
The Relationship between the Spatial Arrangement of Pigments and Exciton Transition Moments in Photosynthetic Light-Harvesting Complexes



Abstract
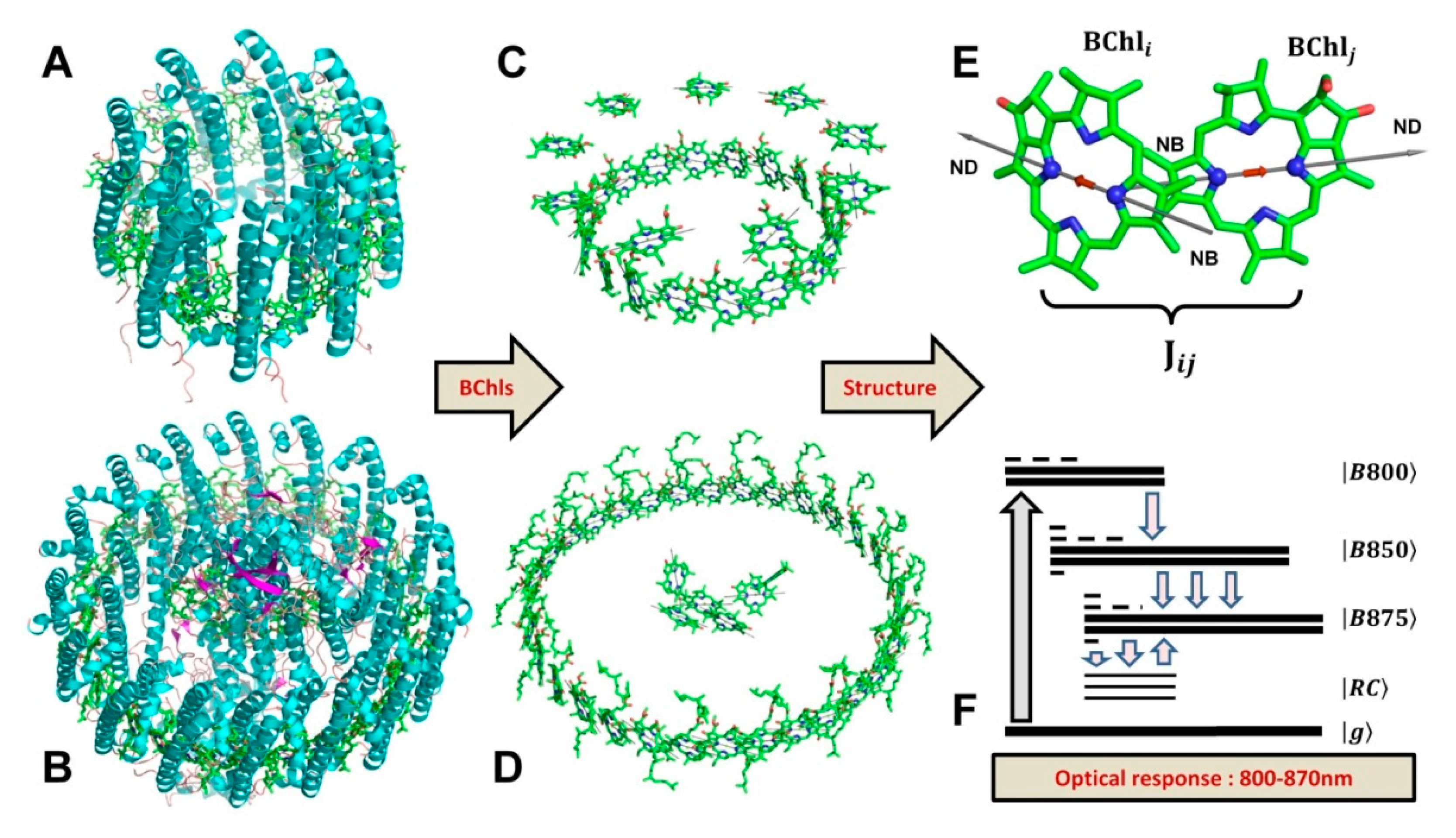
:1. Introduction
2. Theory
3. Results
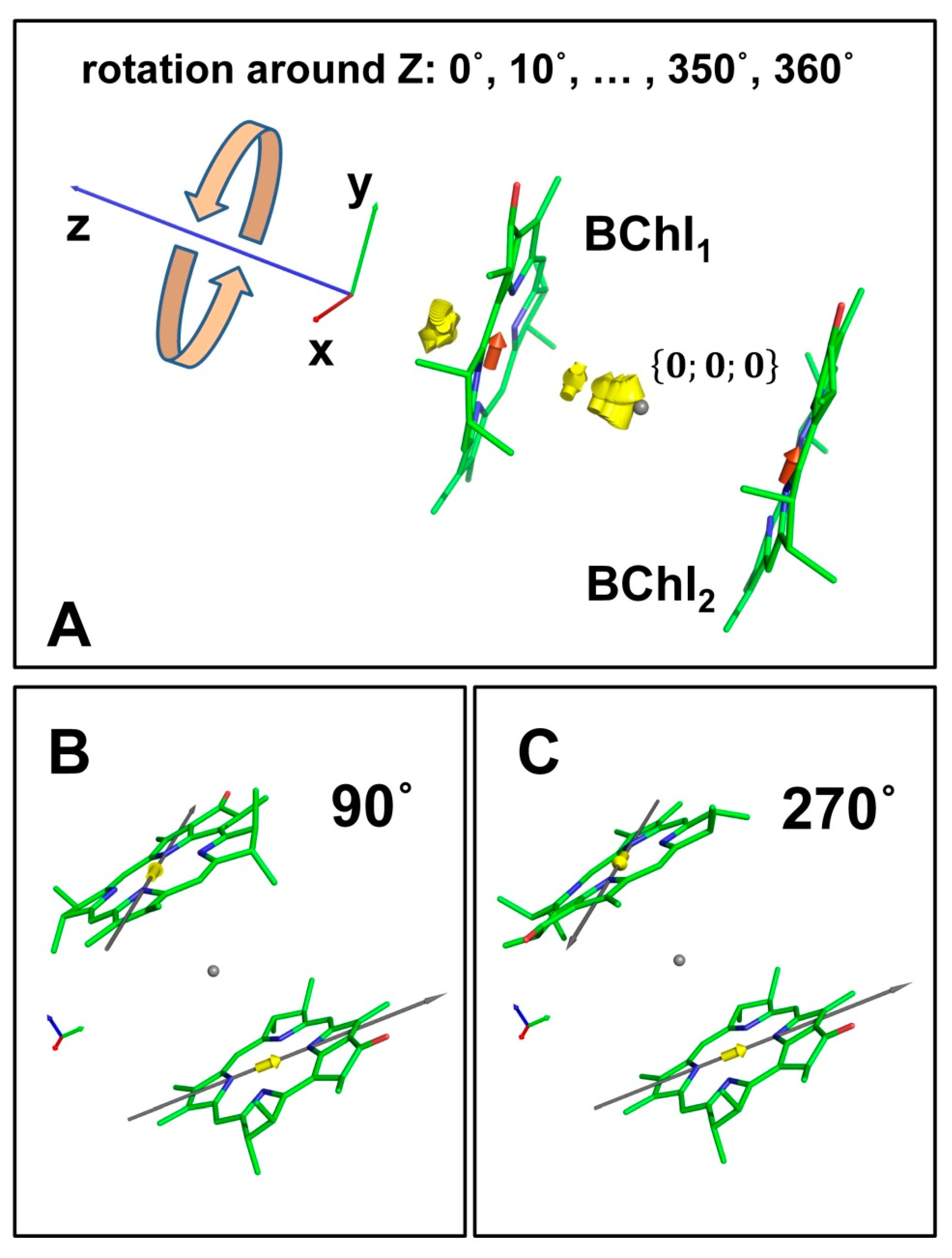
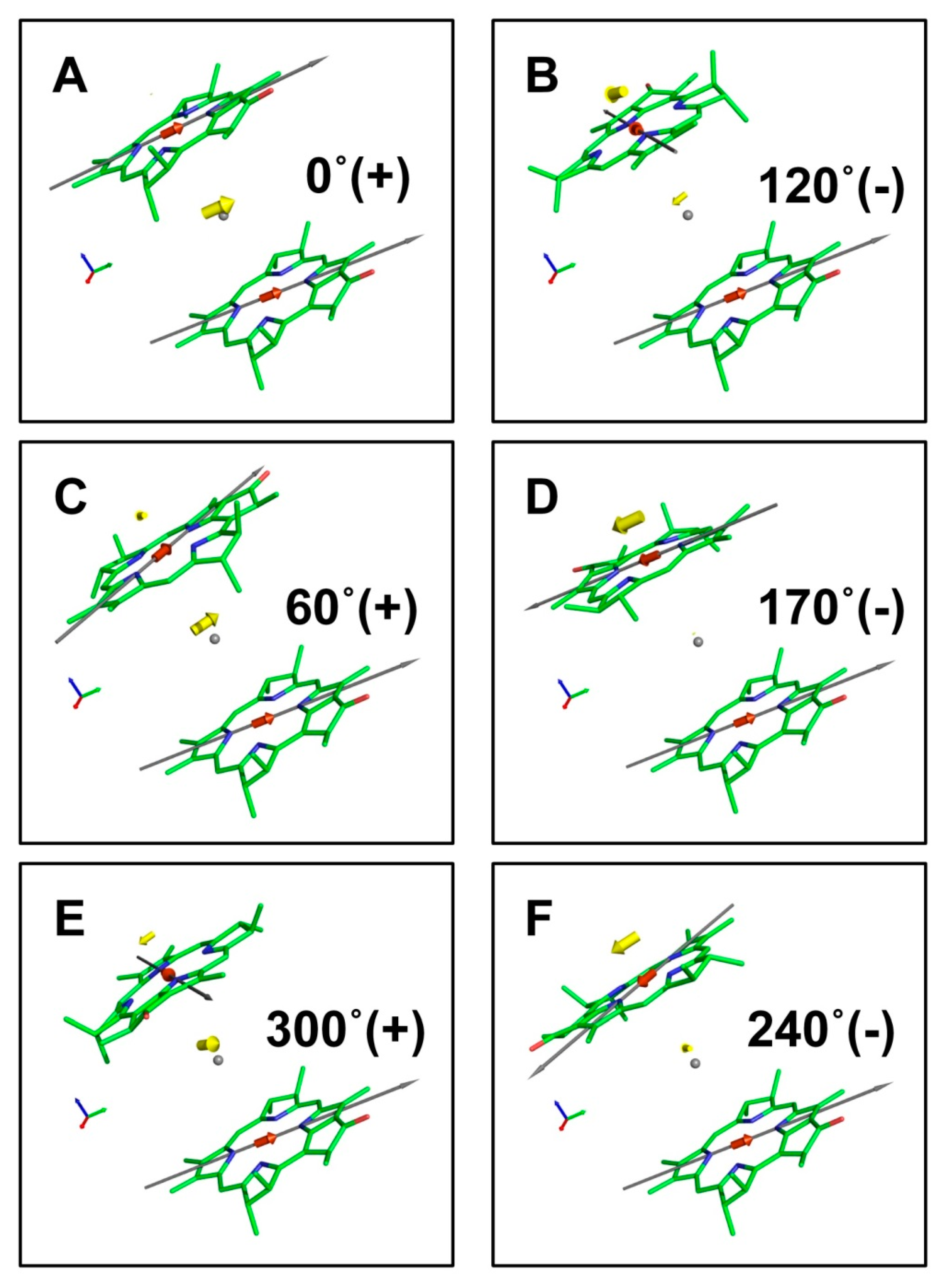
3.1. Dimer of BChl Molecules
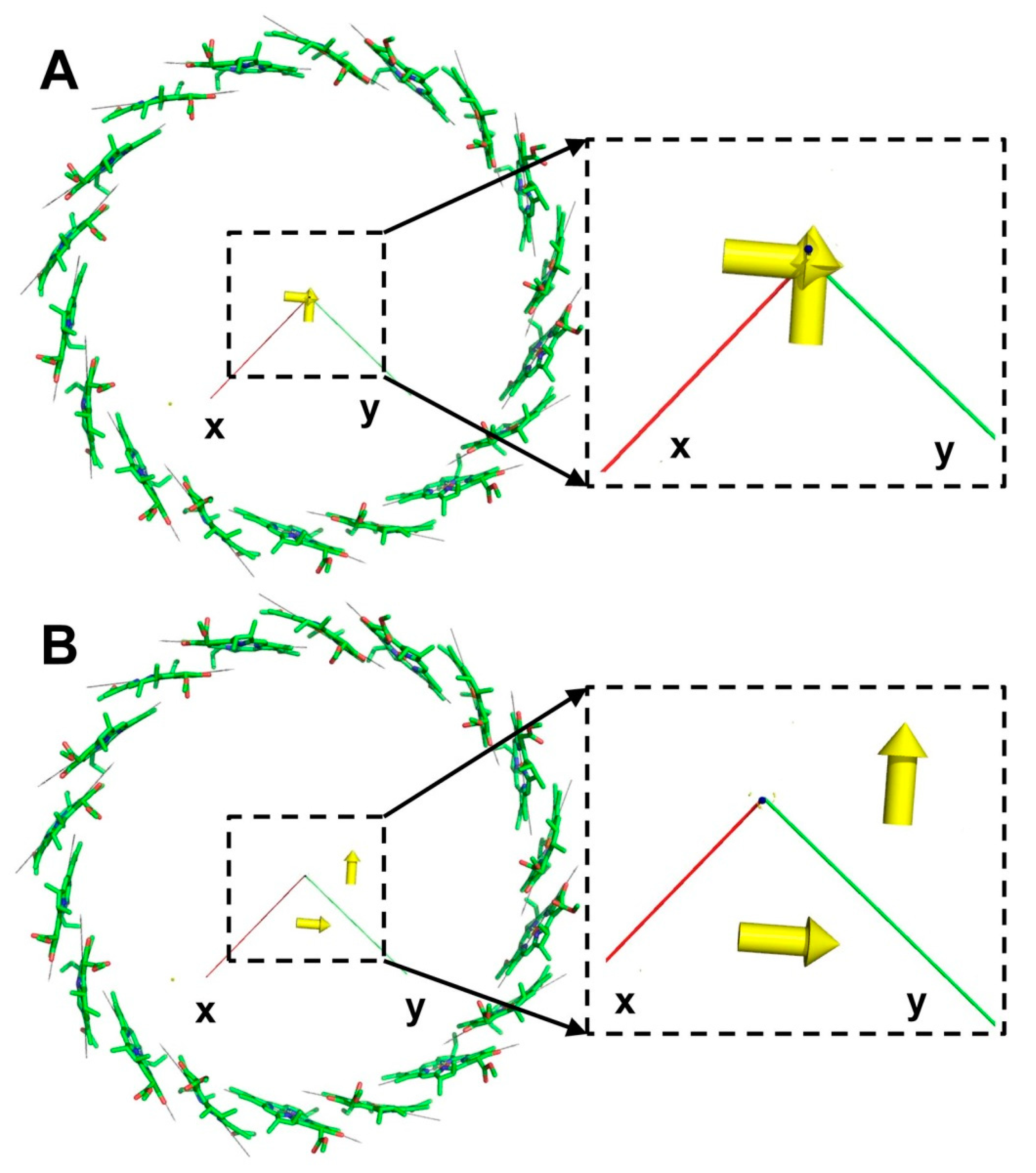
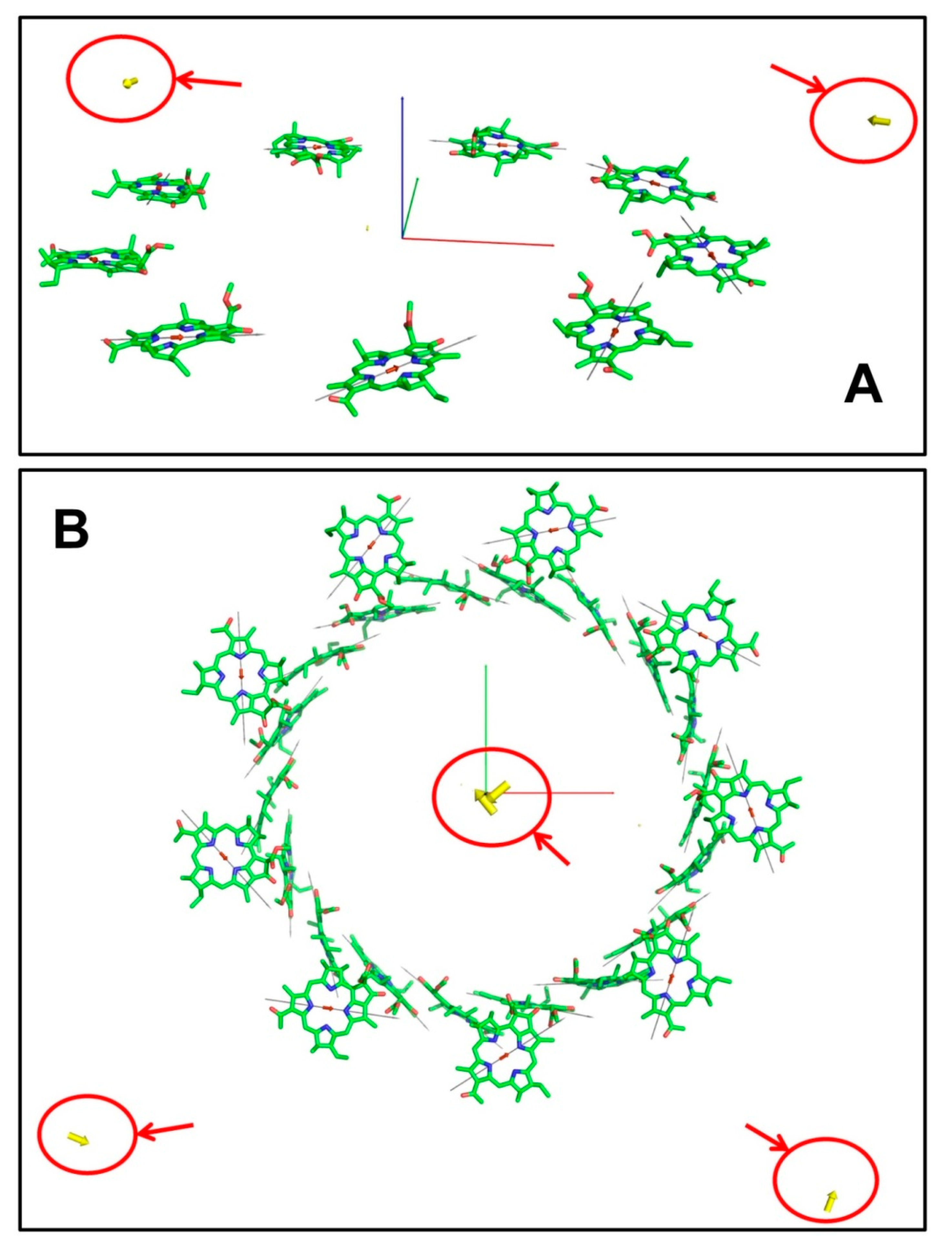
3.2. B850 Ring
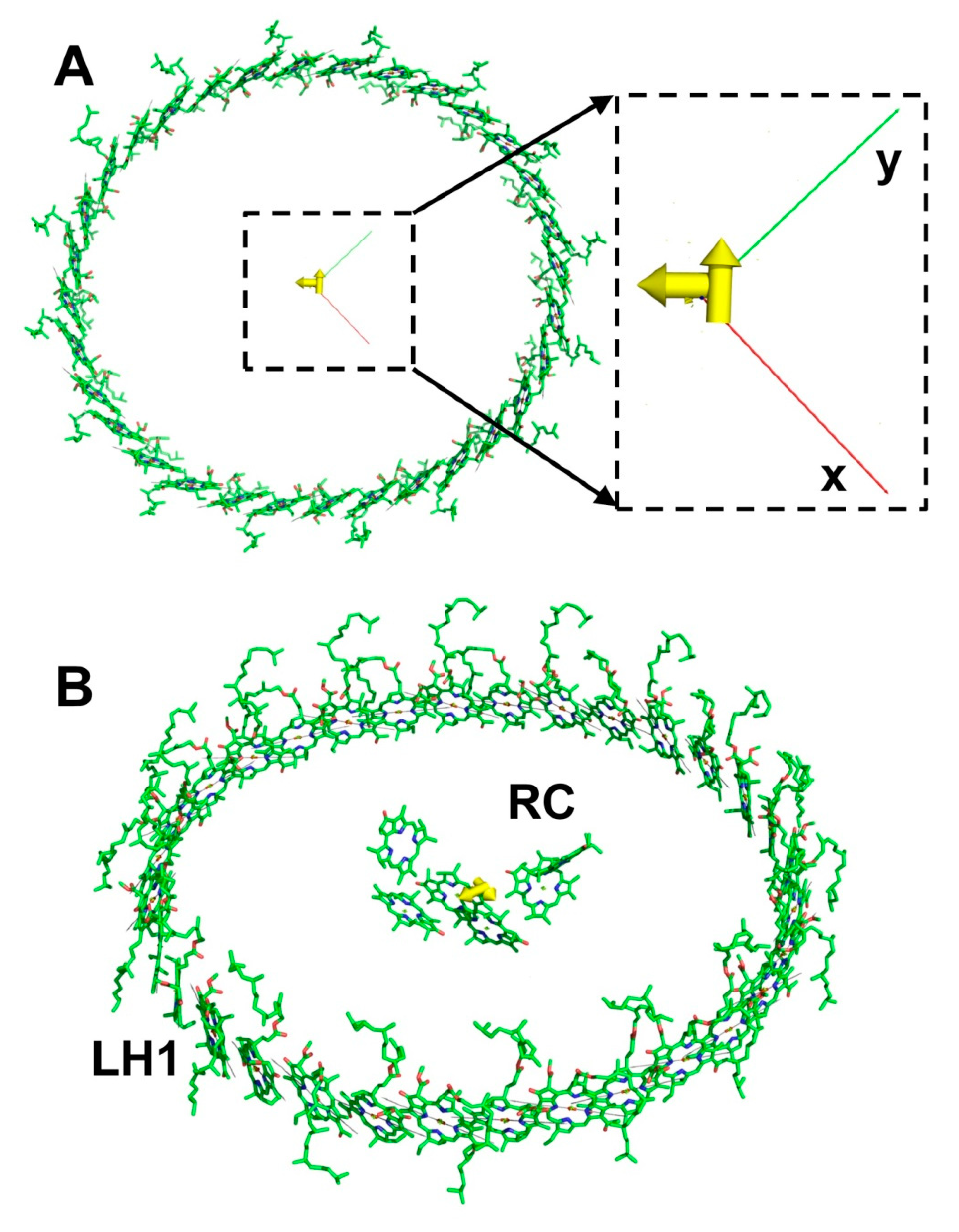
3.3. LH2 and LH1 Complexes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0° | 30° | 60° | 120° | 170° | 240° | 300° | 330° | |
|---|---|---|---|---|---|---|---|---|
| 188.41 | 163.14 | 94.18 | −94.24 | −185.53 | −94.17 | 94.21 | 163.18 | |
| 7.06 | 7.05 | 7.01 | 0.91 | 0.47 | 0.91 | 7.01 | 7.05 | |
| 0.47 | 0.54 | 0.91 | 7.01 | 7.06 | 7.01 | 0.91 | 0.54 | |
| 0.01 | 0.23 | 0.84 | 0.84 | 0.04 | 0.84 | 0.84 | 0.23 | |
| 3.23 | 3.01 | 2.41 | 2.41 | 3.21 | 2.41 | 2.41 | 3.01 |
| B601 | C502 | D602 | E503 | F603 | G504 | H604 | I505 | J605 | K506 | L606 | M507 | N607 | O508 | P608 | R509 | S609 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A501 | 388.9 | −53.7 | 14.0 | −6.4 | 3.0 | −2.0 | 1.2 | −1.0 | 0.7 | −1.1 | 0.9 | −2.0 | 2.5 | −6.6 | 12.5 | −52.3 | 333.3 |
| B601 | 360.5 | −35.2 | 12.4 | −4.1 | 2.6 | −1.1 | 1.0 | −0.4 | 0.8 | −0.4 | 1.1 | −1.0 | 3.1 | −4.2 | 14.4 | −35.8 | |
| C502 | 366.1 | −53.1 | 14.4 | −6.7 | 3.2 | −2.0 | 1.2 | −1.1 | 0.7 | −1.0 | 1.0 | −2.0 | 2.5 | −6.6 | 12.3 | ||
| D602 | 350.3 | −35.2 | 12.6 | −4.0 | 2.5 | −0.9 | 0.9 | −0.3 | 0.7 | −0.3 | 1.1 | −1.0 | 3.0 | −3.8 | |||
| E503 | 354.2 | −52.0 | 13.9 | −6.3 | 2.8 | −1.9 | 1.0 | −0.9 | 0.6 | −1.0 | 0.9 | −1.9 | 2.3 | ||||
| F603 | 381.7 | −37.6 | 12.8 | −4.1 | 2.5 | −0.9 | 0.9 | −0.4 | 0.7 | −0.4 | 1.2 | −1.0 | |||||
| G504 | 381.5 | −54.9 | 14.7 | −6.8 | 3.2 | −2.0 | 1.2 | −1.1 | 0.8 | −1.1 | 1.0 | ||||||
| H604 | 402.3 | −39.7 | 13.7 | −4.4 | 2.7 | −1.2 | 1.1 | −0.5 | 0.8 | −0.5 | |||||||
| I505 | 378.4 | −53.7 | 14.1 | −6.5 | 3.1 | −2.0 | 1.2 | −1.0 | 0.7 | ||||||||
| J605 | 340.9 | −33.1 | 11.9 | −3.9 | 2.4 | −1.0 | 0.9 | −0.4 | |||||||||
| K506 | 364.2 | −53.2 | 14.5 | −6.6 | 3.1 | −2.0 | 1.2 | ||||||||||
| L606 | 359.7 | −36.2 | 12.4 | −4.0 | 2.5 | −0.9 | |||||||||||
| M507 | 379.9 | −53.5 | 14.2 | −6.5 | 3.0 | ||||||||||||
| N607 | 355.2 | −36.4 | 12.8 | −4.0 | |||||||||||||
| O508 | 378.3 | −54.9 | 14.4 | ||||||||||||||
| P608 | 389.6 | −36.6 | |||||||||||||||
| R509 | 362.4 |
| C702 | E703 | G704 | I705 | K706 | M707 | O708 | R709 | |
|---|---|---|---|---|---|---|---|---|
| A701 | −24.9 | −2.8 | −0.8 | −0.3 | −0.3 | −0.7 | −2.7 | −23.7 |
| C702 | −24.2 | −2.9 | −0.7 | −0.3 | −0.3 | −0.7 | −2.7 | |
| E703 | −24.4 | −2.8 | −0.7 | −0.3 | −0.3 | −0.7 | ||
| G704 | −25.5 | −2.9 | −0.7 | −0.3 | −0.3 | |||
| I705 | −24.2 | −2.6 | −0.7 | −0.3 | ||||
| K706 | −23.3 | −2.6 | −0.7 | |||||
| M707 | −22.7 | −2.6 | ||||||
| O708 | −23.4 |
References
- Jang, S.J.; Mennucci, B. Delocalized excitons in natural light-harvesting complexes. Rev. Mod. Phys. 2018, 90, 035003. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.C.; Ritz, T.; Damjanovic, A.; Autenrieth, F.; Schulten, K. Photosynthetic apparatus of purple bacteria. Q. Rev. Biophys. 2002, 35, 1–62. [Google Scholar] [CrossRef] [Green Version]
- Croce, R.; van Amerongen, H. Natural strategies for photosynthetic light harvesting. Nat. Chem. Biol. 2014, 10, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Cogdell, R.J.; Gall, A.; Kohler, J. The architecture and function of the light-harvesting apparatus of purple bacteria: From single molecules to in vivo membranes. Q. Rev. Biophys. 2006, 39, 227–324. [Google Scholar] [CrossRef] [PubMed]
- Mirkovic, T.; Ostroumov, E.E.; Anna, J.M.; van Grondelle, R.; Govindjee; Scholes, G.D. Light Absorption and Energy Transfer in the Antenna Complexes of Photosynthetic Organisms. Chem. Rev. 2017, 117, 249–293. [Google Scholar] [CrossRef]
- Van Grondelle, R.; Novoderezhkin, V.I. Energy transfer in photosynthesis: Experimental insights and quantitative models. Phys. Chem. Chem. Phys. 2006, 8, 793–807. [Google Scholar] [CrossRef]
- McLuskey, K.; Prince, S.M.; Cogdell, R.J.; Isaacs, N.W. The crystallographic structure of the B800-820 LH3 light-harvesting complex from the purple bacteria Rhodopseudomonas acidophila strain 7050. Biochemistry 2001, 40, 8783–8789. [Google Scholar] [CrossRef] [PubMed]
- Koepke, J.; Hu, X.C.; Muenke, C.; Schulten, K.; Michel, H. The crystal structure of the light-harvesting complex II (B800-850) from Rhodospirillum molischianum. Structure 1996, 4, 581–597. [Google Scholar] [CrossRef] [Green Version]
- McDermott, G.; Prince, S.M.; Freer, A.A.; Hawthornthwaitelawless, A.M.; Papiz, M.Z.; Cogdell, R.J.; Isaacs, N.W. Crystal-structure of an integral membrane light-harvesting complex from photosynthetic bacteria. Nature 1995, 374, 517–521. [Google Scholar] [CrossRef]
- Qian, P.; Siebert, C.A.; Wang, P.Y.; Canniffe, D.P.; Hunter, C.N. Cryo-EM structure of the Blastochloris viridis LH1-RC complex at 2.9 angstrom. Nature 2018, 556, 203–208. [Google Scholar] [CrossRef]
- Ma, F.; Romero, E.; Jones, M.R.; Novoderezhkin, V.I.; van Grondelle, R. Vibronic Coherence in the Charge Separation Process of the Rhodobacter sphaeroides Reaction Center. J. Phys. Chem. Lett. 2018, 9, 1827–1832. [Google Scholar] [CrossRef] [Green Version]
- Mamedov, M.; Govindjee; Nadtochenko, V.; Semenov, A. Primary electron transfer processes in photosynthetic reaction centers from oxygenic organisms. Photosynth. Res. 2015, 125, 51–63. [Google Scholar] [CrossRef]
- Unterkofler, S.; Pflock, T.; Southall, J.; Cogdell, R.J.; Kohler, J. Fluorescence Blinking of the RC-LH1 Complex from Rhodopseudomonas palustris. ChemPhysChem 2011, 12, 711–716. [Google Scholar] [CrossRef]
- Gardiner, A.T.; Naydenova, K.; Castro-Hartmann, P.; Nguyen-Phan, T.C.; Russo, C.J.; Sader, K.; Hunter, C.N.; Cogdell, R.J.; Qian, P. The 2.4 angstrom cryo-EM structure of a heptameric light-harvesting 2 complex reveals two carotenoid energy transfer pathways. Sci. Adv. 2021, 7, eabe4650. [Google Scholar] [CrossRef]
- Thyrhaug, E.; Schroter, M.; Bukarte, E.; Kuhn, O.; Cogdell, R.; Hauer, J.; Zigmantas, D. Intraband dynamics and exciton trapping in the LH2 complex of Rhodopseudomonas acidophila. J. Chem. Phys. 2021, 154, 045102. [Google Scholar] [CrossRef] [PubMed]
- Caycedo-Soler, F.; Lim, J.; Oviedo-Casado, S.; van Hulst, N.F.; Huelga, S.F.; Plenio, M.B. Theory of Excitonic Delocalization for Robust Vibronic Dynamics in LH2. J. Phys. Chem. Lett. 2018, 9, 3446–3453. [Google Scholar] [CrossRef] [Green Version]
- Jordanides, X.J.; Scholes, G.D.; Fleming, G.R. The mechanism of energy transfer in the bacterial photosynthetic reaction center. J. Phys. Chem. B 2001, 105, 1652–1669. [Google Scholar] [CrossRef]
- Dahlbom, M.; Pullerits, T.; Mukamel, S.; Sandstrom, V. Exciton delocalization in the B850 light-harvesting complex: Comparison of different measures. J. Phys. Chem. B 2001, 105, 5515–5524. [Google Scholar] [CrossRef] [Green Version]
- Van Oijen, A.M.; Ketelaars, M.; Kohler, J.; Aartsma, T.J.; Schmidt, J. Spectroscopy of individual light-harvesting 2 complexes of Rhodopseudomonas acidophila: Diagonal disorder, intercomplex heterogeneity, spectral diffusion, and energy transfer in the B800 band. Biophys. J. 2000, 78, 1570–1577. [Google Scholar] [CrossRef] [Green Version]
- Novoderezhkin, V.I.; Rutkauskas, D.; van Grondelle, R. Multistate conformational model of a single LH2 complex: Quantitative picture of time-dependent spectral fluctuations. Chem. Phys. 2007, 341, 45–56. [Google Scholar] [CrossRef]
- Jimenez, R.; Dikshit, S.N.; Bradforth, S.E.; Fleming, G.R. Electronic excitation transfer in the LH2 complex of Rhodobacter sphaeroides. J. Phys. Chem. 1996, 100, 6825–6834. [Google Scholar] [CrossRef]
- Pishchalnikov, R.Y.; Razjivin, A.P. From localized excited states to excitons: Changing of conceptions of primary photosynthetic processes in the twentieth century. Biochem.-Mosc. 2014, 79, 242–250. [Google Scholar] [CrossRef]
- Van Grondelle, R. Excitation-energy transfer, trapping and annihilation in photosynthetic systems. Biochim. Biophys. Acta 1985, 811, 147–195. [Google Scholar] [CrossRef]
- Pearlstein, R.M. Exciton migration and trapping in photosynthesis. Photochem. Photobiol. 1982, 35, 835–844. [Google Scholar] [CrossRef]
- Paillotin, G. Transport and capture of electronic excitation energy in the photosynthetic apparatus. J. Theor. Biol. 1972, 36, 223–235. [Google Scholar] [CrossRef]
- Zankel, K.L.; Reed, D.W.; Clayton, R.K. Fluorescence and photochemical quenching in photosynthetic reaction centers. Proc. Natl. Acad. Sci. USA 1968, 61, 1243–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danielius, R.V.; Mineyev, A.P.; Razjivin, A.P. The cooperativity phenomena in a pigment-protein complex of light-harvesting antenna revealed by picosecond absorbance difference spectroscopy. Febs Lett. 1989, 250, 183–186. [Google Scholar] [CrossRef] [Green Version]
- Razjivin, A.; Leupold, D.; Novoderezhkin, V.; Belozersky, A.N. Exciton Delocalization in Antenna Complexes; Springer: Dordrecht, The Netherlands, 1998; pp. 15–20. [Google Scholar]
- Novoderezhkin, V.I.; Razjivin, A.P. Exciton dynamics in circular aggregates—Application to antenna of photosynthetic purple bacteria. Biophys. J. 1995, 68, 1089–1100. [Google Scholar] [CrossRef] [Green Version]
- Roszak, A.W.; Howard, T.D.; Southall, J.; Gardiner, A.T.; Law, C.J.; Isaacs, N.W.; Cogdell, R.J. Crystal structure of the RC-LH1 core complex from Rhodopseudomonas palustris. Science 2003, 302, 1969–1972. [Google Scholar] [CrossRef]
- Pishchalnikov, R.Y.; Shubin, V.V.; Razjivin, A.P. The role of vibronic modes in formation of red antenna states of cyanobacterial PSI. Photosynth. Res. 2020, 146, 75–86. [Google Scholar] [CrossRef]
- Pishchalnikov, R.Y.; Shubin, V.V.; Razjivin, A.P. Spectral differences between monomers and trimers of photosystem I depend on the interaction between peripheral chlorophylls of neighboring monomers in trimer. Phys. Wave Phenom. 2017, 25, 185–195. [Google Scholar] [CrossRef]
- Brixner, T.; Hildner, R.; Kohler, J.; Lambert, C.; Wurthner, F. Exciton Transport in Molecular Aggregates—From Natural Antennas to Synthetic Chromophore Systems. Adv. Energy Mater. 2017, 7, 1700236. [Google Scholar] [CrossRef] [Green Version]
- Renger, T.; Madjet, M.E.A.; Busch, M.S.A.; Adolphs, J.; Muh, F. Structure-based modeling of energy transfer in photosynthesis. Photosynth. Res. 2013, 116, 367–388. [Google Scholar] [CrossRef] [Green Version]
- Mukamel, S.; Abramavicius, D. Many-body approaches for simulating coherent nonlinear spectroscopies of electronic and vibrational excitons. Chem. Rev. 2004, 104, 2073–2098. [Google Scholar] [CrossRef]
- Mukamel, S. Principles of Nonlinear Optical Spectroscopy; Oxford University Press: New York, NY, USA; Oxford, UK, 1995; Volume 6, p. 543. [Google Scholar]
- Jang, S.; Dempster, S.E.; Silbey, R.J. Characterization of the static disorder in the B850 band of LH2. J. Phys. Chem. B 2001, 105, 6655–6665. [Google Scholar] [CrossRef]
- Ma, F.; Romero, E.; Jones, M.R.; Novoderezhkin, V.I.; van Grondelle, R. Both electronic and vibrational coherences are involved in primary electron transfer in bacterial reaction center. Nat. Commun. 2019, 10, 933. [Google Scholar] [CrossRef]
- Krueger, B.P.; Scholes, G.D.; Fleming, G.R. Calculation of couplings and energy-transfer pathways between the pigments of LH2 by the ab initio transition density cube method. J. Phys. Chem. B 1998, 102, 5378–5386. [Google Scholar] [CrossRef]
- Georgakopoulou, S.; Frese, R.N.; Johnson, E.; Koolhaas, C.; Cogdell, R.J.; van Grondelle, R.; van der Zwan, G. Absorption and CD spectroscopy and modeling of various LH2 complexes from purple bacteria. Biophys. J. 2002, 82, 2184–2197. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.; Newton, M.D.; Silbey, R.J. Multichromophoric forster resonance energy transfer from B800 to B850 in the light harvesting complex 2: Evidence for subtle energetic optimization by purple bacteria. J. Phys. Chem. B 2007, 111, 6807–6814. [Google Scholar] [CrossRef]
- Montemayor, D.; Rivera, E.; Jang, S.J. Computational Modeling of Exciton-Bath Hamiltonians for Light Harvesting 2 and Light Harvesting 3 Complexes of Purple Photosynthetic Bacteria at Room Temperature. J. Phys. Chem. B 2018, 122, 3815–3825. [Google Scholar] [CrossRef] [PubMed]
- Sundstrom, V.; Pullerits, T.; van Grondelle, R. Photosynthetic light-harvesting: Reconciling dynamics and structure of purple bacterial LH2 reveals function of photosynthetic unit. J. Phys. Chem. B 1999, 103, 2327–2346. [Google Scholar] [CrossRef]
- Zerlauskiene, O.; Trinkunas, G.; Gall, A.; Robert, B.; Urboniene, V.; Valkunas, L. Static and Dynamic Protein Impact on Electronic Properties of Light-Harvesting Complex LH2. J. Phys. Chem. B 2008, 112, 15883–15892. [Google Scholar] [CrossRef]
- Novoderezhkin, V.; Wendling, M.; van Grondelle, R. Intra- and interband transfers in the B800-B850 antenna of Rhodospirillum molischianum: Redfield theory modeling of polarized pump-probe kinetics. J. Phys. Chem. B 2003, 107, 11534–11548. [Google Scholar] [CrossRef]
- Novoderezhkin, V.I.; Stuart, T.A.C.; van Grondelle, R. Dynamics of Exciton Relaxation in LH2 Antenna Probed by Multipulse Nonlinear Spectroscopy. J. Phys. Chem. A 2011, 115, 3834–3844. [Google Scholar] [CrossRef]
- Trinkunas, G.; Herek, J.L.; Polivka, T.; Sundstrom, V.; Pullerits, T. Exciton delocalization probed by excitation annihilation in the light-harvesting antenna LH2. Phys. Rev. Lett. 2001, 86, 4167–4170. [Google Scholar] [CrossRef]
- Jang, S.J.; Silbey, R.J. Single complex line shapes of the B850 band of LH2. J. Chem. Phys. 2003, 118, 9324–9336. [Google Scholar] [CrossRef]
- Schlau-Cohen, G.S.; Wang, Q.; Southall, J.; Cogdell, R.J.; Moerner, W.E. Single-molecule spectroscopy reveals photosynthetic LH2 complexes switch between emissive states. Proc. Natl. Acad. Sci. USA 2013, 110, 10899–10903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harel, E.; Engel, G.S. Quantum coherence spectroscopy reveals complex dynamics in bacterial light-harvesting complex 2 (LH2). Proc. Natl. Acad. Sci. USA 2012, 109, 706–711. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.C.; Silbey, R.J. Coherence in the B800 ring of purple bacteria LH2. Phys. Rev. Lett. 2006, 96, 028103. [Google Scholar] [CrossRef]
- Rancova, O.; Abramavicius, D. Static and Dynamic Disorder in Bacterial Light-Harvesting Complex LH2: A 2DES Simulation Study. J. Phys. Chem. B 2014, 118, 7533–7540. [Google Scholar] [CrossRef] [PubMed]
- Segatta, F.; Cupellini, L.; Jurinovich, S.; Mukamel, S.; Dapor, M.; Taioli, S.; Garavelli, M.; Mennucci, B. A Quantum Chemical Interpretation of Two-Dimensional Electronic Spectroscopy of Light-Harvesting Complexes. J. Am. Chem. Soc. 2017, 139, 7558–7567. [Google Scholar] [CrossRef] [PubMed]
- Zigmantas, D.; Read, E.L.; Mancal, T.; Brixner, T.; Gardiner, A.T.; Cogdell, R.J.; Fleming, G.R. Two-dimensional electronic spectroscopy of the B800-B820 light-harvesting complex. Proc. Natl. Acad. Sci. USA 2006, 103, 12672–12677. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, M.; Novoderezhkin, V.I.; Romero, E.; Augulis, R.; Pandit, A.; Zigmantasc, D.; van Grondelle, R. The nature of coherences in the B820 bacteriochlorophyll dimer revealed by two-dimensional electronic spectroscopy. Phys. Chem. Chem. Phys. 2014, 16, 9930–9939. [Google Scholar] [CrossRef]
- Madjet, M.E.; Abdurahman, A.; Renger, T. Intermolecular Coulomb couplings from ab initio electrostatic potentials: Application to optical transitions of strongly coupled pigments in photosynthetic antennae and reaction centers. J. Phys. Chem. B 2006, 110, 17268–17281. [Google Scholar] [CrossRef]
- Novoderezhkin, V.I.; Rutkauskas, D.; van Grondelle, R. Dynamics of the emission spectrum of a single LH2 complex: Interplay of slow and fast nuclear motions. Biophys. J. 2006, 90, 2890–2902. [Google Scholar] [CrossRef] [Green Version]
- Nelson, T.R.; White, A.J.; Bjorgaard, J.A.; Sifain, A.E.; Zhang, Y.; Nebgen, B.; Fernandez-Alberti, S.; Mozyrsky, D.; Roitberg, A.E.; Tretiak, S. Non-adiabatic Excited-State Molecular Dynamics: Theory and Applications for Modeling Photophysics in Extended Molecular Materials. Chem. Rev. 2020, 120, 2215–2287. [Google Scholar] [CrossRef] [PubMed]
- Romero, E.; Novoderezhkin, V.I.; van Grondelle, R. Quantum design of photosynthesis for bio-inspired solar-energy conversion. Nature 2017, 543, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Renger, T. Semiclassical Modified Redfield and Generalized Förster theories of exciton relaxation/transfer in light-harvesting complexes: The quest for the principle of detailed balance. J. Phys. Chem. B 2021, 125, 6406–6416. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pishchalnikov, R.Y.; Chesalin, D.D.; Razjivin, A.P. The Relationship between the Spatial Arrangement of Pigments and Exciton Transition Moments in Photosynthetic Light-Harvesting Complexes. Int. J. Mol. Sci. 2021, 22, 10031. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810031
Pishchalnikov RY, Chesalin DD, Razjivin AP. The Relationship between the Spatial Arrangement of Pigments and Exciton Transition Moments in Photosynthetic Light-Harvesting Complexes. International Journal of Molecular Sciences. 2021; 22(18):10031. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810031
Chicago/Turabian StylePishchalnikov, Roman Y., Denis D. Chesalin, and Andrei P. Razjivin. 2021. "The Relationship between the Spatial Arrangement of Pigments and Exciton Transition Moments in Photosynthetic Light-Harvesting Complexes" International Journal of Molecular Sciences 22, no. 18: 10031. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810031