
Mitogen Kinase Kinase (MKK7) Controls Cytokine Production In Vitro and In Vivo in Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Conditional MKK7 Mice Generation
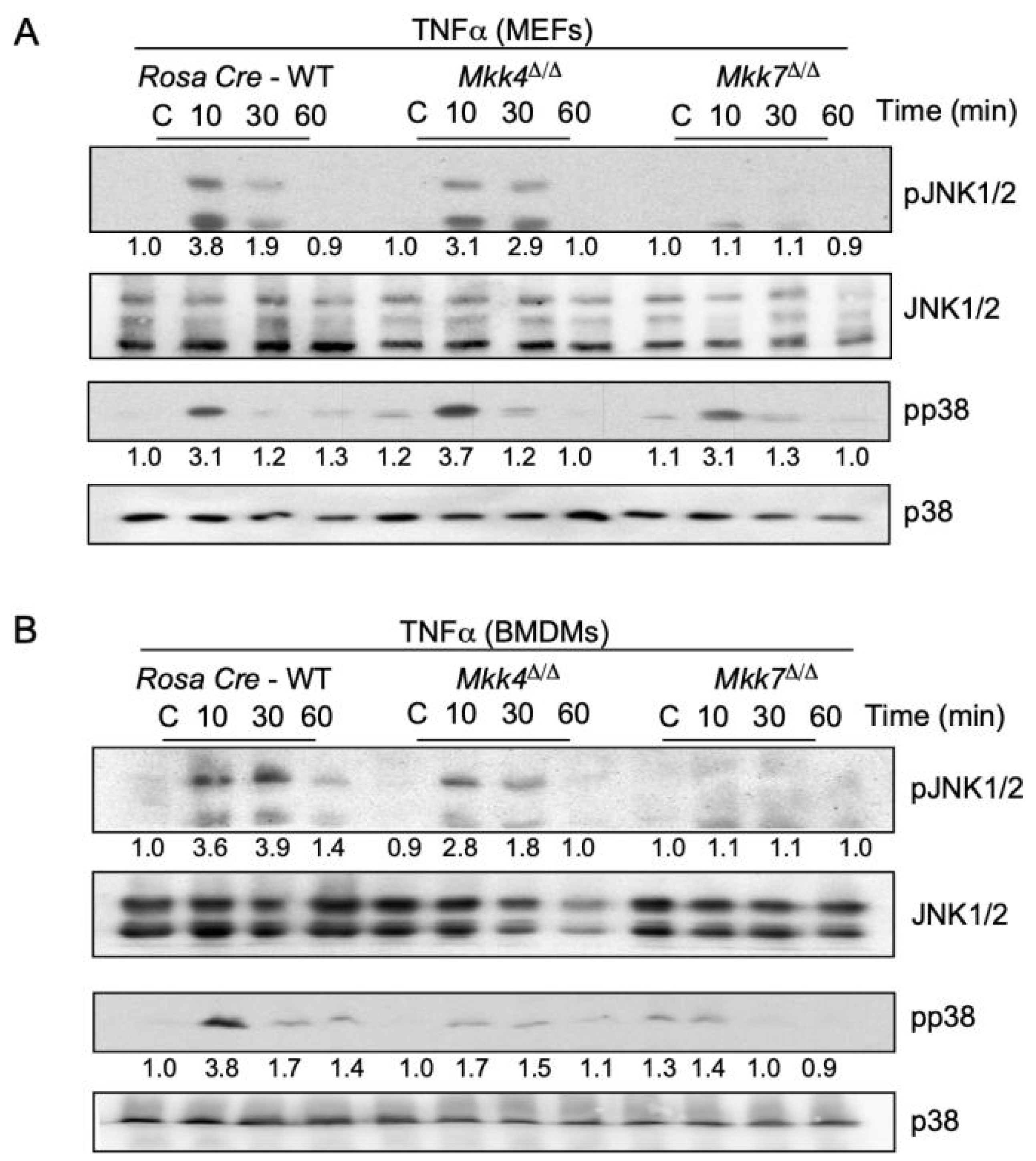
2.2. MKK7 Is Required for TNFα-Stimulated MAP Kinase Activation
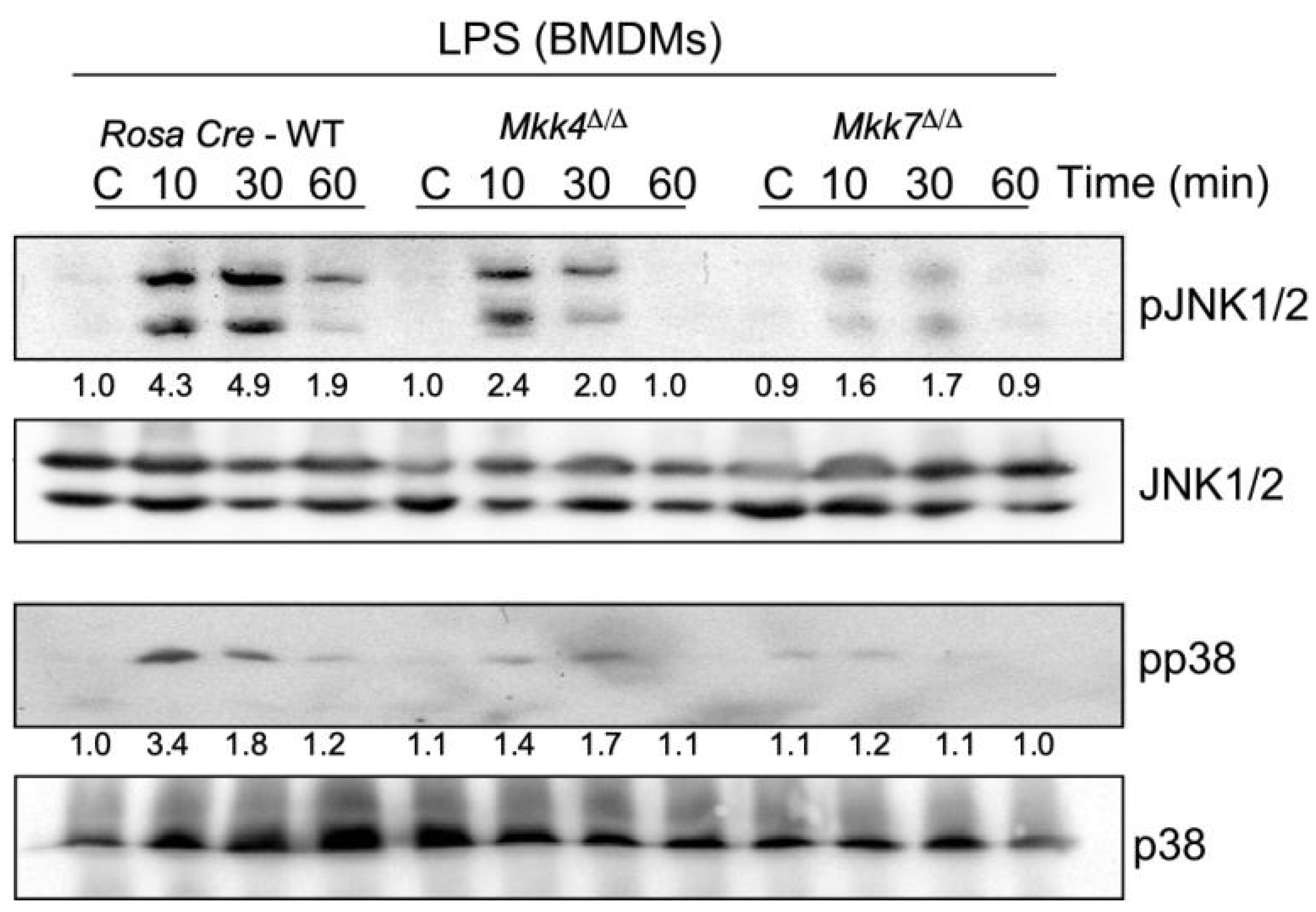
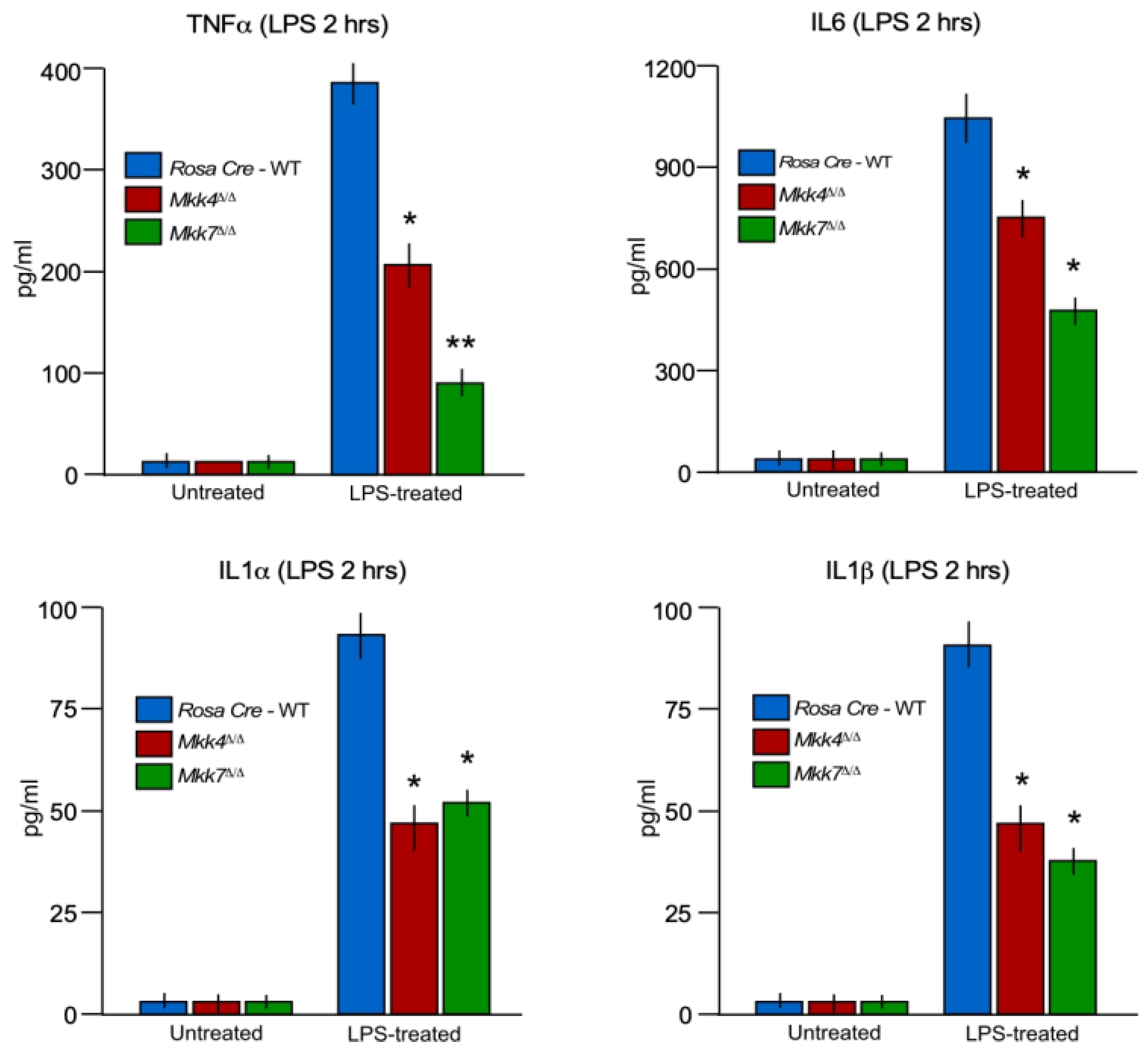
2.3. MKK7 Plays an Important Role in LPS-Induced MAP Kinase Activation and Cytokine Production
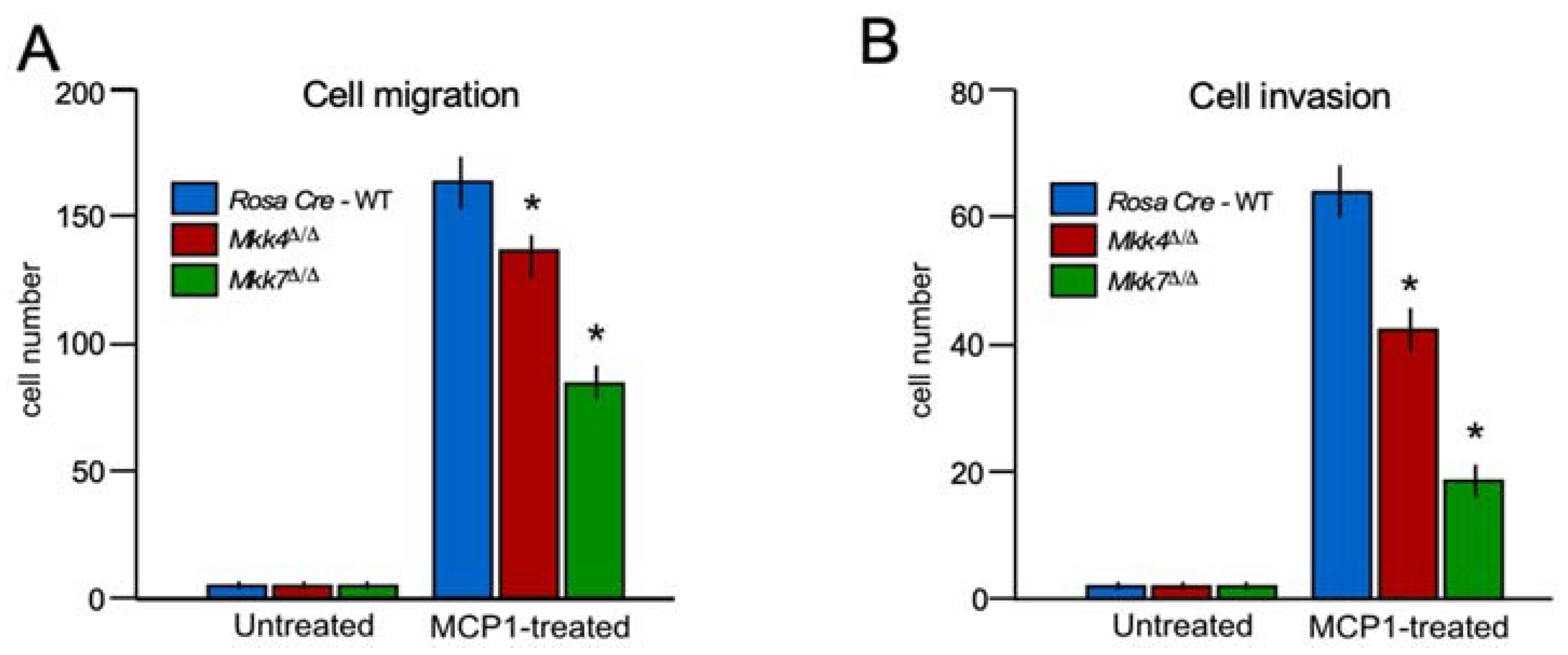
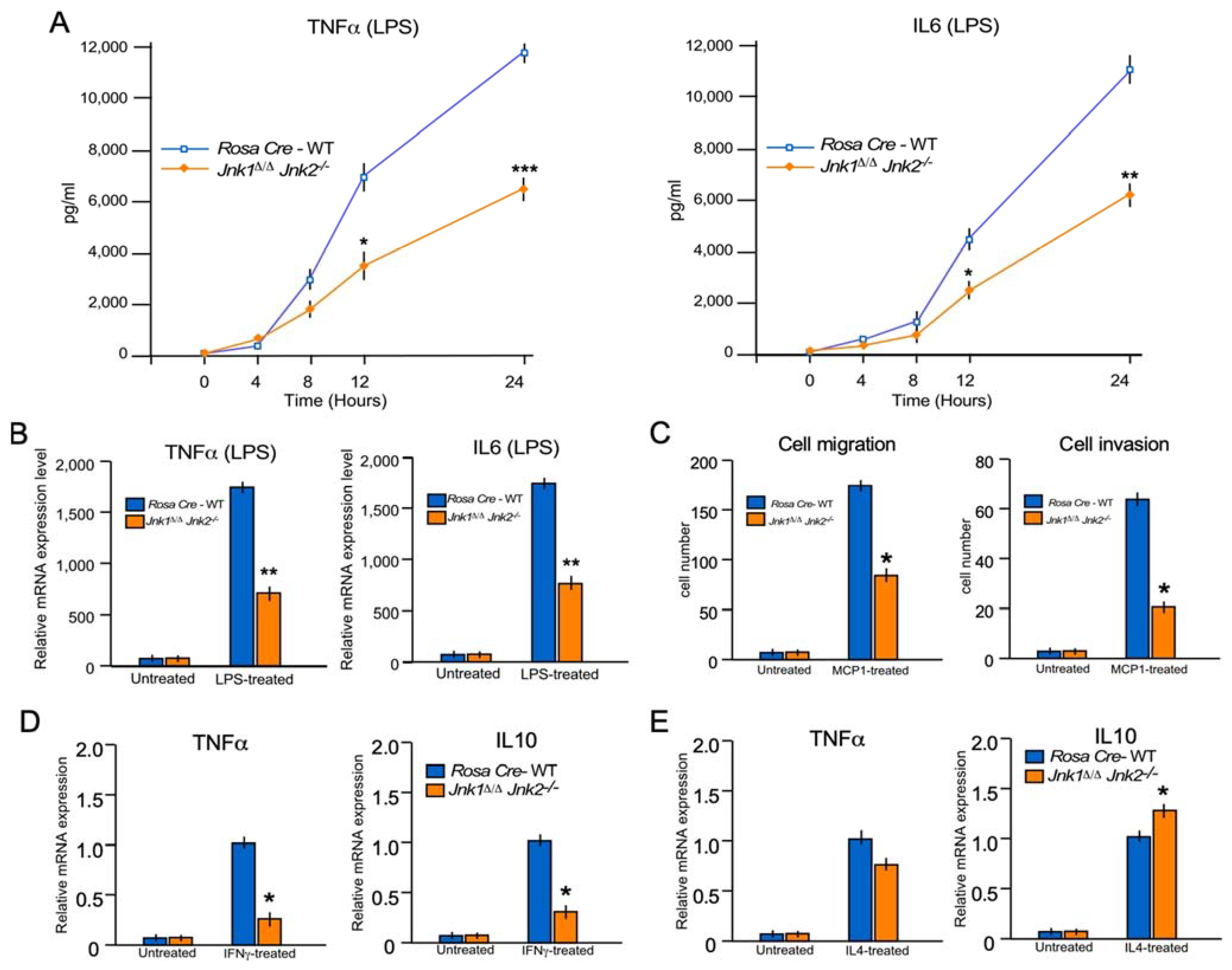
2.4. MKK 7 Controls Cell Migration
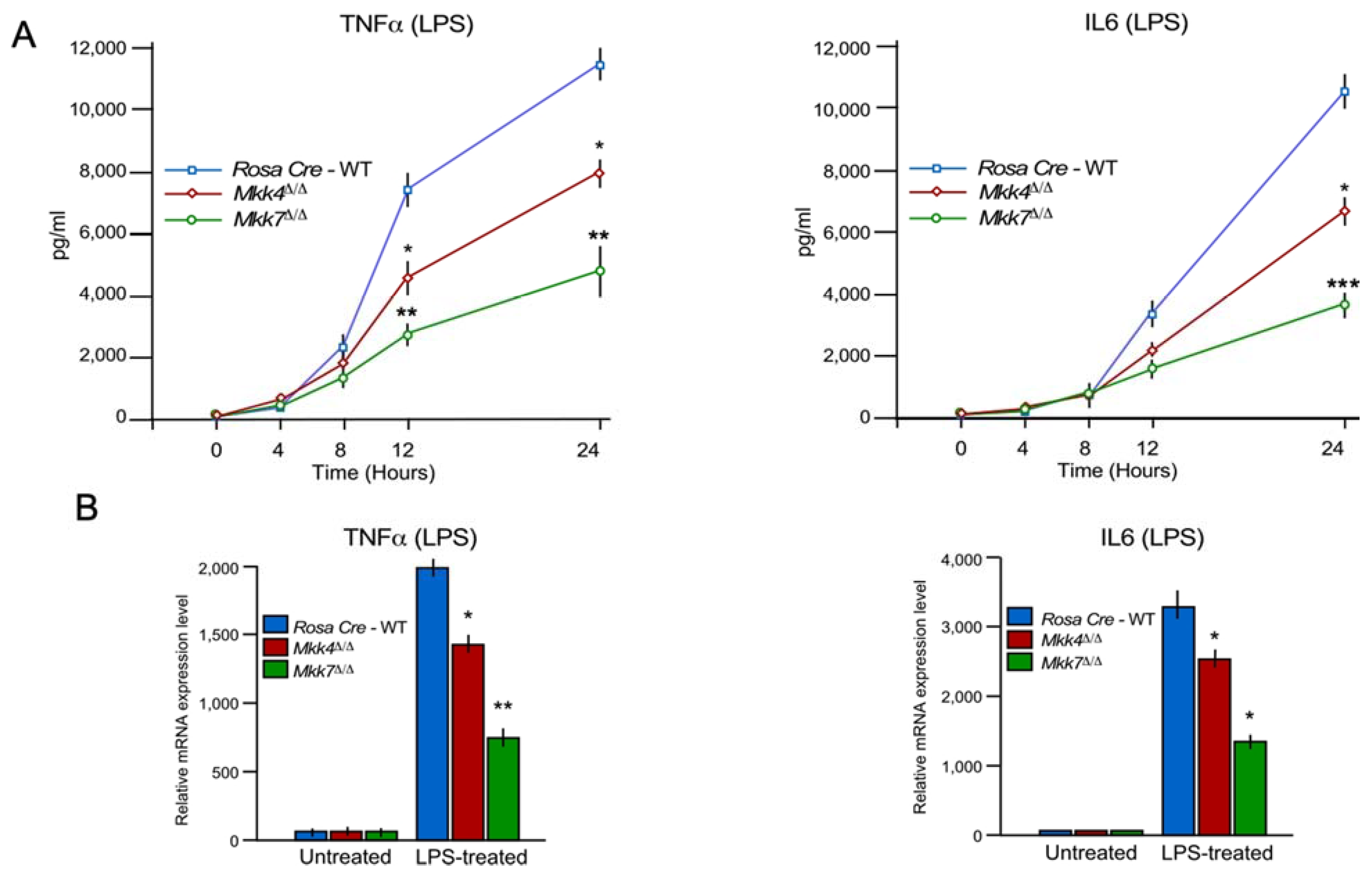
2.5. MKK7 Contributes to Inflammatory Cytokine Production In Vivo
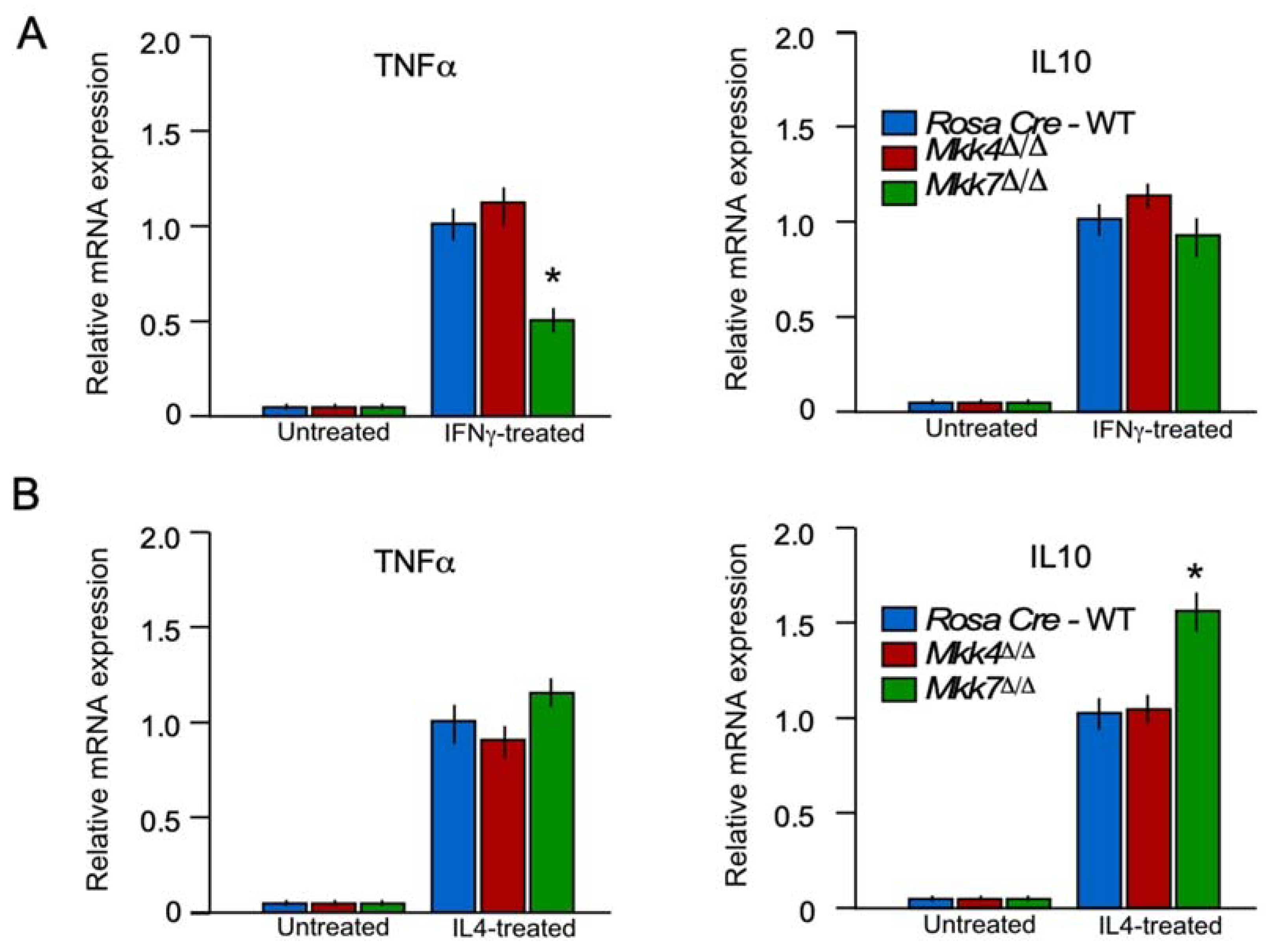
2.6. MKK7 Is Required for M1 Macrophage Polarization
2.7. JNK1/2 Mimics the Phenotype of MKK7 during Inflammatory Cytokine Production In Vitro
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Culture
4.3. Cell Migration and Invasion Assays
4.4. RNA Preparation and Quantitative Polymerase Chain Reaction
4.5. Immunoblot Analysis
4.6. Analysis of Blood
4.7. Antibodies and Reagents
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Dong, C. Regulatory mechanisms of mitogen-activated kinase signaling. Cell Mol. Life. Sci. 2007, 64, 2771–2789. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; Avruch, J. Mammalian MAPK signal transduction pathways activated by stress and inflammation: A 10-year update. Physiol. Rev. 2012, 92, 689–737. [Google Scholar] [CrossRef] [Green Version]
- Davis, R.J. Signal transduction by the JNK group of MAP kinases. Cell 2000, 103, 239–252. [Google Scholar] [CrossRef] [Green Version]
- Craige, S.M.; Reif, M.M.; Kant, S. Mixed—Lineage Protein kinases (MLKs) in inflammation, metabolism, and other disease states. Biochim. Biophys. Acta 2016, 1862, 1581–1586. [Google Scholar] [CrossRef] [PubMed]
- Craige, S.M.; Chen, K.; Blanton, R.M.; Keaney, J.F., Jr.; Kant, S. JNK and cardiometabolic dysfunction. Biosci. Rep. 2019, 39, BSR20190267. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Destrument, A.; Tournier, C. Physiological roles of MKK4 and MKK7: Insights from animal models. Biochim. Biophys. Acta 2007, 1773, 1349–1357. [Google Scholar] [CrossRef] [PubMed]
- Burton, J.C.; Antoniades, W.; Okalova, J.; Roos, M.M.; Grimsey, N.J. Atypical p38 Signaling, Activation, and Implications for Disease. Int. J. Mol. Sci. 2021, 22, 4183. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Tournier, C.; Wysk, M.; Lu, H.T.; Xu, J.; Davis, R.J.; Flavell, R.A. Targeted disruption of the MKK4 gene causes embryonic death, inhibition of c-Jun NH2-terminal kinase activation, and defects in AP-1 transcriptional activity. Proc. Natl. Acad. Sci. USA 1997, 94, 3004–3009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brancho, D.; Tanaka, N.; Jaeschke, A.; Ventura, J.J.; Kelkar, N.; Tanaka, Y.; Kyuuma, M.; Takeshita, T.; Flavell, R.A.; Davis, R.J. Mechanism of p38 MAP kinase activation in vivo. Genes Dev. 2003, 17, 1969–1978. [Google Scholar] [CrossRef] [Green Version]
- Cuenda, A.; Rousseau, S. p38 MAP-kinases pathway regulation, function and role in human diseases. Biochim. Biophys. Acta 2007, 1773, 1358–1375. [Google Scholar] [CrossRef] [Green Version]
- Tournier, C.; Dong, C.; Turner, T.K.; Jones, S.N.; Flavell, R.A.; Davis, R.J. MKK7 is an essential component of the JNK signal transduction pathway activated by proinflammatory cytokines. Genes Dev. 2001, 15, 1419–1426. [Google Scholar] [CrossRef] [Green Version]
- Hubner, A.; Mulholland, D.J.; Standen, C.L.; Karasarides, M.; Cavanagh-Kyros, J.; Barrett, T.; Chi, H.; Greiner, D.L.; Tournier, C.; Sawyers, C.L.; et al. JNK and PTEN cooperatively control the development of invasive adenocarcinoma of the prostate. Proc. Natl. Acad. Sci. USA 2012, 109, 12046–12051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kant, S.; Standen, C.L.; Morel, C.; Jung, D.Y.; Kim, J.K.; Swat, W.; Flavell, R.A.; Davis, R.J. A Protein Scaffold Coordinates SRC-Mediated JNK Activation in Response to Metabolic Stress. Cell Rep. 2017, 20, 2775–2783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, N.C.; Rise, M.L.; Christian, S.L. A Comparison of the Innate and Adaptive Immune Systems in Cartilaginous Fish, Ray-Finned Fish, and Lobe-Finned Fish. Front. Immunol. 2019, 10, 2292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canovas, B.; Nebreda, A.R. Diversity and versatility of p38 kinase signalling in health and disease. Nat. Rev. Mol. Cell Biol. 2021, 22, 346–366. [Google Scholar] [CrossRef]
- Kant, S.; Swat, W.; Zhang, S.; Zhang, Z.Y.; Neel, B.G.; Flavell, R.A.; Davis, R.J. TNF-stimulated MAP kinase activation mediated by a Rho family GTPase signaling pathway. Genes Dev. 2011, 25, 2069–2078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabio, G.; Davis, R.J. TNF and MAP kinase signalling pathways. Semin. Immunol. 2014, 26, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Jaeschke, A.; Davis, R.J. Metabolic stress signaling mediated by mixed-lineage kinases. Mol. Cell 2007, 27, 498–508. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Nadarajah, B.; Robinson, A.C.; McColl, B.W.; Jin, J.W.; Dajas-Bailador, F.; Boot-Handford, R.P.; Tournier, C. Targeted deletion of the mitogen-activated protein kinase kinase 4 gene in the nervous system causes severe brain developmental defects and premature death. Mol. Cell Biol. 2007, 27, 7935–7946. [Google Scholar] [CrossRef] [Green Version]
- Atri, C.; Guerfali, F.Z.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 1801. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.C.; Zou, X.B.; Chai, Y.F.; Yao, Y.M. Macrophage polarization in inflammatory diseases. Int. J. Biol. Sci. 2014, 10, 520–529. [Google Scholar] [CrossRef]
- Nishina, H.; Fischer, K.D.; Radvanyi, L.; Shahinian, A.; Hakem, R.; Rubie, E.A.; Bernstein, A.; Mak, T.W.; Woodgett, J.R.; Penninger, J.M. Stress-signalling kinase Sek1 protects thymocytes from apoptosis mediated by CD95 and CD3. Nature 1997, 385, 350–353. [Google Scholar] [CrossRef] [PubMed]
- Justiz Vaillant, A.A.; Sabir, S.; Jan, A. Physiology, Immune Response. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Taniguchi, K.; Karin, M. NF-kappaB, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 2018, 18, 309–324. [Google Scholar] [CrossRef]
- Tournier, C.; Whitmarsh, A.J.; Cavanagh, J.; Barrett, T.; Davis, R.J. Mitogen-activated protein kinase kinase 7 is an activator of the c-Jun NH2-terminal kinase. Proc. Natl. Acad. Sci. USA 1997, 94, 7337–7342. [Google Scholar] [CrossRef] [Green Version]
- Gallo, K.A.; Johnson, G.L. Mixed-lineage kinase control of JNK and p38 MAPK pathways. Nat. Rev. Mol. Cell Biol. 2002, 3, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Das, M.; Sabio, G.; Jiang, F.; Rincon, M.; Flavell, R.A.; Davis, R.J. Induction of hepatitis by JNK-mediated expression of TNF-alpha. Cell 2009, 136, 249–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Justus, C.R.; Leffler, N.; Ruiz-Echevarria, M.; Yang, L.V. In vitro cell migration and invasion assays. J. Vis. Exp. 2014, 752, 10–24. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2^(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- Kant, S.; Barrett, T.; Vertii, A.; Noh, Y.H.; Jung, D.Y.; Kim, J.K.; Davis, R.J. Role of the mixed-lineage protein kinase pathway in the metabolic stress response to obesity. Cell Rep. 2013, 4, 681–688. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caliz, A.D.; Yoo, H.-J.; Vertii, A.; Dolan, A.C.; Tournier, C.; Davis, R.J.; Keaney, J.F., Jr.; Kant, S. Mitogen Kinase Kinase (MKK7) Controls Cytokine Production In Vitro and In Vivo in Mice. Int. J. Mol. Sci. 2021, 22, 9364. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179364
Caliz AD, Yoo H-J, Vertii A, Dolan AC, Tournier C, Davis RJ, Keaney JF Jr., Kant S. Mitogen Kinase Kinase (MKK7) Controls Cytokine Production In Vitro and In Vivo in Mice. International Journal of Molecular Sciences. 2021; 22(17):9364. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179364
Chicago/Turabian StyleCaliz, Amada D., Hyung-Jin Yoo, Anastassiia Vertii, Ana C. Dolan, Cathy Tournier, Roger J. Davis, John F. Keaney, Jr., and Shashi Kant. 2021. "Mitogen Kinase Kinase (MKK7) Controls Cytokine Production In Vitro and In Vivo in Mice" International Journal of Molecular Sciences 22, no. 17: 9364. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179364