
The Hidden Side of NCAM Family: NCAM2, a Key Cytoskeleton Organization Molecule Regulating Multiple Neural Functions


Abstract
:1. Introduction
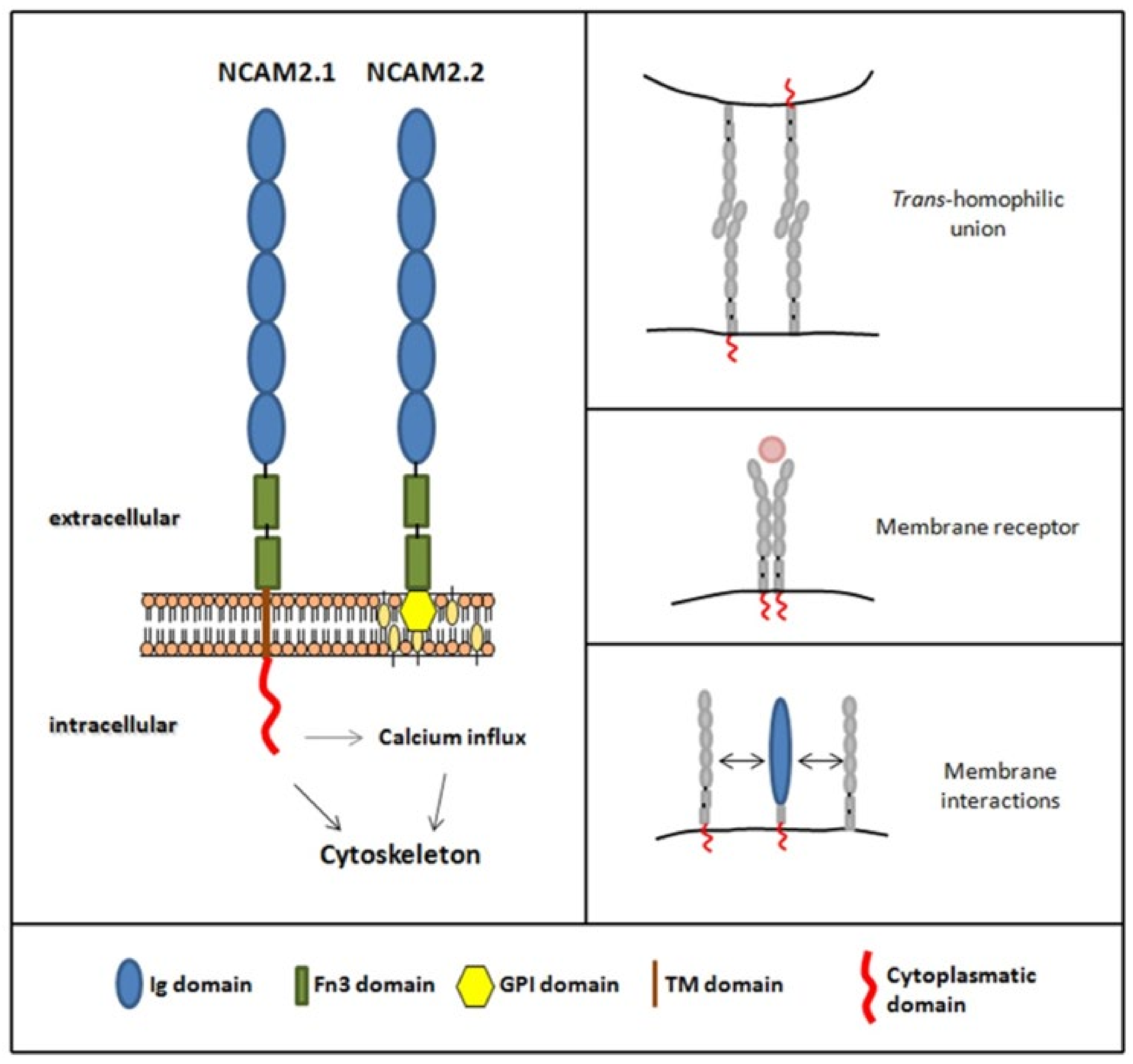
2. NCAM2 Expression and Interactors
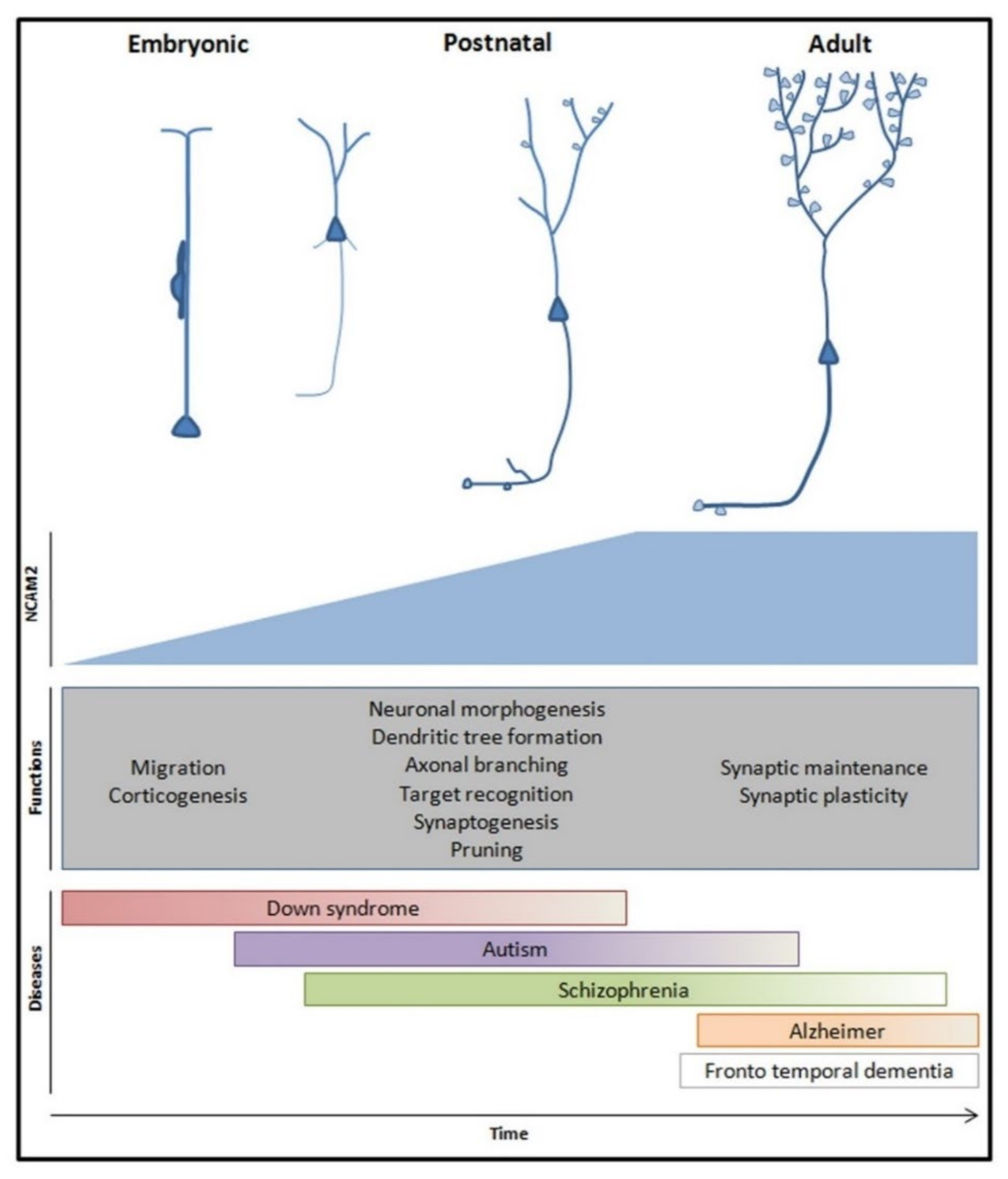
3. NCAM2 in Neuronal Cell Fate Determination and Differentiation
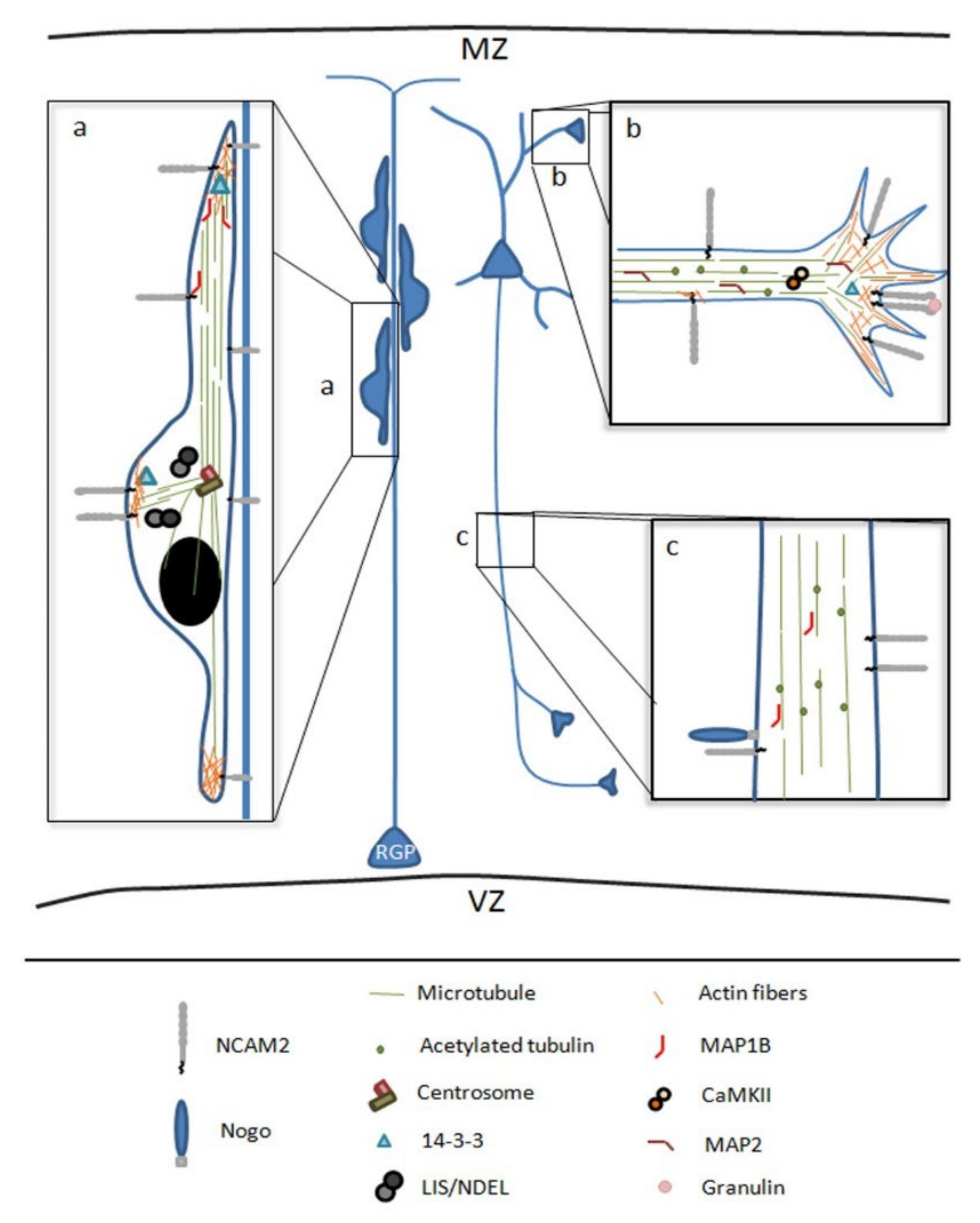
3.1. NCAM2 Role in Neuronal Migration and Corticogenesis
3.2. Neuronal Differentiation
3.2.1. NCAM2 in Dendritic Tree Development
3.2.2. NCAM2 in Axon Formation and Development
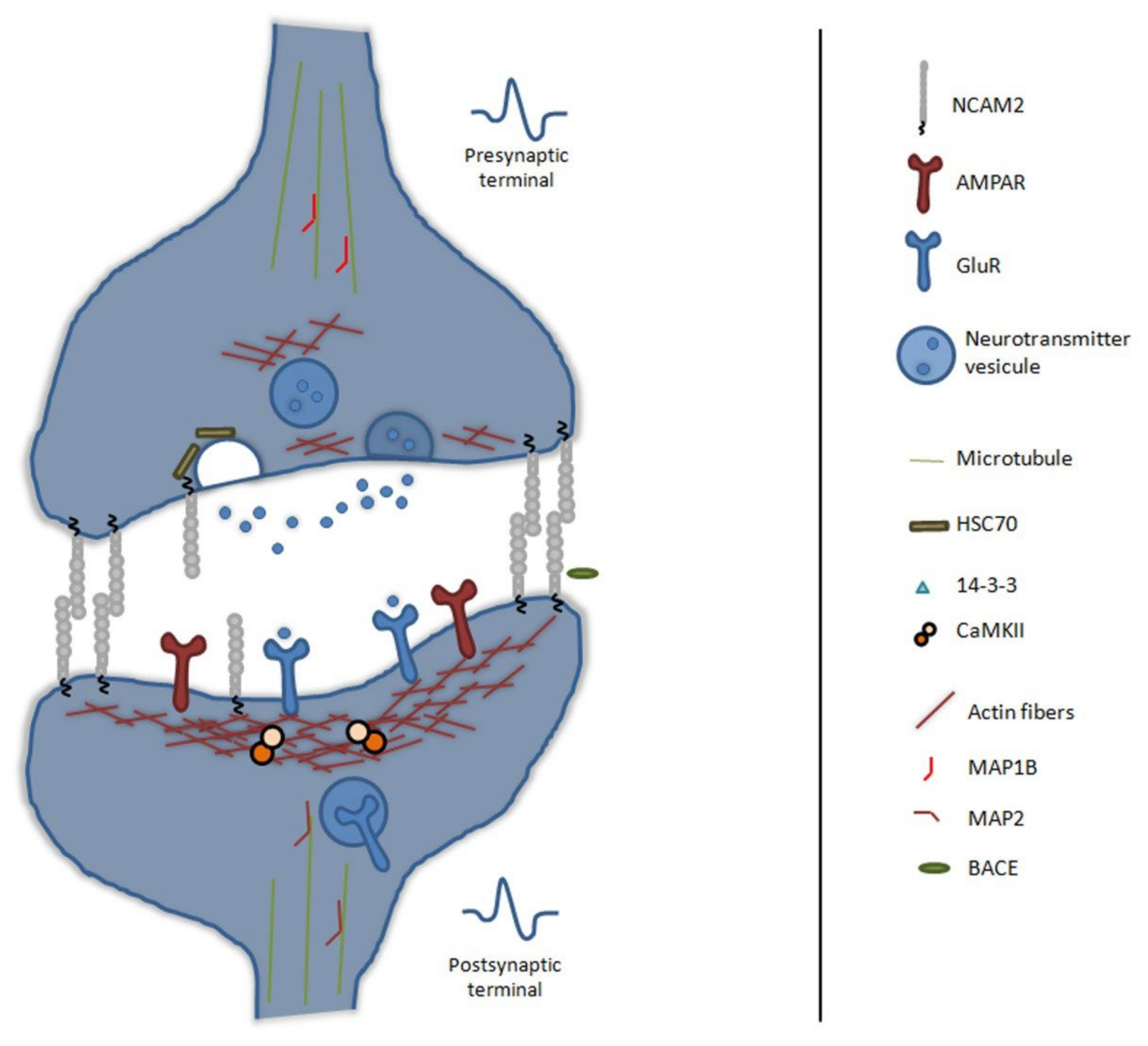
3.2.3. NCAM2 in Synaptogenesis and Synaptic Plasticity
3.3. NCAM2 in Calcium Signaling and Homeostasis
4. NCAM2 in Neuronal Diseases
4.1. Neurodevelopment Diseases
4.2. Neurodegenerative Diseases
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Shapiro, L.; Love, J.; Colman, D.R. Adhesion Molecules in the Nervous System: Structural Insights into Function and Diversity. Annu. Rev. Neurosci. 2007, 30, 451–474. [Google Scholar] [CrossRef]
- Hansen, S.M.; Berezin, V.; Bock, E. Signaling mechanisms of neurite outgrowth induced by the cell adhesion molecules NCAM and N-Cadherin. Cell. Mol. Life Sci. 2008, 65, 3809–3821. [Google Scholar] [CrossRef] [PubMed]
- Shima, Y.; Kawaguchi, S.-Y.; Kosaka, K.; Nakayama, M.; Hoshino, M.; Nabeshima, Y.; Hirano, T.; Uemura, T. Opposing roles in neurite growth control by two seven-pass transmembrane cadherins. Nat. Neurosci. 2007, 10, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Maness, P.F.; Schachner, M. Neural recognition molecules of the immunoglobulin superfamily: Signaling transducers of axon guidance and neuronal migration. Nat. Neurosci. 2006, 10, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Kleene, R.; Mzoughi, M.; Joshi, G.; Kalus, I.; Bormann, U.; Schulze, C.; Xiao, M.-F.; Dityatev, A.; Schachner, M. NCAM-Induced Neurite Outgrowth Depends on Binding of Calmodulin to NCAM and on Nuclear Import of NCAM and fak Fragments. J. Neurosci. 2010, 30, 10784–10798. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-Y.; Mo, J.; Han, S.B.; Choi, S.; Moon, B.; Rhyu, I.; Sun, W.; Kim, H. The expression of non-clustered protocadherins in adult rat hippocampal formation and the connecting brain regions. Neuroscience 2010, 170, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Missaire, M.; Hindges, R. The role of cell adhesion molecules in visual circuit formation: From neurite outgrowth to maps and synaptic specificity. Dev. Neurobiol. 2015, 75, 569–583. [Google Scholar] [CrossRef] [Green Version]
- Gennarini, G.; Bizzoca, A.; Picocci, S.; Puzzo, D.; Corsi, P.; Furley, A.J.W. The role of Gpi-anchored axonal glycoproteins in neural development and neurological disorders. Mol. Cell. Neurosci. 2017, 81, 49–63. [Google Scholar] [CrossRef]
- Sakurai, T. The role of cell adhesion molecules in brain wiring and neuropsychiatric disorders. Mol. Cell. Neurosci. 2017, 81, 4–11. [Google Scholar] [CrossRef]
- Jin, Y.; Li, H. Revisiting Dscam diversity: Lessons from clustered protocadherins. Experientia 2019, 76, 667–680. [Google Scholar] [CrossRef]
- Jan, Y.-N.; Jan, L. Branching out: Mechanisms of dendritic arborization. Nat. Rev. Neurosci. 2010, 11, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Xu, Y.; Liu, J.; Xu, Q.; Ye, H. Down syndrome cell adhesion molecule and its functions in neural development. Neurosci. Bull. 2011, 27, 45–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchinson, K.M.; Vonhoff, F.; Duch, C. Dscam1 Is Required for Normal Dendrite Growth and Branching But Not for Dendritic Spacing in Drosophila Motoneurons. J. Neurosci. 2014, 34, 1924–1931. [Google Scholar] [CrossRef] [PubMed]
- Namba, T.; Funahashi, Y.; Nakamuta, S.; Xu, C.; Takano, T.; Kaibuchi, K. Extracellular and Intracellular Signaling for Neuronal Polarity. Physiol. Rev. 2015, 95, 995–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baig, D.N.; Yanagawa, T.; Tabuchi, K. Distortion of the normal function of synaptic cell adhesion molecules by genetic variants as a risk for autism spectrum disorders. Brain Res. Bull. 2017, 129, 82–90. [Google Scholar] [CrossRef]
- Schmid, R.; Maness, P.F. L1 and NCAM adhesion molecules as signaling coreceptors in neuronal migration and process outgrowth. Curr. Opin. Neurobiol. 2008, 18, 245–250. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Yu, H.; Jiang, S.; Liao, J.; Lu, T.; Wang, L.; Zhang, D.; Yue, W. Evidence for Association of Cell Adhesion Molecules Pathway and NLGN1 Polymorphisms with Schizophrenia in Chinese Han Population. PLoS ONE 2015, 10, e0144719. [Google Scholar] [CrossRef] [Green Version]
- Tost, H.; Bilek, E.; Meyer-Lindenberg, A. Brain connectivity in psychiatric imaging genetics. NeuroImage 2012, 62, 2250–2260. [Google Scholar] [CrossRef]
- Wang, K.S.; Liu, X.F.; Aragam, N. A genome-wide meta-analysis identifies novel loci associated with schizophrenia and bipolar disorder. Schizophr. Res. 2010, 124, 192–199. [Google Scholar] [CrossRef]
- Chapman, N.H.; Estes, A.; Munson, J.; Bernier, R.; Webb, S.J.; Rothstein, J.H.; Minshew, N.J.; Dawson, G.; Schellenberg, G.D.; Wijsman, E.M. Genome-scan for IQ discrepancy in autism: Evidence for loci on chromosomes 10 and 16. Qual. Life Res. 2010, 129, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Morrow, E.; Yoo, S.; Flavell, S.; Kim, T.K.; Lin, Y.; Hill, R.S.; Mukaddes, N.M.; Balkhy, S.; Gascon, G.; Hashmi, A.; et al. Identifying autism loci and genes by tracing recent shared ancestry. Science 2008, 321, 218–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depienne, C.; Bouteiller, D.; Keren, B.; Cheuret, E.; Poirier, K.; Trouillard, O.; Benyahia, B.; Quelin, C.; Carpentier, W.; Julia, S.; et al. Sporadic Infantile Epileptic Encephalopathy Caused by Mutations in PCDH19 Resembles Dravet Syndrome but Mainly Affects Females. PLoS Genet. 2009, 5, e1000381. [Google Scholar] [CrossRef]
- Hussman, J.P.; Chung, R.-H.; Griswold, A.J.; Jaworski, J.M.; Salyakina, D.; Ma, D.; Konidari, I.; Whitehead, P.L.; Vance, J.M.; Martin, E.R.; et al. A noise-reduction GWAS analysis implicates altered regulation of neurite outgrowth and guidance in autism. Mol. Autism 2011, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- Neale, B.M.; Kou, Y.; Liu, L.; Ma’Ayan, A.; Samocha, K.E.; Sabo, A.; Lin, C.F.; Stevens, S.; Wang, L.-S.; Makarov, V.; et al. Patterns and rates of exonic de novo mutations in autism spectrum disorders. Nature 2012, 485, 242–245. [Google Scholar] [CrossRef]
- Cukier, H.N.; Lee, J.M.; Ma, D.; Young, J.I.; Mayo, V.; Butler, B.L.; Ramsook, S.S.; Rantus, J.A.; Abrams, A.J.; Whitehead, P.L.; et al. The Expanding Role of MBD Genes in Autism: Identification of aMECP2Duplication and Novel Alterations inMBD5,MBD6, andSETDB1. Autism Res. 2012, 5, 385–397. [Google Scholar] [CrossRef] [Green Version]
- Kenny, E.M.; Cormican, P.; Furlong, S.; Heron, E.; Kenny, G.; Fahey, C.; Kelleher, E.; Ennis, S.; Tropea, D.; Morris, D.W.; et al. Excess of rare novel loss-of-function variants in synaptic genes in schizophrenia and autism spectrum disorders. Mol. Psychiatry 2014, 19, 872–879. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Schroer, R.; Yan, J.; Song, W.; Yang, C.; Bockholt, A.; Cook, E.H., Jr.; Charles, C.S.; Steve, E.S.; Sommer, S.S.; et al. High frequency of neurexin 1β signal peptide structural variants in patients with autism. Neurosci. Lett. 2006, 409, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Szatmari, P.; Paterson, A.; Zwaigenbaum, L.; Roberts, W.; Brian, J.; Liu, X.Q.; Vincent, J.; Skaug, J.; Thompson, A.; Meyer, K.; et al. Mapping autism risk loci using genetic linkage and chromosomal rearrangements. Nat. Genet. 2007, 39, 319. [Google Scholar]
- Zweier, C.; de Jong, E.K.; Zweier, M.; Orrico, A.; Ousager, L.B.; Collins, A.L.; Bijlsma, E.K.; Oortveld, M.A.; Ekici, A.B.; Reis, A.; et al. CNTNAP2 and NRXN1 Are Mutated in Autosomal-Recessive Pitt-Hopkins-like Mental Retardation and Determine the Level of a Common Synaptic Protein in Drosophila. Am. J. Hum. Genet. 2009, 85, 655–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glessner, J.T.; Wang, K.; Cai, G.; Korvatska, O.; Kim, C.E.; Wood, S.; Zhang, H.; Estes, A.; Brune, C.W.; Hakonarson, H.; et al. Autism genome-wide copy number variation reveals ubiquitin and neuronal genes. Nature 2009, 459, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Xiong, Z.; Zhang, L.; Liu, Y.; Lu, L.; Peng, Y.; Guo, H.; Zhao, J.; Xia, K.; Hu, Z. Variations analysis of NLGN3 and NLGN4X gene in Chinese autism patients. Mol. Biol. Rep. 2014, 41, 4133–4140. [Google Scholar] [CrossRef]
- Jamain, S.; Radyushkin, K.; Hammerschmidt, K.; Granon, S.; Boretius, S.; Varoqueaux, F.; Ramanantsoa, N.; Gallego, J.; Ronnenberg, A.; Winter, D.; et al. Reduced social interaction and ultrasonic communication in a mouse model of monogenic heritable autism. Proc. Natl. Acad. Sci. USA 2008, 105, 1710–1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Hu, Z.; He, Y.; Xiong, Z.; Long, Z.; Peng, Y.; Fengxiao, B.; Jie, L.; Guanglei, X.; Xia, K.; et al. Association analysis of CNTNAP2 poly-morphisms with autism in the Chinese Han population. Psychiatr. Genet. 2010, 20, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, A.J.O.; Bishop, D.V.M.; Ang, Q.W.; Pennell, C.E.; Fisher, S. CNTNAP2variants affect early language development in the general population. Genes Brain Behav. 2011, 10, 451–456. [Google Scholar] [CrossRef]
- Peñagarikano, O.; Abrahams, B.S.; Herman, E.I.; Winden, K.D.; Gdalyahu, A.; Dong, H.; Sonnenblick, L.I.; Gruver, R.; Almajano, J.; Geschwind, D.H. Absence of CNTNAP2 leads to epilepsy, neuronal migration abnormalities, and core autism-related deficits. Cell 2011, 147, 235–246. [Google Scholar] [CrossRef] [Green Version]
- Gdalyahu, A.; Lázaro, M.; Penagarikano, O.; Golshani, P.; Trachtenberg, J.T.; Gescwind, D.H. The Autism Related Protein Contactin-Associated Protein-Like 2 (CNTNAP2) Stabilizes New Spines: An In Vivo Mouse Study. PLoS ONE 2015, 10, e0125633. [Google Scholar] [CrossRef]
- Varea, O.; Martin-De-Saavedra, M.D.; Kopeikina, K.J.; Schürmann, B.; Fleming, H.J.; Fawcett-Patel, J.M.; Bach, A.; Jang, S.; Peles, E.; Kim, E.; et al. Synaptic abnormalities and cytoplasmic glutamate receptor aggregates in contactin associated protein-like 2/Caspr2 knockout neurons. Proc. Natl. Acad. Sci. USA 2015, 112, 6176–6181. [Google Scholar] [CrossRef] [Green Version]
- Atz, M.E.; Rollins, B.; Vawter, M.P. NCAM1 association study of bipolar disorder and schizophrenia: Polymorphisms and alternatively spliced isoforms lead to similarities and differences. Psychiatr. Genet. 2007, 17, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, P.F.; Keefe, R.S.; Lange, L.A.; Lange, E.M.; Stroup, T.S.; Lieberman, J.; Maness, P.F. NCAM1 and Neurocognition in Schizophrenia. Biol. Psychiatry 2007, 61, 902–910. [Google Scholar] [CrossRef]
- Deng, S.-P.; Hu, W.; Calhoun, V.D.; Wang, Y.-P. Integrating Imaging Genomic Data in the Quest for Biomarkers of Schizophrenia Disease. IEEE/ACM Trans. Comput. Biol. Bioinform. 2018, 15, 1480–1491. [Google Scholar] [CrossRef]
- Wedervang-Resell, K.; Ueland, T.; Aukrust, P.; Friis, S.; Holven, K.B.; Johannessen, C.H.; Lekva, T.; Lonning, V.; Smelror, R.E.; Szabo, A.; et al. Reduced levels of circulating adhesion molecules in adolescents with early-onset psychosis. NPJ Schizophr. 2020, 6, 1–8. [Google Scholar] [CrossRef]
- Nikitczuk, J.S.; Patil, S.B.; Matikainen-Ankney, B.A.; Scarpa, J.; Shapiro, M.L.; Benson, D.L.; Huntley, G.W. N-cadherin regulates molecular organization of excitatory and inhibitory synaptic circuits in adult hippocampus in vivo. Hippocampus 2014, 24, 943–962. [Google Scholar] [CrossRef] [Green Version]
- Avdic, U.; Ahl, M.; Andersson, M.; Ekdahl, C.T. Levetiracetam and N-Cadherin Antibody Alleviate Brain Pathology Without Reducing Early Epilepsy Development After Focal Non-convulsive Status Epilepticus in Rats. Front. Neurol. 2021, 12, 630154. [Google Scholar] [CrossRef] [PubMed]
- Takano, K.; Nakagawa, E.; Inoue, K.; Kamada, F.; Kure, S.; Goto, Y.-I. A loss-of-function mutation in theFTSJ1 gene causes nonsyndromic X-linked mental retardation in a japanese family. Am. J. Med Genet. Part B Neuropsychiatr. Genet. 2008, 147B, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, E.; Nakagawa, E.; Nakatani, K.; Sasaki, M.; Goto, Y.-I. Novel AGTR2 missense mutation in a Japanese boy with severe mental retardation, pervasive developmental disorder, and epilepsy. Brain Dev. 2012, 34, 776–779. [Google Scholar] [CrossRef]
- Hayase, Y.; Amano, S.; Hashizume, K.; Tominaga, T.; Miyamoto, H.; Kanno, Y.; Ueno-Inoue, Y.; Inoue, T.; Yamada, M.; Ogata, S.; et al. Down syndrome cell adhesion molecule like-1 (DSCAML1) links the GABA system and seizure susceptibility. Acta Neuropathol. Commun. 2020, 8, 1–17. [Google Scholar] [CrossRef]
- Wang, W.; Wang, L.; Luo, J.; Xi, Z.; Wang, X.; Chen, G.; Chu, L. Role of a Neural Cell Adhesion Molecule Found in Cerebrospinal Fluid as a Potential Biomarker for Epilepsy. Neurochem. Res. 2012, 37, 819–825. [Google Scholar] [CrossRef]
- Cho, S.; Muthukumar, A.K.; Stork, T.; Coutinho-Budd, J.C.; Freeman, M.R. Focal adhesion molecules regulate astrocyte morphology and glutamate transporters to suppress seizure-like behavior. Proc. Natl. Acad. Sci. USA 2018, 115, 11316–11321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsogiannis, M.D.; Pancho, A.; Aerts, T.; Sachse, S.M.; Vanlaer, R.; Noterdaeme, L.; Schmucker, D.; Seuntjens, E. Subtle Roles of Down Syndrome Cell Adhesion Molecules in Embryonic Forebrain Development and Neuronal Migration. Front. Cell Dev. Biol. 2021, 8, 1859. [Google Scholar] [CrossRef]
- Chmielewska, J.J.; Kuzniewska, B.; Milek, J.; Urbanska, K.; Dziembowska, M. Neuroligin 1, 2, and 3 Regulation at the Synapse: FMRP-Dependent Translation and Activity-Induced Proteolytic Cleavage. Mol. Neurobiol. 2018, 56, 2741–2759. [Google Scholar] [CrossRef] [Green Version]
- Alves-Sampaio, A.; Troca-Marín, J.A.; Montesinos, M.L. NMDA-Mediated Regulation of DSCAM Dendritic Local Translation Is Lost in a Mouse Model of Down’s Syndrome. J. Neurosci. 2010, 30, 13537–13548. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.-Y.; Xu, L.; Wang, J.; Hong, Y.; Wang, Y.; Zhu, Q.; Wang, D.; Zhang, X.-Y.; Liu, C.-Y.; Fang, K.-H.; et al. Suppressing the DSCAM/PAK1 pathway reverses neurogenesis deficits in Down Syndrome patient iPSC-derived cerebral organoids. J. Clin. Investig. 2021, 131, e135763. [Google Scholar] [CrossRef] [PubMed]
- Pebusque, M.J.; Coulier, F.; Birnbaum, D.; Pontarotti, P. Ancient large-scale genome duplications: Phylogenetic and linkage analyses shed light on chordate genome evolution. Mol. Biol. Evol. 1998, 15, 1145–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makino, T.; McLysaght, A. Ohnologs in the human genome are dosage balanced and frequently associated with disease. Proc. Natl. Acad. Sci. USA 2010, 107, 9270–9274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiselyov, V.V.; Skladchikova, G.; Hinsby, A.M.; Jensen, P.H.; Kulahin, N.; Soroka, V.; Pedersen, N.; Tsetlin, V.; Poulsen, F.M.; Berezin, V.; et al. Structural Basis for a Direct Interaction between FGFR1 and NCAM and Evidence for a Regulatory Role of ATP. Structure 2003, 11, 691–701. [Google Scholar] [CrossRef] [Green Version]
- Mehrabian, M.; Hildebrandt, H.; Schmitt-Ulms, G. NCAM1 Polysialylation: The Prion Protein’s Elusive Reason for Being? ASN Neuro 2016, 8. [Google Scholar] [CrossRef] [Green Version]
- Angata, K.; Huckaby, V.; Ranscht, B.; Terskikh, A.; Marth, J.D.; Fukuda, M. Polysialic Acid-Directed Migration and Differentiation of Neural Precursors Are Essential for Mouse Brain Development. Mol. Cell. Biol. 2007, 27, 6659–6668. [Google Scholar] [CrossRef] [Green Version]
- Sytnyk, V.; Leshchyns’Ka, I.; Schachner, M. Neural Cell Adhesion Molecules of the Immunoglobulin Superfamily Regulate Synapse Formation, Maintenance, and Function. Trends Neurosci. 2017, 40, 295–308. [Google Scholar] [CrossRef]
- Dityatev, A.; Dityateva, G.; Sytnyk, V.; Delling, M.; Toni, N.; Nikonenko, I.; Muller, D.; Schachner, M. Polysialylated Neural Cell Adhesion Molecule Promotes Remodeling and Formation of Hippocampal Synapses. J. Neurosci. 2004, 24, 9372–9382. [Google Scholar] [CrossRef] [Green Version]
- Muller, D.; Mendez, P.; DeRoo, M.; Klauser, P.; Steen, S.; Poglia, L. Role of NCAM in Spine Dynamics and Synaptogenesis. Adv. Exp. Med. Biol. 2010, 663, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.D.; Tiwari, Y.; Kaplan, W.; Heath, A.; Mitchell, P.B.; Schofield, P.R.; Fullerton, J.M. Characterisation of Genetic Variation in ST8SIA2 and Its Interaction Region in NCAM1 in Patients with Bipolar Disorder. PLoS ONE 2014, 9, e92556. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Xiao, M.; Ji, S.; Tang, J.; Xu, L.; Li, X.; Li, M.; Wang, H.-Z.; Jiang, H.-Y.; Zhang, D.-F.; et al. Promoter variant rs2301228 on the neural cell adhesion molecule 1 gene confers risk of schizophrenia in Han Chinese. Schizophr. Res. 2014, 160, 88–96. [Google Scholar] [CrossRef]
- Di Cristo, G.; Chattopadhyaya, B.; Kuhlman, S.; Fu, Y.; Bélanger, M.-C.; Wu, C.Z.; Rutishauser, U.; Maffei, L.; Huang, Z.J. Activity-dependent PSA expression regulates inhibitory maturation and onset of critical period plasticity. Nat. Neurosci. 2007, 10, 1569–1577. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyaya, B.; Baho, E.; Huang, Z.J.; Schachner, M.; Di Cristo, G. Neural Cell Adhesion Molecule-Mediated Fyn Activation Promotes GABAergic Synapse Maturation in Postnatal Mouse Cortex. J. Neurosci. 2013, 33, 5957–5968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alenius, M.; Bohm, S. Identification of a Novel Neural Cell Adhesion Molecule-related Gene with a Potential Role in Selective Axonal Projection. J. Biol. Chem. 1997, 272, 26083–26086. [Google Scholar] [CrossRef] [Green Version]
- Yoshihara, Y.; Kawasaki, M.; Tamada, A.; Fujita, H.; Hayashi, H.; Kagamiyama, H.; Mori, K. OCAM: A New Member of the Neural Cell Adhesion Molecule Family Related to Zone-to-Zone Projection of Olfactory and Vomeronasal Axons. J. Neurosci. 1997, 17, 5830–5842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paoloni-Giacobino, A.; Chen, H.; Antonarakis, S.E. Cloning of a Novel Human Neural Cell Adhesion Molecule Gene (NCAM2) That Maps to Chromosome Region 21q21 and Is Potentially Involved in Down Syndrome. Genomics 1997, 43, 43–51. [Google Scholar] [CrossRef]
- Ichinohe, N.; Yoshihara, Y.; Hashikawa, T.; Rockland, K.S. Developmental study of dendritic bundles in layer 1 of the rat granular retrosplenial cortex with special reference to a cell adhesion molecule, OCAM. Eur. J. Neurosci. 2003, 18, 1764–1774. [Google Scholar] [CrossRef]
- Parcerisas, A.; Pujadas, L.; Ortega-Gascó, A.; Perelló-Amorós, B.; Viais, R.; Hino, K.; Figueiro-Silva, J.; La Torre, A.; Trullás, R.; Simó, S.; et al. NCAM2 Regulates Dendritic and Axonal Differentiation through the Cytoskeletal Proteins MAP2 and 14-3-3. Cereb. Cortex 2020, 30, 3781–3799. [Google Scholar] [CrossRef]
- Parcerisas, A.; Ortega-Gascó, A.; Hernaiz-Llorens, M.; Odena, M.A.; Ulloa, F.; de Oliveira, E.; Bosch, M.; Pujadas, L.; Soriano, E. New Partners Identified by Mass Spectrometry Assay Reveal Functions of NCAM2 in Neural Cytoskeleton Organization. Int. J. Mol. Sci. 2021, 22, 7404. [Google Scholar] [CrossRef]
- Niethammer, P.; Delling, M.; Sytnyk, V.; Dityatev, A.; Fukami, K.; Schachner, M. Cosignaling of NCAM via lipid rafts and the FGF receptor is required for neuritogenesis. J. Cell Biol. 2002, 157, 521–532. [Google Scholar] [CrossRef]
- Kamiguchi, H. The region-specific activities of lipid rafts during axon growth and guidance. J. Neurochem. 2006, 98, 330–335. [Google Scholar] [CrossRef]
- Huttlin, E.L.; Jedrychowski, M.P.; Elias, J.E.; Goswami, T.; Rad, R.; Beausoleil, S.A.; Villen, J.; Haas, W.; Sowa, M.E.; Gygi, S.P. A Tissue-Specific Atlas of Mouse Protein Phosphorylation and Expression. Cell 2010, 143, 1174–1189. [Google Scholar] [CrossRef] [Green Version]
- Kulahin, N.; Walmod, P.S. The Neural Cell Adhesion Molecule NCAM2/OCAM/RNCAM, a Close Relative to NCAM. In Structure and Function of the Neural Cell Adhesion Molecule NCAM; Springer: New York, NY, USA, 2009; Volume 663, pp. 403–420. [Google Scholar]
- Kim, W.H.; Watanabe, H.; Lomoio, S.; Tesco, G. Spatiotemporal processing of neural cell adhesion molecules 1 and 2 by BACE1 in vivo. J. Biol. Chem. 2021, 296, 100372. [Google Scholar] [CrossRef]
- Leshchyns’Ka, I.; Liew, H.T.; Shepherd, C.; Halliday, G.M.; Stevens, C.H.; Ke, Y.D.; Ittner, L.M.; Sytnyk, V. Aβ-dependent reduction of NCAM2-mediated synaptic adhesion contributes to synapse loss in Alzheimer’s disease. Nat. Commun. 2015, 6, 8836. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.K.; Yanagisawa, M. Glycobiology of neural stem cells. CNS Neurol. Disord. Drug Targets 2006, 5, 415–423. [Google Scholar] [CrossRef]
- Kulahin, N.; Kristensen, O.; Rasmussen, K.K.; Olsen, L.; Rydberg, P.; Vestergaard, B.; Kastrup, J.S.; Berezin, V.; Bock, E.; Walmod, P.S.; et al. Structural Model and trans-Interaction of the Entire Ectodomain of the Olfactory Cell Adhesion Molecule. Structure 2011, 19, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Deleyrolle, L.; Sabourin, J.-C.; Rothhut, B.; Fujita, H.; Guichet, P.-O.; Teigell, M.; Ripoll, C.; Chauvet, N.; Perrin, F.; Mamaeva, D.; et al. OCAM Regulates Embryonic Spinal Cord Stem Cell Proliferation by Modulating ErbB2 Receptor. PLoS ONE 2015, 10, e0122337. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, K.K.; Falkesgaard, M.H.; Winther, M.; Roed, N.K.; Quistgaard, C.L.; Teisen, M.N.; Edslev, S.M.; Petersen, D.L.; Aljubouri, A.; Christensen, C.; et al. NCAM2 Fibronectin type-III domains form a rigid structure that binds and activates the Fibroblast Growth Factor Receptor. Sci. Rep. 2018, 8, 8957. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.; Husi, H.; Yu, L.; Brandon, J.M.; Anderson, C.N.G.; Blackstock, W.P.; Choudhary, J.; Grant, S. Molecular characterization and comparison of the components and multiprotein complexes in the postsynaptic proteome. J. Neurochem. 2006, 97, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Kislinger, T.; Cox, B.; Kannan, A.; Chung, C.; Hu, P.; Ignatchenko, A.; Scott, M.S.; Gramolini, A.O.; Morris, Q.; Hallett, M.T.; et al. Global Survey of Organ and Organelle Protein Expression in Mouse: Combined Proteomic and Transcriptomic Profiling. Cell 2006, 125, 173–186. [Google Scholar] [CrossRef] [Green Version]
- Sheng, L.; Leshchyns’Ka, I.; Sytnyk, V. Neural Cell Adhesion Molecule 2 Promotes the Formation of Filopodia and Neurite Branching by Inducing Submembrane Increases in Ca2+ Levels. J. Neurosci. 2015, 35, 1739–1752. [Google Scholar] [CrossRef] [Green Version]
- Urbanska, M.; Blazejczyk, M.; Jaworski, J. Molecular Basis of Dendritic Arborization. Available online: https://pubmed.ncbi.nlm.nih.gov/18511961/ (accessed on 3 August 2021).
- Da Silva, J.S.; Dotti, C.G. Breaking the neuronal sphere: Regulation of the actin cytoskeleton in neuritogenesis. Nat. Rev. Neurosci. 2002, 3, 694–704. [Google Scholar] [CrossRef]
- Cheng, P.-L.; Poo, M.-M. Early Events in Axon/Dendrite Polarization. Annu. Rev. Neurosci. 2012, 35, 181–201. [Google Scholar] [CrossRef] [PubMed]
- Takano, T.; Xu, C.; Funahashi, Y.; Namba, T.; Kaibuchi, K. Neuronal polarization. Development 2015, 142, 2088–2093. [Google Scholar] [CrossRef] [Green Version]
- Kawauchi, T. Cell Adhesion and Its Endocytic Regulation in Cell Migration during Neural Development and Cancer Metastasis. Int. J. Mol. Sci. 2012, 13, 4564–4590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farah, C.A.; Leclerc, N. HMWMAP2: New perspectives on a pathway to dendritic identity. Cell Motil. Cytoskelet. 2008, 65, 515–527. [Google Scholar] [CrossRef]
- Dalva, M.B.; McClelland, A.C.; Kayser, M.S. Cell adhesion molecules: Signalling functions at the synapse. Nat. Rev. Neurosci. 2007, 8, 206–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seong, E.; Yuan, L.; Arikkath, J. Cadherins and catenins in dendrite and synapse morphogenesis. Cell Adhes. Migr. 2015, 9, 202–213. [Google Scholar] [CrossRef] [Green Version]
- Ichinohe, N.; Knight, A.; Ogawa, M.; Ohshima, T.; Mikoshiba, K.; Yoshihara, Y.; Terashima, T.; Rockland, K. Unusual Patch-Matrix Organization in the Retrosplenial Cortex of the reeler Mouse and Shaking Rat Kawasaki. Cereb. Cortex 2008, 18, 1125–1138. [Google Scholar] [CrossRef]
- Borisovska, M.; McGinley, M.J.; Bensen, A.; Westbrook, G.L. Loss of olfactory cell adhesion molecule reduces the synchrony of mitral cell activity in olfactory glomeruli. J. Physiol. 2011, 589, 1927–1941. [Google Scholar] [CrossRef]
- Walz, A.; Mombaerts, P.; Greer, C.A.; Treloar, H.B. Disrupted compartmental organization of axons and dendrites within olfactory glomeruli of mice deficient in the olfactory cell adhesion molecule, OCAM. Mol. Cell. Neurosci. 2006, 32, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kawauchi, T.; Hoshino, M. Molecular Pathways Regulating Cytoskeletal Organization and Morphological Changes in Migrating Neurons. Dev. Neurosci. 2008, 30, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Hippenmeyer, S. Molecular Pathways Controlling the Sequential Steps of Cortical Projection Neuron Migration. Cell. Mol. Control. Neuronal Migr. 2014, 800, 1–24. [Google Scholar] [CrossRef]
- Reiner, O.; Sapir, T. LIS1 functions in normal development and disease. Curr. Opin. Neurobiol. 2013, 23, 951–956. [Google Scholar] [CrossRef]
- Tsai, L.-H.; Gleeson, J.G. Nucleokinesis in Neuronal Migration. Neuron 2005, 46, 383–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallee, R.B. The cellular roles of the lissencephaly gene LIS1, and what they tell us about brain development. Genes Dev. 2006, 20, 1384–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobyns, W.B. Lissencephaly: A human brain malformation associated with deletion of the LIS1 gene located at chromosome 17p13. JAMA 1993, 270, 2838–2842. [Google Scholar] [CrossRef]
- Niethammer, M.; Smith, D.S.; Ayala, R.; Peng, J.; Ko, J.; Lee, M.-S.; Morabito, M.; Tsai, L.-H. NUDEL Is a Novel Cdk5 Substrate that Associates with LIS1 and Cytoplasmic Dynein. Neuron 2000, 28, 697–711. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, S.; Shionoya, A.; Ishida, M.; Gambello, M.J.; Yingling, J.; Wynshaw-Boris, A.; Hirotsune, S. A LIS1/NUDEL/Cytoplasmic Dynein Heavy Chain Complex in the Developing and Adult Nervous System. Neuron 2000, 28, 681–696. [Google Scholar] [CrossRef] [Green Version]
- Toyooka, K.; Shionoya, A.; Gambello, M.J.; Cardoso, C.; Leventer, R.; Ward, H.L.; Ayala, R.; Tsai, L.-H.; Dobyns, W.; Ledbetter, D.; et al. 14-3-3ε is important for neuronal migration by binding to NUDEL: A molecular explanation for Miller–Dieker syndrome. Nat. Genet. 2003, 34, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Hirotsune, S.; Wynshaw-Boris, A. The essential role of LIS1, NDEL1 and Aurora—A in polarity formation and microtubule organization during neurogensis. Cell Adhes. Migr. 2010, 4, 180–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambello, M.J.; Darling, D.L.; Yingling, J.; Tanaka, T.; Gleeson, J.G.; Wynshaw-Boris, A. Multiple Dose-Dependent Effects of Lis1 on Cerebral Cortical Development. J. Neurosci. 2003, 23, 1719–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Billault, C.; Del Rio, J.A.; Ureña, J.M.; Jiménez-Mateos, E.M.; Barallobre, M.J.; Pascual, M.; Pujadas, L.; Simó, S.; La Torre, A.; Gavin, R.; et al. A role of MAP1B in Reelin-dependent Neuronal Migration. Cereb. Cortex 2005, 15, 1134–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasahara, K.; Watanabe, K.; Kozutsumi, Y.; Oohira, A.; Yamamoto, T.; Sanai, Y. Association of GPI-anchored protein TAG-1 with src-family kinase Lyn in lipid rafts of cerebellar granule cells. Neurochem. Res. 2002, 27, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, K.; Watanabe, K.; Takeuchi, K.; Kaneko, H.; Oohira, A.; Yamamoto, T.; Sanai, Y. Involvement of Gangliosides in Glycosylphosphatidylinositol-anchored Neuronal Cell Adhesion Molecule TAG-1 Signaling in Lipid Rafts. J. Biol. Chem. 2000, 275, 34701–34709. [Google Scholar] [CrossRef] [Green Version]
- Al-Bassam, J.; Ozer, R.S.; Safer, D.; Halpain, S.; Milligan, R.A. MAP2 and tau bind longitudinally along the outer ridges of microtubule protofilaments. J. Cell Biol. 2002, 157, 1187–1196. [Google Scholar] [CrossRef]
- Caceres, A.; Mautino, J.; Kosik, K.S. Suppression of MAP2 in cultured cerebeller macroneurons inhibits minor neurite formation. Neuron 1992, 9, 607–618. [Google Scholar] [CrossRef]
- Harada, A.; Teng, J.; Takei, Y.; Oguchi, K.; Hirokawa, N. MAP2 is required for dendrite elongation, PKA anchoring in dendrites, and proper PKA signal transduction. J. Cell Biol. 2002, 158, 541–549. [Google Scholar] [CrossRef] [Green Version]
- Khuchua, Z.; Wozniak, D.; Bardgett, M.; Yue, Z.; McDonald, M.; Boero, J.; Hartman, R.; Sims, H.; Strauss, A. Deletion of the n-terminus of murine map2 by gene targeting disrupts hippocampal ca1 neuron architecture and alters contextual memory. Neuroscience 2003, 119, 101–111. [Google Scholar] [CrossRef]
- Fink, C.C.; Bayer, K.-U.; Myers, J.W.; Ferrell, J.E.; Schulman, H.; Meyer, T. Selective Regulation of Neurite Extension and Synapse Formation by the β but not the α Isoform of CaMKII. Neuron 2003, 39, 283–297. [Google Scholar] [CrossRef] [Green Version]
- Sogawa, Y.; Yoshimura, Y.; Yamauchi, T. Investigation of the Ca2+-independent form of Ca2+/calmodulin-dependent protein kinase II in neurite outgrowth. Brain Res. Protoc. 2001, 8, 159–169. [Google Scholar] [CrossRef]
- Bodrikov, V.; Sytnyk, V.; Leshchyns’Ka, I.; Hertog, J.D.; Schachner, M. NCAM induces CaMKIIα-mediated RPTPα phosphorylation to enhance its catalytic activity and neurite outgrowth. J. Cell Biol. 2008, 182, 1185–1200. [Google Scholar] [CrossRef] [Green Version]
- Jansen, S.; Melková, K.; Trosanova, Z.; Hanakova, K.; Zachrdla, M.; Novacek, J.; Župa, E.; Zdrahal, Z.; Hritz, J.; Žídek, L. Quantitative mapping of microtubule-associated protein 2c (MAP2c) phosphorylation and regulatory protein 14-3-3ζ-binding sites reveals key differences between MAP2c and its homolog Tau. J. Biol. Chem. 2017, 292, 6715–6727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sluchanko, N.; Gusev, N.B. 14-3-3 Proteins and regulation of cytoskeleton. Biochemistry 2010, 75, 1528–1546. [Google Scholar] [CrossRef] [PubMed]
- Kent, C.B.; Shimada, T.; Ferraro, G.B.; Ritter, B.; Yam, P.T.; McPherson, P.S.; Charron, F.; Kennedy, T.E.; Fournier, A.E. 14-3-3 Proteins Regulate Protein Kinase A Activity to Modulate Growth Cone Turning Responses. J. Neurosci. 2010, 30, 14059–14067. [Google Scholar] [CrossRef] [Green Version]
- Agarwal-Mawal, A.; Qureshi, H.Y.; Cafferty, P.W.; Yuan, Z.; Han, D.; Lin, R.; Paudel, H.K. 14-3-3 Connects Glycogen Synthase Kinase-3β to Tau within a Brain Microtubule-associated Tau Phosphorylation Complex. J. Biol. Chem. 2003, 278, 12722–12728. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Lv, P.; Sun, Z.; Han, L.; Luo, B.; Zhou, W. 14-3-3ζ up-regulates hypoxia-inducible factor-1α in hepatocellular carcinoma via activation of PI3K/Akt/NF-кB signal transduction pathway. Int. J. Clin. Exp. Pathol. 2015, 8, 15845–15853. [Google Scholar]
- Poplawski, G.; Tranziska, A.-K.; Leshchyns’Ka, I.; Meier, I.D.; Streichert, T.; Sytnyk, V.; Schachner, M. L1CAM increases MAP2 expression via the MAPK pathway to promote neurite outgrowth. Mol. Cell. Neurosci. 2012, 50, 169–178. [Google Scholar] [CrossRef]
- Katherine, K.; Dent, W.E. Developing vertebrate CNS. Nat. Rev. Neurosci. 2014, 15, 7–18. [Google Scholar] [CrossRef]
- Homma, N.; Takei, Y.; Tanaka, Y.; Nakata, T.; Terada, S.; Kikkawa, M.; Noda, Y.; Hirokawa, N. Kinesin superfamily protein 2A (KIF2A) functions in suppression of collateral branch extension. Cell 2003, 114, 229–239. [Google Scholar] [CrossRef] [Green Version]
- Garnham, C.P.; Roll-Mecak, A. The chemical complexity of cellular microtubules: Tubulin post-translational modification enzymes and their roles in tuning microtubule functions. Cytoskeleton 2012, 69, 442–463. [Google Scholar] [CrossRef] [Green Version]
- Halpain, S.; Dehmelt, L. The MAP1 family of microtubule-associated proteins. Genome Biol. 2006, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Cleveland, D. Neuronal Intermediate Filaments. Annu. Rev. Neurosci. 1996, 19, 187–217. [Google Scholar] [CrossRef]
- Dent, E.W.; Gupton, S.L.; Gertler, F.B. The Growth Cone Cytoskeleton in Axon Outgrowth and Guidance. Cold Spring Harb. Perspect. Biol. 2010, 3, a001800. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.A.; Wilson, M.H.; Giraud, J.; Xie, Z.; Tseng, H.-C.; England, C.; Herscovitz, H.; Tsai, L.-H.; Delalle, I. Capzb2 Interacts with β-Tubulin to Regulate Growth Cone Morphology and Neurite Outgrowth. PLoS Biol. 2009, 7, e1000208. [Google Scholar] [CrossRef] [Green Version]
- Mohan, R.; John, A. Microtubule-associated proteins as direct crosslinkers of actin filaments and microtubules. IUBMB Life 2015, 67, 395–403. [Google Scholar] [CrossRef]
- Sánchez-Huertas, C.; Lüders, J. The Augmin Connection in the Geometry of Microtubule Networks. Curr. Biol. 2015, 25, R294–R299. [Google Scholar] [CrossRef] [Green Version]
- Kuijpers, M.; Hoogenraad, C.C. Centrosomes, microtubules and neuronal development. Mol. Cell. Neurosci. 2011, 48, 349–358. [Google Scholar] [CrossRef]
- Sheng, L.; Leshchyns’Ka, I.; Sytnyk, V. Cell adhesion and intracellular calcium signaling in neurons. Cell Commun. Signal. 2013, 11, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leshchyns’Ka, I.; Sytnyk, V. Reciprocal Interactions between Cell Adhesion Molecules of the Immunoglobulin Superfamily and the Cytoskeleton in Neurons. Front. Cell Dev. Biol. 2016, 4, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waites, C.L.; Craig, A.M.; Garner, C. Mechanisms of Vertebrate Synaptogenesis. Annu. Rev. Neurosci. 2005, 28, 251–274. [Google Scholar] [CrossRef] [Green Version]
- Stanika, R.; Campiglio, M.; Pinggera, A.; Lee, A.; Striessnig, J.; Flucher, B.E.; Obermair, G.J. Splice variants of the CaV1.3 L-type calcium channel regulate dendritic spine morphology. Sci. Rep. 2016, 6, 34528. [Google Scholar] [CrossRef] [Green Version]
- Leitch, B.; Szostek, A.; Lin, R.; Shevtsova, O. Subcellular distribution of L-type calcium channel subtypes in rat hippocampal neurons. Neuroscience 2009, 164, 641–657. [Google Scholar] [CrossRef]
- Sheng, L.; Leshchyns’Ka, I.; Sytnyk, V. Neural Cell Adhesion Molecule 2 (NCAM2)-Induced c-Src-Dependent Propagation of Submembrane Ca2+ Spikes Along Dendrites Inhibits Synapse Maturation. Cereb. Cortex 2019, 29, 1439–1459. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Bosch, M.; Hayashi, Y. The Roles of CaMKII and F-Actin in the Structural Plasticity of Dendritic Spines: A Potential Molecular Identity of a Synaptic Tag? Physiology 2009, 24, 357–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, K.-I.; Narayanan, R.; Lee, S.H.; Murata, K.; Hayashi, Y. The role of CaMKII as an F-actin-bundling protein crucial for maintenance of dendritic spine structure. Proc. Natl. Acad. Sci. USA 2007, 104, 6418–6423. [Google Scholar] [CrossRef] [Green Version]
- Hell, J.W. CaMKII: Claiming Center Stage in Postsynaptic Function and Organization. Neuron 2014, 81, 249–265. [Google Scholar] [CrossRef] [Green Version]
- Dillon, C.; Goda, Y. The Actin Cytoskeleton: Integrating Form and Function at the Synapse. Annu. Rev. Neurosci. 2005, 28, 25–55. [Google Scholar] [CrossRef]
- Eyang, Y.; Ecalakos, N. Presynaptic long-term plasticity. Front. Synaptic Neurosci. 2013, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Edwards, M.; Zwolak, A.; Schafer, D.A.; Sept, D.; Dominguez, R.; Cooper, J.A. Capping protein regulators fine-tune actin assembly dynamics. Nat. Rev. Mol. Cell Biol. 2014, 15, 677–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimura, Y.; Yamauchi, Y.; Shinkawa, T.; Taoka, M.; Donai, H.; Takahashi, N.; Isobe, T.; Yamauchi, T. Molecular constituents of the postsynaptic density fraction revealed by proteomic analysis using multidimensional liquid chromatography-tandem mass spectrometry. J. Neurochem. 2003, 88, 759–768. [Google Scholar] [CrossRef]
- Kitanishi, T.; Sakai, J.; Kojima, S.; Saitoh, Y.; Inokuchi, K.; Fukaya, M.; Watanabe, M.; Matsuki, N.; Yamada, M.K. Activity-dependent localization in spines of the F-actin capping protein CapZ screened in a rat model of dementia. Genes Cells 2010, 15, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Tang, X.; Vitriol, E.; Chen, G.; Zheng, J.Q. Actin Capping Protein Is Required for Dendritic Spine Development and Synapse Formation. J. Neurosci. 2011, 31, 10228–10233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, M.; Hikita, T.; Taya, S.; Uraguchi-Asaki, J.; Toyooka, K.; Wynshaw-Boris, A.; Ujike, H.; Inada, T.; Takao, K.; Miyakawa, T.; et al. Identification of YWHAE, a gene encoding 14-3-3epsilon, as a possible susceptibility gene for schizophrenia. Hum. Mol. Genet. 2008, 17, 3212–3222. [Google Scholar] [CrossRef] [Green Version]
- Wachi, T.; Cornell, B.; Toyooka, K. Complete ablation of the 14-3-3epsilon protein results in multiple defects in neuropsychiatric behaviors. Behav. Brain Res. 2017, 319, 31–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foote, M.; Qiao, H.; Graham, K.; Wu, Y.; Zhou, Y. Inhibition of 14-3-3 Proteins Leads to Schizophrenia-Related Behavioral Phenotypes and Synaptic Defects in Mice. Biol. Psychiatry 2015, 78, 386–395. [Google Scholar] [CrossRef] [Green Version]
- Dent, E.W. Of microtubules and memory: Implications for microtubule dynamics in dendrites and spines. Mol. Biol. Cell 2017, 28, 1–8. [Google Scholar] [CrossRef]
- Caceres, A.; Payne, M.R.; Binder, L.I.; Steward, O. Immunocytochemical localization of actin and microtubule-associated protein MAP2 in dendritic spines. Proc. Natl. Acad. Sci. USA 1983, 80, 1738–1742. [Google Scholar] [CrossRef] [Green Version]
- Conde, C.; Cáceres, A. Microtubule assembly, organization and dynamics in axons and dendrites. Nat. Rev. Neurosci. 2009, 10, 319–332. [Google Scholar] [CrossRef]
- Tortosa, E.; Montenegro-Venegas, C.; Benoist, M.; Härtel, S.; González-Billault, C.; Esteban, J.A.; Avila, J. Microtubule-associated Protein 1B (MAP1B) Is Required for Dendritic Spine Development and Synaptic Maturation. J. Biol. Chem. 2011, 286, 40638–40648. [Google Scholar] [CrossRef] [Green Version]
- Sousa, R.; Lafer, E.M. The role of molecular chaperones in clathrin mediated vesicular trafficking. Front. Mol. Biosci. 2015, 2, 26. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, S.N.; Martin, M.D.; Akoury, E.; Assimon, V.A.; Borysov, S.; Nordhues, B.A.; Sabbagh, J.J.; Cockman, M.; Gestwicki, J.E.; Zweckstetter, M.; et al. The active Hsc70/tau complex can be exploited to enhance tau turnover without damaging microtubule dynamics. Hum. Mol. Genet. 2015, 24, 3971–3981. [Google Scholar] [CrossRef] [Green Version]
- Leshchyns’Ka, I.; Sytnyk, V.; Richter, M.; Andreyeva, A.; Puchkov, D.; Schachner, M. The Adhesion Molecule CHL1 Regulates Uncoating of Clathrin-Coated Synaptic Vesicles. Neuron 2006, 52, 1011–1025. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Raiker, S.J.; Venkatesh, K.; Geary, R.; Robak, L.A.; Zhang, Y.; Yeh, H.H.; Shrager, P.; Giger, R.J. Synaptic Function for the Nogo-66 Receptor NgR1: Regulation of Dendritic Spine Morphology and Activity-Dependent Synaptic Strength. J. Neurosci. 2008, 28, 2753–2765. [Google Scholar] [CrossRef] [Green Version]
- Raiker, S.J.; Lee, H.; Baldwin, K.; Duan, Y.; Shrager, P.; Giger, R.J. Oligodendrocyte-Myelin Glycoprotein and Nogo Negatively Regulate Activity-Dependent Synaptic Plasticity. J. Neurosci. 2010, 30, 12432–12445. [Google Scholar] [CrossRef] [PubMed]
- Zagrebelsky, M.; Schweigreiter, R.; Bandtlow, C.E.; Schwab, M.E.; Korte, M. Nogo-A Stabilizes the Architecture of Hippocampal Neurons. J. Neurosci. 2010, 30, 13220–13234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pernet, V.; Schwab, M.E. The role of Nogo—A in axonal plasticity, regrowth and repair. Cell Tissue Res. 2012, 349, 97–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajor, M.; Kaczmarek, L. Proteolytic Remodeling of the Synaptic Cell Adhesion Molecules (CAMs) by Metzincins in Synaptic Plasticity. Neurochem. Res. 2012, 38, 1113–1121. [Google Scholar] [CrossRef] [Green Version]
- Krueger, D.D.; Tuffy, L.P.; Papadopoulos, T.; Brose, N. The role of neurexins and neuroligins in the formation, maturation, and function of vertebrate synapses. Curr. Opin. Neurobiol. 2012, 22, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Hirano, S.; Takeichi, M. Cadherins in Brain Morphogenesis and Wiring. Physiol. Rev. 2012, 92, 597–634. [Google Scholar] [CrossRef]
- Molloy, C.A.; Keddache, M.; Martin, L.J. Evidence for linkage on 21q and 7q in a subset of autism characterized by developmental regression. Mol. Psychiatry 2005, 10, 741–746. [Google Scholar] [CrossRef] [Green Version]
- Scholz, C.; Steinemann, D.; Mälzer, M.; Roy, M.; Arslan-Kirchner, M.; Illig, T.; Schmidtke, J.; Stuhrmann, M. NCAM2 deletion in a boy with macrocephaly and autism: Cause, association or predisposition? Eur. J. Med. Genet. 2016, 59, 493–498. [Google Scholar] [CrossRef]
- Winther, M.; Berezin, V.; Walmod, P.S. NCAM2/OCAM/RNCAM: Cell adhesion molecule with a role in neuronal com-partmentalization. Int. J. Biochem. Cell Biol. 2012, 44, 441–446. [Google Scholar] [CrossRef]
- Petit, F.; Plessis, G.; DeCamp, M.; Cuisset, J.-M.; Blyth, M.; Pendlebury, M.; Andrieux, J. 21q21 deletion involving NCAM2: Report of 3 cases with neurodevelopmental disorders. Eur. J. Med. Genet. 2015, 58, 44–46. [Google Scholar] [CrossRef] [PubMed]
- Han, M.-R.; Schellenberg, G.D.; Wang, L.-S.; Initiative, T.A.D.N. Genome-wide association reveals genetic effects on human Aβ 42 and τ protein levels in cerebrospinal fluids: A case control study. BMC Neurol. 2010, 10, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, R.; Kamino, K.; Yamamoto, M.; Nuripa, A.; Kida, T.; Kazui, H.; Hashimoto, R.; Tanaka, T.; Kudo, T.; Yamagata, H.; et al. The DYRK1A gene, encoded in chromosome 21 Down syndrome critical region, bridges between β-amyloid production and tau phosphorylation in Alzheimer disease. Hum. Mol. Genet. 2007, 16, 15–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niederhoffer, K.Y.; Fahiminiya, S.; Eydoux, P.; Mawson, J.; Nishimura, G.; Jerome-Majewska, L.A.; Patel, M.S. Diagnosis of Van den Ende-Gupta syndrome: Approach to the Marden-Walker-like spectrum of disorders. Am. J. Med. Genet. Part A 2016, 170, 2310–2321. [Google Scholar] [CrossRef]
- Griesi-Oliveira, K.; Suzuki, A.M.; Alves, A.Y.; Mafra, A.C.C.N.; Yamamoto, G.L.; Ezquina, S.; Magalhães, Y.T.; Forti, F.L.; Sertie, A.L.; Zachi, E.C.; et al. Actin cytoskeleton dynamics in stem cells from autistic individuals. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Torrico, B.; Antón-Galindo, E.; Fernàndez-Castillo, N.; Rojo-Francàs, E.; Ghorbani, S.; Pineda-Cirera, L.; Hervás, A.; Rueda, I.; Moreno, E.; Fullerton, J.M.; et al. Involvement of the 14-3-3 Gene Family in Autism Spectrum Disorder and Schizophrenia: Genetics, Transcriptomics and Functional Analyses. J. Clin. Med. 2020, 9, 1851. [Google Scholar] [CrossRef] [PubMed]
- Mukaetova-Ladinska, E.B.; Arnold, H.; Jaros, E.; Perry, R.; Perry, E. Depletion of MAP2 expression and laminar cytoarchitectonic changes in dorsolateral prefrontal cortex in adult autistic individuals. Neuropathol. Appl. Neurobiol. 2004, 30, 615–623. [Google Scholar] [CrossRef]
- Pham, E.; Crews, L.; Ubhi, K.; Hansen, L.; Adame, A.; Cartier, A.; Salmon, D.; Galasko, D.; Michael, S.; Savas, J.N.; et al. Progressive accumulation of amyloid-β oligomers in Alzheimer’s disease and in amyloid precursor protein transgenic mice is accompanied by selective alterations in synaptic scaffold proteins. FEBS J. 2010, 277, 3051–3067. [Google Scholar] [CrossRef] [Green Version]
- Penzes, P.; VanLeeuwen, J.-E. Impaired regulation of synaptic actin cytoskeleton in Alzheimer’s disease. Brain Res. Rev. 2011, 67, 184–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheff, S.W.; Price, D.A. Synaptic pathology in Alzheimer’s disease: A review of ultrastructural studies. Neurobiol. Aging 2003, 24, 1029–1046. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J. Alzheimer’s Disease Is a Synaptic Failure. Science 2002, 298, 789–791. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.R.; Damiano, J.; Franceschetti, S.; Carpenter, S.; Canafoglia, L.; Morbin, M.; Rossi, G.; Pareyson, D.; Mole, S.; Staropoli, J.F.; et al. Strikingly Different Clinicopathological Phenotypes Determined by Progranulin-Mutation Dosage. Am. J. Hum. Genet. 2012, 90, 1102–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Autism Spectrum Disorder | |||
|---|---|---|---|
| Molecule | Type of Study | Implications | References |
| CDHs | Genetic in humans | CNVs and SNPs in CDHs genes found in ASD patients. | [19,20] |
| PDCH | Genetic in humans | Alterations in PCDH9, PCDH10 and PCDH19 genes in patients with autism. | [21,22] |
| FAT1 | Genetic in humans | Genetic modifications in FAT1 in Autism Spectrum Disorder patients. | [23,24,25,26] |
| NRXN | Genetic in humans | Mutations and CNVs in NRXN1-3 genes are associated with ASD. | [27,28] |
| Experimental in mouse | NRXN1 deletion causes electrophysiological and behavioral changes consistent with cognitive impairments. | [29] | |
| NLGN | Genetic in humans | Genetic modifications in NLGN1-4 genes found in ASD patients. | [30,31] |
| Experimental in mice models | Ngln3 and Ngln4 KO mice displayed reduced social interaction | [32] | |
| CNTNAP2 | Genetic in humans | Genetic alterations in ASD patients | [33,34] |
| Experimental in mice models | CNTNAP2 deficient mice present deficits in communication and social interaction; and repetitive behaviors. | [35,36,37] | |
| CNTN | Genetic in humans | CNTN3–6 are considered as gene risk for ASD. | [21,30] |
| Schizophrenia | |||
| Molecule | Type of Study | Implications | References |
| NCAM1 | Genetic in humans | SNPs in NCAM1 in schizophrenia and bipolar patients. | [38,39] |
| NLGN1 | GWAS study | NLGN1 contributed to schizophrenia susceptibility in Han Chinese population. | [17,40] |
| Selectin | Proteomic in humans | Reduced levels in plasma from adolescents with early-onset psycosis. | [41] |
| VCAM1 | Proteomic in humans | Reduced levels in plasma from adolescents with early-onset psycosis. | [41] |
| Epilepsy | |||
| Molecule | Type of Study | Implications | References |
| N-cadherin | Mouse model | N-cadherin reduction changes mature excitatory and inhibitory circuits and contribute to significant impairment in spatial memory | [42] |
| Mouse model | N-cadherin antibody alleviate brain pathology | [43] | |
| DSCAML1 | Genetic in humans | single nucleotide substitution resulting in its loss of function of DSCAML1. | [44,45] |
| Rat model | GABAergic neurons were reduced and neurons’ excitability was enhanced. | [46] | |
| NCAM1 | Proteomics in CSF | NCAM1 concentration in CSF is lower in epilepsy patients. | [47] |
| β-integrin | Mouse model | Seizure activity and nervous system hyperexcitability. | [48] |
| Fragile X Syndrome | |||
| Molecule | Type of Study | Implications | References |
| DSCAML1 | Mouse model | Dosage variations of DSCAML1 disrupt cell–cell and cell–environment interactions crucial for neuronal migration and brain formation | [49] |
| NLGNs | Mouse model | FMRP controls the synaptic level of NLGNs | [50] |
| Down Syndrome | |||
| Molecule | Type of Study | Implications | References |
| DSCAML1 | Mouse model | Overexpression of DSCAM led to the inhibition of dendritic branching, a phenotype observed in DS patients. | [51] |
| IPSC from DS patients | DSCAM/PAK1 pathway suppression reverses neurogenesis deficits. | [52] | |
| Extracellular Region | |
|---|---|
| Interaction | References |
| Fibroblast Growth Factor Receptor (FGFR) | [80] |
| Epidermal Growth Factor Receptor (EGFR) | [79] |
| Nogo | [70] |
| Granulin | [70] |
| Prion protein (Prp) | [81,82] |
| Beta-site APP cleaving enzyme 1 (BACE1) | [75] |
| Post-translational modification | |
| N-glycosylation | [78] |
| Proteolytic cleavage | [75,76] |
| Intracellular region | |
| Interaction | References |
| Proto-oncogene tyrosine-protein kinase Src | [83] |
| Calcium/calmodulin-dependent protein kinase type II (CaMKII) | [69,83] |
| Microtubule-associated protein 2 (MAP2) | [69] |
| Actin | [70] |
| Tubulin | [69] |
| 14-3-3 family proteins | [69,81,82] |
| Microtubule-associated protein 1B (MAP1B) | [70] |
| F-actin-capping complex (CAPZ) | [70] |
| Heat shock cognate 71 protein (HSC70) | [70] |
| Postranslational modification | |
| Palmitoylation | [70,74] |
| Phosphorylation | [73] |
| Neurodevelopmental Disorder | |||
|---|---|---|---|
| Disorder | Type of Study | Implications | References |
| Autism Spectrum Disorders | Genetic in humans | Genetic studies associate alterations and deletions in Ncam2 with ASD. | [23,164,165] |
| Down Syndrome | Genetic in humans | Increased expression in DS patients due to the location of Ncam2 in the 21 chromosome. | [67,166] |
| Other neurodevelopmental disorders | Genetic in humans | Deletions Ncam2 are found in patients with neurodevelpmental disorders. | [167] |
| Neurodegenerative diseases | |||
| Disorder | Implications | References | |
| Alzheimer’s Disease | Genetic in humans | Alterations in Ncam2 found in AD patients. | [168,169] |
| Experimental with human and mouse samples. | β-amyloid induces proteolysis of synaptic NCAM2. | [76] | |
| Frontotemporal dementia | Experimental with mouse tissue samples | NCAM2 proposed as a candidate receptor for GRN | [70] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parcerisas, A.; Ortega-Gascó, A.; Pujadas, L.; Soriano, E. The Hidden Side of NCAM Family: NCAM2, a Key Cytoskeleton Organization Molecule Regulating Multiple Neural Functions. Int. J. Mol. Sci. 2021, 22, 10021. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810021
Parcerisas A, Ortega-Gascó A, Pujadas L, Soriano E. The Hidden Side of NCAM Family: NCAM2, a Key Cytoskeleton Organization Molecule Regulating Multiple Neural Functions. International Journal of Molecular Sciences. 2021; 22(18):10021. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810021
Chicago/Turabian StyleParcerisas, Antoni, Alba Ortega-Gascó, Lluís Pujadas, and Eduardo Soriano. 2021. "The Hidden Side of NCAM Family: NCAM2, a Key Cytoskeleton Organization Molecule Regulating Multiple Neural Functions" International Journal of Molecular Sciences 22, no. 18: 10021. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810021