.; Cogliati, B.; Vinken, M. Connexin and pannexin signaling in gastrointestinal and liver disease. Transl. Res. 2015, 166, 332–343. [Google Scholar] [CrossRef] [Green Version]
- Horvath, A.; Upham, B.L.; Ganev, V.; Trosko, J.E. Determination of the epigenetic effects of ochratoxin in a human kidney and a rat liver epithelial cell line. Toxicon 2002, 40, 273–282. [Google Scholar] [CrossRef]
- Osgood, R.S.; Upham, B.L.; Bushel, P.R.; Velmurugan, K.; Xiong, K.-N.; Bauer, A.K. Secondhand smoke-prevalent polycyclic aromatic hydrocarbon binary mixture-induced specific mitogenic and pro-inflammatory cell signaling events in lung epithelial cells. Toxicol. Sci. 2017, 157, 156–171. [Google Scholar] [CrossRef] [Green Version]
- Kubincová, P.; Sychrová, E.; Raška, J.; Basu, A.; Yawer, A.; Dydowiczová, A.; Babica, P.; Sovadinová, I. Polycyclic aromatic hydrocarbons and endocrine disruption: Role of testicular gap junctional intercellular communication and connexins. Toxicol. Sci. 2019, 169, 70–83. [Google Scholar] [CrossRef]
- Sovadinova, I.; Babica, P.; Boke, H.; Kumar, E.; Wilke, A.; Park, J.-S.; Trosko, J.E.; Upham, B.L. Phosphatidylcholine specific PLC-induced dysregulation of gap junctions, a robust cellular response to environmental toxicants, and prevention by resveratrol in a rat liver cell model. PLoS ONE 2015, 10, e0124454. [Google Scholar] [CrossRef] [Green Version]
- Upham, B.L.; Rummel, A.M.; Carbone, J.M.; Trosko, J.E.; Ouyang, Y.; Crawford, R.B.; Kaminski, N.E. Cannabinoids inhibit gap junctional intercellular communication and activate ERK in a rat liver epithelial cell line. Int. J. Cancer 2003, 104, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.Z.; Ao, P.; Boynton, A.L. Platelet-derived growth factor-induced disruption of gap junctional communication and phosphorylation of connexin43 involves protein kinase C and mitogen-activated protein kinase. J. Cell. Physiol. 1998, 176, 332–341. [Google Scholar] [CrossRef]
- Rivedal, E.; Opsahl, H. Role of PKC and MAP kinase in EGF- and TPA-induced connexin43 phosphorylation and inhibition of gap junction intercellular communication in rat liver epithelial cells. Carcinogenesis 2001, 22, 1543–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruciani, V.; Mikalsen, S.O. Connexins, gap junctional intercellular communication and kinases. Biol. Cell 2002, 94, 433–443. [Google Scholar] [CrossRef] [Green Version]
- Lampe, P.D.; Lau, A.F. The effects of connexin phosphorylation on gap junctional communication. Int. J. Biochem. Cell Biol. 2004, 36, 1171–1186. [Google Scholar] [CrossRef] [Green Version]
- Albright, C.D.; Grimley, P.M.; Jones, R.T.; Fontana, J.A.; Keenan, K.P.; Resau, J.H. Cell-to-cell communication: A differential response to TGF-beta in normal and transformed (BEAS-2B) human bronchial epithelial cells. Carcinogenesis 1991, 12, 1993–1999. [Google Scholar] [CrossRef] [PubMed]
- Budunova, I.V.; Mittelman, L.A.; Miloszewska, J. Role of protein kinase C in the regulation of gap junctional communication. Teratog. Carcinog. Mutagen. 1994, 14, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Criswell, K.A.; Lochcaruso, R.; Stuenkel, E.L. Lindane inhibition of gap junctional communication in myometrial myocytes is partially dependent on phosphoinositide-generated second messengers. Toxicol. Appl. Pharmacol. 1995, 130, 280–293. [Google Scholar] [CrossRef]
- Husøy, T.; Cruciani, V.; Sanner, T.; Mikalsen, S.O. Phosphorylation of connexin43 and inhibition of gap junctional communication in 12-O-tetradecanoylphorbol-13-acetate-exposed R6 fibroblasts: Minor role of protein kinase C beta I and mu. Carcinogenesis 2001, 22, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Koo, S.K.; Kim, D.Y.; Park, S.D.; Kang, K.W.; Joe, C.O. PKC phosphorylation disrupts gap junctional communication at G0/S phase in clone 9 cells. Mol. Cell. Biochem. 1997, 167, 41–49. [Google Scholar] [CrossRef]
- Upham, B.L.; Blaha, L.; Babica, P.; Park, J.-S.; Sovadinova, I.; Pudrith, C.; Rummel, A.M.; Weis, L.M.; Sai, K.; Tithof, P.K.; et al. Tumor promoting properties of a cigarette smoke prevalent polycyclic aromatic hydrocarbon as indicated by the inhibition of gap junctional intercellular communication via phosphatidylcholine-specific phospholipase C. Cancer Sci. 2008, 99, 696–705. [Google Scholar] [CrossRef] [Green Version]
- Machala, M.; Bláha, L.; Vondráček, J.; Trosko, J.E.; Scott, J.; Upham, B.L. Inhibition of gap junctional intercellular communication by noncoplanar polychlorinated biphenyls: Inhibitory potencies and screening for potential mode(s) of action. Toxicol. Sci. 2003, 76, 102–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.C.; Trosko, J.E.; Kung, H.J.; Bombick, D.; Matsumura, F. Potential role of the src gene product in inhibition of gap-junctional communication in NIH/3T3 cells. Proc. Natl. Acad. Sci. USA 1985, 82, 5360–5364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottrell, G.T.; Lin, R.; Warn-Cramer, B.J.; Lau, A.F.; Burt, J.M. Mechanism of v-Src- and mitogen-activated protein kinase-induced reduction of gap junction communication. Am. J. Physiol. Cell Physiol. 2003, 284, C511–C520. [Google Scholar] [CrossRef]
- Filson, A.J.; Azarnia, R.; Beyer, E.C.; Loewenstein, W.R.; Brugge, J.S. Tyrosine phosphorylation of a gap junction protein correlates with inhibition of cell-to-cell communication. Cell Growth Differ. Mol. Biol. J. Am. Assoc. Cancer Res. 1990, 1, 661–668. [Google Scholar]
- Lin, R.; Warn-Cramer, B.J.; Kurata, W.E.; Lau, A.F. v-Src phosphorylation of connexin 43 on Tyr247 and Tyr265 disrupts gap junctional communication. J. Cell Biol. 2001, 154, 815–827. [Google Scholar] [CrossRef]
- Pahujaa, M.; Anikin, M.; Goldberg, G.S. Phosphorylation of connexin43 induced by Src: Regulation of gap junctional communication between transformed cells. Exp. Cell Res. 2007, 313, 4083–4090. [Google Scholar] [CrossRef]
- Ghoshal, S.; Weber, W.J.; Rummel, A.M.; Trosko, J.E.; Upham, B.L. Epigenetic toxicity of a mixture of polycyclic aromatic hydrocarbons on gap junctional intercellular communication before and after biodegradation. Environ. Sci. Technol. 1999, 33, 1044–1050. [Google Scholar] [CrossRef]
- Abbaci, M.; Barberi-Heyob, M.; Blondel, W.; Guillemin, F.; Didelon, J. Advantages and limitations of commonly used methods to assay the molecular permeability of gap junctional intercellular communication. Biotechniques 2008, 45, 33–62. [Google Scholar] [CrossRef]
- Maes, M.; Yanguas, S.C.; Willebrords, J.; Vinken, M. Models and methods for in vitro testing of hepatic gap junctional communication. Toxicol. In Vitro 2015, 30, 569–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyyathurai, J.; Himpens, B.; Bultynck, G.; D’hondt, C. Calcium wave propagation triggered by local mechanical stimulation as a method for studying gap junctions and hemichannels. In Gap Junction Protocols; Methods in Molecular Biology; Vinken, M., Johnstone, S.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; Volume 1437, pp. 203–211. ISBN 978-1-4939-3664-9. [Google Scholar]
- Brümmer, F.; Zempel, G.; Bühle, P.; Stein, J.C.; Hülser, D.F. Retinoic acid modulates gap junctional permeability: A comparative study of dye spreading and ionic coupling in cultured cells. Exp. Cell Res. 1991, 196, 158–163. [Google Scholar] [CrossRef] [Green Version]
- Sidorova, T.S.; Matesic, D.F. Protective effect of the natural product, chaetoglobosin K, on lindane- and dieldrin-induced changes in astroglia: Identification of activated signaling pathways. Pharm. Res. 2008, 25, 1297–1308. [Google Scholar] [CrossRef]
- Choi, S.J.; Kim, S.W.; Lee, J.B.; Lim, H.J.; Kim, Y.J.; Tian, C.; So, H.S.; Park, R.; Choung, Y.-H. Gingko biloba extracts protect auditory hair cells from cisplatin-induced ototoxicity by inhibiting perturbation of gap junctional intercellular communication. Neuroscience 2013, 244, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Pulukuri, S.; Sitaramayya, A. Retinaldehyde, a potent inhibitor of gap junctional intercellular communication. Cell Commun. Adhes. 2004, 11, 25–33. [Google Scholar] [CrossRef]
- Traoré, A.; Baudrimont, I.; Dano, S.; Sanni, A.; Larondelle, Y.; Schneider, Y.J.; Creppy, E.E.; Traore, A.; Baudrimont, I.; Dano, S.; et al. Epigenetic properties of the diarrhetic marine toxin okadaic acid: Inhibition of the gap junctional intercellular communication in a human intestine epithelial cell line. Arch. Toxicol. 2003, 77, 657–662. [Google Scholar] [CrossRef]
- Creppy, E.E.; Traore, A.; Baudrimont, I.; Cascante, M.; Carratu, M.R. Recent advances in the study of epigenetic effects induced by the phycotoxin okadaic acid. Toxicology 2002, 181, 433–439. [Google Scholar] [CrossRef]
- Morita, H.; Katsuno, T.; Hoshimoto, A.; Hirano, N.; Saito, Y.; Suzuki, Y. Connexin 26-mediated gap junctional intercellular communication suppresses paracellular permeability of human intestinal epithelial cell monolayers. Exp. Cell Res. 2004, 298, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ey, B.; Eyking, A.; Gerken, G.; Podolsky, D.K.; Cario, E. TLR2 mediates gap junctional intercellular communication through connexin-43 in intestinal epithelial barrier injury. J. Biol. Chem. 2009, 284, 22332–22343. [Google Scholar] [CrossRef] [Green Version]
- Pitts, J.D.; Hamilton, A.E.; Kam, E.; Burk, R.R.; Murphy, J.P. Retinoic acid inhibits junctional communication between animal cells. Carcinogenesis 1986, 7, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Ayashi, T.; Hasler, C.; Oh, S.Y.; Madhukar, B.; Chang, C.C.; Trosko, J.; Hayashi, T. A human kidney epithelial cell culture as an in vitro model to study chemical modulation of intercellular communication. In Vitro Mol. Toxicol. J. Basic Appl. Res. 1996, 9, 3–17. [Google Scholar]
- Mally, A.; Decker, M.; Bekteshi, M.; Dekant, W. Ochratoxin A alters cell adhesion and gap junction intercellular communication in MDCK cells. Toxicology 2006, 223, 15–25. [Google Scholar] [CrossRef]
- Budunova, I.V.; Williams, G.M.; Spray, D.C. Effect of tumor-promoting stimuli on gap junction permeability and connexin43 expression in Arl-18 rat liver cell line. Arch. Toxicol. 1993, 67, 565–572. [Google Scholar] [CrossRef]
- Hu, D.; Zou, H.; Han, T.; Xie, J.; Dai, N.; Zhuo, L.; Gu, J.; Bian, J.; Yuan, Y.; Liu, X.; et al. Gap junction blockage promotes cadmium-induced apoptosis in BRL 3A derived from Buffalo rat liver cells. J. Vet. Sci. 2016, 17, 63–70. [Google Scholar] [CrossRef]
- Zou, H.; Liu, X.; Han, T.; Hu, D.; Yuan, Y.; Gu, J.; Bian, J.; Liu, Z. Alpha-lipoic acid protects against cadmium-induced hepatotoxicity via calcium signalling and gap junctional intercellular communication in rat hepatocytes. J. Toxicol. Sci. 2015, 40, 469–477. [Google Scholar] [CrossRef] [Green Version]
- Zou, H.; Hu, D.; Han, T.; Zhao, H.; Xie, J.; Liu, X.; Wang, Y.; Gu, J.; Yuan, Y.; Bian, J.; et al. Salidroside ameliorates Cd-induced calcium overload and gap junction dysfunction in BRL 3A rat liver cells. Biol. Trace Elem. Res. 2015, 164, 90–98. [Google Scholar] [CrossRef]
- Zou, H.; Liu, X.; Han, T.; Hu, D.; Wang, Y.; Yuan, Y.; Gu, J.; Bian, J.; Zhu, J.; Liu, Z.-P.Z. Salidroside protects against cadmium-induced hepatotoxicity in rats via GJIC and MAPK pathways. PLoS ONE 2015, 10, e0129788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Lin, L.; Luan, T.; Chan Gilbert, Y.S.; Lan, C. Effects of organophosphorus pesticides and their ozonation byproducts on gap junctional intercellular communication in rat liver cell line. Food Chem. Toxicol. 2007, 45, 2057–2063. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Cai, Z.; Chen, H.; Cao, L.; Chen, B.; Lin, B. Involvement of gap junctions in propylthiouracil-induced cytotoxicity in BRL-3A cells. Exp. Ther. Med. 2019, 17, 2799–2806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, F.; Li, S.; Gan, X.; Wang, R.; Chen, Z. Propofol inhibits gap junctions by attenuating sevoflurane-induced cytotoxicity against rat liver cells in vitro. Eur. J. Anaesthesiol. 2014, 31, 219–224. [Google Scholar] [CrossRef]
- Wang, R.; Huang, F.; Chen, Z.; Li, S. Downregulation of connexin 32 attenuates hypoxia/reoxygenation injury in liver cells. J. Biochem. Mol. Toxicol. 2015, 29, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Zhuo, L.; Han, T.; Hu, D.; Yang, X.; Wang, Y.; Yuan, Y.; Gu, J.; Bian, J.; Liu, X.; et al. Autophagy and gap junctional intercellular communication inhibition are involved in cadmium-induced apoptosis in rat liver cells. Biochem. Biophys. Res. Commun. 2015, 459, 713–719. [Google Scholar] [CrossRef]
- Carruba, G.; Cocciadiferro, L.; Bellavia, V.; Rizzo, S.; Tsatsanis, C.; Spandidos, D.; Muti, P.; Smith, C.; Mehta, P.; Castagnetta, L. Intercellular communication and human hepatocellular carcinoma. Ann. N. Y. Acad. Sci. 2004, 1028, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Benane, S.G.; Nelson, G.B.; Ross, J.A.; Blackman, C.F. Benzo{[}a]pyrene and dibenzo{[}a,l]pyrene do not alter gap junction communication in rat liver epithelial cells. Polycycl. Aromat. Compd. 1999, 17, 53–62. [Google Scholar] [CrossRef]
- Thompson, D.C.; Barhoumi, R.; Burghardt, R.C. Comparative toxicity of eugenol and its quinone methide metabolite in cultured liver cells using kinetic fluorescence bioassays. Toxicol. Appl. Pharmacol. 1998, 149, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Meola, S.; Meola, R.; Barhoumi, R.; Miles, J.M.; Burghardt, R.C. Laser cytometric analysis of permethrin toxicity in insect and mammalian epithelial cells. Toxic Subst. Mech. 1997, 16, 237–249. [Google Scholar]
- Zhang, J.; Grindstaff, R.D.; Thai, S.-F.; Murray, S.A.; Kohan, M.; Blackman, C.F. Chloral hydrate decreases gap junction communication in rat liver epithelial cells. Cell Biol. Toxicol. 2011, 27, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, V.M.; Rook, M.B.; Traub, O.; Hertzberg, E.L.; Sáez, J.C. On the mechanisms of cell uncoupling induced by a tumor promoter phorbol ester in clone 9 cells, a rat liver epithelial cell line. Eur. J. Cell Biol. 1993, 62, 384–396. [Google Scholar]
- Na, M.R.; Koo, S.K.; Kim, D.Y.; Park, S.D.; Rhee, S.K.; Kang, K.W.; Joe, C.O. In vitro inhibition of gap junctional intercellular communication by chemical carcinogens. Toxicology 1995, 98, 199–206. [Google Scholar] [CrossRef]
- Griffin, G.D.; Williams, M.W.; Gailey, P.C. Cellular communication in clone 9 cells exposed to magnetic fields. Radiat. Res. 2000, 153, 690–698. [Google Scholar] [CrossRef]
- Barhoumi, R.; Mouneimne, Y.; Ramos, K.S.; Safe, S.H.; Phillips, T.D.; Centonze, V.E.; Ainley, C.; Gupta, M.S.; Burghardt, R.C. Analysis of benzo{[}a]pyrene partitioning and cellular homeostasis in a rat liver cell line. Toxicol. Sci. 2000, 53, 264–270. [Google Scholar] [CrossRef] [Green Version]
- Ottinger, S.; Barhoumi, R.; McKenzie, K.S.; McDonald, T.; Burgardt, R.; Huebner, H.J.; Phillips, T.D.; Burghardt, R.; Huebner, H.J.; Phillips, T.D.; et al. FIA/MS analysis of temporally ozonated benzo{[}a]pyrene and pyrene and their reaction products: Inhibition of gap junction-mediated intercellular communication. Chemosphere 2005, 60, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, R.W.; Barhoumi, R.; Miles, J.M.; Burghardt, R.C. Attenuation of gossypol cytotoxicity by cyclic AMP in a rat liver cell line. Toxicol. Appl. Pharmacol. 1998, 151, 311–318. [Google Scholar] [CrossRef]
- Sharovskaja, J.J.; Vaiman, A.V.; Solomatina, N.A.; Kobliakov, V.A. Inhibition of gap junction intercellular communications in cell culture by polycyclic aromatic hydrocarbons (PAH) in the absence of PAH metabolism. Biochemistry 2004, 69, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Sharovskaya, J.; Kobliakova, I.; Solomatina, N.; Kobliakov, V. Effect of some carcinogenic and non-carcinogenic polycyclic aromatic hydrocarbons on gap junction intercellular communication in hepatoma cell cultures. Eur. J. Cell Biol. 2006, 85, 387–397. [Google Scholar] [CrossRef]
- Sharovskaya, Y.Y.; Vajman, A.V.; Solomatina, N.A.; Kobliakov, V.A. Inhibition of gap junction intercellular communications in cell culture by polycyclic aromatic hydrocarbons (PAH) in the absence of PAH metabolism. Biokhimiya 2004, 69, 511–518. [Google Scholar]
- Yang, Y.; Qin, S.-K.; Wu, Q.; Wang, Z.-S.; Zheng, R.-S.; Tong, X.-H.; Liu, H.; Tao, L.; He, X.-D. Connexin-dependent gap junction enhancement is involved in the synergistic effect of sorafenib and all-trans retinoic acid on HCC growth inhibition. Oncol. Rep. 2014, 31, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Jansen, L.A.M.; Jongen, W.M.F. The use of initiated cells as a test system for the detection of inhibitors of gap junctional intercellular communication. Carcinogenesis 1996, 17, 333–339. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Yang, S.; Yu, W.; Chen, Q.; Shen, J.; Hu, Y.; Shi, J.; Wu, X.; Li, J.; Li, N. Effects of continuous renal replacement therapy on intestinal mucosal barrier function during extracorporeal membrane oxygenation in a porcine model. J. Cardiothorac. Surg. 2014, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Kamendulis, L.M.; Isenberg, J.S.; Smith, J.H.; Pugh, G.J.; Lington, A.W.; Klaunig, J.E. Comparative effects of phthalate monoesters on gap junctional intercellular communication and peroxisome proliferation in rodent and primate hepatocytes. J. Toxicol. Environ. Health. A 2002, 65, 569–588. [Google Scholar] [CrossRef]
- Dukic, A.R.; McClymont, D.W.; Tasken, K. Cell-based high-throughput assay for gap junction communication suitable for assessing connexin 43-ezrin interaction disruptors using IncuCyte ZOOM. SLAS Discov. Adv. Life Sci. R D 2017, 22, 77–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenne, K.; Fransson-Steen, R.; Honkasalo, S.; Wärngård, L. Two inhibitors of gap junctional intercellular communication, TPA and endosulfan: Different effects on phosphorylation of connexin 43 in the rat liver epithelial cell line, IAR 20. Carcinogenesis 1994, 15, 1161–1165. [Google Scholar] [CrossRef] [PubMed]
- Mesnil, M.; Montesano, R.; Yamasaki, H. Intercellular communication of transformed and non-transformed rat liver epithelial cells: Modulation by TPA. Exp. Cell Res. 1986, 165, 391–402. [Google Scholar] [CrossRef]
- Leithet, E.; Rivedal, E. Ubiquitination and down-regulation of gap junction protein connexin-43 in response to 12-O-tetradecanoylphorbol 13-acetate treatment. J. Biol. Chem. 2004, 279, 50089–50096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivedal, E.; Leithe, E. Connexin43 synthesis, phosphorylation, and degradation in regulation of transient inhibition of gap junction intercellular communication by the phorbol ester TPA in rat liver epithelial cells. Exp. Cell Res. 2005, 302, 143–152. [Google Scholar] [CrossRef]
- Rivedal, E.; Witz, G. Metabolites of benzene are potent inhibitors of gap-junction intercellular communication. Arch. Toxicol. 2005, 79, 303–311. [Google Scholar] [CrossRef]
- Kato, Y.; Kenne, K.; Haraguchi, K.; Masuda, Y.; Kimura, R.; Wärngård, L. Inhibition of cell-cell communication by methylsulfonyl metabolites of polychlorinated biphenyl congeners in rat liver epithelial IAR 20 cells. Arch. Toxicol. 1998, 72, 178–182. [Google Scholar] [CrossRef]
- Kato, Y.; Kenne, K. Inhibition of cell-cell communication by commercial chlorinated paraffins in rat liver epithelial IAR 20 cells. Pharmacol. Toxicol. 1996, 79, 23–28. [Google Scholar] [CrossRef]
- Hori, T.; Asamoto, M.; Krutovskikh, V.; Iwahori, Y.; Maeda, M.; ToriyamaBaba, H.; Takasuka, N.; Tsuda, H. Triazine derivatives inhibit rat hepatocarcinogenesis but do not enhance gap junctional intercellular communication. Jpn. J. Cancer Res. 1997, 88, 12–17. [Google Scholar] [CrossRef]
- Bager, Y.; Lindebro, M.C.; Martel, P.; Chaumontet, C.; Warngard, L. Altered function, localization and phosphorylation of gap junctions in rat liver epithelial, IAR 20, cells after treatment with PCBs or TCDD. Environ. Toxicol. Pharmacol. 1997, 3, 257–266. [Google Scholar] [CrossRef]
- Leithe, E.; Cruciani, V.; Sanner, T.; Mikalsen, S.-O.; Rivedal, E. Recovery of gap junctional intercellular communication after phorbol ester treatment requires proteasomal degradation of protein kinase C. Carcinogenesis 2003, 24, 1239–1245. [Google Scholar] [CrossRef] [Green Version]
- Leithe, E.; Rivedal, E. Epidermal growth factor regulates ubiquitination, internalization and proteasome-dependent degradation of connexin43. J. Cell Sci. 2004, 117, 1211–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirnes, S.; Leithe, E.; Rivedal, E. The detergent resistance of Connexin43 is lost upon TPA or EGF treatment and is an early step in gap junction endocytosis. Biochem. Biophys. Res. Commun. 2008, 373, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Mercier, T.; Honikman-Leban, E.; Chaumontet, C.; Martel, P.; Shahin, M.M. Studies on the modulating effects of retinoic acid and retinol acetate using dye transfer and metabolic cooperation assays. Fundam. Appl. Toxicol. 1993, 21, 270–276. [Google Scholar] [CrossRef]
- Chaumontet, C.; Bex, V.; Véran, F.; Martel, P. The vitamin E analog tocopherol succinate strongly inhibits gap junctional intercellular communication in rat liver epithelial cells (IAR203). J. Nutr. Biochem. 2008, 19, 263–268. [Google Scholar] [CrossRef]
- Bex, V.; Mercier, T.; Chaumontet, C.; Gaillard-Sanchez, I.; Flechon, B.; Mazet, F.; Traub, O.; Martel, P. Retinoic acid enhances connexin43 expression at the post-transcriptional level in rat liver epithelial cells. Cell Biochem. Funct. 1995, 13, 69–77. [Google Scholar] [CrossRef]
- Lampe, P.D. Analyzing phorbol ester effects on gap junctional communication: A dramatic inhibition of assembly. J. Cell Biol. 1994, 127, 1895–1905. [Google Scholar] [CrossRef] [Green Version]
- Apostoli, P.; Huard, C.; Chaumontet, C.; Martel, P.; Alessio, L.; Mazzoleni, G. Effects of four inorganic lead compounds on the proliferation and junctional coupling of cultured REL liver cells. Am. J. Ind. Med. 2000, 38, 340–348. [Google Scholar] [CrossRef]
- Huard, C.; Druesne, N.; Guyonnet, D.; Thomas, M.; Pagniez, A.; Le Bon, A.-M.; Martel, P.; Chaumontet, C. Diallyl disulfide (DADS) enhances gap-junctional intercellular communication by both direct and indirect mechanisms in rat liver cells. Carcinogenesis 2004, 25, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Chaumontet, C.; Droumaguet, C.; Bex, V.; Heberden, C.; Gaillard-Sanchez, I.; Martel, P. Flavonoids (apigenin, tangeretin) counteract tumor promoter-induced inhibition of intercellular communication of rat liver epithelial cells. Cancer Lett. 1997, 114, 207–210. [Google Scholar] [CrossRef]
- Chaumontet, C.; Suschetet, M.; Honikman-Leban, E.; Krutovskikh, V.A.; Berges, R.; Le Bon, A.M.; Heberden, C.; Shahin, M.M.; Yamasaki, H.; Martel, P. Lack of tumor-promoting effects of flavonoids: Studies on rat liver preneoplastic foci and on in vivo and in vitro gap junctional intercellular communication. Nutr. Cancer 1996, 26, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.P.; Peng, A.; Hossain, M.Z.; Fan, Y.; Jagdale, A.; Boynton, A.L. Tumor promotion by hydrogen peroxide in rat liver epithelial cells. Carcinogenesis 1999, 20, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Lau, A.F.; Kanemitsu, M.Y.; Kurata, W.E.; Danesh, S.; Boynton, A.L. Epidermal growth factor disrupts gap-junctional communication and induces phosphorylation of connexin43 on serine. Mol. Biol. Cell 1992, 3, 865–874. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.P.; Peng, A.; Golard, A.; Hossain, M.Z.; Huang, R.; Liu, Y.G.; Boynton, A.L. Hydrogen peroxide promotes transformation of rat liver non-neoplastic epithelial cells through activation of epidermal growth factor receptor. Mol. Carcinog. 2001, 30, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.Z.; Jagdale, A.B.; Ao, P.; Boynton, A.L. Mitogen-activated protein kinase and phosphorylation of connexin43 are not sufficient for the disruption of gap junctional communication by platelet-derived growth factor and tetradecanoylphorbol acetate. J. Cell. Physiol. 1999, 179, 87–96. [Google Scholar] [CrossRef]
- Kanemitsu, M.Y.; Lau, A.F. Epidermal growth factor stimulates the disruption of gap junctional communication and connexin43 phosphorylation independent of 12-0- tetradecanoylphorbol 13-acetate-sensitive protein kinase C: The possible involvement of mitogen-activated protein kinase. Mol. Biol. Cell 1993, 4, 837–848. [Google Scholar] [CrossRef]
- Kang, N.J.; Lee, K.M.; Kim, J.H.; Lee, B.K.; Kwon, J.Y.; Lee, K.W.; Lee, H.J. Inhibition of gap junctional intercellular communication by the green tea polyphenol (-)-epigallocatechin gallate in normal rat liver epithelial cells. J. Agric. Food Chem. 2008, 56, 10422–10427. [Google Scholar] [CrossRef]
- Upham, B.L.; Masten, S.J.; Lockwood, B.R.; Trosko, J.E. Nongenotoxic effects of polycyclic aromatic hydrocarbons and their oxygenation by-products on the intercellular communication of rat liver epithelial cells. Fundam. Appl. Toxicol. 1994, 23, 470–475. [Google Scholar] [CrossRef]
- Kang, K.S.; Kang, B.C.; Lee, B.J.; Che, J.H.; Li, G.X.; Trosko, J.E.; Lee, Y.S. Preventive effect of epicatechin and ginsenoside Rb2 on the inhibition of gap junctional intercellular communication by TPA and H2O2. Cancer Lett. 2000, 152, 97–106. [Google Scholar] [CrossRef]
- Abdelmohsen, K.; Stuhlmann, D.; Daubrawa, F.; Klotz, L.-O. Dicumarol is a potent reversible inhibitor of gap junctional intercellular communication. Arch. Biochem. Biophys. 2005, 434, 241–247. [Google Scholar] [CrossRef]
- Fransson, R.; Nicotera, P.; Wärngård, L.; Ahlborg, U.G. Changes in cytosolic CA2+ are not involved in DDT-induced loss of gap junctional communication in WB-F344 cells. Cell Biol. Toxicol. 1990, 6, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Hii, C.S.; Ferrante, A.; Schmidt, S.; Rathjen, D.A.; Robinson, B.S.; Poulos, A.; Murray, A.W. Inhibition of gap junctional communication by polyunsaturated fatty acids in WB cells: Evidence that connexin 43 is not hyperphosphorylated. Carcinogenesis 1995, 16, 1505–1511. [Google Scholar] [CrossRef]
- Lee, K.M.; Kwon, J.Y.; Lee, K.W.; Lee, H.J. Ascorbic acid 6-palmitate suppresses gap-junctional intercellular communication through phosphorylation of connexin 43 via activation of the MEK-ERK pathway. Mutat. Res. Mol. Mech. Mutagen. 2009, 660, 51–56. [Google Scholar] [CrossRef]
- Upham, B.L.; Deocampo, N.D.; Wurl, B.; Trosko, J.E. Inhibition of gap junctional intercellular communication by perfluorinated fatty acids is dependent on the chain length of the fluorinated tail. Int. J. Cancer 1998, 78, 491–495. [Google Scholar] [CrossRef]
- Hakulinen, P.; Rintala, E.; Mäki-Paakkanen, J.; Komulainen, H. Altered expression of connexin43 in the inhibition of gap junctional intercellular communication by chlorohydroxyfuranones in WB-F344 rat liver epithelial cells. Toxicol. Appl. Pharmacol. 2006, 212, 146–155. [Google Scholar] [CrossRef]
- Flodström, S.; Warngård, L.; Hemming, H.; Ahlborg, U.G. Chlorobenzilate-induced effects on enzyme-altered foci in rat liver and intercellular communication in rat liver WB-F344 epithelial cells. Cancer Lett. 1988, 43, 161–166. [Google Scholar] [CrossRef]
- Sai, K.; Upham, B.L.; Kang, K.S.; Hasegawa, R.; Inoue, T.; Trosko, J.E. Inhibitory effect of pentachlorophenol on gap junctional intercellular communication in rat liver epithelial cells in vitro. Cancer Lett. 1998, 130, 9–17. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, N.J.; Lee, B.K.; Lee, K.W.; Lee, H.J. Gallic acid, a metabolite of the antioxidant propyl gallate, inhibits gap junctional intercellular communication via phosphorylation of connexin 43 and extracellular-signal-regulated kinase1/2 in rat liver epithelial cells. Mutat. Res. 2008, 638, 175–183. [Google Scholar] [CrossRef]
- Forejtníková, H.; Lunerová, K.; Kubínová, R.; Jankovská, D.; Marek, R.; Kares, R.; Suchý, V.; Vondrácek, J.; Machala, M. Chemoprotective and toxic potentials of synthetic and natural chalcones and dihydrochalcones in vitro. Toxicology 2005, 208, 81–93. [Google Scholar] [CrossRef]
- Nishikawa, A.; Sai, K.; Okazaki, K.; Son, H.-Y.; Kanki, K.; Nakajima, M.; Kinae, N.; Nohmi, T.; Trosko, J.E.; Inoue, T.; et al. MX, a by-product of water chlorination, lacks in vivo genotoxicity in gpt delta mice but inhibits gap junctional intercellular communication in rat WB cells. Environ. Mol. Mutagen. 2006, 47, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, P.-J.; Jao, J.-C.; Tsai, J.-L.; Chang, W.-T.; Jeng, K.-S.; Kuo, K.-K. Inorganic arsenic trioxide induces gap junction loss in association with the downregulation of connexin43 and E-cadherin in rat hepatic “stem-like” cells. Kaohsiung J. Med. Sci. 2014, 30, 57–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, C.S.; Oh, S.Y.; Schmidt, S.A.; Clark, K.J.; Murray, A.W. Lysophosphatidic acid inhibits gap-junctional communication and stimulates phosphorylation of connexin-43 in WB cells: Possible involvement of the mitogen-activated protein kinase cascade. Biochem. J. 1994, 303 Pt 2, 475–479. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.H.; Cho, M.H.; Cho, J.H. Effects of cadmium on gap junctional intercellular communication in WB-F344 rat liver epithelial cells. Hum. Exp. Toxicol. 2001, 20, 577–583. [Google Scholar] [CrossRef]
- Masten, S.J.; Tian, M.; Upham, B.L.; Trosko, J.E. Effect of selected pesticides and their ozonation by-products on gap junctional intercellular communication using rat liver epithelial cell lines. Chemosphere 2001, 44, 457–465. [Google Scholar] [CrossRef]
- Alpatova, A.L.; Shan, W.; Babica, P.; Upham, B.L.; Rogensues, A.R.; Masten, S.J.; Drown, E.; Mohanty, A.K.; Alocilja, E.C.; Tarabara, V.V. Single-walled carbon nanotubes dispersed in aqueous media via non-covalent functionalization: Effect of dispersant on the stability, cytotoxicity, and epigenetic toxicity of nanotube suspensions. Water Res. 2010, 44, 505–520. [Google Scholar] [CrossRef]
- Upham, B.L.; Guzvić, M.; Scott, J.; Carbone, J.M.; Blaha, L.; Coe, C.; Li, L.L.; Rummel, A.M.; Trosko, J.E. Inhibition of gap junctional intercellular communication and activation of mitogen-activated protein kinase by tumor-promoting organic peroxides and protection by resveratrol. Nutr. Cancer 2007, 57, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Flodström, S.; Hemming, H.; Wärngård, L.; Ahlborg, U.G. Promotion of altered hepatic foci development in rat liver, cytochrome P450 enzyme induction and inhibition of cell-cell communication by DDT and some structurally related organohalogen pesticides. Carcinogenesis 1990, 11, 1413–1417. [Google Scholar] [CrossRef]
- Babica, P.; Čtveráčková, L.; Lenčešová, Z.; Trosko, J.E.; Upham, B.L. Chemopreventive agents attenuate rapid inhibition of gap junctional intercellular communication induced by environmental toxicants. Nutr. Cancer 2016, 68, 827–837. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.; Ruch, R.J.; Vang, O. Resveratrol reverses tumor-promoter-induced inhibition of gap-junctional intercellular communication. Biochem. Biophys. Res. Commun. 2000, 275, 804–809. [Google Scholar] [CrossRef]
- Matesic, D.F.; Abifadel, D.N.; Garcia, E.L.; Jann, M.W. Effect of thioridazine on gap junction intercellular communication in connexin 43-expressing cells. Cell Biol. Toxicol. 2006, 22, 257–268. [Google Scholar] [CrossRef]
- Lee, C.-H.; Chen, I.-H.; Lee, C.-R.; Chi, C.-H.; Tsai, M.-C.; Tsai, J.-L.; Lin, H.-F. Inhibition of gap junctional Intercellular communication in WB-F344 rat liver epithelial cells by triphenyltin chloride through MAPK and PI3-kinase pathways. J. Occup. Med. Toxicol. 2010, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Hasler, C.M.; Frick, M.A.; Bennink, M.R.; Trosko, J.E. TPA-induced inhibition of gap junctional intercellular communication is not mediated through free radicals. Toxicol. Appl. Pharmacol. 1990, 103, 389–398. [Google Scholar] [CrossRef]
- Lee, B.K.; Chung, M.-Y.; Lee, K.W. Benzo[a]pyrene-7,8-diol-9,10-epoxide inhibits gap junction intercellular communication via phosphorylation of tumor progression locus 2 in WB-F344 rat liver epithelial cells. Mol. Carcinog. 2015, 54, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Upham, B.L.; Weis, L.M.; Rummel, A.M.; Masten, S.J.; Trosko, J.E. The effects of anthracene and methylated anthracenes on gap junctional intercellular communication in rat liver epithelial cells. Fundam. Appl. Toxicol. 1996, 34, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Upham, B.L.; Weis, L.M.; Trosko, J.E. Modulated gap junctional intercellular communication as a biomarker of PAH epigenetic toxicity: Structure-function relationship. Environ. Health Perspect. 1998, 106 (Suppl. 4), 975–981. [Google Scholar] [CrossRef]
- Weis, L.M.; Rummel, A.M.; Masten, S.J.; Trosko, J.E.; Upham, B.L. Bay or baylike regions of polycyclic aromatic hydrocarbons were potent inhibitors of gap junctional intercellular communication. Environ. Health Perspect. 1998, 106, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Rummel, A.M.; Trosko, J.E.; Wilson, M.R.; Upham, B.L. Polycyclic aromatic hydrocarbons with bay-like regions inhibited gap junctional intercellular communication and stimulated MAPK activity. Toxicol. Sci. 1999, 49, 232–240. [Google Scholar] [CrossRef] [Green Version]
- Bláha, L.; Kapplová, P.; Vondráček, J.; Upham, B.; Machala, M. Inhibition of gap-junctional intercellular communication by environmentally occuring polycyclic aromatic hydrocarbons. Toxicol. Sci. 2002, 65, 43–51. [Google Scholar] [CrossRef]
- Marvanová, S.; Vondrácek, J.; Penccíková, K.; Trilecová, L.; Krcmárr, P.; Topinka, J.; Nováková, Z.; Milcová, A.; Machala, M. Toxic effects of methylated benz[a]anthracenes in liver cells. Chem. Res. Toxicol. 2008, 21, 503–512. [Google Scholar] [CrossRef]
- Svihálková-Sindlerová, L.; Machala, M.; Pencíková, K.; Marvanová, S.; Neca, J.; Topinka, J.; Sevastyanova, O.; Kozubík, A.; Vondrácek, J.; Švihálková-Šindlerová, L.; et al. Dibenzanthracenes and benzochrysenes elicit both genotoxic and nongenotoxic events in rat liver “stem-like” cells. Toxicology 2007, 232, 147–159. [Google Scholar] [CrossRef]
- Kabátková, M.; Svobodová, J.; Pěnčíková, K.; Mohatad, D.S.; Šmerdová, L.; Kozubík, A.; Machala, M.; Vondráček, J.; Kabatkova, M.; Svobodova, J.; et al. Interactive effects of inflammatory cytokine and abundant low-molecular-weight PAHs on inhibition of gap junctional intercellular communication, disruption of cell proliferation control, and the AhR-dependent transcription. Toxicol. Lett. 2015, 232, 113–121. [Google Scholar] [CrossRef]
- Novakova, K.; Blaha, L.; Babica, P. Tumor promoting effects of cyanobacterial extracts are potentiated by anthropogenic contaminants-Evidence from in vitro study. Chemosphere 2012, 89, 30–37. [Google Scholar] [CrossRef]
- Luster-Teasley, S.L.; Ganey, P.E.; DiOrio, M.; Ward, J.S., 3rd; Maleczka, R.E.J.; Trosko, J.E.; Masten, S.J. Effect of byproducts from the ozonation of pyrene: Biphenyl-2,2′,6,6′-tetracarbaldehyde and biphenyl-2,2′,6,6′-tetracarboxylic acid on gap junction intercellular communication and neutrophil function. Environ. Toxicol. Chem. 2005, 24, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Herner, H.A.; Trosko, J.E.; Masten, S.J. The epigenetic toxicity of pyrene and related ozonation byproducts containing an aldehyde functional group. Environ. Sci. Technol. 2001, 35, 3576–3583. [Google Scholar] [CrossRef] [PubMed]
- Machala, M.; Svihálková-Sindlerová, L.; Pencíková, K.; Krcmár, P.; Topinka, J.; Milcová, A.; Nováková, Z.; Kozubík, A.; Vondrácek, J. Effects of methylated chrysenes on AhR-dependent and -independent toxic events in rat liver epithelial cells. Toxicology 2008, 247, 93–101. [Google Scholar] [CrossRef]
- Vondrácek, J.; Chramostová, K.; Plísková, M.; Bláha, L.; Brack, W.; Kozubík, A.; Machala, M.; Vondracek, J.; Chramostova, K.; Pliskova, M.; et al. Induction of aryl hydrocarbon receptor-mediated and estrogen receptor-mediated activities, and modulation of cell proliferation by dinaphthofurans. Environ. Toxicol. Chem. 2004, 23, 2214–2220. [Google Scholar] [CrossRef] [Green Version]
- Vondráček, J.; Švihálková-Šindlerová, L.; Pěnčíková, K.; Krčmář, P.; Andrysík, Z.; Chramostová, K.; Marvanová, S.; Valovičová, Z.; Kozubík, A.; Gábelová, A.; et al. 7H-Dibenzo[c,g]carbazole and 5,9-dimethyldibenzo[c,g]carbazole exert multiple toxic events contributing to tumor promotion in rat liver epithelial “stem-like” cells. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 2006, 596, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.G.; Trosko, J.E. Concentration/response effect of 2,2′, 4,4′, 5,5′-hexabromobiphenyl on cell-cell communication in vitro: Assessment by fluorescence redistribution after photobleaching (“FRAP”). Cell Biol. Toxicol. 1988, 4, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Hamers, T.; Kamstra, J.H.; Cenijn, P.H.; Pencikova, K.; Palkova, L.; Simeckova, P.; Vondracek, J.; Andersson, P.L.; Stenberg, M.; Machala, M. In vitro toxicity profiling of ultrapure non-dioxin-like polychlorinated biphenyl congeners and their relative toxic contribution to PCB mixtures in humans. Toxicol. Sci. 2011, 121, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Simecková, P.; Vondrácek, J.; Andrysík, Z.; Zatloukalová, J.; Krcmár, P.; Kozubík, A.; Machala, M. The 2,2′,4,4′,5,5′-hexachlorobiphenyl-enhanced degradation of connexin 43 involves both proteasomal and lysosomal activities. Toxicol. Sci. 2009, 107, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Machala, M.; Bláha, L.; Lehmler, H.-J.; Plísková, M.; Májková, Z.; Kapplová, P.; Sovadinová, I.; Vondrácek, J.; Malmberg, T.; Robertson, L.W. Toxicity of hydroxylated and quinoid PCB metabolites: Inhibition of gap junctional intercellular communication and activation of aryl hydrocarbon and estrogen receptors in hepatic and mammary cells. Chem. Res. Toxicol. 2004, 17, 340–347. [Google Scholar] [CrossRef]
- Satoh, A.Y.; Trosko, J.E.; Masten, S.J. Epigenetic toxicity of hydroxylated biphenyls and hydroxylated polychlorinated biphenyls on normal rat liver epithelial cells. Environ. Sci. Technol. 2003, 37, 2727–2733. [Google Scholar] [CrossRef]
- Andrysik, Z.; Prochazkova, J.; Kabatkova, M.; Umannova, L.; Simeckova, P.; Kohoutek, J.; Kozubik, A.; Machala, M.; Vondracek, J.; Andrysík, Z.; et al. Aryl hydrocarbon receptor-mediated disruption of contact inhibition is associated with connexin43 downregulation and inhibition of gap junctional intercellular communication. Arch. Toxicol. 2013, 87, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Upham, B.L.; Trosko, J.E. A paradigm shift in the understanding of oxidative stress and its implications to exposure of low-level ionizing radiation. Acta Med. Nagasaki. 2005, 50, 63–68. [Google Scholar]
- Oh, S.-Y.; Schmidt, S.A.; Murray, A.W. Epidermal growth factor inhibits gap junctional communication and stimulates serine-phosphorylation of connexin43 in WB cells by a protein kinase c-independent mechanism. Cell Commun. Adhes. 1993, 1, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.-H.; Cho, S.-D.; Hu, H.; Kim, S.-H.; Lee, S.K.; Lee, Y.-S.; Kang, K.-S. The roles of ERK1/2 and p38 MAP kinases in the preventive mechanisms of mushroom Phellinus linteus against the inhibition of gap junctional intercellular communication by hydrogen peroxide. Carcinogenesis 2002, 23, 1163–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upham, B.L.; Kang, K.S.; Cho, H.Y.; Trosko, J.E. Hydrogen peroxide inhibits gap junctional intercellular communication in glutathione sufficient but not glutathione deficient cells. Carcinogenesis 1997, 18, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-J.; Seo, S.G.; Choi, K.; Kim, J.E.; Kang, H.; Chung, M.-Y.; Lee, K.W.; Lee, H.J. Recovery effect of onion peel extract against H2O2-induced inhibition of gap-junctional intercellular communication is mediated through quercetin. J. Food Sci. 2014, 79, H1011–H1017. [Google Scholar] [CrossRef]
- Kim, J.-S.; Ha, T.-Y.; Ahn, J.; Kim, H.-K.; Kim, S. Pterostilbene from Vitis coignetiae protect H2O2-induced inhibition of gap junctional intercellular communication in rat liver cell line. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2009, 47, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Lee, K.W.; Lee, H.J. Abies nephrolepis leaf phenolics prevent the inhibition of gap junction intercellular communication by hydrogen peroxide in rat liver epithelial cells. Biofactors 2004, 21, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Choi, S.H.; Kim, J.; Lee, B.K.; Lee, K.W.; Lee, H.J. Differential regulation of the hydrogen-peroxide-induced inhibition of gap-junction intercellular communication by resveratrol and butylated hydroxyanisole. Mutat. Res. 2009, 671, 40–44. [Google Scholar] [CrossRef]
- Lee, D.E.; Shin, B.J.; Hur, H.J.; Kim, J.H.; Kim, J.; Kang, N.J.; Kim, D.O.; Lee, C.Y.; Lee, K.W.; Lee, H.J. Quercetin, the active phenolic component in kiwifruit, prevents hydrogen peroxide-induced inhibition of gap-junction intercellular communication. Br. J. Nutr. 2010, 104, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Lee, B.K.; Lee, K.W.; Lee, H.J. Resveratrol counteracts gallic acid-induced down-regulation of gap-junction intercellular communication. J. Nutr. Biochem. 2009, 20, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.-W.; Cho, S.-D.; Ahn, N.-S.; Yang, S.-R.; Park, J.-S.; Jo, E.-H.; Hwang, J.-W.; Aruoma, O.I.; Lee, Y.-S.; Kang, K.-S. Effects of the histone deacetylases inhibitors sodium butyrate and trichostatin A on the inhibition of gap junctional intercellular communication by H2O2- and 12-O-tetradecanoylphorbol-13-acetate in rat liver epithelial cells. Cancer Lett. 2006, 241, 301–308. [Google Scholar] [CrossRef]
- Lee, K.W.; Hur, H.J.; Lee, H.J.; Lee, C.Y.; Ki, W.L.; Haeng, J.H.; Hyong, J.L.; Chang, Y.L.; Lee, K.W.; Hur, H.J.; et al. Antiproliferative effects of dietary phenolic substances and hydrogen peroxide. J. Agric. Food Chem. 2005, 53, 1990–1995. [Google Scholar] [CrossRef]
- Lee, K.W.; Hwang, E.-S.; Kang, N.J.; Kim, K.H.; Lee, H.J. Extraction and chromatographic separation of anticarcinogenic fractions from cacao bean husk. BioFactors 2005, 23, 141–150. [Google Scholar] [CrossRef]
- Kim, J.-S.; Lee, W.-M.; Rhee, H.C.; Kim, S. Red paprika (Capsicum annuum L.) and its main carotenoids, capsanthin and β-carotene, prevent hydrogen peroxide-induced inhibition of gap-junction intercellular communication. Chem. Biol. Interact. 2016, 254, 146–155. [Google Scholar] [CrossRef]
- Hu, W.; Jones, P.D.; Upham, B.L.; Trosko, J.E.; Lau, C.; Giesy, J.P. Inhibition of gap junctional intercellular communication by perfluorinated compounds in rat liver and dolphin kidney epithelial cell lines in vitro and Sprague-Dawley rats in vivo. Toxicol. Sci. 2002, 68, 429–436. [Google Scholar] [CrossRef]
- Upham, B.L.; Park, J.-S.; Babica, P.; Sovadinova, I.; Rummel, A.M.; Trosko, J.E.; Hirose, A.; Hasegawa, R.; Kanno, J.; Sai, K. Structure-activity-dependent regulation of cell communication by perfluorinated fatty acids using in vivo and in vitro model systems. Environ. Health Perspect. 2009, 117, 545–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madhukar, B.V.; de Feijter-Rupp, H.L.; Trosko, J.E. Pulse treatment with the tumor promoter TPA delays the onset of desensitization response and prolongs the inhibitory effect on gap junctional intercellular communication of a rat liver epithelial cell line WB F-344. Cancer Lett. 1996, 106, 117–123. [Google Scholar] [CrossRef]
- Sun, H.; Liu, G.-T. Chemopreventive effect of bicyclol on malignant transformation of WB-F344 rat liver epithelial cells and its effect on related signal transduction in vitro. Cancer Lett. 2006, 236, 239–249. [Google Scholar] [CrossRef]
- Kang, K.S.; Yun, J.W.; Yoon, B.; Lim, Y.K.; Lee, Y.S. Preventive effect of germanium dioxide on the inhibition of gap junctional intercellular communication by TPA. Cancer Lett. 2001, 166, 147–153. [Google Scholar] [CrossRef]
- Oh, S.Y.; Madhukar, B.V.; Trosko, J.E. Inhibition of gap junctional blockage by palmitoyl carnitine and TMB-8 in a rat liver epithelial cell line. Carcinogenesis 1988, 9, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-R.; Park, J.-S.; Jo, E.-H.; Hwang, J.-W.; Kim, S.-J.; Ra, J.-C.; Aruoma, O.I.; Lee, Y.-S.; Kang, K.-S. Reversal of the TPA-induced inhibition of gap junctional intercellular communication by Chaga mushroom (Inonotus obliquus) extracts: Effects on MAP kinases. Biofactors 2006, 27, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Prochazka, L.; Turanek, J.; Tesarik, R.; Knotigova, P.; Polaskova, P.; Andrysik, Z.; Kozubik, A.; Zak, F.; Sova, P.; Neuzil, J.; et al. Apoptosis and inhibition of gap-junctional intercellular communication induced by LA-12, a novel hydrophobic platinum(IV) complex. Arch. Biochem. Biophys. 2007, 462, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Čtveráčková, L.; Jančula, D.; Raška, J.; Babica, P.; Sovadinová, I. Structure-dependent effects of phthalates on intercellular and intracellular communication in liver oval cells. Int. J. Mol. Sci. 2020, 21, 6069. [Google Scholar] [CrossRef]
- Babica, P.; Zurabian, R.; Kumar, E.R.; Chopra, R.; Mianecki, M.J.; Park, J.-S.; Jasa, L.; Trosko, J.E.; Upham, B.L. Methoxychlor and vinclozolin induce rapid changes in intercellular and intracellular signaling in liver progenitor cells. Toxicol. Sci. 2016, 153, 174–185. [Google Scholar] [CrossRef]
- Ye, Y.-X.; Bombick, D.; Hirst, K.; Zhang, G.; Chang, C.-C.; Trosko, J.E.; Akera, T. The modulation of gap junctional communication by Gossypol in various mammalian cell lines in vitro. Toxicol. Sci. 1990, 14, 817–832. [Google Scholar] [CrossRef]
- Suzuki, J.; Na, H.K.; Upham, B.L.; Chang, C.C.; Trosko, J.E. lambda-Carrageenan-induced inhibition of gap-junctional intercellular communication in rat liver epithelial cells. Nutr. Cancer Int. J. 2000, 36, 122–128. [Google Scholar] [CrossRef]
- Upham, B.L.; Koski, T.R.; Rummel, A.M.; Wilson, M.R.; Horvath, A.; Trosko, J.E. Differential roles of 2, 6, and 8 carbon ceramides on the modulation of gap junctional communication and apoptosis during carcinogenesis. Cancer Lett. 2003, 191, 27–34. [Google Scholar] [CrossRef]
- Ale-Agha, N.; Stahl, W.; Sies, H. (-)-Epicatechin effects in rat liver epithelial cells: Stimulation of gap junctional communication and counteraction of its loss due to the tumor promoter 12-O-tetradecanoylphorbol-13-acetate. Biochem. Pharmacol. 2002, 63, 2145–2149. [Google Scholar] [CrossRef]
- Guan, X.; Ruch, R.J. Gap junction endocytosis and lysosomal degradation of connexin43-P2 in WB-F344 rat liver epithelial cells treated with DDT and lindane. Carcinogenesis 1996, 17, 1791–1798. [Google Scholar] [CrossRef] [PubMed]
- Sigler, K.; Ruch, R.J. Enhancement of gap junctional intercellular communication in tumor promoter-treated cells by components of green tea. Cancer Lett. 1993, 69, 15–19. [Google Scholar] [CrossRef]
- Hu, J.; Speisky, H.; Cotgreave, I.A. The inhibitory effects of boldine, glaucine, and probucol on TPA-induced down regulation of gap junction function. Relationships to intracellular peroxides, protein kinase C translocation, and connexin 43 phosphorylation. Biochem. Pharmacol. 1995, 50, 1635–1643. [Google Scholar] [CrossRef]
- Roemer, E.; Lammerich, H.-P.; Conroy, L.L.; Weisensee, D. Characterization of a gap-junctional intercellular communication (GJIC) assay using cigarette smoke. Toxicol. Lett. 2013, 219, 248–253. [Google Scholar] [CrossRef] [Green Version]
- Pierucci, F.; Frati, A.; Squecco, R.; Lenci, E.; Vicenti, C.; Slavik, J.; Francini, F.; Machala, M.; Meacci, E. Non-dioxin-like organic toxicant PCB153 modulates sphingolipid metabolism in liver progenitor cells: Its role in Cx43-formed gap junction impairment. Arch. Toxicol. 2017, 91, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Hayashi, T.; Kyoizumi, S.; Kusunoki, Y.; Nakachi, K.; MacPhee, D.G.; Trosko, J.E.; Kataoka, K.; Yorioka, N. Anisomycin downregulates gap-junctional intercellular communication via the p38 MAP-kinase pathway. J. Cell Sci. 2004, 117, 2087–2096. [Google Scholar] [CrossRef] [Green Version]
- Kuslikis, B.I.; Trosko, J.E.; Braselton, W.E.J. Mutagenicity and effect on gap-junctional intercellular communication of 4,4′-methylenebis(2-chloroaniline) and its oxidized metabolites. Mutagenesis 1991, 6, 19–24. [Google Scholar] [CrossRef]
- Jone, C.; Trosko, J.E.; Chang, C.C. Characterization of a rat liver epithelial cell line to detect inhibitors of metabolic cooperation. In Vitro Cell. Dev. Biol. J. Tissue Cult. Assoc. 1987, 23, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Bokkala, S.; Reis, H.M.; Rubin, E.; Joseph, S.K. Effect of angiotensin II and ethanol on the expression of connexin 43 in WB rat liver epithelial cells. Biochem. J. 2001, 357, 769–777. [Google Scholar] [CrossRef]
- Ren, P.; Ruch, R.J. Inhibition of gap junctional intercellular communication by barbiturates in long-term primary cultured rat hepatocytes is correlated with liver tumour promoting activity. Carcinogenesis 1996, 17, 2119–2124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerashchenko, B.I.; Howell, R.W. Cell proximity is a prerequisite for the proliferative response of bystander cells co-cultured with cells irradiated with gamma-rays. Cytom. A 2003, 56, 71–80. [Google Scholar] [CrossRef]
- Ren, P.; Mehta, P.P.; Ruch, R.J. Inhibition of gap junctional intercellular communication by tumor promoters in connexin43 and connexin32-expressing liver cells: Cell specificity and role of protein kinase C. Carcinogenesis 1998, 19, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Matesic, D.F.; Rupp, H.L.; Bonney, W.J.; Ruch, R.J.; Trosko, J.E. Changes in gap-junction permeability, phosphorylation, and number mediated by phorbol ester and non-phorbol-ester tumor promoters in rat liver epithelial cells. Mol. Carcinog. 1994, 10, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Bonney, W.J.; Ruch, R.J. Changes in gap junction permeability, gap junction number, and connexin43 expression in lindane-treated rat liver epithelial cells. Toxicol. Appl. Pharmacol. 1995, 130, 79–86. [Google Scholar] [CrossRef]
- Hasler, C.M.; Bennink, M.R.; Trosko, J.E. Inhibition of gap junction-mediated intercellular communication by alpha-linolenate. Am. J. Physiol. 1991, 261, C161–C168. [Google Scholar] [CrossRef] [PubMed]
- Hasler, C.M.; Trosko, J.E.; Bennink, M.R. Incorporation of n-3 fatty acids into WB-F344 cell phospholipids inhibits gap junctional intercellular communication. Lipids 1991, 26, 788–792. [Google Scholar] [CrossRef]
- Fransson-Steen, R.; Wärngård, L. Inhibitory effects of endosulfan on gap junctional intercellular communication in WB-F344 rat liver cells and primary rat hepatocytes. Carcinogenesis 1992, 13, 657–662. [Google Scholar] [CrossRef]
- Ehrhardt, C.; Kneuer, C.; Fiegel, J.; Hanes, J.; Schaefer, U.F.; Kim, K.-J.; Lehr, C.-M. Influence of apical fluid volume on the development of functional intercellular junctions in the human epithelial cell line 16HBE14o-: Implications for the use of this cell line as an in vitro model for bronchial drug absorption studies. Cell Tissue Res. 2002, 308, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Yeh, S.-L.; Hu, M.-L. Oxidized beta-carotene inhibits gap junction intercellular communication in the human lung adenocarcinoma cell line A549. Food Chem. Toxicol. 2003, 41, 1677–1684. [Google Scholar] [CrossRef]
- Dydowiczová, A.; Brózman, O.; Babica, P.; Sovadinová, I. Improved multiparametric scrape loading-dye transfer assay for a simultaneous high-throughput analysis of gap junctional intercellular communication, cell density and viability. Sci. Rep. 2020, 10, 730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, X.; Hardenbrook, J.; Fernstrom, M.J.; Chaudhuri, R.; Malkinson, A.M.; Ruch, R.J. Down-regulation by butylated hydroxytoluene of the number and function of gap junctions in epithelial cell lines derived from mouse lung and rat liver. Carcinogenesis 1995, 16, 2575–2582. [Google Scholar] [CrossRef]
- Brózman, O.; Novák, J.; Bauer, A.K.; Babica, P. Airborne PAHs inhibit gap junctional intercellular communication and activate MAPKs in human bronchial epithelial cell line. Environ. Toxicol. Pharmacol. 2020, 79, 103422. [Google Scholar] [CrossRef]
- Aylsworth, C.F.; Trosko, J.E.; Chang, C.C.; Benjamin, K.; Lockwood, E. Synergistic inhibition of metabolic cooperation by oleic acid or 12-0-tetradecanoylphorbol-13-acetate and dichlorodiphenyltrichlorethane (DDT) in Chinese hamster V79 cells: Implication of a role for protein kinase C in the regulation of gap junctional in. Cell Biol. Toxicol. 1989, 5, 27–37. [Google Scholar] [CrossRef]
- Wärngård, L.; Flodström, S. Effects of tetradecanoyl phorbol acetate, pyrethroids and DDT in the V79. Cell Biol. Toxicol. 1989, 5, 67–75. [Google Scholar] [CrossRef]
- Wärngård, L.; Flodström, S.; Ljungquist, S.; Ahlborg, U.G. Interaction between quercetin, TPA and DDT in the V79 metabolic cooperation assay. Carcinogenesis 1987, 8, 1201–1205. [Google Scholar] [CrossRef]
- Zeilmaker, M.J.; Yamasaki, H. Inhibition of junctional intercellular communication as a possible short-term test to detect tumor-promoting agents: Results with nine chemicals tested by dye transfer assay in Chinese hamster V79 cells. Cancer Res. 1986, 46, 6180–6186. [Google Scholar]
- Lin, Z.X.; Kavanagh, T.; Trosko, J.E.; Chang, C.C. Inhibition of gap junctional intercellular communication in human teratocarcinoma cells by organochlorine pesticides. Toxicol. Appl. Pharmacol. 1986, 83, 10–19. [Google Scholar] [CrossRef]
- Tsushimoto, G.; Chang, C.C.; Trosko, J.E.; Matsumura, F. Cytotoxic, mutagenic, and cell-cell communication inhibitory properties of DDT, lindane, and chlordane on Chinese hamster cells in vitro. Arch. Environ. Contam. Toxicol. 1983, 12, 721–729. [Google Scholar] [CrossRef]
- Lee, I.-K.; Rhee, S.-K. Inhibitory effect of bisphenol A on gap junctional intercellular communication in an epithelial cell line of rat mammary tissue. Arch. Pharm. Res. 2007, 30, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Tai, M.-H.; Upham, B.L.; Olson, L.K.; Tsao, M.-S.; Reed, D.N.J.; Trosko, J.E. Cigarette smoke components inhibited intercellular communication and differentiation in human pancreatic ductal epithelial cells. Int. J. Cancer 2007, 120, 1855–1862. [Google Scholar] [CrossRef] [PubMed]
- Carruba, G.; Stefano, R.; Cocciadiferro, L.; Saladino, F.; Di Cristina, A.; Tokar, E.; Quader, S.T.A.; Webber, M.M.; Castagnetta, L. Intercellular communication and human prostate carcinogenesis. Ann. N. Y. Acad. Sci. 2002, 963, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Carruba, G.; Webber, M.M.; Quader, S.T.A.; Amoroso, M.; Cocciadiferro, L.; Saladino, F.; Trosko, J.E.; Castagnetta, L.A.M. Regulation of cell-to-cell communication in non-tumorigenic and malignant human prostate epithelial cells. Prostate 2002, 50, 73–82. [Google Scholar] [CrossRef]
- Tateno, C.; Ito, S.; Tanaka, M.; Yoshitake, A. Effects of pyrethroid insecticides on gap junctional intercellular communications in Balb/c3T3 cells by dye-transfer assay. Cell Biol. Toxicol. 1993, 9, 215–221. [Google Scholar] [CrossRef]
- Zhang, Q.; Wu, S.; Liu, L.; Hou, X.; Jiang, J.; Wei, X.; Hao, W. Effects of bisphenol A on gap junctions in HaCaT cells as mediated by the estrogen receptor pathway. J. Appl. Toxicol. 2019, 39, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Swierenga, S.H.H.; Yamasaki, H.; Piccoli, C.; Robertson, L.; Bourgon, L.; Marceau, N.; Fitzgerald, D.J. Effects of intercellular communication in human keratinocytes and liver-derived cells of polychlorinated biphenyl congeners with differing in vivo promotion activities. Carcinogenesis 1990, 11, 921–926. [Google Scholar] [CrossRef]
- Choung, Y.-H.; Choi, S.J.; Joo, J.S.; Lee, J.B.; Lee, H.K.; Lee, S.J. Green tea prevents down-regulation of gap junction intercellular communication in human keratinocytes treated with PMA. Eur. Arch. Otorhinolaryngol. 2011, 268, 885–892. [Google Scholar] [CrossRef] [Green Version]
- Defamie, N.; Mograbi, B.; Roger, C.; Cronier, L.; Malassine, A.; Brucker-Davis, F.; Fenichel, P.; Segretain, D.; Pointis, G. Disruption of gap junctional intercellular communication by lindane is associated with aberrant localization of connexin43 and zonula occludens-1 in 42GPA9 Sertoli cells. Carcinogenesis 2001, 22, 1537–1542. [Google Scholar] [CrossRef] [Green Version]
- Fiorini, C.; Gilleron, J.; Carette, D.; Valette, A.; Tilloy, A.; Chevalier, S.; Segretain, D.; Pointis, G. Accelerated internalization of junctional membrane proteins (connexin 43, N-cadherin and ZO-1) within endocytic vacuoles: An early event of DDT carcinogenicity. Biochim. Biophys. Acta 2008, 1778, 56–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yawer, A.; Sychrová, E.; Labohá, P.; Raška, J.; Jambor, T.; Babica, P.; Sovadinová, I. Endocrine-disrupting chemicals rapidly affect intercellular signaling in Leydig cells. Toxicol. Appl. Pharmacol. 2020, 404, 115177. [Google Scholar] [CrossRef] [PubMed]
- Aravindakshan, J.; Cyr, D.G. Nonylphenol alters connexin 43 levels and connexin 43 phosphorylation via an inhibition of the p38-mitogen-activated protein kinase pathway. Biol. Reprod. 2005, 72, 1232–1240. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Siragam, V.; Chen, J.; Fridman, M.D.; Hamilton, R.M.; Sun, Y. High-throughput measurement of gap junctional intercellular communication. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H1708–H1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofgaard, J.P.; Mollerup, S.; Holstein-Rathlou, N.-H.; Nielsen, M.S. Quantification of gap junctional intercellular communication based on digital image analysis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R243–R247. [Google Scholar] [CrossRef] [Green Version]
- Begandt, D.; Bader, A.; Antonopoulos, G.C.; Schomaker, M.; Kalies, S.; Meyer, H.; Ripken, T.; Ngezahayo, A. Gold nanoparticle-mediated (GNOME) laser perforation: A new method for a high-throughput analysis of gap junction intercellular coupling. J. Bioenerg. Biomembr. 2015, 47, 441–449. [Google Scholar] [CrossRef]
- Homolya, L.; Holló, Z.; Germann, U.A.; Pastan, I.; Gottesman, M.M.; Sarkadi, B. Fluorescent cellular indicators are extruded by the multidrug resistance protein. J. Biol. Chem. 1993, 268, 21493–21496. [Google Scholar] [CrossRef]
- Picoli, C.; Soleilhac, E.; Journet, A.; Barette, C.; Comte, M.; Giaume, C.; Mouthon, F.; Fauvarque, M.-O.; Charveriat, M. High-content screening identifies new inhibitors of connexin 43 gap junctions. Assay Drug Dev. Technol. 2019, 17, 240–248. [Google Scholar] [CrossRef]
- Li, Z.; Yan, Y.; Powers, E.A.; Ying, X.; Janjua, K.; Garyantes, T.; Baron, B. Identification of gap junction blockers using automated fluorescence microscopy imaging. J. Biomol. Screen. 2003, 8, 489–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, N.; Bathany, C.; Hua, S.Z. Assay for molecular transport across gap junction channels in one-dimensional cell arrays. Lab Chip 2011, 11, 1096–1101. [Google Scholar] [CrossRef]
- Chen, S.; Lee, L.P. Non-invasive microfluidic gap junction assay. Integr. Biol. 2010, 2, 130–138. [Google Scholar] [CrossRef]
- Haq, N.; Grose, D.; Ward, E.; Chiu, O.; Tigue, N.; Dowell, S.J.; Powell, A.J.; Chen, M.X. A high-throughput assay for connexin 43 (Cx43, GJA1) gap junctions using codon-optimized aequorin. Assay Drug Dev. Technol. 2013, 11, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Choi, E.J.; Lee, J. A new high-throughput screening-compatible gap junctional intercellular communication assay. BMC Biotechnol. 2015, 15, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeo, J.H.; Lee, J. An iodide-yellow fluorescent protein-gap junction-intercellular communication assay. JOVE J. Vis. Exp. 2019, 144, e58966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danish, A.; Gedschold, R.; Hinz, S.; Schiedel, A.C.; Thimm, D.; Bedner, P.; Steinhäuser, C.; Müller, C.E. A cellular assay for the identification and characterization of connexin gap junction modulators. Int. J. Mol. Sci. 2021, 22, 1417. [Google Scholar] [CrossRef]
- El-Fouly, M.H.; Trosko, J.E.; Chang, C.-C. Scrape-loading and dye transfer: A rapid and simple technique to study gap junctional intercellular communication. Exp. Cell Res. 1987, 168, 422–430. [Google Scholar] [CrossRef]
- Babica, P.; Sovadinová, I.; Upham, B.L. Scrape loading/dye transfer assay. In Gap Junction Protocols; Vinken, M., Johnstone, S.R., Eds.; Springer: New York, NY, USA, 2016; Volume 1437, pp. 133–144. ISBN 978-1-4939-3664-9. [Google Scholar]
- Tsao, M.S.; Smith, J.D.; Nelson, K.G.; Grisham, J.W. A diploid epithelial cell line from normal adult rat liver with phenotypic properties of “oval” cells. Exp. Cell Res. 1984, 154, 38–52. [Google Scholar] [CrossRef]
- Shafritz, D.A.; Dabeva, M.D. Liver stem cells and model systems for liver repopulation. J. Hepatol. 2002, 36, 552–564. [Google Scholar] [CrossRef]
- De Feijter, A.W.; Ray, J.S.; Weghorst, C.M.; Klaunig, J.E.; Goodman, J.I.; Chang, C.C.; Ruch, R.J.; Trosko, J.E. Infection of rat liver epithelial cells with v-Ha-ras: Correlation between oncogene expression, gap junctional communication, and tumorigenicity. Mol. Carcinog. 1990, 3, 54–67. [Google Scholar] [CrossRef]
- Coleman, W.B.; McCullough, K.D.; Esch, G.L.; Faris, R.A.; Hixson, D.C.; Smith, G.J.; Grisham, J.W. Evaluation of the differentiation potential of WB-F344 rat liver epithelial stem-like cells in vivo. Differentiation to hepatocytes after transplantation into dipeptidylpeptidase-IV-deficient rat liver. Am. J. Pathol. 1997, 151, 353–359. [Google Scholar]
- Grisham, J.W.; Coleman, W.B.; Smith, G.J. Isolation, culture, and transplantation of rat hepatocytic precursor (stem-like) cells. Proc. Soc. Exp. Biol. Med. 1993, 204, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Malouf, N.N.; Coleman, W.B.; Grisham, J.W.; Lininger, R.A.; Madden, V.J.; Sproul, M.; Anderson, P.A. Adult-derived stem cells from the liver become myocytes in the heart in vivo. Am. J. Pathol. 2001, 158, 1929–1935. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.-Y.; Xue, X.-W.; Shi, Z.-M.; Wang, P.-Y.; Wu, X.-R.; Wang, X.-J. Oleanolic acid and ursolic acid inhibit proliferation in transformed rat hepatic oval cells. World J. Gastroenterol. 2014, 20, 1348–1356. [Google Scholar] [CrossRef]
- Labine, M.; Minuk, G.Y. Long-term, low-dose exposure to microcystin toxin does not increase the risk of liver tumor development or growth in mice. Hepatol. Res. 2015, 45, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Liu, G. Chemopreventive effect of dimethyl dicarboxylate biphenyl on malignant transformation of WB-F344 rat liver epithelial cells. Acta Pharmacol. Sin. 2005, 26, 1339–1344. [Google Scholar] [CrossRef]
- Oh, S.Y.; Dupont, E.; Madhukar, B.V.; Briand, J.P.; Chang, C.C.; Beyer, E.; Trosko, J.E. Characterization of gap junctional communication-deficient mutants of a rat liver epithelial cell line. Eur. J. Cell Biol. 1993, 60, 250–255. [Google Scholar] [PubMed]
- DeoCampo, N.D.; Wilson, M.R.; Trosko, J.E. Cooperation of bcl-2 and myc in the neoplastic transformation of normal rat liver epithelial cells is related to the down-regulation of gap junction-mediated intercellular communication. Carcinogenesis 2000, 21, 1501–1506. [Google Scholar]
- Hayashi, T.; Nomata, K.; Chang, C.C.; Ruch, R.J.; Trosko, J.E. Cooperative effects of v-myc and c-Ha-ras oncogenes on gap junctional intercellular communication and tumorigenicity in rat liver epithelial cells. Cancer Lett. 1998, 128, 145–154. [Google Scholar] [CrossRef]
- Jou, Y.S.; Layhe, B.; Matesic, D.F.; Chang, C.C.; de Feijter, A.W.; Lockwood, L.; Welsch, C.W.; Klaunig, J.E.; Trosko, J.E. Inhibition of gap junctional intercellular communication and malignant transformation of rat liver epithelial cells by neu oncogene. Carcinogenesis 1995, 16, 311–317. [Google Scholar] [CrossRef]
- Hooth, M.J.; Coleman, W.B.; Presnell, S.C.; Borchert, K.M.; Grisham, J.W.; Smith, G.J. Spontaneous neoplastic transformation of WB-F344 rat liver epithelial cells. Am. J. Pathol. 1998, 153, 1913–1921. [Google Scholar] [CrossRef] [Green Version]
- de Feijter, A.W.; Matesic, D.F.; Ruch, R.J.; Guan, X.J.; Chang, C.C.; Trosko, J.E.; de Feijter, A.W. Localization and function of the connexin 43 gap-junction protein in normal and various oncogene-expressing rat liver epithelial cells. Mol. Carcinog. 1996, 16, 203–212. [Google Scholar] [CrossRef]
- Rae, R.S.; Mehta, P.P.; Chang, C.C.; Trosko, J.E.; Ruch, R.J. Neoplastic phenotype of gap-junctional intercellular communication-deficient WB rat liver epithelial cells and its reversal by forced expression of connexin 32. Mol. Carcinog. 1998, 22, 120–127. [Google Scholar] [CrossRef]
- Jou, Y.S.; Matesic, D.; Dupont, E.; Lu, S.C.; Rupp, H.L.; Madhukar, B.V.; Oh, S.Y.; Trosko, J.E.; Chang, C.C. Restoration of gap-junctional intercellular communication in a communication-deficient rat liver cell mutant by transfection with connexin 43 cDNA. Mol. Carcinog. 1993, 8, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Upham, B.L.; Suzuki, J.; Chen, G.; Wang, Y.; McCabe, L.R.; Chang, C.-C.; Krutovskikh, V.A.; Yamasaki, H.; Trosko, J.E. Reduced gap junctional intercellular communication and altered biological effects in mouse osteoblast and rat liver oval cell lines transfected with dominant-negative connexin 43. Mol. Carcinog. 2003, 37, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Caruso, R.L.; Upham, B.L.; Harris, C.; Trosko, J.E. Biphasic lindane-induced oxidation of glutathione and inhibition of gap junctions in myometrial cells. Toxicol. Sci. 2005, 86, 417–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SciRap In Vitro Web-Based Tool. Available online: http://www.scirap.org/ (accessed on 16 June 2021).
- Bláha, L.; Babica, P.; Hilscherová, K.; Upham, B.L. Inhibition of gap-junctional intercellular communication and activation of mitogen-activated protein kinases by cyanobacterial extracts-indications of novel tumor-promoting cyanotoxins? Toxicon 2010, 55, 126–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corvi, R.; Madia, F. EURL ECVAM Genotoxicity and Carcinogenicity Consolidated Database of Ames Positive Chemicals; European Commission: Brussels, Belgium, 2018. [Google Scholar]
- Madia, F.; Kirkland, D.; Morita, T.; White, P.; Asturiol, D.; Corvi, R. EURL ECVAM genotoxicity and carcinogenicity database of substances eliciting negative results in the Ames test: Construction of the database. Mutat. Res. Toxicol. Environ. Mutagen. 2020, 854–855, 503199. [Google Scholar] [CrossRef]
- Warren, S.T.; Doolittle, D.J.; Chang, C.C.; Goodmann, J.I.; Trosko, J.E. Evaluation of the carcinogenic potential of 2,4-dinitrofluorobenzene and its implications regarding mutagenicity testing. Carcinogenesis 1982, 3, 139–145. [Google Scholar] [CrossRef]
- IARC Monographs. Available online: https://monographs.iarc.fr/list-of-classifications (accessed on 10 October 2020).
- Shi, H.; Hardesty, J.E.; Jin, J.; Head, K.Z.; Falkner, K.C.; Cave, M.C.; Prough, R.A. Concentration dependence of human and mouse aryl hydrocarbon receptor responsiveness to polychlorinated biphenyl exposures: Implications for aroclor mixtures. Xenobiotica 2019, 49, 1414–1422. [Google Scholar] [CrossRef]
- Baker, N.A.; Shoemaker, R.; English, V.; Larian, N.; Sunkara, M.; Morris, A.J.; Walker, M.; Yiannikouris, F.; Cassis, L.A. Effects of adipocyte aryl hydrocarbon receptor deficiency on PCB-induced disruption of glucose homeostasis in lean and obese mice. Environ. Health Perspect. 2015, 123, 944–950. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.A.; Karounos, M.; English, V.; Fang, J.; Wei, Y.; Stromberg, A.; Sunkara, M.; Morris, A.J.; Swanson, H.I.; Cassis, L.A. Coplanar polychlorinated biphenyls impair glucose homeostasis in lean C57BL/6 mice and mitigate beneficial effects of weight loss on glucose homeostasis in obese mice. Environ. Health Perspect. 2013, 121, 105–110. [Google Scholar] [CrossRef]
- Hennig, B.; Meerarani, P.; Slim, R.; Toborek, M.; Daugherty, A.; Silverstone, A.E.; Robertson, L.W. Proinflammatory properties of coplanar PCBs: In vitro and in vivo evidence. Toxicol. Appl. Pharmacol. 2002, 181, 174–183. [Google Scholar] [CrossRef]
- Hao, J.-L.; Suzuki, K.; Lu, Y.; Hirano, S.; Fukuda, K.; Kumagai, N.; Kimura, K.; Nishida, T. Inhibition of gap junction–mediated intercellular communication by TNF-α in cultured human corneal fibroblasts. Investig. Ophthalmol. Vis. Sci. 2005, 46, 1195–1200. [Google Scholar] [CrossRef]
- Hu, J.; Cotgreave, I.A. Differential regulation of gap junctions by proinflammatory mediators in vitro. J. Clin. Investig. 1997, 99, 2312–2316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Rijen, H.V.; van Kempen, M.J.; Postma, S.; Jongsma, H.J. Tumour necrosis factor alpha alters the expression of connexin43, connexin40, and connexin37 in human umbilical vein endothelial cells. Cytokine 1998, 10, 258–264. [Google Scholar] [CrossRef]
- Osgood, R.S.; Upham, B.L.; Hill, T., 3rd; Helms, K.L.; Velmurugan, K.; Babica, P.; Bauer, A.K.; Hill III, T.; Helms, K.L.; Velmurugan, K.; et al. Polycyclic aromatic hydrocarbon-induced signaling events relevant to inflammation and tumorigenesis in lung cells are dependent on molecular structure. PLoS ONE 2013, 8, e65150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romo, D.; Velmurugan, K.; Upham, B.L.; Dwyer-Nield, L.D.; Bauer, A.K. Dysregulation of gap junction function and cytokine production in response to non-genotoxic polycyclic aromatic hydrocarbons in an in vitro lung cell model. Cancers 2019, 11, 572. [Google Scholar] [CrossRef] [Green Version]
- Siegrist, K.J.; Romo, D.; Upham, B.L.; Armstrong, M.; Quinn, K.; Vanderlinden, L.; Osgood, R.S.; Velmurugan, K.; Elie, M.; Manke, J.; et al. Early mechanistic events induced by low molecular weight polycyclic aromatic hydrocarbons in mouse lung epithelial cells: A role for eicosanoid signaling. Toxicol. Sci. 2019, 169, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Hill, T., 3rd; Osgood, R.S.; Velmurugan, K.; Alexander, C.-M.; Upham, B.L.; Bauer, A.K. Bronchoalveolar lavage fluid utilized ex vivo to validate in vivo findings: Inhibition of gap junction activity in lung tumor promotion is Toll-like receptor 4-dependent. J. Mol. Biomark. Diagn. 2013, 5, 22168. [Google Scholar] [CrossRef]
- Raska, J.; Ctverackova, L.; Dydowiczova, A.; Sovadinova, I.; Blaha, L.; Babica, P. Tumor-promoting cyanotoxin microcystin-LR does not induce procarcinogenic events in adult human liver stem cells. Toxicol. Appl. Pharmacol. 2018, 345, 103–113. [Google Scholar] [CrossRef]
- Kubickova, B.; Babica, P.; Hilscherová, K.; Šindlerová, L. Effects of cyanobacterial toxins on the human gastrointestinal tract and the mucosal innate immune system. Environ. Sci. Eur. 2019, 31, 31. [Google Scholar] [CrossRef] [Green Version]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef] [PubMed]
- Vondráček, J.; Machala, M. Environmental ligands of the aryl hydrocarbon receptor and their effects in models of adult liver progenitor cells. Stem Cells Int. 2016, 2016, 4326194. [Google Scholar] [CrossRef] [Green Version]
- Bauer, A.K.; Velmurugan, K.; Plöttner, S.; Siegrist, K.J.; Romo, D.; Welge, P.; Brüning, T.; Xiong, K.-N.; Käfferlein, H.U. Environmentally prevalent polycyclic aromatic hydrocarbons can elicit co-carcinogenic properties in an in vitro murine lung epithelial cell model. Arch. Toxicol. 2018, 92, 1311–1322. [Google Scholar] [CrossRef] [Green Version]
- Kass, L.; Gomez, A.L.; Altamirano, G.A. Relationship between agrochemical compounds and mammary gland development and breast cancer. Mol. Cell. Endocrinol. 2020, 508, 110789. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.J.; Grulke, C.M.; Edwards, J.; McEachran, A.D.; Mansouri, K.; Baker, N.C.; Patlewicz, G.; Shah, I.; Wambaugh, J.F.; Judson, R.S.; et al. The comptox chemistry dashboard: A community data resource for environmental chemistry. J. Cheminform. 2017, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- US EPA. Oncologic 9/8. Available online: https://www.epa.gov/tsca-screening-tools/oncologictm-expert-system-evaluate-carcinogenic-potential-chemicals (accessed on 10 April 2021).
- Sakai, A.; Iwase, Y.; Nakamura, Y.; Sasaki, K.; Tanaka, N.; Umeda, M. Use of a cell transformation assay with established cell lines, and a metabolic cooperation assay with V79 cells for the detection of tumour promoters: A review. Altern. Lab. Anim. 2002, 30, 33–59. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, M.; Rosenkranz, H.S.; Klopman, G. Intercellular communication, tumor promotion and non-genotoxic carcinogenesis: Relationships based upon structural considerations. Mutat. Res. 1997, 381, 171–188. [Google Scholar] [CrossRef]
- Hernández-Guerra, M.; Hadjihambi, A.; Jalan, R. Gap junctions in liver disease: Implications for pathogenesis and therapy. J. Hepatol. 2019, 70, 759–772. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, E.; Faris, R.A.; Spray, D.C.; Monfils, B.; Abreu, S.; Danishefsky, I.; Reid, L.M. Correlation of expression of connexin mRNA isoforms with degree of cellular differentiation. Cell Adhes. Commun. 1996, 4, 223–235. [Google Scholar] [CrossRef]
- Neveu, M.J.; Sattler, C.A.; Sattler, G.L.; Hully, J.R.; Hertzberg, E.L.; Paul, D.L.; Nicholson, B.J.; Pitot, H.C. Differences in the expression of connexin genes in rat hepatomas in vivo and in vitro. Mol. Carcinog. 1994, 11, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Piechocki, M.P.; Burk, R.D.; Ruch, R.J. Regulation of connexin32 and connexin43 gene expression by DNA methylation in rat liver cells. Carcinogenesis 1999, 20, 401–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.S.; Babica, P.; Sovadinová, I.; Trosko, J.E.; Chang, C.C.; Upham, B. The Effects of Polycyclic Aromatic Hydrocarbons on Adult Human Epithelial Liver Stem Cell Line. In Proceedings of the First Midwest Conference on Stem Cell Biology and Therapy-Book of Abstracts, Rochester, MI, USA, 9–11 May 2008; p. 52. [Google Scholar]
- Bauer, A.K.; Upham, B.L.; Rondini, E.A.; Tennis, M.A.; Velmuragan, K.; Wiese, D. Toll-like receptor expression in human non-small cell lung carcinoma: Potential prognostic indicators of disease. Oncotarget 2017, 8, 91860–91875. [Google Scholar] [CrossRef] [Green Version]
- Ouzzani, M.; Hammady, H.; Fedorowicz, Z.; Elmagarmid, A. Rayyan—a web and mobile app for systematic reviews. Syst. Rev. 2016, 5, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watford, S.; Ly Pham, L.; Wignall, J.; Shin, R.; Martin, M.T.; Friedman, K.P. ToxRefDB version 2.0: Improved utility for predictive and retrospective toxicology analyses. Reprod. Toxicol. 2019, 89, 145–158. [Google Scholar] [CrossRef] [PubMed]



| Method | Tracer | Cell Line | Set-Up | HTS/HCA/HCS Features | Advantages +/Disadvantages − |
|---|---|---|---|---|---|
| Dye transfer assays | |||||
| Multiparametric SL-DT assay [259] | LY | WB-F344 HBE1 Beas-2B TM3 TM4 | 24-96-wp | Platform ready for automated cell seeding or compound adding. Automatic fixed cell imaging. | + Endpoints: GJIC, cell density and viability + Applicable for a variety of adherent cell types + Automated image acquisition and analysis + No specialized cell model, equipment or technical skills needed − Invasive − For cells forming nearly confluent monolayers − Not applicable for Cx channels excluding LY |
| Microinjection [280] | HPTS | HeLa HEK293 HL-1 | 35-mm Petri dish | Robotic microinjection system. Automatic live cell imaging. | + Precise and quantitative + Instantaneous delivery + Automated microinjection of a large of cells + Permits the correlation of morphological + Enables kinetic studies (the transfer rate from one cell to another) and functional data from individual cells − Manual image analysis − Low throughput/time consuming − Specialized microinjection equipment needed − Invasive − Unsuitable for rapid effects or requiring a continuous application of the stimulus |
| Electroporation [281] | LY | Cx43-C6 | Electroporation slides | Manual live-cell imaging by light and fluorescence microscopy. Low throughput. | + A rapid and objective quantification with a high degree of reproducibility + Applicable to a large variety of adherent cell types + Semiautomatic image analysis − A specialized electroporation equipment needed − Low throughput − Unsuitable for poorly adherent cells − Cell health concerns − Not applicable for Cx channels excluding LY |
| GNOME-LP/DT [282] | LY | GM-7373 RBE4 | 24-96-wp | Automatic fixed cell imaging. | + Automated image analysis + Non-invasive + Semiautomatic image analysis + Applicable for 3D models or tissue − Low throughput − Inappropriate to investigate cell assemblies or low-density cultures − Specialized microscopic equipment needed − Not applicable for Cx channels excluding LY |
| Parachute assay | Calcein [243] | WB F344 | 96-wp | Platform ready for automated cell seeding or compound adding. Automatic live cell imaging. | + Automated image acquisition and analysis + Semiautomated HCS − Relatively long (2–5 h) time-lapse image acquisition − Nonspecific dye transfer of calcein has been observed [97] − Calcein can be actively pumped out by MRPs [283] − Trypsinization of the donor cells − Formation of GJ channels between donor and recipient cells during exposure |
| Calcein/Vybrant DiI [284] | U251 | ||||
| Preloading assay | Calcein | IAR-20 [139] | 384-wp | Automated cell seeding. Automated compound adding. | + Automated image acquisition and analysis + The high level of automation with very few manual steps − Special live microscopy equipment needed − Relatively long (2 h) time-lapse image acquisition − Nonspecific dye transfer of calcein has been observed [97] − Calcein can be actively pumped out by MRPs [283] − Trypsinizations of the preloaded cells − Formation of GJ channels between donor and recipient cells during (or after) exposure |
| Cxs-C6 [285] | |||||
| Microfluidic assay | CFDA [286] | NRK-49F | Chip | Microfluidic platform. Automatic live cell imaging. | + Non-invasive + In situ monitoring of GJIC + Rapid screening + Applicable for studying the kinetics of gap junction channels diffusion + Ease-of-use + Can be scaled up for high-throughput applications + Low reagent consumption − Specialized microfluidic equipment needed − Only suitable for monolayer forming cells − A limited number of studied cells − Nonspecific dye transfer of calcein has been observed [97] − Calcein can be actively pumped out by MRPs [283] |
| Calcein [287] | C6 Cx43-C6 HeLa | ||||
| Metabolic cooperation assays | |||||
| Cx43 GJ aequorin assay [288] | Calcium | HeLa CHO HEK29 U2OS (transduced with Cx43 and cytoAeq/α1-ARs or TRPV1) | 384-wp | Platform ready for automated cell seeding and compound adding. Luminescence plate reader. | + HTS screening assay (miniaturization) + Non-invasive + Does not require complex equipment and analysis − Genetically modified cells needed − High probability of false-positive hits − Optimized only for Cx43 |
| I--YFPQL assay [289,290] | Iodide | LN215 HOS (transduced or non-transducted with iodide transporter/iodide sensor protein) | 96-wp | Platform ready for automated cell seeding and compound adding. Fluorescence plate reader. | + HTS screening assay (miniaturization) + Non-invasive + Short assay time (10 s) + Does not require complex equipment and analysis − Genetically modified cells needed − High probability of false-positive hits − Optimized only for Cx43 |
| cAMP based assay [291] | cAMP | HeLa (transfected with Cx43 and A2AAR or GloSensor-20F) | 96-wp | Platform ready for automated cell seeding and compound adding. Luminescence plate reader. | + HTS screening assay (miniaturization) + Non-invasive + Short assay time (1 s) + Does not require complex equipment and analysis + Applicable for studying the kinetics of gap junction channels diffusion − Genetically modified cells needed − High probability of false-positive hits − Optimized only for Cx43 |
| SL-DT Assay | Total Chemicals | |||
|---|---|---|---|---|
| Carcinogenicity | ↓ a | — or —/↓ or E b | ||
| IARC | Group 1, 2A and 2B | 33 | 10 | 72 |
| Group 3 | 16 | 13 | ||
| Sensitivity | 77% (33/43) | |||
| Specificity | 45% (13/29) | |||
| Accuracy | 64% (46/72) | |||
| Group 1 | 15 | 5 | 20 | |
| Sensitivity | 75% (15/20) | |||
| CompTox | + c | 59 | 23 | 82 |
| Sensitivity | 73% (60/82) | |||
| OncoLogic | Low-moderate, Moderate, Moderate-high, High | 58 | 29 | 143 |
| Low, Marginal, Marginal to High-moderate, Low to Moderate to Marginal, Low to Moderate, Marginal to Low moderate | 43 | 13 | ||
| Sensitivity | 67% (58/87) | |||
| Specificity | 23% (13/56) | |||
| Accuracy | 50% (71/143) | |||
| MC Assay | ↓ a | 15 | 0 | 31 |
| —, E b | 11 | 5 | ||
| Sensitivity | 100% (15/15) | |||
| Specificity | 31% (5/16) | |||
| Accuracy | 65% (20/31) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sovadinová, I.; Upham, B.L.; Trosko, J.E.; Babica, P. Applicability of Scrape Loading-Dye Transfer Assay for Non-Genotoxic Carcinogen Testing. Int. J. Mol. Sci. 2021, 22, 8977. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168977
Sovadinová I, Upham BL, Trosko JE, Babica P. Applicability of Scrape Loading-Dye Transfer Assay for Non-Genotoxic Carcinogen Testing. International Journal of Molecular Sciences. 2021; 22(16):8977. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168977
Chicago/Turabian StyleSovadinová, Iva, Brad L. Upham, James E. Trosko, and Pavel Babica. 2021. "Applicability of Scrape Loading-Dye Transfer Assay for Non-Genotoxic Carcinogen Testing" International Journal of Molecular Sciences 22, no. 16: 8977. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168977