
Long-Term Hypoxia Maintains a State of Dedifferentiation and Enhanced Stemness in Fetal Cardiovascular Progenitor Cells


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
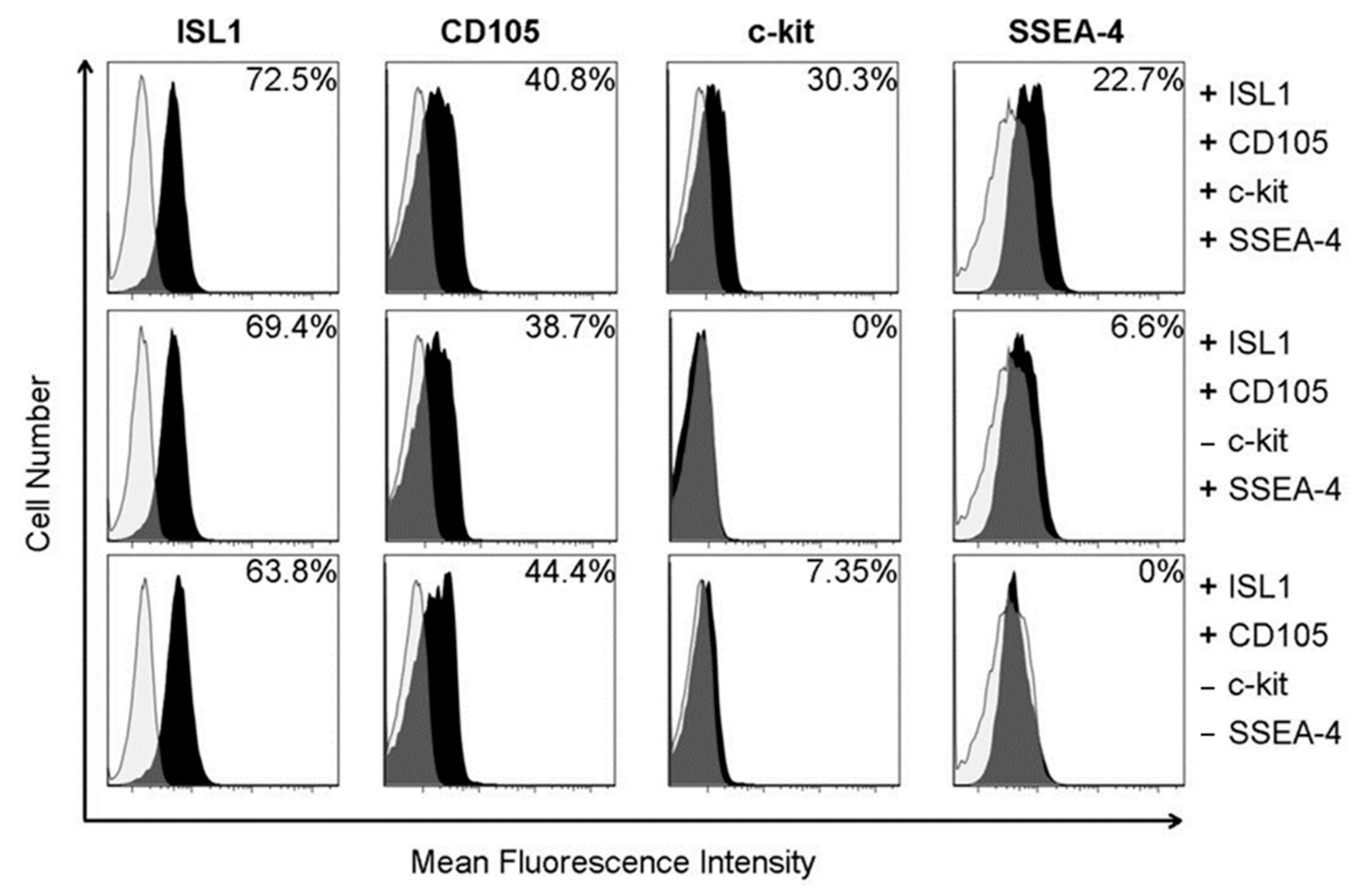
2.1. Cardiovascular Progenitor Cell Characterization
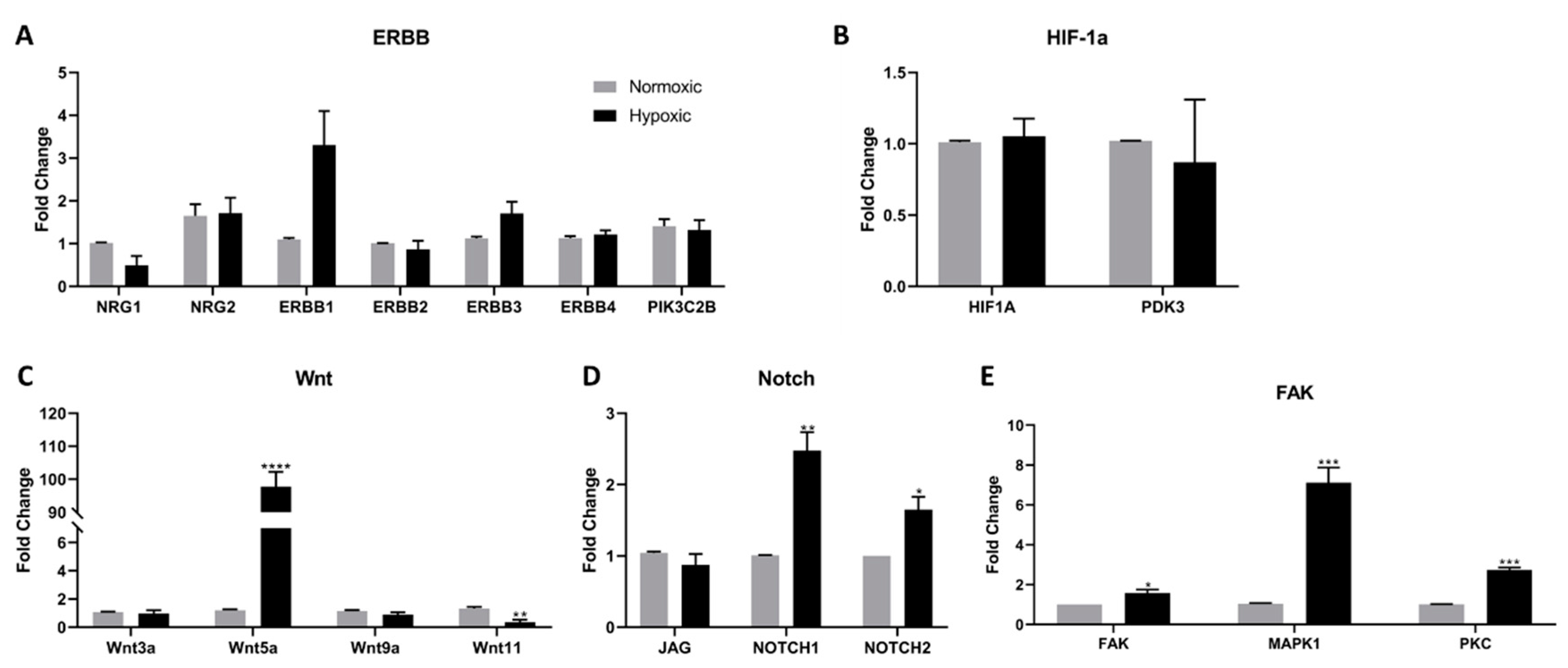
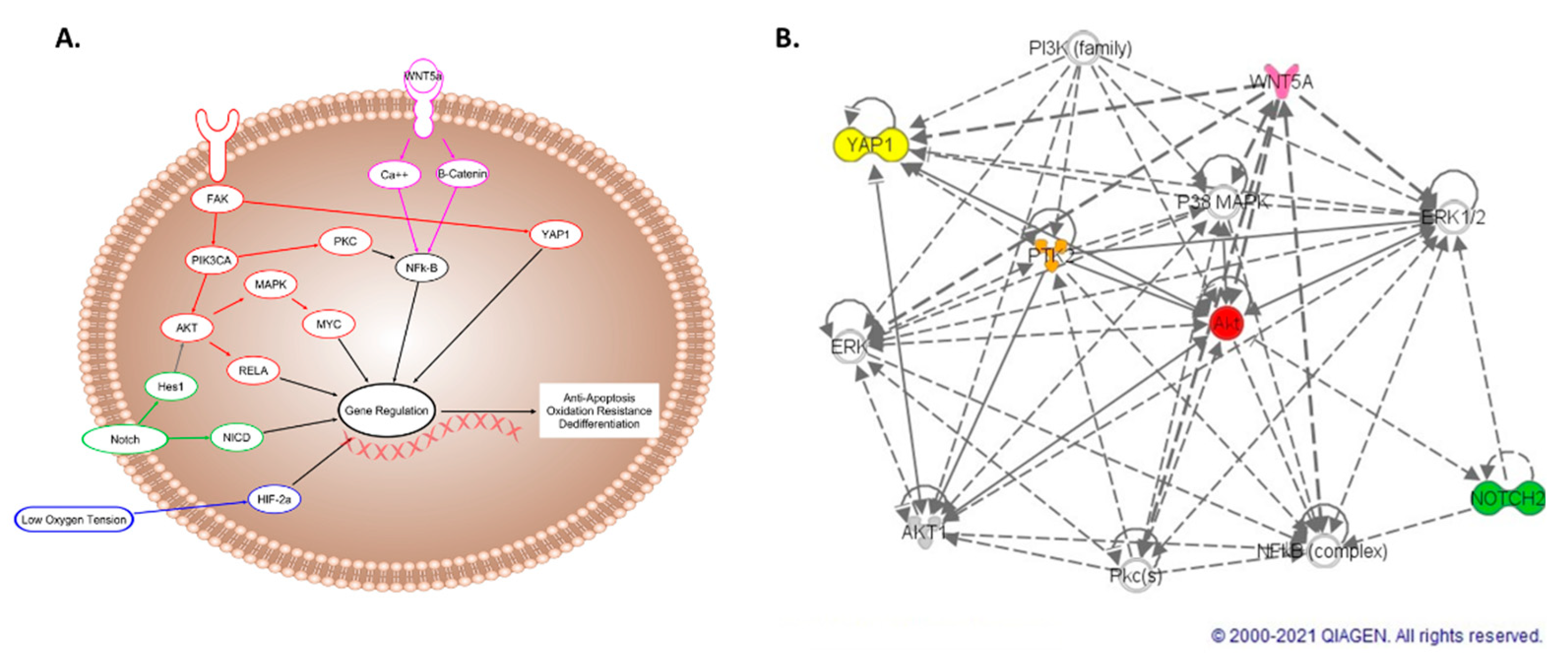
2.2. Wnt, Notch, and FAK Function Synergistically to Promote Cell Survival
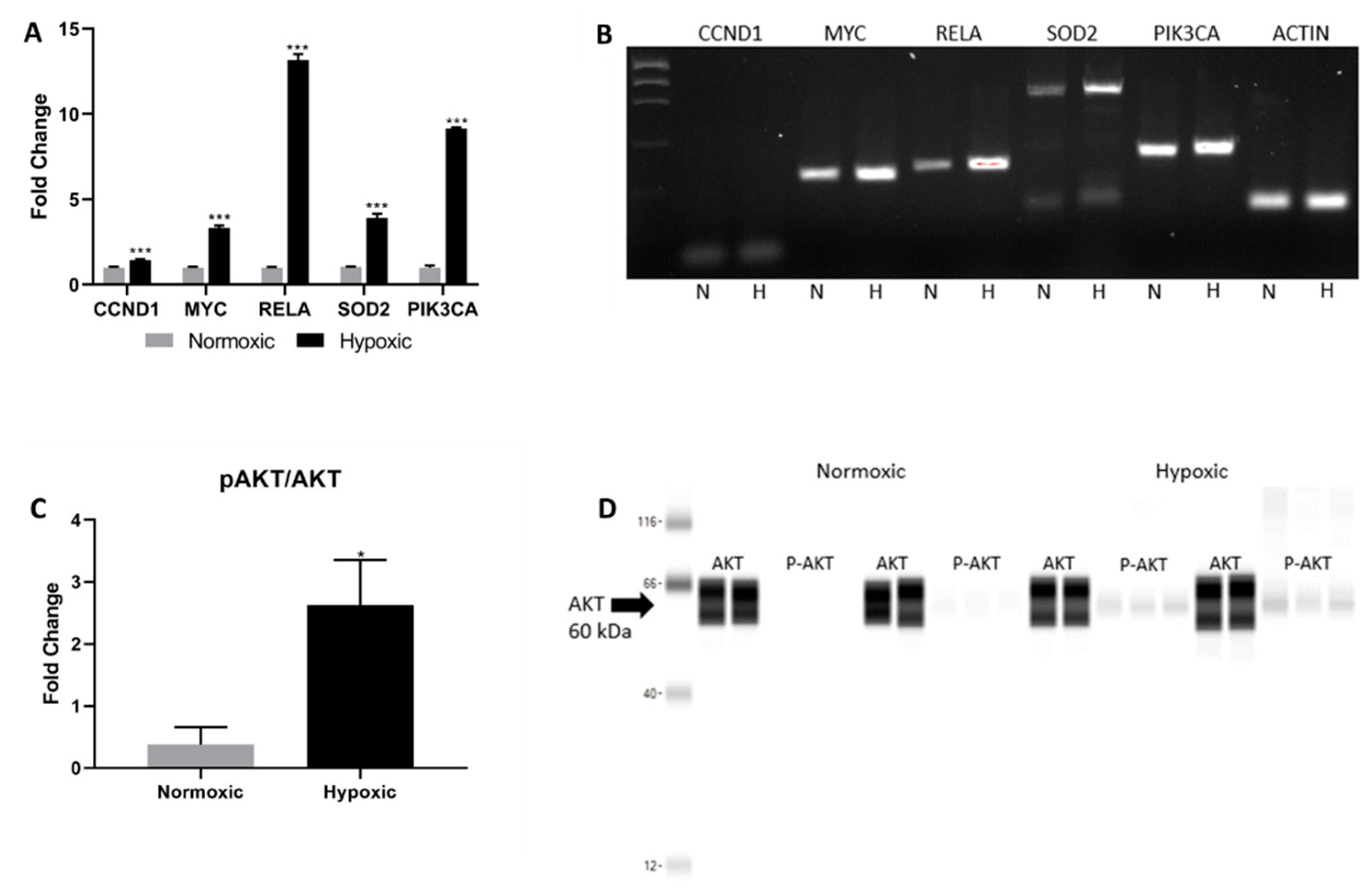
2.3. FAK and Notch Directly Upregulate PI3K/AKT in the Hypoxic Niche
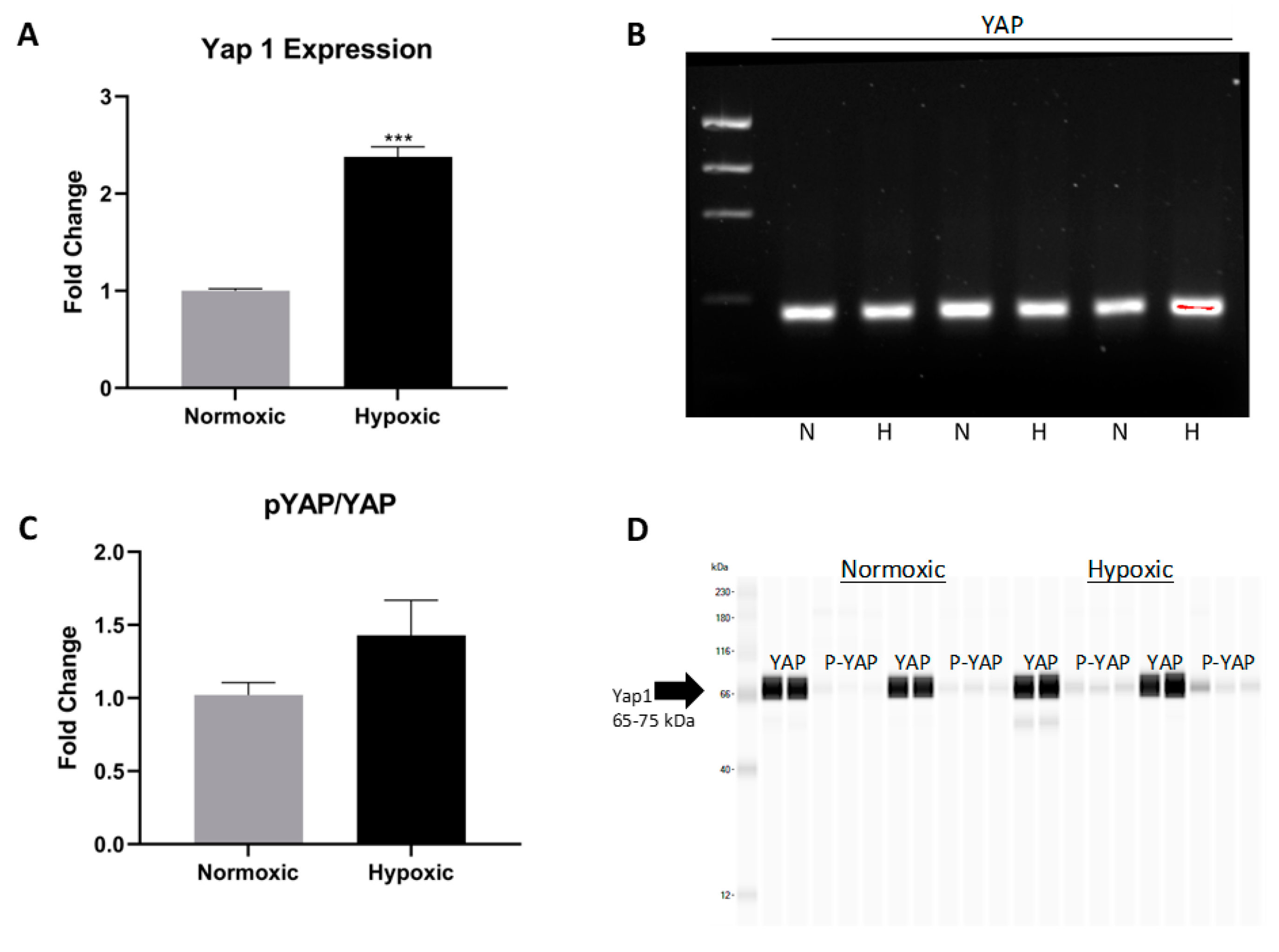
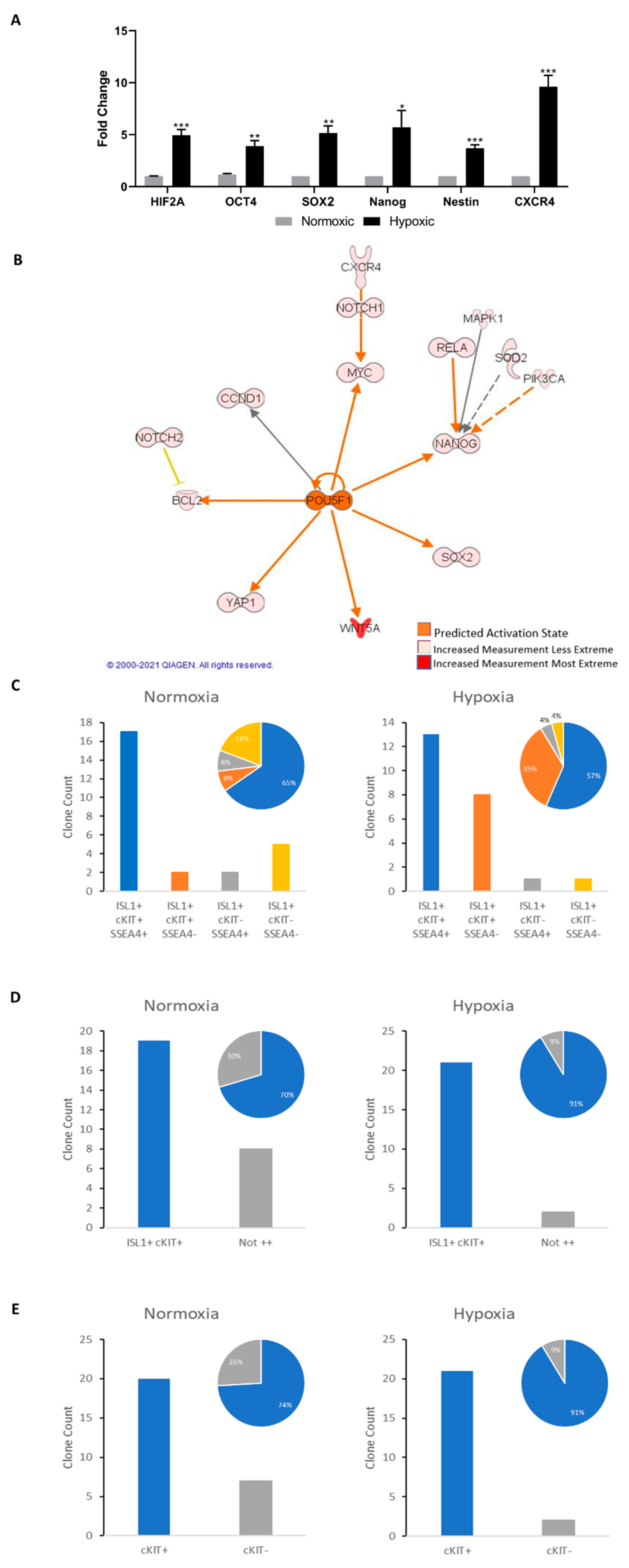
2.4. Islet-1+ Cardiovascular Progenitor Cells Remain Dedifferentiated under Long-Term Hypoxic Conditions
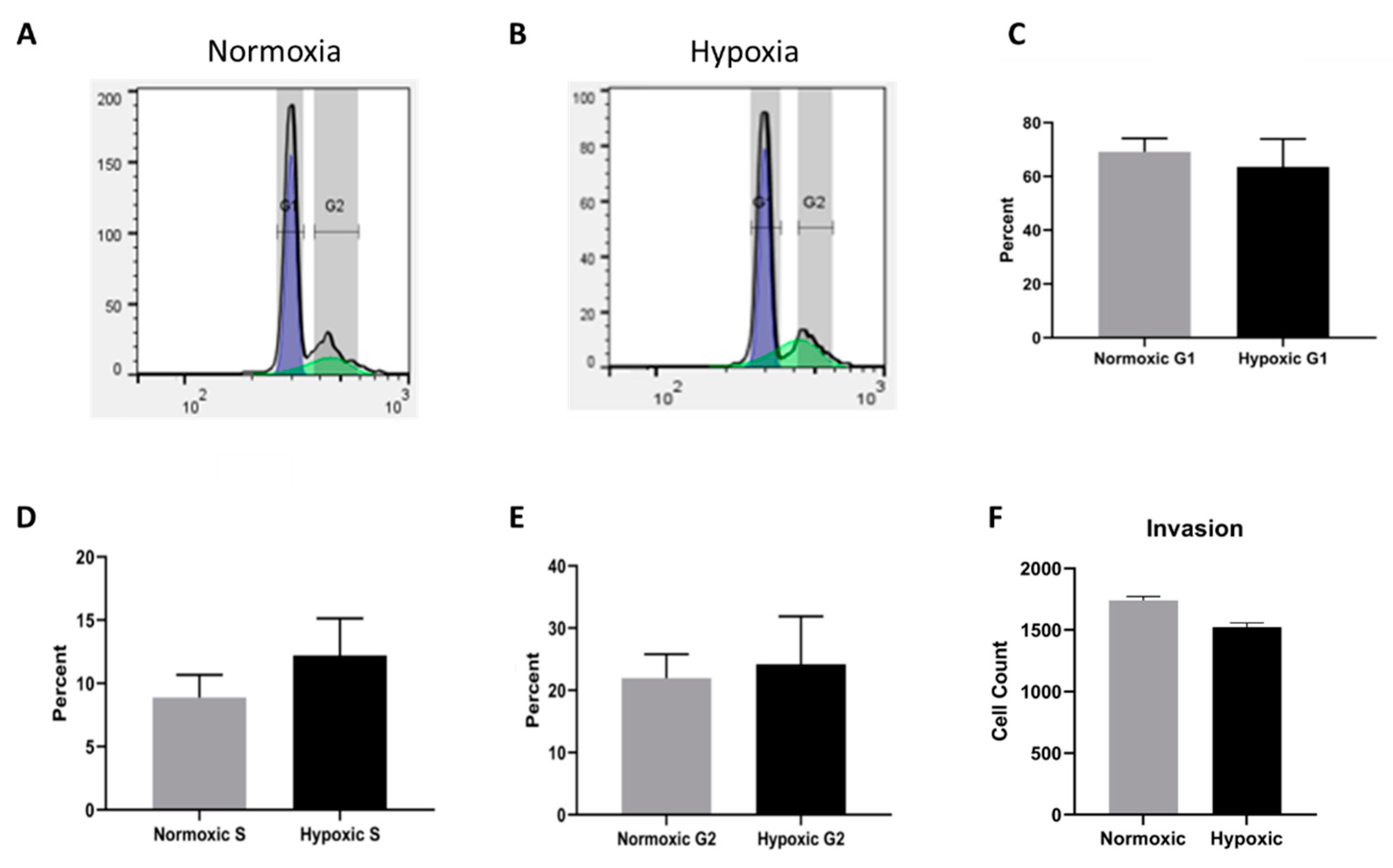
2.5. Cell Cycle Progression and Migration Are Unaffected by Long Term Hypoxia
3. Discussion
4. Materials and Methods
4.1. Animal Selection
4.2. Cell Isolation
4.3. Hypoxic Cell Culture
4.4. Quantitative RT-PCR
4.5. Western Blot
4.6. Flow Cytometry
4.7. Cell Cycle
4.8. Transwell Invasion Assay
4.9. Ingenuity Pathway Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AKT | Ak Strand Transforming |
| BCL2BSA | B-Cell Lymphoma 2Bovine Serum Albumin |
| CCND-1 | Cyclin D1 |
| c-Kit | KIT Proto-Oncogene: Receptor Tyrosine Kinase |
| CPC | Cardiovascular Progenitor Cell |
| CXCR4 | CXC Chemokine Receptor 4 |
| DNA | Deoxyribonucleic Acid |
| EDTA | Ethylenediamine Tetra-Acetic Acid |
| ERBB | Epidermal Growth Factor |
| FAK | Focal Adhesion Kinase |
| HES1 | Hes Family BHLB Transcription Factor 1 |
| HIF-1 | Hypoxia-Inducible Factor 1 |
| HIF-2 | Hypoxia-Inducible Factor 2 |
| HMOX | Heme Oxygenase |
| IPA | Ingenuity Pathway Analysis |
| IRB | Institutional Review Board |
| ISL-1 | Islet-1 |
| MAPK | Mitogen Activated Protein Kinase |
| MYC | MYC Proto-Oncogene |
| Nanog | Nanog Homeobox |
| NFKB | Nuclear Factor Kappa Light Chain Enhance of Activated B Cells |
| NRG | Neuregulin |
| OCT4 | Octamer-Binding Transcription Factor 4 |
| PBS | Phosphate Buffered Saline |
| PDK | Pyruvate Dehydrogenase Kinase |
| PIK3C2B | Phosphatidylinositol-4-Phosphate 3-Kinase C2 Domain Containing Beta Polypeptide |
| PIK3CA | Phosphatidylinositol-4,5-Bisphosphonate 3-Kinase Catalytic Subunit Alpha |
| PKCPOU5F1 | Protein Kinase CPou Domain, Class 5, Transcription Factor 1 |
| RELA | V-Rel Avian Reticuloendotheliosis Viral Oncogene Homolog A |
| RIPA | Radioimmunoprecipitation Assay |
| RNA | Ribonucleic Acid |
| RT-PCR | Reverse Transcription Polymerase Chain Reaction |
| SDF-1 | Stromal Cell Derived Factor 1 |
| SOD2 | Superoxide Dismutase 2 |
| SOX2 | SRY-Related HMG Box |
| SSEA-4 | Stage Specific Embryonic Antigen 4 |
| Wnt | Wingless Tyrosine Kinase |
| Yap1 | Yes1 Associated Transcriptional Regulator |
References
- Barreto, S.; Hamel, L.; Schiatti, T.; Yang, Y.; George, V. Cardiac Progenitor Cells from Stem Cells: Learning from Genetics and Biomaterials. Cells 2019, 8, 1536. [Google Scholar] [CrossRef] [Green Version]
- Duran, A.; Reidell, O.; Stachelscheid, H.; Klose, K.; Gossen, M.; Falk, V.; Röll, W.; Stamm, C. Regenerative Medicine/Cardiac Cell Therapy: Pluripotent Stem Cells. Thorac. Cardiovasc. Surg. 2018, 66, 53–62. [Google Scholar] [CrossRef]
- Hernandez, I.; Baio, J.M.; Tsay, E.; Martinez, A.F.; Fuentes, T.I.; Bailey, L.L.; Hasaniya, N.W.; Kearns-Jonker, M. Short-Term Hypoxia Improves Early Cardiac Progenitor Cell Function in Vitro. Am. J. Stem. Cells 2018, 7, 1–17. [Google Scholar]
- Hu, J.; Wang, Y.; Jiao, J.; Liu, Z.; Zhao, C.; Zhou, Z.; Zhang, Z.; Forde, K.; Wang, L.; Wang, J.; et al. Patient-Specific Cardiovascular Progenitor Cells Derived from Integration-Free Induced Pluripotent Stem Cells for Vascular Tissue Regeneration. Biomaterials 2015, 73, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Ezashi, T.; Das, P.; Roberts, R.M. Low O2 Tensions and the Prevention of Differentiation of HES Cells. Proc. Natl. Acad. Sci. USA 2005, 102, 4783–4788. [Google Scholar] [CrossRef] [Green Version]
- Petruzzelli, R.; Christensen, D.R.; Parry, K.L.; Sanchez-Elsner, T.; Houghton, F.D. HIF-2α Regulates NANOG Expression in Human Embryonic Stem Cells Following Hypoxia and Reoxygenation through the Interaction with an Oct-Sox Cis Regulatory Element. PLoS ONE 2014, 9, e108309. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Ding, R.; Ha, Y.; Jia, Y.; Liao, X.; Wang, S.; Li, R.; Shen, Z.; Xiong, H.; Guo, J.; et al. Hypoxia-Stressed Cardiomyocytes Promote Early Cardiac Differentiation of Cardiac Stem Cells through HIF-1α/Jagged1/Notch1 Signaling. Acta Pharm. Sinica. B 2018, 8, 795–804. [Google Scholar] [CrossRef]
- Korski, K.I.; Kubli, D.A.; Wang, B.J.; Khalafalla, F.G.; Monsanto, M.M.; Firouzi, F.; Echeagaray, O.H.; Kim, T.; Adamson, R.M.; Dembitsky, W.P.; et al. Hypoxia Prevents Mitochondrial Dysfunction and Senescence in Human C-Kit + Cardiac Progenitor Cells: Hypoxia Blunts Senescence of Cardiac Stem Cells. Stem. Cells 2019, 37, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Gersten, M.; Zhou, D.; Azad, P.; Haddad, G.G.; Subramaniam, S. Wnt Pathway Activation Increases Hypoxia Tolerance during Development. PLoS ONE 2014, 9, e103292. [Google Scholar] [CrossRef]
- Huang, X.; Trinh, T.; Aljoufi, A.; Broxmeyer, H.E. Hypoxia Signaling Pathway in Stem Cell Regulation: Good and Evil. Curr. Stem. Cell Rep. 2018, 4, 149–157. [Google Scholar] [CrossRef]
- Hou, J.; Wang, L.; Long, H.; Wu, H.; Wu, Q.; Zhong, T.; Chen, X.; Zhou, C.; Guo, T.; Wang, T. Hypoxia Preconditioning Promotes Cardiac Stem Cell Survival and Cardiogenic Differentiation in Vitro Involving Activation of the HIF-1α/Apelin/APJ Axis. Stem. Cell Res. Ther. 2017, 8, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, A.I.; Mallon, B.S.; Matsui, T.; Ogawa, W.; Rosenzweig, A.; Okamoto, T.; Macklin, W.B. Akt-Mediated Survival of Oligodendrocytes Induced by Neuregulins. J. Neurosci. 2000, 20, 7622–7630. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.W.; Schoeberl, B.; Jasper, P.J.; Niepel, M.; Nielsen, U.B.; Lauffenburger, D.A.; Sorger, P.K. Input–Output Behavior of ErbB Signaling Pathways as Revealed by a Mass Action Model Trained against Dynamic Data. Mol. Syst. Biol. 2009, 5, 239. [Google Scholar] [CrossRef]
- Foulquier, S.; Daskalopoulos, E.P.; Lluri, G.; Hermans, K.C.M.; Deb, A.; Blankesteijn, W.M. WNT Signaling in Cardiac and Vascular Disease. Pharmacol. Rev. 2018, 70, 68–141. [Google Scholar] [CrossRef]
- Mohammed, M.K.; Shao, C.; Wang, J.; Wei, Q.; Wang, X.; Collier, Z.; Tang, S.; Liu, H.; Zhang, F.; Huang, J.; et al. Wnt/β-Catenin Signaling Plays an Ever-Expanding Role in Stem Cell Self-Renewal, Tumorigenesis and Cancer Chemoresistance. Genes Dis. 2016, 3, 11–40. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, B. Wnt/β-Catenin Signaling Pathway Governs a Full Program for Dopaminergic Neuron Survival, Neurorescue and Regeneration in the MPTP Mouse Model of Parkinson’s Disease. IJMS 2018, 19, 3743. [Google Scholar] [CrossRef] [Green Version]
- Manning, B.D.; Toker, A. AKT/PKB Signaling: Navigating the Network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef] [Green Version]
- Abeyrathna, P.; Su, Y. The Critical Role of Akt in Cardiovascular Function. Vasc. Pharmacol. 2015, 74, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhang, W.; Wang, Y.; Zhao, D.; Wu, M.; Fan, J.; Li, J.; Gong, Y.; Dan, N.; Yang, D.; et al. The Diacylglycerol Kinase α (DGKα)/Akt/NF-ΚB Feedforward Loop Promotes Esophageal Squamous Cell Carcinoma (ESCC) Progression via FAK-Dependent and FAK-Independent Manner. Oncogene 2019, 38, 2533–2550. [Google Scholar] [CrossRef]
- Kozlovskaja-Gumbrienė, A.; Yi, R.; Alexander, R.; Aman, A.; Jiskra, R.; Nagelberg, D.; Knaut, H.; McClain, M.; Piotrowski, T. Proliferation-Independent Regulation of Organ Size by Fgf/Notch Signaling. eLife 2017, 6, e21049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, P.; Mele, D.; Caliceti, C.; Pannella, M.; Fortini, C.; Clementz, A.G.; Morelli, M.B.; Aquila, G.; Ameri, P.; Ferrari, R. The Role of Notch in the Cardiovascular System: Potential Adverse Effects of Investigational Notch Inhibitors. Front. Oncol. 2015, 4. [Google Scholar] [CrossRef] [Green Version]
- Prasad, S.M.; Czepiel, M.; Cetinkaya, C.; Smigielska, K.; Weli, S.C.; Lysdahl, H.; Gabrielsen, A.; Petersen, K.; Ehlers, N.; Fink, T.; et al. Continuous Hypoxic Culturing Maintains Activation of Notch and Allows Long-Term Propagation of Human Embryonic Stem Cells without Spontaneous Differentiation. Cell Prolif. 2009, 42, 63–74. [Google Scholar] [CrossRef]
- Palomero, T.; Dominguez, M.; Ferrando, A.A. The Role of the PTEN/AKT Pathway in NOTCH1-Induced Leukemia. Cell Cycle 2008, 7, 965–970. [Google Scholar] [CrossRef]
- Zheng, Y.; Pan, D. The Hippo Signaling Pathway in Development and Disease. Dev. Cell 2019, 50, 264–282. [Google Scholar] [CrossRef]
- Misra, J.R.; Irvine, K.D. The Hippo Signaling Network and Its Biological Functions. Annu. Rev. Genet. 2018, 52, 65–87. [Google Scholar] [CrossRef]
- Camberos, V.; Baio, J.; Bailey, L.; Hasaniya, N.; Lopez, L.V.; Kearns-Jonker, M. Effects of Spaceflight and Simulated Microgravity on YAP1 Expression in Cardiovascular Progenitors: Implications for Cell-Based Repair. IJMS 2019, 20, 2742. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Li, X.; He, Y.; Li, W.; Wang, Y.; Wang, H.; Jiang, S.; Xin, Y. YAP1 enhances cell proliferation, migration, and invasion of gastric cancer in vitro and in vivo. Oncotarget 2016, 7, 81062–81076. [Google Scholar] [CrossRef] [Green Version]
- Koo, J.H.; Plouffe, S.W.; Meng, Z.; Lee, D.H.; Yang, D.; Lim, D.S.; Wang, C.Y.; Guan, K.L. Induction of AP-1 by YAP/TAZ contributes to cell proliferation and organ growth. Genes Dev. 2020, 34, 72–86. [Google Scholar] [CrossRef]
- Kanazawa, H.; Imoto, K.; Okada, M.; Yamawaki, H. Canstatin Inhibits Hypoxia-Induced Apoptosis through Activation of Integrin/Focal Adhesion Kinase/Akt Signaling Pathway in H9c2 Cardiomyoblasts. PLoS ONE 2017, 12, e0173051. [Google Scholar] [CrossRef] [Green Version]
- Borreguero-Muñoz, N.; Fletcher, G.C.; Aguilar-Aragon, M.; Elbediwy, A.; Vincent-Mistiaen, Z.I.; Thompson, B.J. The Hippo Pathway Integrates PI3K–Akt Signals with Mechanical and Polarity Cues to Control Tissue Growth. PLoS Biol. 2019, 17, e3000509. [Google Scholar] [CrossRef] [PubMed]
- Forristal, C.E.; Wright, K.L.; Hanley, N.A.; Oreffo, R.O.C.; Houghton, F.D. Hypoxia Inducible Factors Regulate Pluripotency and Proliferation in Human Embryonic Stem Cells Cultured at Reduced Oxygen Tensions. Reproduction 2010, 139, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, P.; Chandel, N.S.; Simon, M.C. Cellular Adaptation to Hypoxia through Hypoxia Inducible Factors and Beyond. Nat. Rev. Mol. Cell Biol. 2020, 21, 268–283. [Google Scholar] [CrossRef]
- Madl, C.M.; Le Savage, B.L.; Dewi, R.E.; Dinh, C.B.; Stowers, R.S.; Khariton, M.; Lampe, K.J.; Nguyen, D.; Chaudhuri, O.; Enejder, A.; et al. Maintenance of Neural Progenitor Cell Stemness in 3D Hydrogels Requires Matrix Remodelling. Nat. Mater 2017, 16, 1233–1242. [Google Scholar] [CrossRef] [Green Version]
- Kang, N.; Choi, S.Y.; Kim, B.N.; Yeo, C.D.; Park, C.K.; Kim, Y.K.; Kim, T.-J.; Lee, S.-B.; Lee, S.H.; Park, J.Y.; et al. Hypoxia-Induced Cancer Stemness Acquisition Is Associated with CXCR4 Activation by Its Aberrant Promoter Demethylation. BMC Cancer 2019, 19, 148. [Google Scholar] [CrossRef] [Green Version]
- Truong, T.T.; Huynh, K.; Nakatsu, M.N.; Deng, S.X. SSEA4 Is a Potential Negative Marker for the Enrichment of Human Corneal Epithelial Stem/Progenitor Cells. Invest. Ophthalmol. Vis. Sci. 2011, 52, 6315. [Google Scholar] [CrossRef]
- Lee, S.H.; Hyeon, D.Y.; Yoon, S.H.; Han, S.M.; Jang, J.W.; Nguyen, M.P.; Chi, X.Z.; An, S.; Hyun, K.G.; Jung, H.J.; et al. RUNX3 methylation drives hypoxia-induced cell proliferation and antiapoptosis in early tumorigenesis. Cell Death Differ. 2021, 28, 1251–1269. [Google Scholar] [CrossRef] [PubMed]
- Shorning, B.Y.; Dass, M.S.; Smalley, M.J.; Pearson, H.B. The PI3K-AKT-MTOR Pathway and Prostate Cancer: At the Crossroads of AR, MAPK, and WNT Signaling. IJMS 2020, 21, 4507. [Google Scholar] [CrossRef]
- Feng, Y.; Liang, Y.; Zhu, X.; Wang, M.; Gui, Y.; Lu, Q.; Gu, M.; Xue, X.; Sun, X.; He, W.; et al. The signaling protein Wnt5a promotes TGFβ1-mediated macrophage polarization and kidney fibrosis by inducing the transcriptional regulators Yap/Taz. J. Biol. Chem. 2018, 293, 19290–19302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, N.G.; Gumbiner, B.M. Adhesion to fibronectin regulates Hippo signaling via the FAK-Src-PI3K pathway. J. Cell Biol. 2015, 210, 503–515. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, L.A.; Squatrito, M.; Northcott, P.; Awan, A.; Holland, E.C.; Taylor, M.D.; Nahlé, Z.; Kenney, A.M. Oncogenic YAP promotes radioresistance and genomic instability in medulloblastoma through IGF2-mediated Akt activation. Oncogene 2012, 31, 1923–1937. [Google Scholar] [CrossRef] [Green Version]
- Kang, W.; Tong, J.H.; Chan, A.W.; Lee, T.L.; Lung, R.W.; Leung, P.P.; So, K.K.; Wu, K.; Fan, D.; Yu, J.; et al. Yes-associated protein 1 exhibits oncogenic property in gastric cancer and its nuclear accumulation associates with poor prognosis. Clin. Cancer Res. 2011, 17, 2130–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.K.; Yonehara, S. Identification of mechanism that couples multisite phosphorylation of Yes-associated protein (YAP) with transcriptional coactivation and regulation of apoptosis. J. Biol. Chem. 2012, 287, 9568–9578. [Google Scholar] [CrossRef] [Green Version]
- Slack-Davis, J.K.; Eblen, S.T.; Zecevic, M.; Boerner, S.A.; Tarcsafalvi, A.; Diaz, H.B.; Marshall, M.S.; Weber, M.J.; Parsons, J.T.; Catling, A.D. PAK1 phosphorylation of MEK1 regulates fibronectin-stimulated MAPK activation. J. Cell Biol. 2003, 162, 281–291. [Google Scholar] [CrossRef] [Green Version]
- Dufour, G.; Demers, M.J.; Gagné, D.; Dydensborg, A.B.; Teller, I.C.; Bouchard, V.; Degongre, I.; Beaulieu, J.F.; Cheng, J.Q.; Fujita, N.; et al. Human intestinal epithelial cell survival and anoikis. Differentiation state-distinct regulation and roles of protein kinase B/Akt isoforms. J. Biol. Chem. 2004, 79, 44113–44122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igishi, T.; Fukuhara, S.; Patel, V.; Katz, B.Z.; Yamada, K.M.; Gutkind, J.S. Divergent signaling pathways link focal adhesion kinase to mitogen-activated protein kinase cascades. Evidence for a role of paxillin in c-Jun NH(2)-terminal kinase activation. J. Biol. Chem. 1999, 274, 30738–30746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Liu, B.; Al-Aidaroos, A.Q.; Shi, H.; Li, L.; Guo, K.; Li, J.; Tan, B.C.; Loo, J.M.; Tang, J.P.; et al. Dual-faced SH3BGRL: Oncogenic in mice, tumor suppressive in humans. Oncogene 2016, 35, 3303–3313. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.H.; Xia, W.; Lee, N.P.; Mruk, D.D.; Lee, W.M.; Cheng, C.Y. Regulation of ectoplasmic specialization dynamics in the seminiferous epithelium by focal adhesion-associated proteins in testosterone-suppressed rat testes. Endocrinology 2005, 146, 1192–1204. [Google Scholar] [CrossRef] [Green Version]
- Aikawa, R.; Nagai, T.; Kudoh, S.; Zou, Y.; Tanaka, M.; Tamura, M.; Akazawa, H.; Takano, H.; Nagai, R.; Komuro, I. Integrins play a critical role in mechanical stress-induced p38 MAPK activation. Hypertension 2002, 39, 233–238. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, Y.; Fritz, D.; Rajshankar, D.; Downey, G.P.; McCulloch, C.A. Interactions of the protein-tyrosine phosphatase-α with the focal adhesion targeting domain of focal adhesion kinase are involved in interleukin-1 signaling in fibroblasts. J. Biol. Chem. 2014, 289, 18427–18441. [Google Scholar] [CrossRef] [Green Version]
- Toruner, M.; Fernandez-Zapico, M.; Sha, J.J.; Pham, L.; Urrutia, R.; Egan, L.J. Antianoikis effect of nuclear factor-kappaB through up-regulated expression of osteoprotegerin, BCL-2, and IAP-1. J. Biol. Chem. 2006, 281, 8686–8696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funakoshi-Tago, M.; Sonoda, Y.; Tanaka, S.; Hashimoto, K.; Tago, K.; Tominaga, S.; Kasahara, T. Tumor necrosis factor-induced nuclear factor kappaB activation is impaired in focal adhesion kinase-deficient fibroblasts. J. Biol. Chem. 2003, 278, 29359–29365. [Google Scholar] [CrossRef] [Green Version]
- Etienne-Manneville, S.; Manneville, J.B.; Adamson, P.; Wilbourn, B.; Greenwood, J.; Couraud, P.O. ICAM-1-coupled cytoskeletal rearrangements and transendothelial lymphocyte migration involve intracellular calcium signaling in brain endothelial cell lines. J. Immunol. 2000, 165, 3375–3383. [Google Scholar] [CrossRef] [Green Version]
- Disatnik, M.H.; Rando, T.A. Integrin-mediated muscle cell spreading. The role of protein kinase c in outside-in and inside-out signaling and evidence of integrin cross-talk. J. Biol. Chem. 1999, 274, 32486–32492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyme, A.; Marivin, A.; Garcia-Marcos, M. GIV/Girdin (Gα-interacting, Vesicle-associated Protein/Girdin) Creates a Positive Feedback Loop That Potentiates Outside-in Integrin Signaling in Cancer Cells. J. Biol. Chem. 2016, 291, 8269–8282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oderup, C.; LaJevic, M.; Butcher, E.C. Canonical and noncanonical Wnt proteins program dendritic cell responses for tolerance. J. Immunol. 2013, 190, 6126–6134. [Google Scholar] [CrossRef] [Green Version]
- Whang, Y.M.; Jo, U.; Sung, J.S.; Ju, H.J.; Kim, H.K.; Park, K.H.; Lee, J.W.; Koh, I.S.; Kim, Y.H. Wnt5a is associated with cigarette smoke-related lung carcinogenesis via protein kinase C. PLoS ONE. 2013, 8, e53012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenei, V.; Sherwood, V.; Howlin, J.; Linnskog, R.; Säfholm, A.; Axelsson, L.; Andersson, T. A t-butyloxycarbonyl-modified Wnt5a-derived hexapeptide functions as a potent antagonist of Wnt5a-dependent melanoma cell invasion. Proc. Natl. Acad. Sci. USA 2009, 106, 19473–19478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.; Choi, M.H.; Chang, M.S.; Baik, J.H. Wnt5a-dopamine D2 receptor interactions regulate dopamine neuron development via extracellular signal-regulated kinase (ERK) activation. J. Biol. Chem. 2011, 286, 15641–15651. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Wen, Q.; Hu, S.; Zhou, X.; Xiong, W.; Du, X.; Zhang, L.; Fu, Y.; Yang, J.; Zhou, C.; et al. IL-36γ Promotes Killing of Mycobacterium tuberculosis by Macrophages via WNT5A-Induced Noncanonical WNT Signaling. J. Immunol. 2019, 203, 922–935. [Google Scholar] [CrossRef]
- Vassallo, I.; Zinn, P.; Lai, M.; Rajakannu, P.; Hamou, M.F.; Hegi, M.E. WIF1 re-expression in glioblastoma inhibits migration through attenuation of non-canonical WNT signaling by downregulating the lncRNA MALAT1. Oncogene 2016, 35, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, T.; Chen, L.; Endo, T.; Tang, L.; Lu, D.; Castro, J.E.; Widhopf, G.F.; Rassenti, L.Z.; Cantwell, M.J.; Prussak, C.E.; et al. Antisera induced by infusions of autologous Ad-CD154-leukemia B cells identify ROR1 as an oncofetal antigen and receptor for Wnt5a. Proc. Natl. Acad. Sci. USA 2008, 105, 3047–3052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, M.; Katoh, M. Transcriptional mechanisms of WNT5A based on NF-kappaB, Hedgehog, TGFbeta, and Notch signaling cascades. Int. J. Mol. Med. 2009, 23, 763–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, A.; Panda, S.; Kudmulwar, D.; Chhatbar, S.P.; Nayak, S.C.; Krishnan, H.H. Hepatitis C virus NS5A binds to the mRNA cap-binding eukaryotic translation initiation 4F (eIF4F) complex and up-regulates host translation initiation machinery through eIF4E-binding protein 1 inactivation. J. Biol. Chem. 2012, 287, 5042–5058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Z.; Zhang, Q.; Feng, Q.; Xu, J.; Teng, T.; Luan, Q.; Shan, C.; Hu, Y.; Hemmings, B.A.; Gao, X.; et al. Deletion of Akt1 causes heart defects and abnormal cardiomyocyte proliferation. Dev. Biol. 2010, 347, 384–391. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Hung, M.C. Regulation of the activity of p38 mitogen-activated protein kinase by Akt in cancer and adenoviral protein E1A-mediated sensitization to apoptosis. Mol. Cell Biol. 2003, 23, 6836–6848. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Ju, J.H.; Lee, K.M.; Shin, I. Akt isoform-specific inhibition of MDA-MB-231 cell proliferation. Cell Signal. 2011, 23, 19–26, Epub 2010 Aug 2. [Google Scholar] [CrossRef]
- Brami-Cherrier, K.; Valjent, E.; Garcia, M.; Pagès, C.; Hipskind, R.A.; Caboche, J. Dopamine induces a PI3-kinase-independent activation of Akt in striatal neurons: A new route to cAMP response element-binding protein phosphorylation. J. Neurosci. 2002, 22, 8911–8921. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.W.; Kim, H.K.; Kim, I.Y.; Yim, M.B.; Chock, P.B. Dual function of protein kinase C (PKC) in 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced manganese superoxide dismutase (MnSOD) expression: Activation of CREB and FOXO3a by PKC-alpha phosphorylation and by PKC-mediated inactivation of Akt, respectively. J. Biol. Chem. 2011, 286, 29681–29690. [Google Scholar]
- Li, Q.; Laumonnier, Y.; Syrovets, T.; Simmet, T. Plasmin triggers cytokine induction in human monocyte-derived macrophages. Arter. Thromb. Vasc. Biol. 2007, 27, 1383–1389. [Google Scholar] [CrossRef]
- Thuille, N.; Heit, I.; Fresser, F.; Krumböck, N.; Bauer, B.; Leuthaeusser, S.; Dammeier, S.; Graham, C.; Copeland, T.D.; Shaw, S.; et al. Critical role of novel Thr-219 autophosphorylation for the cellular function of PKCtheta in T lymphocytes. EMBO J. 2005, 24, 3869–3880. [Google Scholar] [CrossRef] [Green Version]
- Pierau, M.; Na, S.Y.; Simma, N.; Lowinus, T.; Marx, A.; Schraven, B.; Bommhardt, U.H. Constitutive Akt1 signals attenuate B-cell receptor signaling and proliferation, but enhance B-cell migration and effector function. Eur. J. Immunol. 2012, 42, 3381–3393. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, C.; Liu, Z.; Liu, X.; Han, C.; Cao, X.; Li, N. Notch signal suppresses Toll-like receptor-triggered inflammatory responses in macrophages by inhibiting extracellular signal-regulated kinase 1/2-mediated nuclear factor κB activation. J. Biol. Chem. 2012, 287, 6208–6217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-García, A.; López-López, S.; García-Ramírez, J.J.; Baladrón, V.; Ruiz-Hidalgo, M.J.; López-Sanz, L.; Ballesteros, Á.; Laborda, J.; Monsalve, E.M.; Díaz-Guerra, M.J. The Tetraspanin TSPAN33 Controls TLR-Triggered Macrophage Activation through Modulation of NOTCH Signaling. J. Immunol. 2016, 197, 3371–3381. [Google Scholar] [CrossRef]
- Xu, M.; Wang, J.; Xu, Z.; Li, R.; Wang, P.; Shang, R.; Cigliano, A.; Ribback, S.; Solinas, A.; Pes, G.M.; et al. SNAI1 Promotes the Cholangiocellular Phenotype, but not Epithelial-Mesenchymal Transition, in a Murine Hepatocellular Carcinoma Model. Cancer Res. 2019, 79, 5563–5574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graziani, I.; Eliasz, S.; De Marco, M.A.; Chen, Y.; Pass, H.I.; De May, R.M.; Strack, P.R.; Miele, L.; Bocchetta, M. Opposite effects of Notch-1 and Notch-2 on mesothelioma cell survival under hypoxia are exerted through the Akt pathway. Cancer Res. 2008, 68, 9678–9685. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Appleby, N.; Fuentes, T.; Longo, L.D.; Bailey, L.L.; Hasaniya, N.; Kearns-Jonker, M. Isolation, Charac-terization, and Spatial Distribution of Cardiac Progenitor Cells in the Sheep Heart. J. Clin. Exp. Cardiol. 2013, S6, 4. [Google Scholar]
- Zhang, Y.; Gao, E.; Guan, H.; Wang, Q.; Zhang, S.; Liu, K.; Yan, F.; Tian, H.; Shan, D.; Xu, H.; et al. Vitamin C treatment of embryos, but not donor cells, improves the cloned embryonic development in sheep. Reprod. Domest. Anim. 2020, 55, 255–265. [Google Scholar] [CrossRef]
- Samiec, M.; Skrzyszowska, M. Assessment of in vitro developmental capacity of porcine nuclear-transferred embryos reconstituted with cumulus oophorus cells undergoing vital diagnostics for apoptosis detection. Ann. Anim. Sci. 2013, 13, 513–529. [Google Scholar] [CrossRef] [Green Version]
- Gavin, W.; Buzzell, N.; Blash, S.; Chen, L.; Hawkins, N.; Miner, K.; Pollock, D.; Porter, C.; Bonzo, D.; Meade, H. Generation of goats by nuclear transfer: A retrospective analysis of a commercial operation (1998–2010). Transgenic Res. 2020, 29, 443–459. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knox, C.; Camberos, V.; Ceja, L.; Monteon, A.; Hughes, L.; Longo, L.; Kearns-Jonker, M. Long-Term Hypoxia Maintains a State of Dedifferentiation and Enhanced Stemness in Fetal Cardiovascular Progenitor Cells. Int. J. Mol. Sci. 2021, 22, 9382. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179382
Knox C, Camberos V, Ceja L, Monteon A, Hughes L, Longo L, Kearns-Jonker M. Long-Term Hypoxia Maintains a State of Dedifferentiation and Enhanced Stemness in Fetal Cardiovascular Progenitor Cells. International Journal of Molecular Sciences. 2021; 22(17):9382. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179382
Chicago/Turabian StyleKnox, Cole, Victor Camberos, Lourdes Ceja, Andrea Monteon, Lorelei Hughes, Lawrence Longo, and Mary Kearns-Jonker. 2021. "Long-Term Hypoxia Maintains a State of Dedifferentiation and Enhanced Stemness in Fetal Cardiovascular Progenitor Cells" International Journal of Molecular Sciences 22, no. 17: 9382. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179382