
Novel Insights into Epigenetic Regulation of IL6 Pathway: In Silico Perspective on Inflammation and Cancer Relationship

 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Gene Expression and Survival Analysis
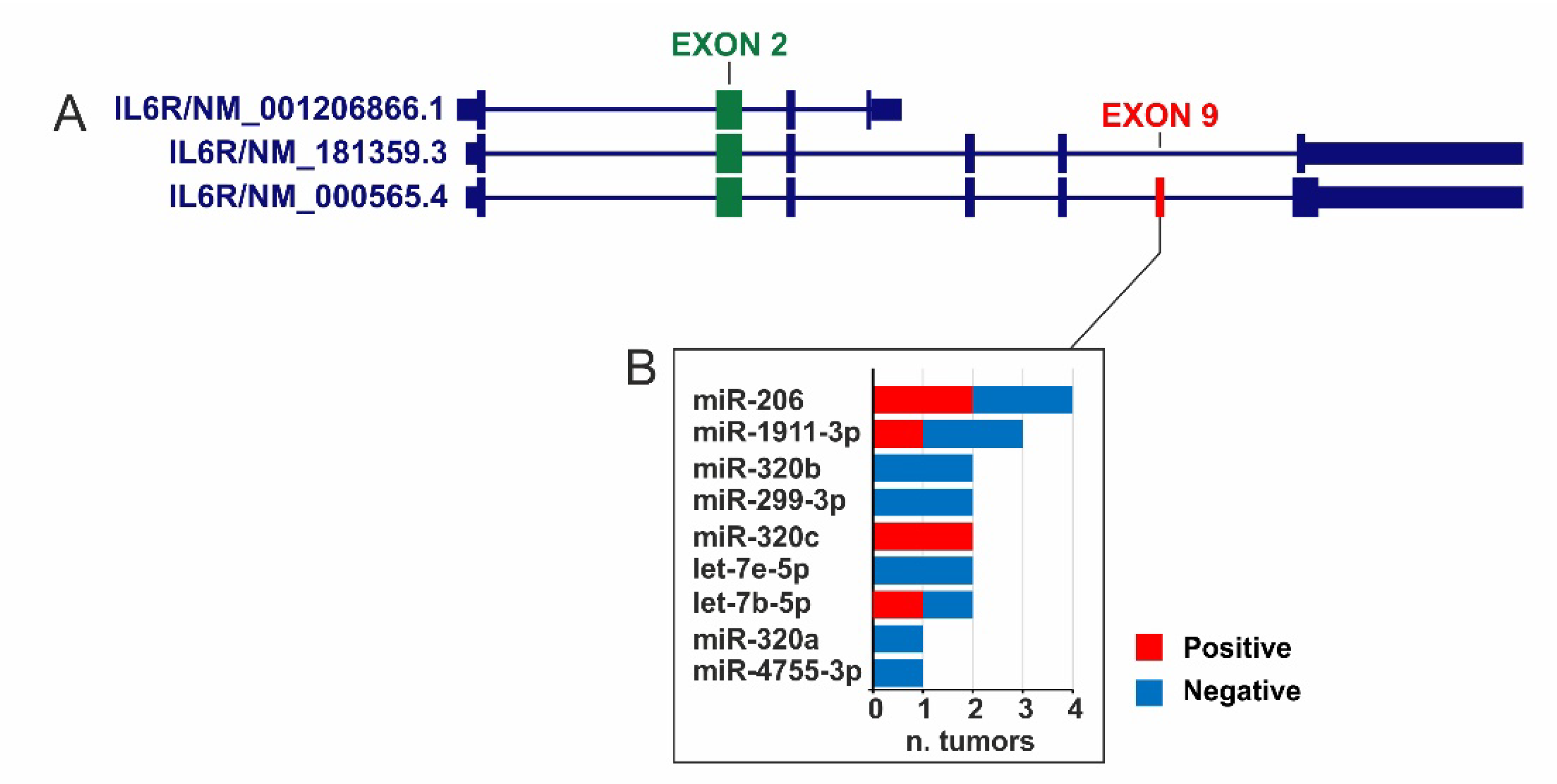
2.2. IL6R Exon Expression and Survival Analysis
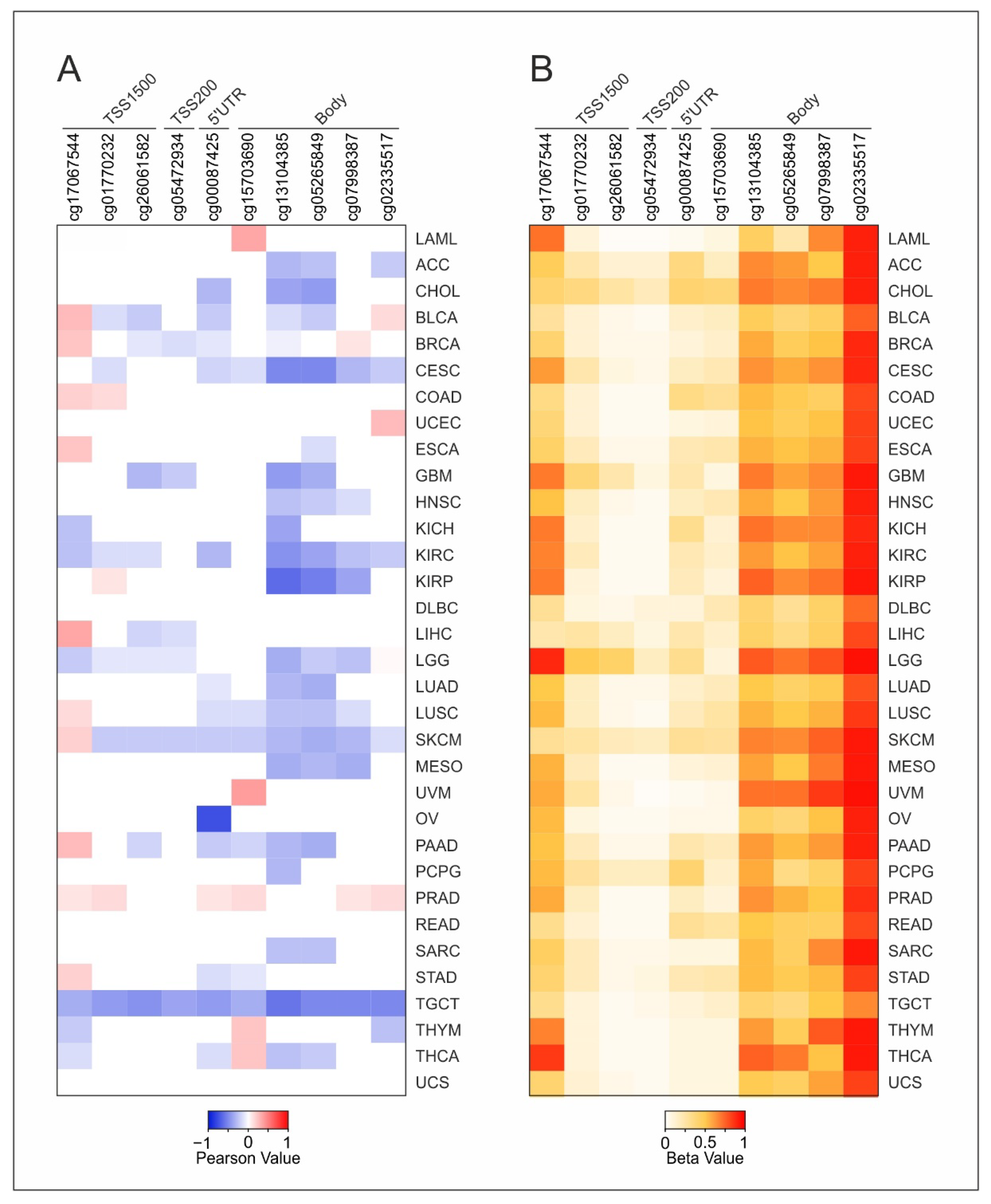
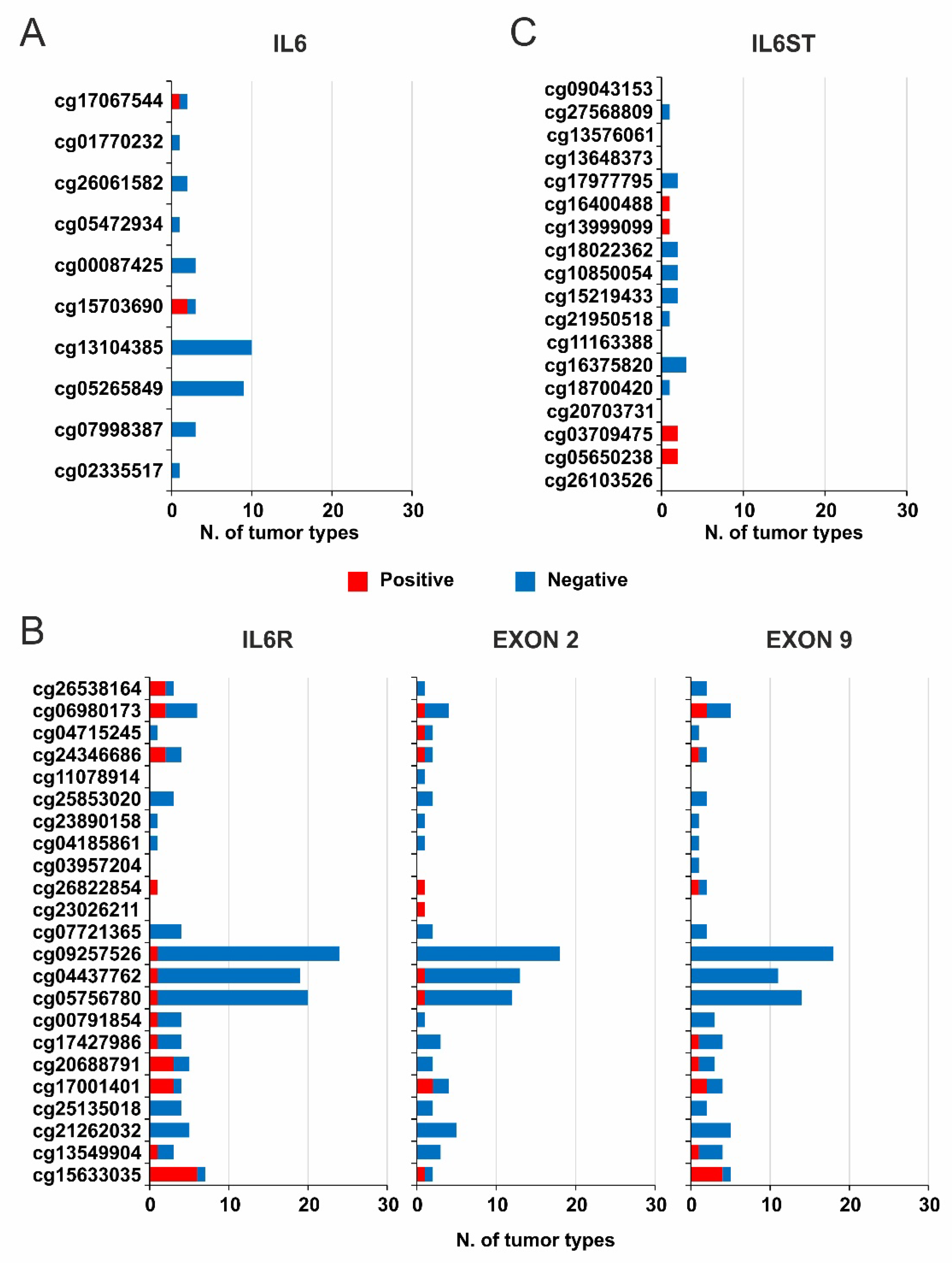
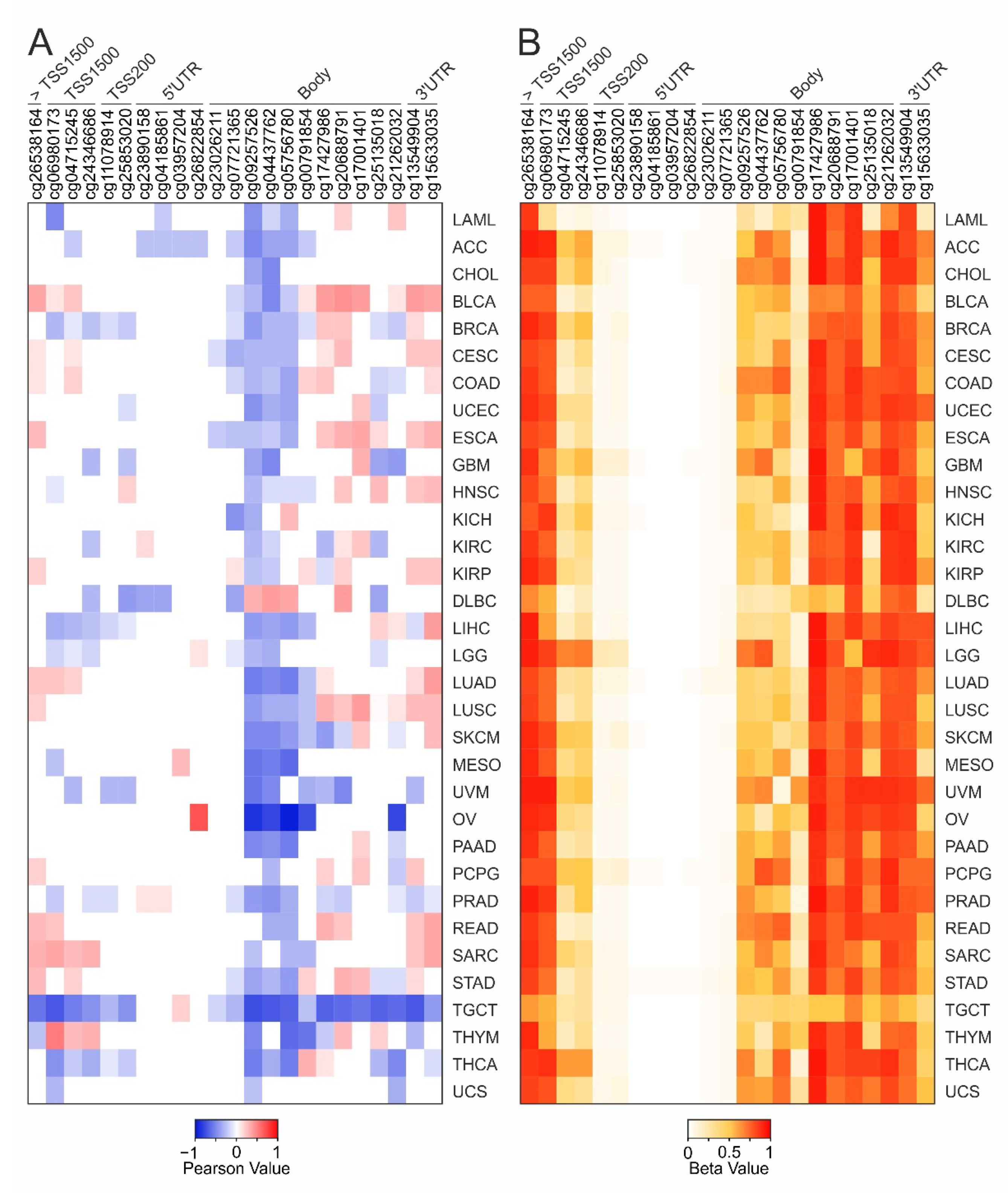
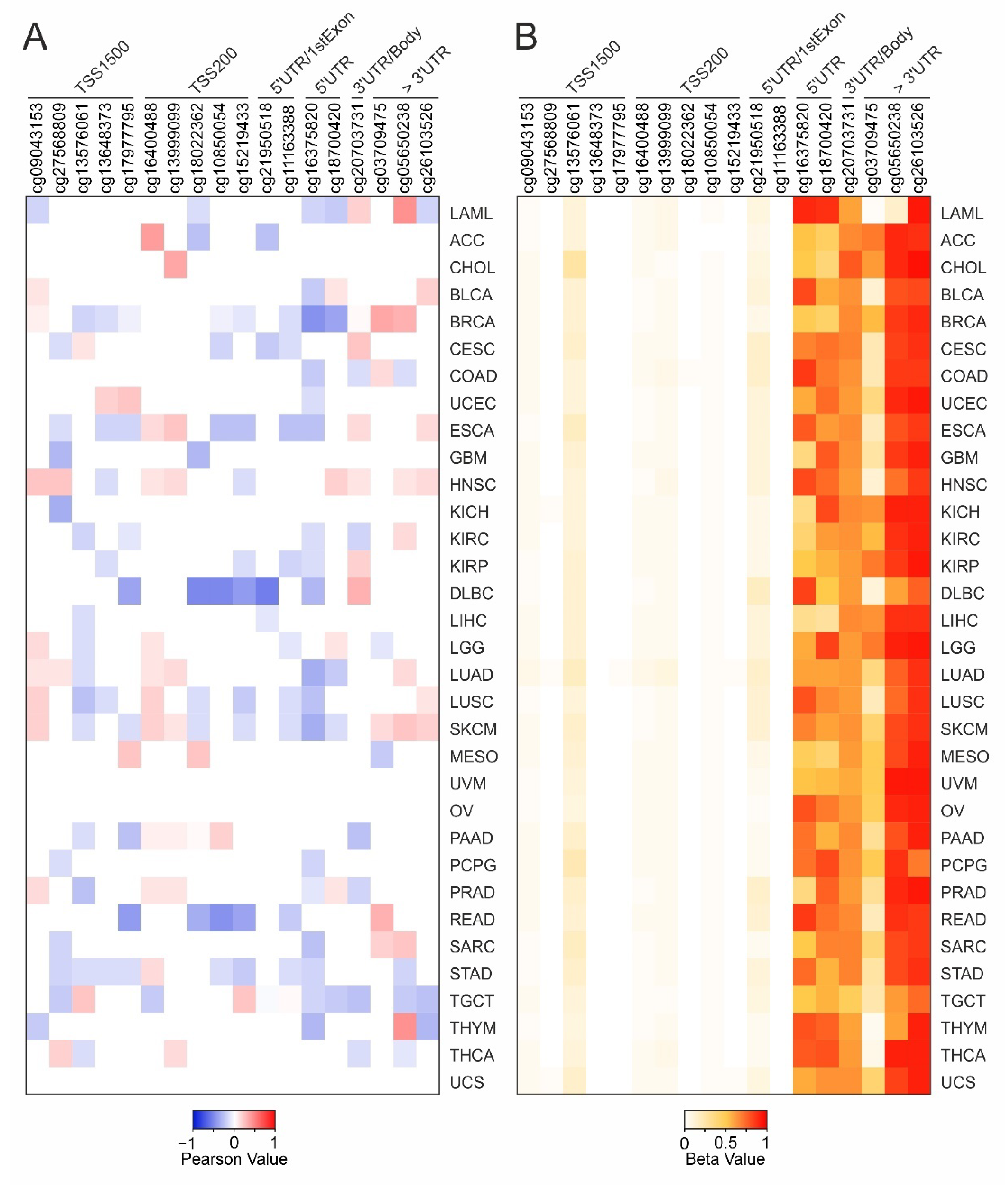
2.3. DNA Methylation and Survival Analysis
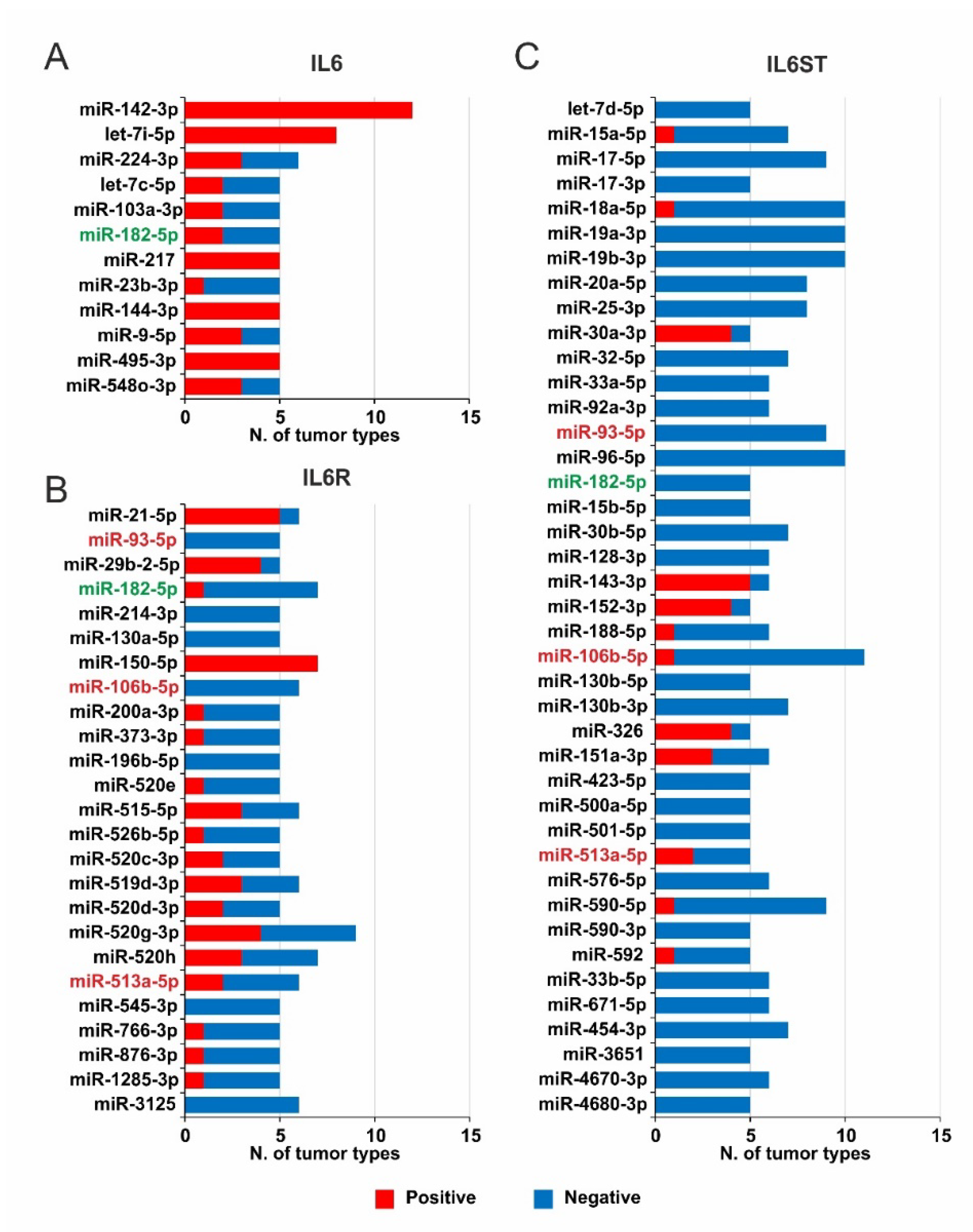
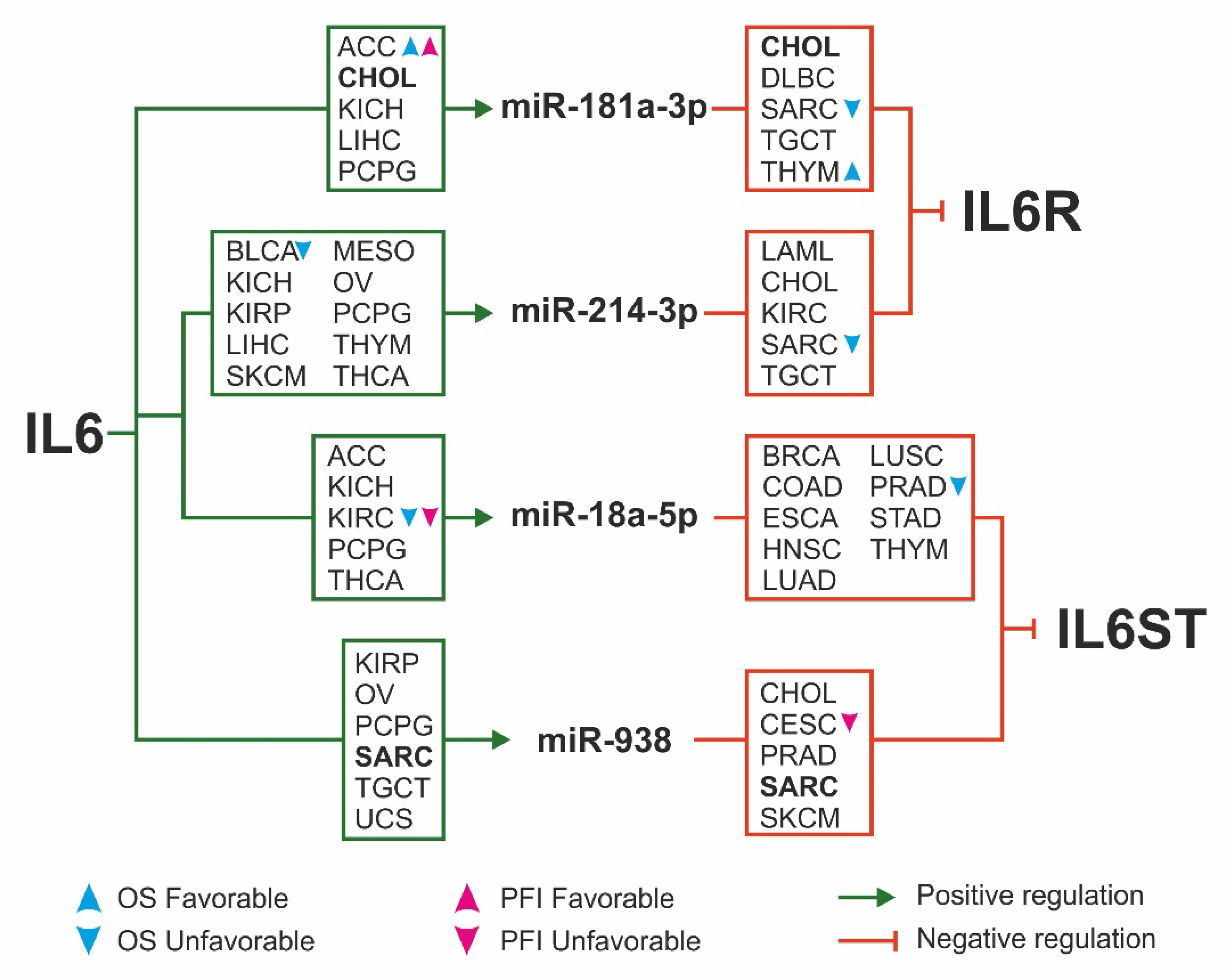
2.4. miRNAs Expression and Survival Analysis
3. Discussion
4. Materials and Methods
4.1. Sample and Datasets
4.2. Analysis of Gene and Exon Expression
4.3. Overall Survival and Progression Free Interval Analyses
4.4. Correlation Analysis
4.5. miRNA Targets Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Setrerrahmane, S.; Xu, H. Tumor-related interleukins: Old validated targets for new anti-cancer drug development. Mol. Cancer 2017, 16, 153. [Google Scholar] [CrossRef] [PubMed]
- Brücher, B.L.; Jamall, I.S. Cell-cell communication in the tumor microenvironment, carcinogenesis, and anticancer treatment. Cell. Physiol. Biochem. 2014, 34, 213–243. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.P.; Li, J.; Tewari, A.K. Inflammation and prostate cancer: The role of interleukin 6 (IL-6). BJU Int. 2014, 113, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Ward, A.C. Role of the interleukin 6 receptor family in epithelial ovarian cancer and its clinical implications. Biochim. Biophys. Acta 2014, 1845, 117–125. [Google Scholar] [CrossRef]
- Kang, S.; Narazaki, M.; Metwally, H.; Kishimoto, T. Historical overview of the interleukin-6 family cytokine. J. Exp. Med. 2020, 217, e20190347. [Google Scholar] [CrossRef] [Green Version]
- Boulanger, M.J.; Chow, D.C.; Brevnova, E.E.; Garcia, K.C. Hexameric structure and assembly of the interleukin-6/IL-6 alpha-receptor/gp130 complex. Science 2003, 300, 2101–2104. [Google Scholar] [CrossRef]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248. [Google Scholar] [CrossRef]
- Comen, E.A.; Bowman, R.L.; Kleppe, M. Underlying Causes and Therapeutic Targeting of the Inflammatory Tumor Microenvironment. Front. Cell Dev. Biol. 2018, 6, 56. [Google Scholar] [CrossRef]
- Bournazou, E.; Bromberg, J. Targeting the tumor microenvironment: JAK-STAT3 signaling. JAKSTAT 2013, 2, e23828. [Google Scholar] [CrossRef] [Green Version]
- George, D.J.; Halabi, S.; Shepard, T.F.; Sanford, B.; Vogelzang, N.J.; Small, E.J.; Kantoff, P.W. The prognostic significance of plasma interleukin-6 levels in patients with metastatic hormone-refractory prostate cancer: Results from cancer and leukemia group B 9480. Clin. Cancer Res. 2005, 11, 1815–1820. [Google Scholar] [CrossRef] [Green Version]
- Lederle, W.; Depner, S.; Schnur, S.; Obermueller, E.; Catone, N.; Just, A.; Fusenig, N.E.; Mueller, M.M. IL-6 promotes malignant growth of skin SCCs by regulating a network of autocrine and paracrine cytokines. Int. J. Cancer 2011, 128, 2803–2814. [Google Scholar] [CrossRef]
- Kusaba, T.; Nakayama, T.; Yamazumi, K.; Yakata, Y.; Yoshizaki, A.; Inoue, K.; Nagayasu, T.; Sekine, I. Activation of STAT3 is a marker of poor prognosis in human colorectal cancer. Oncol. Rep. 2006, 15, 1445–1451. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, J.; Wang, X.; Liu, X.; Li, H.; Lv, Q.; Zhu, J.; Wei, B.; Tang, Y. STAT3, a Poor Survival Predicator, Is Associated with Lymph Node Metastasis from Breast Cancer. J. Breast Cancer 2013, 16, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Macha, M.A.; Matta, A.; Kaur, J.; Chauhan, S.S.; Thakar, A.; Shukla, N.K.; Gupta, S.D.; Ralhan, R. Prognostic significance of nuclear pSTAT3 in oral cancer. Head Neck 2011, 33, 482–489. [Google Scholar] [CrossRef]
- McFarland, B.C.; Hong, S.W.; Rajbhandari, R.; Twitty, G.B., Jr.; Gray, G.K.; Yu, H.; Benveniste, E.N.; Nozell, S.E. NF-κB-induced IL-6 ensures STAT3 activation and tumor aggressiveness in glioblastoma. PLoS ONE 2013, 8, e78728. [Google Scholar]
- Qian, D.; Yan, S.; Pan, X. Association of IL-6 -597 G/A Polymorphism with Cancer Risk: Evidence from a Meta-Analysis. Crit. Rev. Eukaryot. Gene Expr. 2017, 27, 211–217. [Google Scholar] [CrossRef]
- Malaponte, G.; Polesel, J.; Candido, S.; Sambataro, D.; Bevelacqua, V.; Anzaldi, M.; Vella, N.; Fiore, V.; Militello, L.; Mazzarino, M.C.; et al. IL-6-174 G > C and MMP-9-1562 C > T polymorphisms are associated with increased risk of deep vein thrombosis in cancer patients. Cytokine 2013, 62, 64–69. [Google Scholar] [CrossRef]
- Baran, P.; Nitz, R.; Grötzinger, J.; Scheller, J.; Garbers, C. Minimal interleukin 6 (IL-6) receptor stalk composition for IL-6 receptor shedding and IL-6 classic signaling. J. Biol. Chem. 2013, 288, 14756–14768. [Google Scholar] [CrossRef] [Green Version]
- Jones, S.A.; Horiuchi, S.; Topley, N.; Yamamoto, N.; Fuller, G.M. The soluble interleukin 6 receptor: Mechanisms of production and implications in disease. FASEB J. 2001, 15, 43–58. [Google Scholar] [CrossRef] [Green Version]
- Candido, S.; Parasiliti Palumbo, G.A.; Pennisi, M.; Russo, G.; Sgroi, G.; Di Salvatore, V.; Libra, M.; Pappalardo, F. EpiMethEx: A tool for large-scale integrated analysis in methylation hotspots linked to genetic regulation. BMC Bioinform. 2019, 19, 385. [Google Scholar]
- You, J.S.; Jones, P.A. Cancer genetics and epigenetics: Two sides of the same coin? Cancer Cell 2012, 22, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasmin, R.; Siraj, S.; Hassan, A.; Khan, A.R.; Abbasi, R.; Ahmad, N. Epigenetic regulation of inflammatory cytokines and associated genes in human malignancies. Mediators Inflamm. 2015, 2015, 201703. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Pinello, N.; Nguyen, T.V.; Thoeng, A.; Nagarajah, R.; Holst, J.; Rasko, J.E.; Wong, J.J. DNA methylation/hydroxymethylation regulate gene expression and alternative splicing during terminal granulopoiesis. Epigenomics 2019, 11, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Maunakea, A.K.; Chepelev, I.; Cui, K.; Zhao, K. Intragenic DNA methylation modulates alternative splicing by recruiting MeCP2 to promote exon recognition. Cell Res. 2013, 23, 1256–1269. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Tian, Y.; Wang, J.; Sun, Z.; Zhu, Y. Genome-wide analysis reveals the association between alternative splicing and DNA methylation across human solid tumors. BMC Med. Genomics 2020, 13, 4. [Google Scholar] [CrossRef]
- Shayevitch, R.; Askayo, D.; Keydar, I.; Ast, G. The importance of DNA methylation of exons on alternative splicing. RNA 2018, 24, 1351–1362. [Google Scholar] [CrossRef] [Green Version]
- Tekpli, X.; Landvik, N.E.; Anmarkud, K.H.; Skaug, V.; Haugen, A.; Zienolddiny, S. DNA methylation at promoter regions of interleukin 1B, interleukin 6, and interleukin 8 in non-small cell lung cancer. Cancer Immunol. Immunother. 2013, 62, 337–345. [Google Scholar] [CrossRef]
- D’Anello, L.; Sansone, P.; Storci, G.; Mitrugno, V.; D’Uva, G.; Chieco, P.; Bonafé, M. Epigenetic control of the basal-like gene expression profile via Interleukin-6 in breast cancer cells. Mol. Cancer 2010, 9, 300. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.C.; Lin, J.H.; Hsu, T.W.; Su, K.; Li, A.F.; Hsu, H.S.; Hung, S.C. IL-6 enriched lung cancer stem-like cell population by inhibition of cell cycle regulators via DNMT1 upregulation. Int. J. Cancer 2015, 136, 547–559. [Google Scholar]
- Tang, L.P.; Cho, C.H.; Hui, W.M.; Huang, C.; Chu, K.M.; Xia, H.H.; Lam, S.K.; Rashid, A.; Wong, B.C.; Chan, A.O. An inverse correlation between Interleukin-6 and select gene promoter methylation in patients with gastric cancer. Digestion 2006, 74, 85–90. [Google Scholar] [CrossRef]
- Yao, Q.; Chen, Y.; Zhou, X. The roles of microRNAs in epigenetic regulation. Curr. Opin. Chem. Biol. 2019, 51, 11–17. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, X.; Cai, Z.; Zhou, J.; Cao, R.; Zhao, Y.; Chen, Z.; Wang, D.; Ruan, W.; Zhao, Q.; et al. A novel class of microRNA-recognition elements that function only within open reading frames. Nat. Struct. Mol. Biol. 2018, 25, 1019–1027. [Google Scholar] [CrossRef]
- Jansson, M.D.; Lund, A.H. MicroRNA and cancer. Mol. Oncol. 2012, 6, 590–610. [Google Scholar] [CrossRef]
- Shenouda, S.K.; Alahari, S.K. MicroRNA function in cancer: Oncogene or a tumor suppressor? Cancer Metastasis Rev. 2009, 28, 369–378. [Google Scholar] [CrossRef]
- Leonardi, G.C.; Candido, S.; Carbone, M.; Colaianni, V.; Garozzo, S.F.; Cinà, D.; Libra, M. microRNAs and thyroid cancer: Biological and clinical significance. Int. J. Mol. Med. 2012, 30, 991–999. [Google Scholar] [CrossRef] [Green Version]
- Candido, S.; Lupo, G.; Pennisi, M.; Basile, M.S.; Anfuso, C.D.; Petralia, M.C.; Gattuso, G.; Vivarelli, S.; Spandidos, D.A.; Libra, M.; et al. The analysis of miRNA expression profiling datasets reveals inverse microRNA patterns in glioblastoma and Alzheimer’s disease. Oncol. Rep. 2019, 42, 911–922. [Google Scholar] [CrossRef] [Green Version]
- Tan, W.; Liu, B.; Qu, S.; Liang, G.; Luo, W.; Gong, C. MicroRNAs and cancer: Key paradigms in molecular therapy. Oncol. Lett. 2018, 15, 2735–2742. [Google Scholar] [CrossRef] [Green Version]
- Christopher, A.F.; Gupta, M.; Bansal, P. Micronome revealed miR-19a/b as key regulator of SOCS3 during cancer related inflammation of oral squamous cell carcinoma. Gene 2016, 594, 30–40. [Google Scholar] [CrossRef]
- Srivastava, S.; Tsongalis, G.J.; Kaur, P. Role of microRNAs in regulation of the TNF/TNFR gene superfamily in chronic lymphocytic leukemia. Clin. Biochem. 2016, 49, 1307–1310. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, L.; Cui, M.; Ye, W.; Zhang, P.; Zhou, S.; Wang, J. miR-302b inhibits cancer-related inflammation by targeting ERBB4, IRF2 and CXCR4 in esophageal cancer. Oncotarget 2017, 8, 49053–49063. [Google Scholar] [CrossRef] [Green Version]
- Lin, K.Y.; Ye, H.; Han, B.W.; Wang, W.T.; Wei, P.P.; He, B.; Li, X.J.; Chen, Y.Q. Genome-wide screen identified let-7c/miR-99a/miR-125b regulating tumor progression and stem-like properties in cholangiocarcinoma. Oncogene 2016, 35, 3376–3386. [Google Scholar] [CrossRef] [Green Version]
- Yokomizo, R.; Yanaihara, N.; Yamaguchi, N.; Saito, M.; Kawabata, A.; Takahashi, K.; Takenaka, M.; Yamada, K.; Shapiro, J.S.; Okamoto, A. MicroRNA-34a/IL-6R pathway as a potential therapeutic target for ovarian high-grade serous carcinoma. Oncotarget 2019, 10, 4880–4893. [Google Scholar] [CrossRef]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Grivennikov, S.I.; Karin, M. Inflammation and oncogenesis: A vicious connection. Curr. Opin. Genet. Dev. 2010, 20, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Burger, R. Impact of interleukin-6 in hematological malignancies. Transfus. Med. Hemother. 2013, 40, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Van Duijneveldt, G.; Griffin, M.D.W.; Putoczki, T.L. Emerging roles for the IL-6 family of cytokines in pancreatic cancer. Clin. Sci. 2020, 134, 2091–2115. [Google Scholar] [CrossRef]
- Silva, E.M.; Mariano, V.S.; Pastrez, P.R.A.; Pinto, M.C.; Castro, A.G.; Syrjanen, K.J.; Longatto-Filho, A. High systemic IL-6 is associated with worse prognosis in patients with non-small cell lung cancer. PLoS ONE 2017, 12, e0181125. [Google Scholar] [CrossRef]
- Shang, G.S.; Liu, L.; Qin, Y.W. IL-6 and TNF-α promote metastasis of lung cancer by inducing epithelial-mesenchymal transition. Oncol. Lett. 2017, 13, 4657–4660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browning, L.; Patel, M.R.; Horvath, E.B.; Tawara, K.; Jorcyk, C.L. IL-6 and ovarian cancer: Inflammatory cytokines in promotion of metastasis. Cancer Manag. Res. 2018, 10, 6685–6693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, B.; Haggeney, T.; Fietz, D.; Indumathy, S.; Loveland, K.L.; Hedger, M.; Kliesch, S.; Weidner, W.; Bergmann, M.; Schuppe, H.C. Specific immune cell and cytokine characteristics of human testicular germ cell neoplasia. Hum. Reprod. 2016, 31, 2192–2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, C.; Shen, J.; Fang, Z.; Qiao, L.; Feng, R.; Lin, X.; Li, S. Abnormally expressed JunB transactivated by IL-6/STAT3 signaling promotes uveal melanoma aggressiveness via epithelial-mesenchymal transition. Biosci. Rep. 2018, 38, BSR20180532. [Google Scholar] [CrossRef] [Green Version]
- Nagarkatti-Gude, N.; Bronkhorst, I.H.; van Duinen, S.G.; Luyten, G.P.; Jager, M.J. Cytokines and chemokines in the vitreous fluid of eyes with uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2012, 53, 6748–6755. [Google Scholar] [CrossRef] [Green Version]
- Stabile, L.P.; Egloff, A.M.; Gibson, M.K.; Gooding, W.E.; Ohr, J.; Zhou, P.; Rothenberger, N.J.; Wang, L.; Geiger, J.L.; Flaherty, J.T.; et al. IL6 is associated with response to dasatinib and cetuximab: Phase II clinical trial with mechanistic correlatives in cetuximab-resistant head and neck cancer. Oral. Oncol. 2017, 69, 38–45. [Google Scholar] [CrossRef]
- Rong, Q.; Gao, Y.; Cai, Q.; Wang, X.; Bai, B.; Ping, L.; He, H.; Rao, H.; Zhang, Y.; Li, Z.; et al. High IL-6 expression in the tumor microenvironment is associated with poor prognosis of patients with extranodal natural/killer T-cell lymphoma (ENKTL). Expert Rev. Anticancer Ther. 2021, 21, 121–127. [Google Scholar] [CrossRef]
- Gholiha, A.R.; Hollander, P.; Glimelius, I.; Hedstrom, G.; Molin, D.; Hjalgrim, H.; Smedby, K.E.; Hashemi, J.; Amini, R.M.; Enblad, G. Revisiting IL-6 expression in the tumor microenvironment of classical Hodgkin lymphoma. Blood Adv. 2021, 5, 1671–1681. [Google Scholar] [CrossRef]
- Cui, G.; Yuan, A.; Sun, Z.; Zheng, W.; Pang, Z. IL-1β/IL-6 network in the tumor microenvironment of human colorectal cancer. Pathol. Res. Pract. 2018, 214, 986–992. [Google Scholar] [CrossRef]
- Fisher, D.T.; Appenheimer, M.M.; Evans, S.S. The two faces of IL-6 in the tumor microenvironment. Semin. Immunol. 2014, 26, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Cao, M.Q.; You, A.B.; Zhu, X.D.; Zhang, W.; Zhang, Y.Y.; Zhang, S.Z.; Zhang, K.W.; Cai, H.; Shi, W.K.; Li, X.L.; et al. miR-182-5p promotes hepatocellular carcinoma progression by repressing FOXO3a. J. Hematol. Oncol. 2018, 11, 12. [Google Scholar] [CrossRef] [Green Version]
- Sang, Y.; Chen, B.; Song, X.; Li, Y.; Liang, Y.; Han, D.; Zhang, N.; Zhang, H.; Liu, Y.; Chen, T.; et al. circRNA_0025202 Regulates Tamoxifen Sensitivity and Tumor Progression via Regulating the miR-182-5p/FOXO3a Axis in Breast Cancer. Mol. Ther. 2019, 27, 1638–1652. [Google Scholar] [CrossRef]
- Jia, X.N.; Yin, S.D.; Wei, Y.; Chen, L. MiR-182-5p inhibited proliferation and migration of ovarian cancer cells by targeting BNIP3. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 7912. [Google Scholar]
- Xie, F.; Li, Y.; Wang, M.; Huang, C.; Tao, D.; Zheng, F.; Zhang, H.; Zeng, F.; Xiao, X.; Jiang, G. Circular RNA BCRC-3 suppresses bladder cancer proliferation through miR-182-5p/p27 axis. Mol. Cancer. 2018, 17, 144. [Google Scholar] [CrossRef]
- Yan, S.; Wang, H.; Chen, X.; Liang, C.; Shang, W.; Wang, L.; Li, J.; Xu, D. MiR-182-5p inhibits colon cancer tumorigenesis, angiogenesis, and lymphangiogenesis by directly downregulating VEGF-C. Cancer Lett. 2020, 488, 18–26. [Google Scholar] [CrossRef]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repečka, K.; McDade, F.; Kamath, A.; Banerjee, A.; Luo, Y.; Rogers, D.; Brooks, A.N.; et al. Visualizing and interpreting cancer genomics data via the Xena platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef]
- Ryan, M.; Wong, W.C.; Brown, R.; Akbani, R.; Su, X.; Broom, B.; Melott, J.; Weinstein, J. TCGASpliceSeq a compendium of alternative mRNA splicing in cancer. Nucleic Acids Res. 2016, 44, D1018–D1022. [Google Scholar] [CrossRef]
- Bland, J.M.; Altman, D.G. The logrank test. BMJ 2004, 328, 1073. [Google Scholar] [CrossRef] [Green Version]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Tokar, T.; Pastrello, C.; Rossos, A.E.M.; Abovsky, M.; Hauschild, A.C.; Tsay, M.; Lu, R.; Jurisica, I. mirDIP 4.1-integrative database of human microRNA target predictions. Nucleic Acids Res. 2018, 46, D360–D370. [Google Scholar] [CrossRef]
- Rennie, W.; Kanoria, S.; Liu, C.; Mallick, B.; Long, D.; Wolenc, A.; Carmack, C.S.; Lu, J.; Ding, Y. STarMirDB: A database of microRNA binding sites. RNA Biol. 2016, 13, 554–560. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor Type | IL6 | IL6R | IL6ST | ||||||
|---|---|---|---|---|---|---|---|---|---|
| T vs. N (FC) | OS * | PFI * | T vs. N (FC) | OS * | PFI * | T vs. N (FC) | OS * | PFI * | |
| Acute Myeloid Leukemia (LAML) | −2.92 | NA | −1.93 | NA | NA | ||||
| Adrenocortical Cancer (ACC) | −1.44 | ||||||||
| Bile Duct Cancer (CHOL) | 5.584 (+) | ||||||||
| Bladder Cancer (BLCA) | 1.89 | −3.51 | −3.30 | 5.39 (−) | |||||
| Breast Cancer (BRCA) | −7.37 | −2.02 | 7.54 (+) | ||||||
| Cervical Cancer (CESC) | 3.04 | −2.61 | −3.32 | ||||||
| Colon Cancer (COAD) | −5.70 | −2.56 | |||||||
| Endometrioid Cancer (UCEC) | 1.79 | −3.30 | −3.21 | ||||||
| Esophageal Cancer (ESCA) | 1.62 | ||||||||
| Glioblastoma (GBM) | 2.66 | 5.89 (−) | 1.40 | ||||||
| Head and Neck Cancer (HNSC) | 2.20 | 4.57 (−) | 1.79 | −2.36 | 5.42 (+) | 5.23 (+) | |||
| Kidney Chromophobe (KICH) | −2.44 | −2.21 | |||||||
| Kidney Clear Cell Carcinoma (KIRC) | 1.59 | 31.46 (−) | 17.13 (−) | 2.00 | 16.31 (+) | 16.09 (+) | 3.11 | 9.10 (+) | 6.09 (+) |
| Kidney Papillary Cell Carcinoma (KIRP) | 5.78 (−) | 4.43 (−) | |||||||
| Large B-cell Lymphoma (DLBC) | 6.39 | −10.49 | |||||||
| Liver Cancer (LIHC) | −1.58 | −1.46 | |||||||
| Lower Grade Glioma (LGG) | −1.92 | 12.38 (−) | 6.77 (−) | 5.04 (−) | 1.41 | ||||
| Lung Adenocarcinoma (LUAD) | −4.08 | 6.50 (+) | −1.66 | 8.90 (+) | |||||
| Lung Squamous Cell Carcinoma (LUSC) | −2.40 | 4.87 (−) | −2.36 | −3.60 | |||||
| Melanoma (SKCM) | 1.42 | ||||||||
| Mesothelioma (MESO) | −2.80 | −3.50 | 4.20 (+) | 11.79 (+) | |||||
| Ocular melanomas (UVM) | −11.90 | 7.52 (−) | 4.19 (−) | −1.46 | |||||
| Ovarian Cancer (OV) | 2.88 | −2.54 | |||||||
| Pancreatic Cancer (PAAD) | 3.21 | 1.42 | |||||||
| Pheochromocytoma and Paraganglioma (PCPG) | −3.97 | −1.74 | |||||||
| Prostate Cancer (PRAD) | −1.87 | ||||||||
| Rectal Cancer (READ) | −5.66 | −2.43 | |||||||
| Sarcoma (SARC) | 5.74 (+) | −15.19 | 6.92 (+) | ||||||
| Stomach Cancer (STAD) | 2.18 | 4.57 (−) | −1.56 | 4.59 (−) | 4.16 (−) | ||||
| Testicular Cancer (TGCT) | 5.19 | 3.47 | |||||||
| Thymoma (THYM) | −11.77 | 3.87 (−) | 2.69 | ||||||
| Thyroid Cancer (THCA) | −2.20 | 3.66 (−) | −1.59 | ||||||
| Uterine Carcinosarcoma (UCS) | 1.52 | −5.38 | −2.89 | ||||||
| Cancer Type | IL6 | IL6R | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| cg13104385 | cg05265849 | cg09257526 | cg04437762 | cg05756780 | ||||||
| OS * | PFI * | OS * | PFI * | OS * | PFI * | OS * | PFI * | OS * | PFI * | |
| Acute Myeloid Leukemia (LAML) | 5.42 (+) | |||||||||
| Adrenocortical Cancer (ACC) | 21.70 (−) | 18.51 (−) | 8.68 (−) | 18.77 (−) | 4.12 (−) | |||||
| Bile Duct Cancer (CHOL) | 6.66 (−) | |||||||||
| Bladder Cancer (BLCA) | 4.72 (−) | |||||||||
| Breast Cancer (BRCA) | 5.31 (+) | |||||||||
| Cervical Cancer (CESC) | 4.25 (−) | 5.53 (−) | 5.28 (−) | 5.90 (−) | ||||||
| Endometrioid Cancer (UCEC) | 7.10 (+) | 5.39 (−) | 4.90 (−) | |||||||
| Glioblastoma (GBM) | 4.58 (+) | |||||||||
| Head and Neck Cancer (HNSC) | 4.60 (−) | |||||||||
| Kidney Clear Cell Carcinoma (KIRC) | 4.93 (−) | 4.09 (−) | ||||||||
| Kidney Papillary Cell Carcinoma (KIRP) | 4.34 (+) | 5.44 (+) | 10.41 (−) | 6.56 (−) | 7.25 (−) | |||||
| Lower Grade Glioma (LGG) | 21.47 (+) | 17.17 (+) | 8.01 (+) | 11.80 (+) | 22.81 (+) | 19.12 (+) | 5.53 (+) | |||
| Lung Adenocarcinoma (LUAD) | 5.18 (−) | 6.92 (−) | 4.03 (−) | 13.40 (−) | 7.39 (−) | |||||
| Lung Squamous Cell Carcinoma (LUSC) | 4.37 (+) | 11.68 (+) | 4.17 (+) | |||||||
| Melanoma (SKCM) | 5.75 (−) | 5.58 (+) | 10.46 (+) | 4.74 (+) | ||||||
| Mesothelioma (MESO) | 5.00 (−) | 9.43 (−) | 14.36 (−) | 27.43 (−) | 9.02 (−) | |||||
| Ocular melanomas (UVM) | 4.28 (−) | 7.39 (−) | 18.55 (−) | 14.88 (−) | ||||||
| Pancreatic Cancer (PAAD) | 4.29 (−) | |||||||||
| Sarcoma (SARC) | 4.15 (+) | 5.06 (−) | ||||||||
| Thymoma (THYM) | 12.19 (+) | |||||||||
| Tumor Type (Abbreviation) | TCGA/GTEx | TCGA † | |||||
|---|---|---|---|---|---|---|---|
| Tumor TCGA (N) | Normal GTEx (N) | Total (N) | Gene expr. (N) | DNA Methylation (N) | miRNA expr. (N) | EXON expr. (N) | |
| Acute Myeloid Leukemia (LAML) | 173 | Blood (337) | 200 | 173 | 194 | 188* | 173 |
| Adrenocortical Cancer (ACC) | 77 | Adrenal Gland (128) | 92 | 79 | 80 | 79 | 79 |
| Bile Duct Cancer (CHOL) | 36 | Liver (110) | 45 | 45 | 45 | 45 | 45 |
| Bladder Cancer (BLCA) | 407 | Bladder (9) | 436 | 426 | 434 | 429 | 426 |
| Breast Cancer (BRCA) | 1099 | Breast (179) | 1247 | 1218 | 888 | 832 | 1218 |
| Cervical Cancer (CESC) | 306 | Cervix Uteri (10) | 313 | 308 | 312 | 311 | 308 |
| Colon Cancer (COAD) | 290 | Colon (308) | 551 | 329 | 337 | 261 | 329 |
| Endometrioid Cancer (UCEC) | 181 | Uterus (78) | 596 | 201 | 478 | 430 | 201 |
| Esophageal Cancer (ESCA) | 182 | Esophagus (653) | 204 | 196 | 202 | 195 | 196 |
| Glioblastoma (GBM) | 166 | Brain (1141) | 629 | 172 | 155 | 5 | 172 |
| Head and Neck Cancer (HNSC) | 520 | Salivary Gland (55) | 604 | 566 | 580 | 529 | 566 |
| Kidney Chromophobe (KICH) | 197 | Kidney (29) | 91 | 91 | 66 | 89 | 91 |
| Kidney Clear Cell Carcinoma (KIRC) | 531 | Kidney (29) | 945 | 606 | 480 | 311 | 606 |
| Kidney Papillary Cell Carcinoma (KIRP) | 289 | Kidney (29) | 352 | 323 | 321 | 321 | 323 |
| Large B-cell Lymphoma (DLBC) | 47 | Blood (337) | 48 | 48 | 48 | 47 | 48 |
| Liver Cancer (LIHC) | 371 | Liver (110) | 438 | 423 | 429 | 420 | 423 |
| Lower Grade Glioma (LGG) | 523 | Brain (1141) | 530 | 530 | 530 | 524 | 530 |
| Lung Adenocarcinoma (LUAD) | 515 | Lung (288) | 706 | 576 | 492 | 495 | 576 |
| Lung Squamous Cell Carcinoma (LUSC) | 498 | Lung (288) | 626 | 553 | 415 | 380 | 553 |
| Melanoma (SKCM) | 469 | Skin (556) | 481 | 474 | 476 | 452 | 474 |
| Mesothelioma (MESO) | 87 | Lung (288) | 87 | 87 | 87 | 87 | 87 |
| Ocular melanomas (UVM) | 79 | Skin (556) | 80 | 80 | 80 | 80 | 80 |
| Ovarian Cancer (OV) | 427 | Ovary (88) | 630 | 308 | 10 | 485 | 308 |
| Pancreatic Cancer (PAAD) | 179 | Pancreas (167) | 196 | 183 | 195 | 182 | 183 |
| Pheochromocytoma & Paraganglioma (PCPG) | 182 | Adrenal Gland (128) | 187 | 187 | 187 | 186 | 187 |
| Prostate Cancer (PRAD) | 496 | Prostate (100) | 566 | 550 | 549 | 544 | 550 |
| Rectal Cancer (READ) | 93 | Colon (308) | 186 | 105 | 106 | 92 | 105 |
| Sarcoma (SARC) | 262 | Muscle (396) | 271 | 265 | 269 | 260 | 265 |
| Stomach Cancer (STAD) | 414 | Stomach (174) | 580 | 450 | 398 | 428 | 450 |
| Testicular Cancer (TGCT) | 154 | Testis (165) | 156 | 156 | 156 | 155 | 156 |
| Thymoma (THYM) | 119 | Blood (337) | 126 | 122 | 126 | 126 | 122 |
| Thyroid Cancer (THCA) | 512 | Thyroid (279) | 580 | 572 | 571 | 569 | 572 |
| Uterine Carcinosarcoma (UCS) | 57 | Uterus (78) | 57 | 57 | 57 | 56 | 57 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Candido, S.; Tomasello, B.M.R.; Lavoro, A.; Falzone, L.; Gattuso, G.; Libra, M. Novel Insights into Epigenetic Regulation of IL6 Pathway: In Silico Perspective on Inflammation and Cancer Relationship. Int. J. Mol. Sci. 2021, 22, 10172. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810172
Candido S, Tomasello BMR, Lavoro A, Falzone L, Gattuso G, Libra M. Novel Insights into Epigenetic Regulation of IL6 Pathway: In Silico Perspective on Inflammation and Cancer Relationship. International Journal of Molecular Sciences. 2021; 22(18):10172. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810172
Chicago/Turabian StyleCandido, Saverio, Barbara Maria Rita Tomasello, Alessandro Lavoro, Luca Falzone, Giuseppe Gattuso, and Massimo Libra. 2021. "Novel Insights into Epigenetic Regulation of IL6 Pathway: In Silico Perspective on Inflammation and Cancer Relationship" International Journal of Molecular Sciences 22, no. 18: 10172. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810172