
Effects of Prenatal Phthalate Exposure and Childhood Exercise on Maternal Behaviors in Female Rats at Postpartum: A Role of Oxtr Methylation in the Hypothalamus

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Effects of Prenatal DEHP Exposure and Childhood Exercise on Anxiety-like Behaviors in Postpartum Dams
2.2. Effects of Prenatal DEHP Exposure and Childhood Exercise on Levels of BDNF, ACTH, and Corticosterone in Postpartum Dams
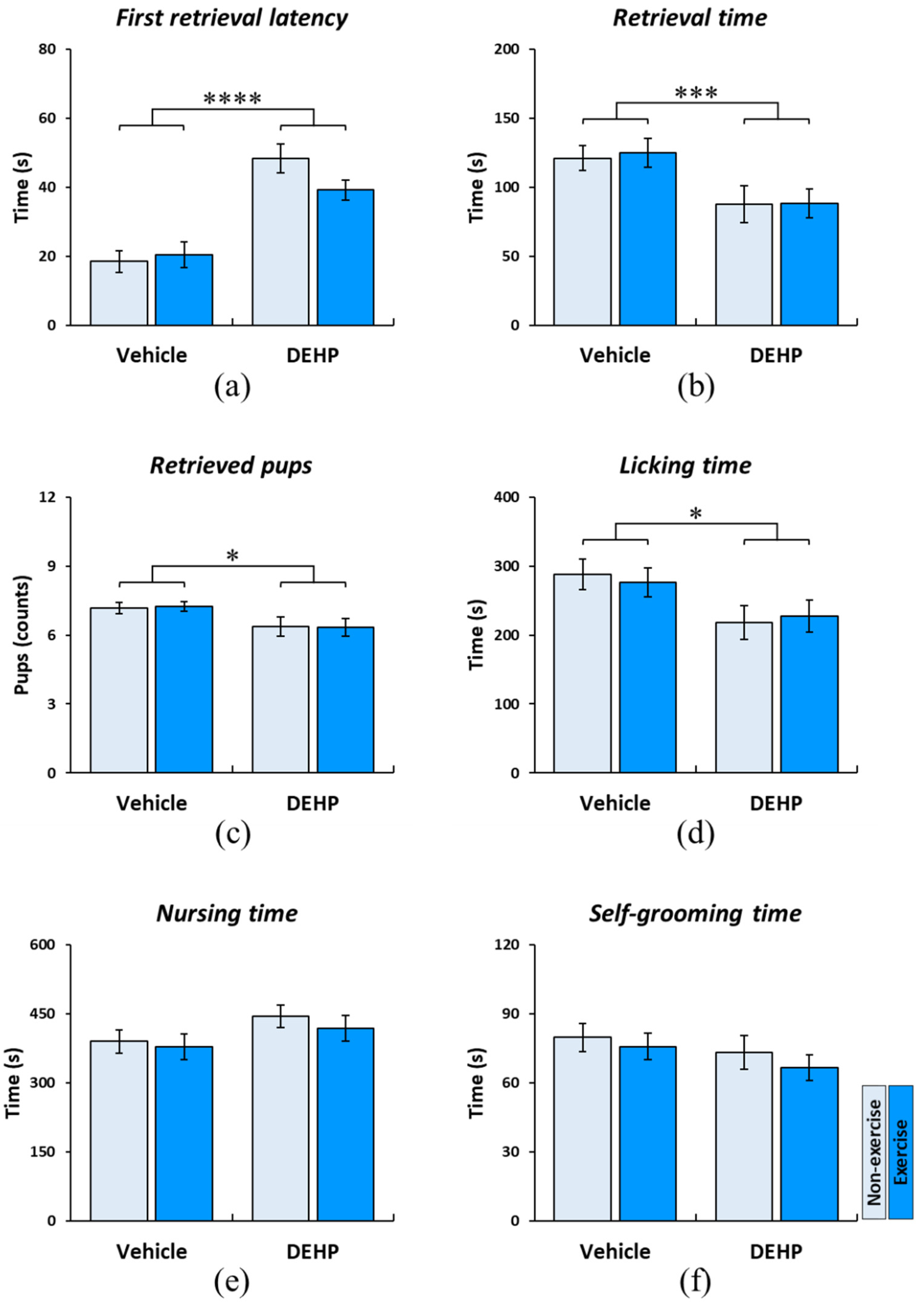
2.3. Effects of Prenatal DEHP Exposure and Childhood Exercise on Maternal Behaviors in Postpartum Dams
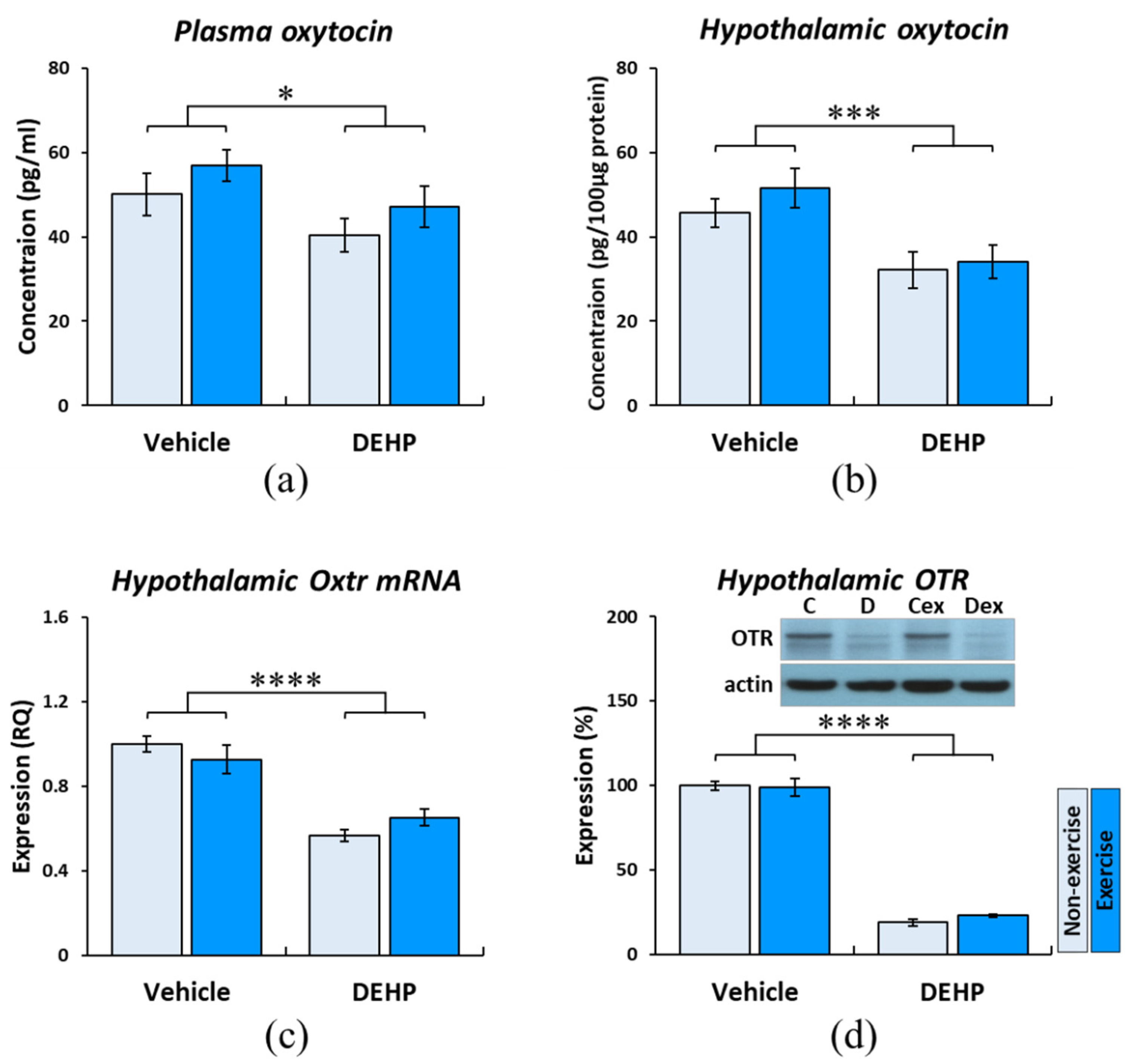
2.4. Effects of Prenatal DEHP Exposure and Childhood Exercise on the Oxytocin Pathway in Postpartum Dams
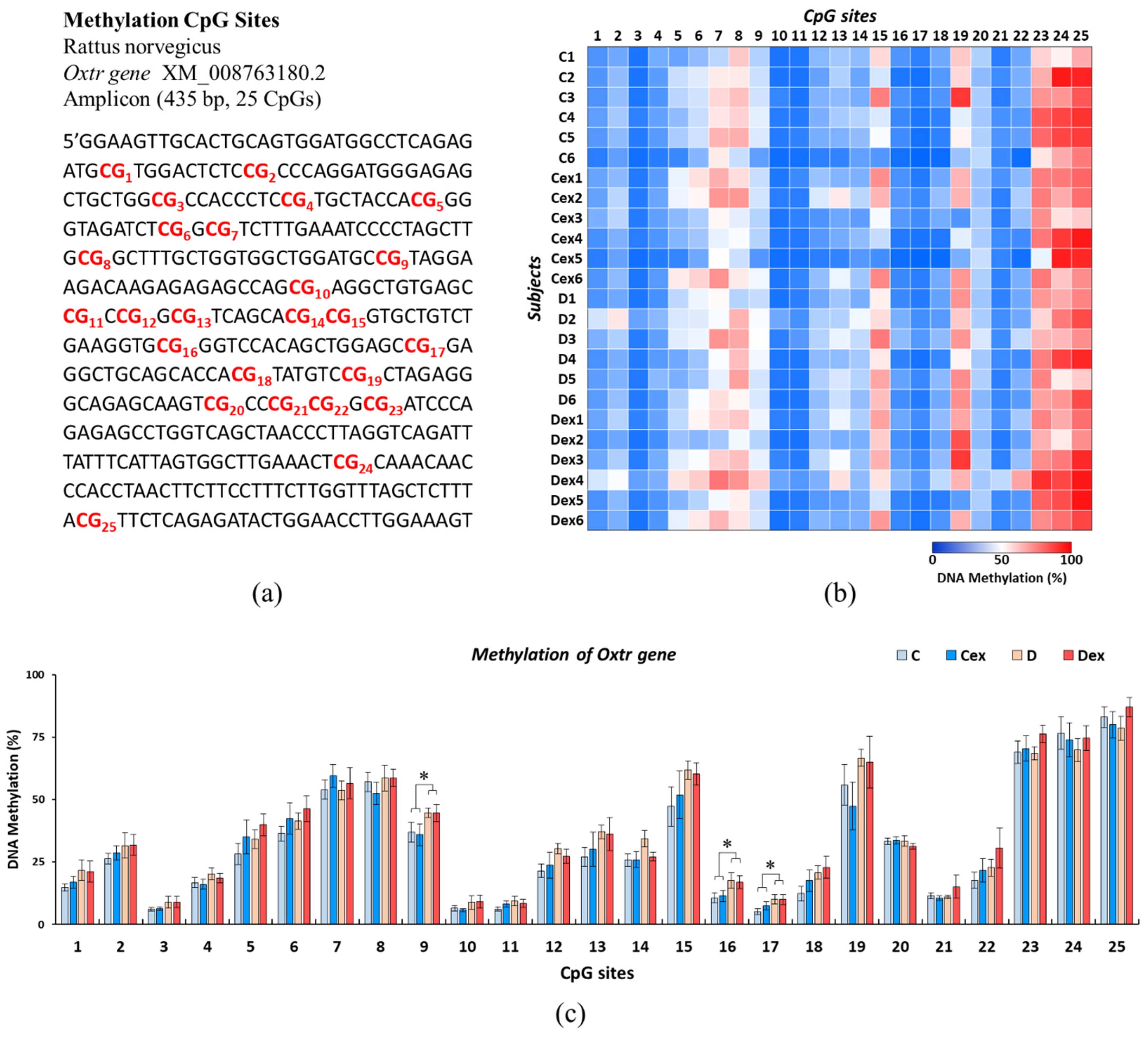
2.5. Effects of Prenatal DEHP Exposure and Childhood Exercise on Oxtr Methylation Levels in Postpartum Dams
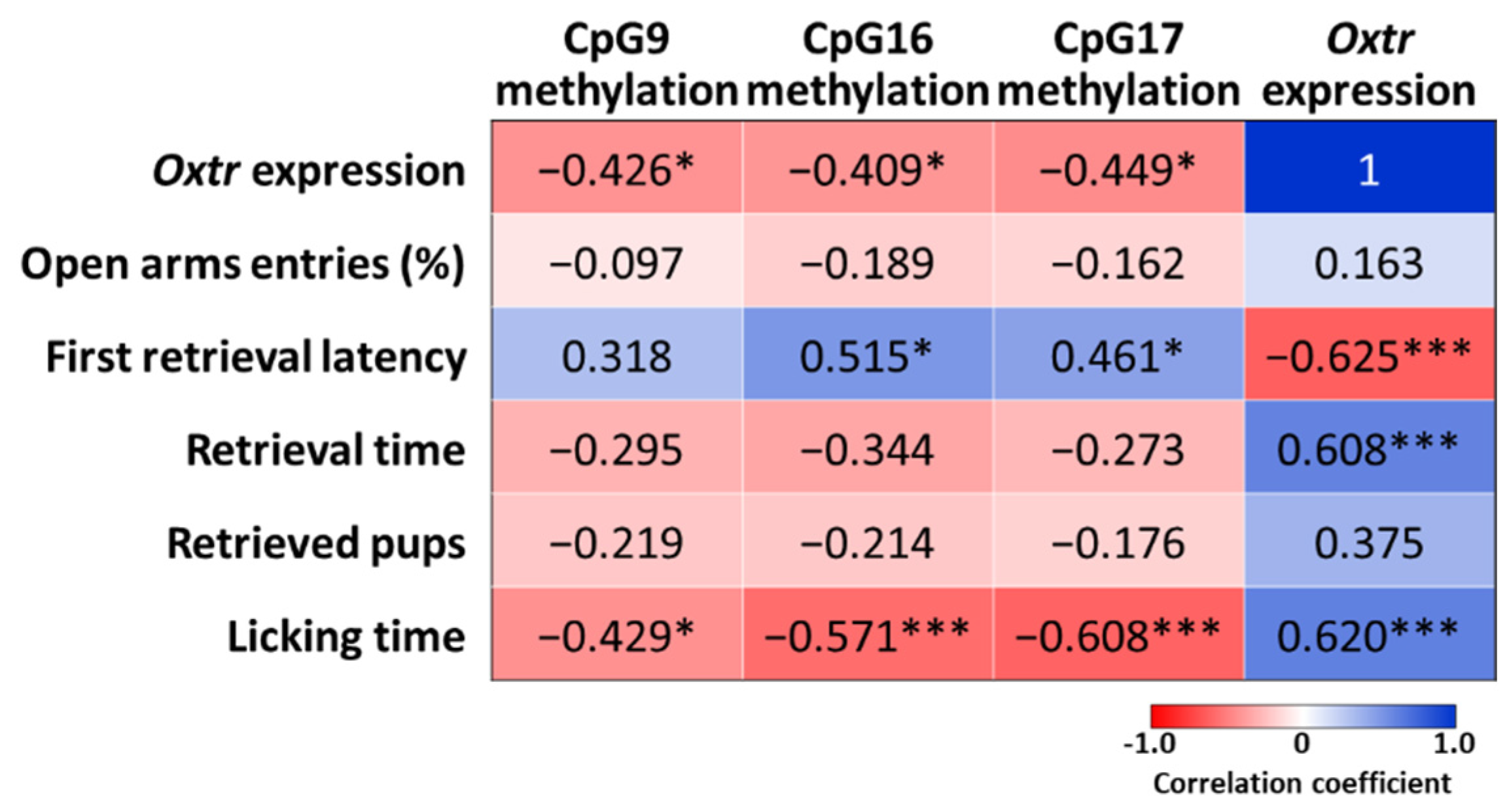
2.6. Correlations between Oxtr Methylation, Oxtr Expression, and Maternal Behaviors
3. Discussion
4. Materials and Methods
4.1. Animals
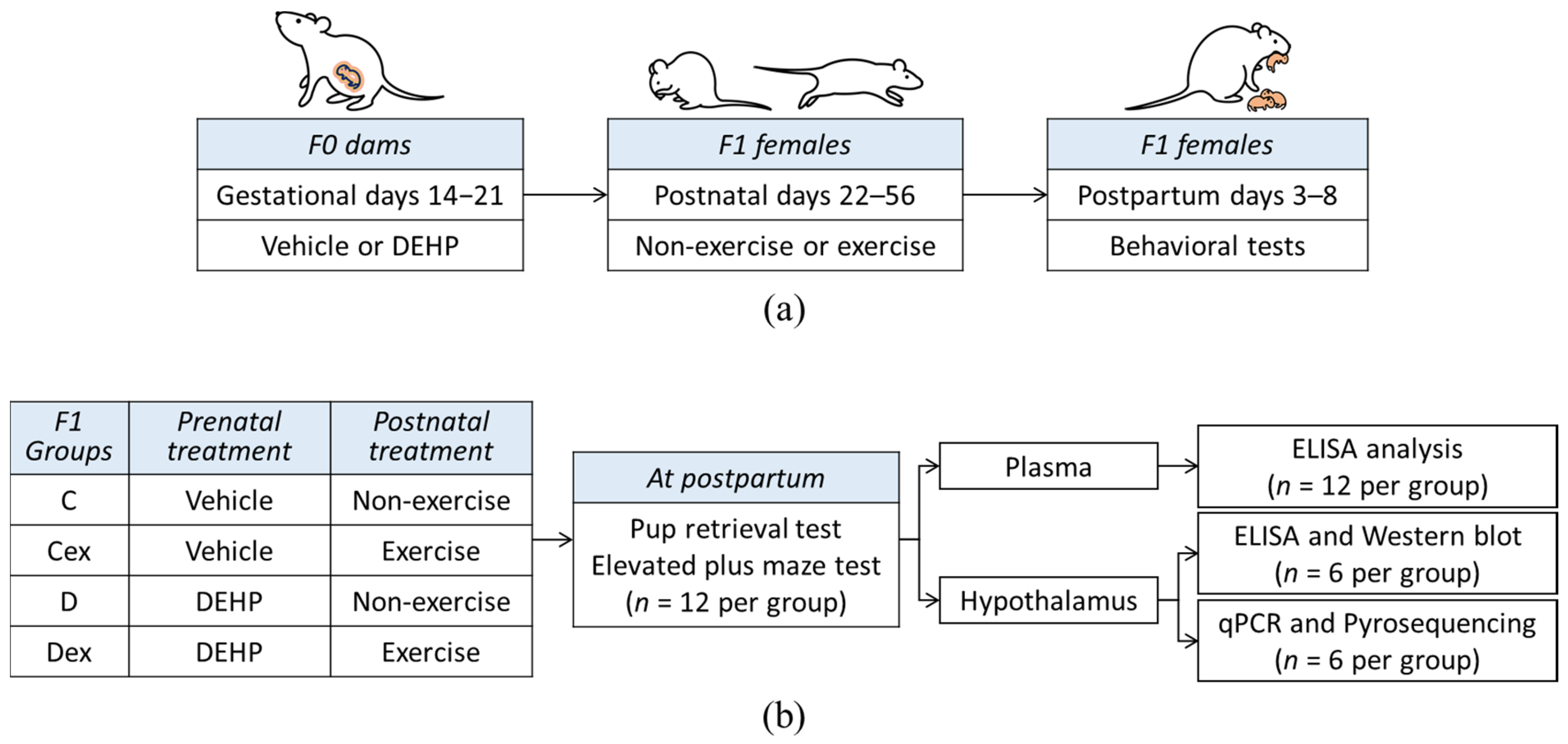
4.2. Experimental Design
4.3. Gestational Administration of DEHP
4.4. Treadmill Running
4.5. Pup Retrieval Test
4.6. Elevated plus Maze
4.7. Blood and Tissue Sample Collection
4.8. Western Blot
4.9. Enzyme-Linked Immunosorbent Assay
4.10. Quantitative Real-Time Polymerase Chain Reaction
4.11. Oxtr Methylation Pyrosequencing
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rilling, J.K.; Young, L.J. The biology of mammalian parenting and its effect on offspring social development. Science 2014, 345, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Strathearn, L. Oxytocin and maternal brain plasticity. New Dir. Child Adolesc. Dev. 2016, 2016, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.; Weller, A.; Zagoory-Sharon, O.; Levine, A. Evidence for a neuroendocrinological foundation of human affiliation: Plasma oxytocin levels across pregnancy and the postpartum period predict mother-infant bonding. Psychol. Sci. 2007, 18, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Fonagy, P.; Koos, O.; Dorsett, K.; Strathearn, L. Maternal oxytocin response predicts mother-to-infant gaze. Brain Res. 2014, 1580, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Guoynes, C.D.; Marler, C.A. An acute dose of intranasal oxytocin rapidly increases maternal communication and maintains maternal care in primiparous postpartum California mice. PLoS ONE 2021, 16, e0244033. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, C.A.; Prange, A.J., Jr. Induction of maternal behavior in virgin rats after intracerebroventricular administration of oxytocin. Proc. Natl. Acad. Sci. USA 1979, 76, 6661–6665. [Google Scholar] [CrossRef] [Green Version]
- van Leengoed, E.; Kerker, E.; Swanson, H.H. Inhibition of post-partum maternal behaviour in the rat by injecting an oxytocin antagonist into the cerebral ventricles. J. Endocrinol. 1987, 112, 275–282. [Google Scholar] [CrossRef]
- Breton, C.; Zingg, H.H. Expression and region-specific regulation of the oxytocin receptor gene in rat brain. Endocrinology 1997, 138, 1857–1862. [Google Scholar] [CrossRef]
- Champagne, F.; Diorio, J.; Sharma, S.; Meaney, M.J. Naturally occurring variations in maternal behavior in the rat are associated with differences in estrogen-inducible central oxytocin receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 12736–12741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, D.D.; Champagne, F.C.; Meaney, M.J. Variations in maternal behaviour are associated with differences in oxytocin receptor levels in the rat. J. Neuroendocrinol. 2000, 12, 1145–1148. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, C.A.; Vadlamudi, S.V.; Boccia, M.L.; Amico, J.A. Maternal behavior deficits in nulliparous oxytocin knockout mice. Genes. Brain Behav. 2006, 5, 274–281. [Google Scholar] [CrossRef]
- Rich, M.E.; deCárdenas, E.J.; Lee, H.J.; Caldwell, H.K. Impairments in the initiation of maternal behavior in oxytocin receptor knockout mice. PLoS ONE 2014, 9, e98839. [Google Scholar] [CrossRef] [Green Version]
- Takayanagi, Y.; Yoshida, M.; Bielsky, I.F.; Ross, H.E.; Kawamata, M.; Onaka, T.; Yanagisawa, T.; Kimura, T.; Matzuk, M.M.; Young, L.J.; et al. Pervasive social deficits, but normal parturition, in oxytocin receptor-deficient mice. Proc. Natl. Acad. Sci. USA 2005, 102, 16096–16101. [Google Scholar] [CrossRef] [Green Version]
- King, L.B.; Walum, H.; Inoue, K.; Eyrich, N.W.; Young, L.J. Variation in the oxytocin receptor gene predicts brain region-specific expression and social attachment. Biol. Psychiatry 2016, 80, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Perkeybile, A.M.; Carter, C.S.; Wroblewski, K.L.; Puglia, M.H.; Kenkel, W.M.; Lillard, T.S.; Karaoli, T.; Gregory, S.G.; Mohammadi, N.; Epstein, L.; et al. Early nurture epigenetically tunes the oxytocin receptor. Psychoneuroendocrinology 2019, 99, 128–136. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maud, C.; Ryan, J.; McIntosh, J.E.; Olsson, C.A. The role of oxytocin receptor gene (OXTR) DNA methylation (DNAm) in human social and emotional functioning: A systematic narrative review. BMC Psychiatry 2018, 18, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beery, A.K.; McEwen, L.M.; MacIsaac, J.L.; Francis, D.D.; Kobor, M.S. Natural variation in maternal care and cross-tissue patterns of oxytocin receptor gene methylation in rats. Horm. Behav. 2016, 77, 42–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregory, S.G.; Connelly, J.J.; Towers, A.J.; Johnson, J.; Biscocho, D.; Markunas, C.A.; Lintas, C.; Abramson, R.K.; Wright, H.H.; Ellis, P.; et al. Genomic and epigenetic evidence for oxytocin receptor deficiency in autism. BMC Med. 2009, 7, 62. [Google Scholar] [CrossRef] [Green Version]
- Bell, A.F.; Carter, C.S.; Steer, C.D.; Golding, J.; Davis, J.M.; Steffen, A.D.; Rubin, L.H.; Lillard, T.S.; Gregory, S.P.; Harris, J.C.; et al. Interaction between oxytocin receptor DNA methylation and genotype is associated with risk of postpartum depression in women without depression in pregnancy. Front. Genet. 2015, 6, 243. [Google Scholar] [CrossRef] [Green Version]
- Ebner, N.C.; Lin, T.; Muradoglu, M.; Weir, D.H.; Plasencia, G.M.; Lillard, T.S.; Pournajafi-Nazarloo, H.; Cohen, R.A.; Sue Carter, C.; Connelly, J.J. Associations between oxytocin receptor gene (OXTR) methylation, plasma oxytocin, and attachment across adulthood. Int. J. Psychophysiol. 2019, 136, 22–32. [Google Scholar] [CrossRef]
- Rubin, L.H.; Connelly, J.J.; Reilly, J.L.; Carter, C.S.; Drogos, L.L.; Pournajafi-Nazarloo, H.; Ruocco, A.C.; Keedy, S.K.; Matthew, I.; Tandon, N.; et al. Sex and diagnosis specific associations between DNA methylation of the oxytocin receptor gene with emotion processing and temporal-limbic and prefrontal brain volumes in psychotic disorders. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2016, 1, 141–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berdasco, M.; Esteller, M. Clinical epigenetics: Seizing opportunities for translation. Nat. Rev. Genet. 2019, 20, 109–127. [Google Scholar] [CrossRef] [PubMed]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A key factor with multipotent impact on brain signaling and synaptic plasticity. Cell. Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Maynard, K.R.; Hobbs, J.W.; Phan, B.N.; Gupta, A.; Rajpurohit, S.; Williams, C.; Rajpurohit, A.; Shin, J.H.; Jaffe, A.E.; Martinowich, K. BDNF-TrkB signaling in oxytocin neurons contributes to maternal behavior. Elife 2018, 7, e33676. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Shahrokh, D.; Hellstrom, I.C.; Wen, X.; Diorio, J.; Breuillaud, L.; Caldji, C.; Meaney, M.J. Brain-derived neurotrophic factor in the nucleus accumbens mediates individual differences in behavioral responses to a natural, social reward. Mol. Neurobiol. 2020, 57, 290–301. [Google Scholar] [CrossRef] [PubMed]
- de Poli, R.A.B.; Lopes, V.H.F.; Lira, F.S.; Zagatto, A.M.; Jimenez-Maldonado, A.; Antunes, B.M. Peripheral BDNF and psycho-behavioral aspects are positively modulated by high-intensity intermittent exercise and fitness in healthy women. Sci. Rep. 2021, 11, 4113. [Google Scholar] [CrossRef]
- Jong, T.R.; Menon, R.; Bludau, A.; Grund, T.; Biermeier, V.; Klampfl, S.M.; Jurek, B.; Bosch, O.J.; Hellhammer, J.; Neumann, I.D. Salivary oxytocin concentrations in response to running, sexual self-stimulation, breastfeeding and the TSST: The Regensburg Oxytocin Challenge (ROC) study. Psychoneuroendocrinology 2015, 62, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Pietrelli, A.; Matković, L.; Vacotto, M.; Lopez-Costa, J.J.; Basso, N.; Brusco, A. Aerobic exercise upregulates the BDNF-Serotonin systems and improves the cognitive function in rats. Neurobiol. Learn. Mem. 2018, 155, 528–542. [Google Scholar] [CrossRef]
- Torner, L.; Plotsky, P.M.; Neumann, I.D.; de Jong, T.R. Forced swimming-induced oxytocin release into blood and brain: Effects of adrenalectomy and corticosterone treatment. Psychoneuroendocrinology 2017, 77, 165–174. [Google Scholar] [CrossRef]
- Ashdown-Franks, G.; Firth, J.; Carney, R.; Carvalho, A.F.; Hallgren, M.; Koyanagi, A.; Rosenbaum, S.; Schuch, F.B.; Smith, L.; Solmi, M.; et al. Exercise as medicine for mental and substance use disorders: A meta-review of the benefits for neuropsychiatric and cognitive outcomes. Sports Med. 2020, 50, 151–170. [Google Scholar] [CrossRef]
- Erickson, K.I.; Hillman, C.; Stillman, C.M.; Ballard, R.M.; Bloodgood, B.; Conroy, D.E.; Macko, R.; Marquez, D.X.; Petruzzello, S.J.; Powell, K.E. Physical activity, cognition, and brain outcomes: A review of the 2018 physical activity guidelines. Med. Sci. Sports Exerc. 2019, 51, 1242–1251. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Pinilla, F.; Zhuang, Y.; Feng, J.; Ying, Z.; Fan, G. Exercise impacts brain-derived neurotrophic factor plasticity by engaging mechanisms of epigenetic regulation. Eur. J. Neurosci. 2011, 33, 383–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seaborne, R.A.; Strauss, J.; Cocks, M.; Shepherd, S.; O’Brien, T.D.; van Someren, K.A.; Bell, P.G.; Murgatroyd, C.; Morton, J.P.; Stewart, C.E.; et al. Human skeletal muscle possesses an epigenetic memory of hypertrophy. Sci. Rep. 2018, 8, 1898. [Google Scholar] [CrossRef]
- Turner, D.C.; Seaborne, R.A.; Sharples, A.P. Comparative transcriptome and methylome analysis in human skeletal muscle anabolism, hypertrophy and epigenetic memory. Sci. Rep. 2019, 9, 4251. [Google Scholar] [CrossRef] [Green Version]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2: The Endocrine Society’s Second Scientific Statement on Endocrine-Disrupting Chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef]
- Martinez-Arguelles, D.B.; Papadopoulos, V. Prenatal phthalate exposure: Epigenetic changes leading to lifelong impact on steroid formation. Andrology 2016, 4, 573–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Högberg, J.; Hanberg, A.; Berglund, M.; Skerfving, S.; Remberger, M.; Calafat, A.M.; Filipsson, A.F.; Jansson, B.; Johansson, N.; Appelgren, M.; et al. Phthalate diesters and their metabolites in human breast milk, blood or serum, and urine as biomarkers of exposure in vulnerable populations. Environ. Health Perspect. 2008, 116, 334–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.; Ku, H.Y.; Su, P.H.; Chen, J.W.; Huang, P.C.; Angerer, J.; Wang, S.L. Phthalate exposure in pregnant women and their children in central Taiwan. Chemosphere 2011, 82, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Carbone, S.; Ponzo, O.J.; Gobetto, N.; Samaniego, Y.A.; Reynoso, R.; Moguilevsky, J.A.; Cutrera, R.A. Effect of di(2-ethylhexyl) phthalate on the neuroendocrine regulation of reproduction in adult male rats and its relationship to anxiogenic behavior: Participation of GABAergic system. Hum. Exp. Toxicol. 2019, 38, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Shao, P.; Wang, Y.; Zhang, M.; Wen, X.; Zhang, J.; Xu, Z.; Hu, M.; Jiang, J.; Liu, T. The interference of DEHP in precocious puberty of females mediated by the hypothalamic IGF-1/PI3K/Akt/mTOR signaling pathway. Ecotoxicol. Environ. Saf. 2019, 181, 362–369. [Google Scholar] [CrossRef]
- Carbone, S.; Ponzo, O.J.; Gobetto, N.; Samaniego, Y.A.; Reynoso, R.; Scacchi, P.; Moguilevsky, J.A.; Cutrera, R. Antiandrogenic effect of perinatal exposure to the endocrine disruptor di-(2-ethylhexyl) phthalate increases anxiety-like behavior in male rats during sexual maturation. Horm. Behav. 2013, 63, 692–699. [Google Scholar] [CrossRef]
- Wang, D.C.; Chen, T.J.; Lin, M.L.; Jhong, Y.C.; Chen, S.C. Exercise prevents the increased anxiety-like behavior in lactational di-(2-ethylhexyl) phthalate-exposed female rats in late adolescence by improving the regulation of hypothalamus-pituitary-adrenal axis. Horm. Behav. 2014, 66, 674–684. [Google Scholar] [CrossRef]
- Kougias, D.G.; Cortes, L.R.; Moody, L.; Rhoads, S.; Pan, Y.X.; Juraska, J.M. Effects of perinatal exposure to phthalates and a high-fat diet on maternal behavior and pup development and social play. Endocrinology 2018, 159, 1088–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinnies, K.M.; Harris, E.P.; Snyder, R.W.; Sumner, S.S.; Rissman, E.F. Direct and transgenerational effects of low doses of perinatal di-(2-ethylhexyl) phthalate (DEHP) on social behaviors in mice. PLoS ONE 2017, 12, e0171977. [Google Scholar] [CrossRef] [PubMed]
- Kardas, F.; Bayram, A.K.; Demirci, E.; Akin, L.; Ozmen, S.; Kendirci, M.; Canpolat, M.; Oztop, D.B.; Narin, F.; Gumus, H.; et al. Increased serum phthalates (MEHP, DEHP) and bisphenol A concentrations in children with autism spectrum disorder: The role of endocrine disruptors in autism etiopathogenesis. J. Child Neurol. 2016, 31, 629–635. [Google Scholar] [CrossRef]
- Ku, H.Y.; Tsai, T.L.; Wang, P.L.; Su, P.H.; Sun, C.W.; Wang, C.J.; Wang, S.L. Prenatal and childhood phthalate exposure and attention deficit hyperactivity disorder traits in child temperament: A 12-year follow-up birth cohort study. Sci. Total. Environ. 2020, 699, 134053. [Google Scholar] [CrossRef] [PubMed]
- Siu, M.T.; Goodman, S.J.; Yellan, I.; Butcher, D.T.; Jangjoo, M.; Grafodatskaya, D.; Rajendram, R.; Lou, Y.; Zhang, R.; Zhao, C.; et al. DNA methylation of the oxytocin receptor across neurodevelopmental disorders. J. Autism Dev. Disord. 2021, 1–14. [Google Scholar] [CrossRef]
- Orso, R.; Creutzberg, K.C.; Wearick-Silva, L.E.; Wendt Viola, T.; Tractenberg, S.G.; Benetti, F.; Grassi-Oliveira, R. How early life stress impact maternal care: A systematic review of rodent studies. Front. Behav. Neurosci. 2019, 13, 197. [Google Scholar] [CrossRef]
- Andrade, A.J.; Grande, S.W.; Talsness, C.E.; Grote, K.; Chahoud, I. A dose-response study following in utero and lactational exposure to di-(2-ethylhexyl)-phthalate (DEHP): Non-monotonic dose-response and low dose effects on rat brain aromatase activity. Toxicology 2006, 227, 185–192. [Google Scholar] [CrossRef]
- Carbone, S.; Samaniego, Y.A.; Cutrera, R.; Reynoso, R.; Cardoso, N.; Scacchi, P.; Moguilevsky, J.A.; Ponzo, O.J. Different effects by sex on hypothalamic-pituitary axis of prepubertal offspring rats produced by in utero and lactational exposure to di-(2-ethylhexyl) phthalate (DEHP). Neurotoxicology 2012, 33, 78–84. [Google Scholar] [CrossRef]
- Gao, N.; Hu, R.; Huang, Y.; Dao, L.; Zhang, C.; Liu, Y.; Wu, L.; Wang, X.; Yin, W.; Gore, A.C.; et al. Specific effects of prenatal DEHP exposure on neuroendocrine gene expression in the developing hypothalamus of male rats. Arch. Toxicol. 2018, 92, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Kaffman, A.; Meaney, M.J. Neurodevelopmental sequelae of postnatal maternal care in rodents: Clinical and research implications of molecular insights. J. Child Psychol. Psychiatry 2007, 48, 224–244. [Google Scholar] [CrossRef]
- Grinevich, V.; Desarménien, M.G.; Chini, B.; Tauber, M.; Muscatelli, F. Ontogenesis of oxytocin pathways in the mammalian brain: Late maturation and psychosocial disorders. Front. Neuroanat. 2015, 8, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Wang, K.; Svoboda, L.K.; Rygiel, C.A.; Neier, K.; Jones, T.R.; Cavalcante, R.G.; Colacino, J.A.; Dolinoy, D.C.; Sartor, M.A. Perinatal DEHP exposure induces sex- and tissue-specific DNA methylation changes in both juvenile and adult mice. Environ. Epigenet. 2021, 7, dvab004. [Google Scholar] [CrossRef]
- Nadeem, A.; Al-Harbi, N.O.; Ahmad, S.F.; Alhazzani, K.; Attia, S.M.; Alsanea, S.; Alhoshani, A.; Mahmood, H.M.; Alfardan, A.S.; Bakheet, S.A. Exposure to the plasticizer, Di-(2-ethylhexyl) phthalate during juvenile period exacerbates autism-like behavior in adult BTBR T + tf/J mice due to DNA hypomethylation and enhanced inflammation in brain and systemic immune cells. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 109, 110249. [Google Scholar] [CrossRef]
- Svoboda, L.K.; Wang, K.; Cavalcante, R.G.; Neier, K.; Colacino, J.A.; Sartor, M.A.; Dolinoy, D.C. Sex-specific programming of cardiac DNA methylation by developmental phthalate exposure. Epigenet. Insights 2020, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Danoff, J.S.; Wroblewski, K.L.; Graves, A.J.; Quinn, G.C.; Perkeybile, A.M.; Kenkel, W.M.; Lillard, T.S.; Parikh, H.I.; Golino, H.F.; Gregory, S.G.; et al. Genetic, epigenetic, and environmental factors controlling oxytocin receptor gene expression. Clin. Epigenetics 2021, 13, 23. [Google Scholar] [CrossRef]
- Harony-Nicolas, H.; Mamrut, S.; Brodsky, L.; Shahar-Gold, H.; Barki-Harrington, L.; Wagner, S. Brain region-specific methylation in the promoter of the murine oxytocin receptor gene is involved in its expression regulation. Psychoneuroendocrinology 2014, 39, 121–131. [Google Scholar] [CrossRef]
- Pérez, P.A.; Toledo, J.; Sosa, L.D.V.; Peinetti, N.; Torres, A.I.; De Paul, A.L.; Gutiérrez, S. The phthalate DEHP modulates the estrogen receptors alpha and beta increasing lactotroph cell population in female pituitary glands. Chemosphere 2020, 258, 127304. [Google Scholar] [CrossRef] [PubMed]
- Phillips, N.L.H.; Roth, T.L. Animal models and their contribution to our understanding of the relationship between environments, epigenetic modifications, and behavior. Genes 2019, 10, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, K.T.; Warren, M.; Roth, T.L. The influence of infant-caregiver experiences on amygdala Bdnf, OXTr, and NPY expression in developing and adult male and female rats. Behav. Brain Res. 2014, 272, 175–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peña, C.J.; Neugut, Y.D.; Champagne, F.A. Developmental timing of the effects of maternal care on gene expression and epigenetic regulation of hormone receptor levels in female rats. Endocrinology 2013, 154, 4340–4351. [Google Scholar] [CrossRef] [Green Version]
- Gouin, J.P.; Zhou, Q.Q.; Booij, L.; Boivin, M.; Côté, S.M.; Hébert, M.; Ouellet-Morin, I.; Szyf, M.; Tremblay, R.E.; Turecki, G.; et al. Associations among oxytocin receptor gene (OXTR) DNA methylation in adulthood, exposure to early life adversity, and childhood trajectories of anxiousness. Sci. Rep. 2017, 7, 7446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unternaehrer, E.; Meyer, A.H.; Burkhardt, S.C.; Dempster, E.; Staehli, S.; Theill, N.; Lieb, R.; Meinlschmidt, G. Childhood maternal care is associated with DNA methylation of the genes for brain-derived neurotrophic factor (BDNF) and oxytocin receptor (OXTR) in peripheral blood cells in adult men and women. Stress 2015, 18, 451–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, S.M.; Doherty, T.S.; Roth, T.L. Pharmacological manipulation of DNA methylation normalizes maternal behavior, DNA methylation, and gene expression in dams with a history of maltreatment. Sci. Rep. 2019, 9, 10253. [Google Scholar] [CrossRef]
- Smith, A.S.; Tabbaa, M.; Lei, K.; Eastham, P.; Butler, M.J.; Linton, L.; Altshuler, R.; Liu, Y.; Wang, Z. Local oxytocin tempers anxiety by activating GABAA receptors in the hypothalamic paraventricular nucleus. Psychoneuroendocrinology 2016, 63, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Shi, C.; Li, X.; Zhang, P.; Liu, B.; Wang, H.; Wang, Y.; Yang, Y.; Wu, Y.; Li, H.; et al. Injection of oxytocin into paraventricular nucleus reverses depressive-like behaviours in the postpartum depression rat model. Behav. Brain Res. 2018, 336, 236–243. [Google Scholar] [CrossRef]
- Cox, E.Q.; Stuebe, A.; Pearson, B.; Grewen, K.; Rubinow, D.; Meltzer-Brody, S. Oxytocin and HPA stress axis reactivity in postpartum women. Psychoneuroendocrinology 2015, 55, 164–172. [Google Scholar] [CrossRef] [Green Version]
- Sabihi, S.; Dong, S.M.; Durosko, N.E.; Leuner, B. Oxytocin in the medial prefrontal cortex regulates maternal care, maternal aggression and anxiety during the postpartum period. Front. Behav. Neurosci. 2014, 8, 258. [Google Scholar] [CrossRef] [Green Version]
- Ruthschilling, C.A.; Albiero, G.; Lazzari, V.M.; Becker, R.O.; de Moura, A.C.; Lucion, A.B.; Almeida, S.; Veiga, A.B.; Giovenardi, M. Analysis of transcriptional levels of the oxytocin receptor in different areas of the central nervous system and behaviors in high and low licking rats. Behav. Brain Res. 2012, 228, 176–184. [Google Scholar] [CrossRef]
- Roepke, T.A.; Yang, J.A.; Yasrebi, A.; Mamounis, K.J.; Oruc, E.; Zama, A.M.; Uzumcu, M. Regulation of arcuate genes by developmental exposures to endocrine-disrupting compounds in female rats. Reprod. Toxicol. 2016, 62, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.A.; Holahan, M.R. Reduced hippocampal dendritic spine density and BDNF expression following acute postnatal exposure to di(2-ethylhexyl) phthalate in male Long Evans rats. PLoS ONE 2014, 9, e109522. [Google Scholar]
- Jobst, A.; Krause, D.; Maiwald, C.; Härtl, K.; Myint, A.M.; Kästner, R.; Obermeier, M.; Padberg, F.; Brücklmeier, B.; Weidinger, E.; et al. Oxytocin course over pregnancy and postpartum period and the association with postpartum depressive symptoms. Arch. Womens Ment. Health 2016, 19, 571–579. [Google Scholar] [CrossRef]
- Schechter, M.; Weller, A.; Pittel, Z.; Gross, M.; Zimmer, A.; Pinhasov, A. Endocannabinoid receptor deficiency affects maternal care and alters the dam’s hippocampal oxytocin receptor and brain-derived neurotrophic factor expression. J. Neuroendocrinol. 2013, 25, 898–909. [Google Scholar] [CrossRef] [PubMed]
- Branchi, I.; Curley, J.P.; D’Andrea, I.; Cirulli, F.; Champagne, F.A.; Alleva, E. Early interactions with mother and peers independently build adult social skills and shape BDNF and oxytocin receptor brain levels. Psychoneuroendocrinology 2013, 38, 522–532. [Google Scholar] [CrossRef] [Green Version]
- Lommatzsch, M.; Hornych, K.; Zingler, C.; Schuff-Werner, P.; Höppner, J.; Virchow, J.C. Maternal serum concentrations of BDNF and depression in the perinatal period. Psychoneuroendocrinology 2006, 31, 388–394. [Google Scholar] [CrossRef]
- Uvnäs-Moberg, K.; Ekström-Bergström, A.; Berg, M.; Buckley, S.; Pajalic, Z.; Hadjigeorgiou, E.; Kotłowska, A.; Lengler, L.; Kielbratowska, B.; Leon-Larios, F.; et al. Maternal plasma levels of oxytocin during physiological childbirth—A systematic review with implications for uterine contractions and central actions of oxytocin. BMC Pregnancy Childbirth 2019, 19, 285. [Google Scholar] [CrossRef] [Green Version]
- Bakos, J.; Hlavacova, N.; Makatsori, A.; Tybitanclova, K.; Zorad, S.; Hinghofer-Szalkay, H.; Johansson, B.B.; Jezova, D. Oxytocin levels in the posterior pituitary and in the heart are modified by voluntary wheel running. Regul. Pept. 2007, 139, 96–101. [Google Scholar] [CrossRef]
- Sun, G.C.; Lee, Y.J.; Lee, Y.C.; Yu, H.F.; Wang, D.C. Exercise prevents the impairment of learning and memory in prenatally phthalate-exposed male rats by improving the expression of plasticity-related proteins. Behav. Brain Res. 2021, 413, 113444. [Google Scholar] [CrossRef]
- Wang, D.C.; Lin, H.T.; Lee, Y.J.; Yu, H.F.; Wu, S.R.; Qamar, M.U. Recovery of BDNF and CB1R in the prefrontal cortex underlying improvement of working memory in prenatal DEHP-exposed male rats after aerobic exercise. Int. J. Mol. Sci. 2020, 21, 3867. [Google Scholar] [CrossRef]
- Szuhany, K.L.; Bugatti, M.; Otto, M.W. A meta-analytic review of the effects of exercise on brain-derived neurotrophic factor. J. Psychiatr. Res. 2015, 60, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Venezia, A.C.; Guth, L.M.; Sapp, R.M.; Spangenburg, E.E.; Roth, S.M. Sex-dependent and independent effects of long-term voluntary wheel running on Bdnf mRNA and protein expression. Physiol. Behav. 2016, 156, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Onakomaiya, M.M.; Porter, D.M.; Oberlander, J.G.; Henderson, L.P. Sex and exercise interact to alter the expression of anabolic androgenic steroid-induced anxiety-like behaviors in the mouse. Horm. Behav. 2014, 66, 283–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanehkar, F.; Rashidy-Pour, A.; Vafaei, A.A.; Sameni, H.R.; Haghighi, S.; Miladi-Gorji, H.; Motamedi, F.; Akhavan, M.M.; Bavarsad, K. Voluntary exercise does not ameliorate spatial learning and memory deficits induced by chronic administration of nandrolone decanoate in rats. Horm. Behav. 2013, 63, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Shelby, M.D. NTP-CERHR monograph on the potential human reproductive and developmental effects of di(2-ethylhexyl) phthalate (DEHP). Ntp. Cerhr. Mon. 2006, 18, v, vii-7, II-iii-xiii passim. [Google Scholar]
- Campioli, E.; Martinez-Arguelles, D.B.; Papadopoulos, V. In utero exposure to the endocrine disruptor di-(2-ethylhexyl) phthalate promotes local adipose and systemic inflammation in adult male offspring. Nutr. Diabetes 2014, 4, e115. [Google Scholar] [CrossRef] [Green Version]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.T.; Chen, Y.C.; Wu, C.W.; Yu, L.; Chen, H.I.; Jen, C.J.; Kuo, Y.M. Glucocorticoid signaling and exercise-induced downregulation of the mineralocorticoid receptor in the induction of adult mouse dentate neurogenesis by treadmill running. Psychoneuroendocrinology 2008, 33, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Carini, L.M.; Nephew, B.C. Effects of early life social stress on endocrinology, maternal behavior, and lactation in rats. Horm. Behav. 2013, 64, 634–641. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Observed Categories | Groups (Mean ± SEM) | Effect (F Value) | ||||
|---|---|---|---|---|---|---|
| C | Cex | D | Dex | DEHP | Exercise | |
| Number of open arms entries (counts) | 9.42 ± 1.47 | 9.92 ± 1.82 | 8.17 ± 1.81 | 8.83 ± 1.87 | 0.444 | 0.111 |
| Number of close arms entries (counts) | 15.33 ± 2.06 | 15.35 ± 1.91 | 16.17 ± 2.39 | 17.25 ± 2.34 | 0.145 | 0.002 |
| Number of open arms entries (%) | 38.47 ± 3.68 | 37.83 ± 2.34 | 31.77 ± 1.88 | 32.06 ± 2.39 | 2.336 | 0.077 |
| Time of open arms entries (s) | 67.00 ± 8.03 | 66.33 ± 9.29 | 63.25 ± 9.66 | 64.92 ± 9.22 | 0.081 | 0.003 |
| Time of close arms entries (s) | 184.50 ± 7.84 | 190.25 ± 6.38 | 193.75 ± 8.05 | 194.50 ± 8.53 | 0.760 | 0.176 |
| Time of open arms entries (%) | 22.33 ± 2.68 | 22.11 ± 3.10 | 21.08 ± 3.22 | 21.64 ± 3.07 | 0.081 | 0.003 |
| Measured Concentrations | Groups (Mean ± SEM) | Effect (F Value) | ||||
|---|---|---|---|---|---|---|
| C | Cex | D | Dex | DEHP | Exercise | |
| Plasma BDNF (pg/mL) | 348.72 ± 31.10 | 375.33 ± 27.92 | 308.97 ± 24.52 | 330.01 ± 24.02 | 2.475 | 0.777 |
| Hypothalamic BDNF (pg/100 μg protein) | 128.26 ± 12.46 | 131.88 ± 9.27 | 121.60 ± 9.17 | 117.63 ± 7.65 | 1.138 | 0.000 |
| Plasma ACTH (pg/mL) | 115.65 ± 5.11 | 112.67 ± 4.92 | 128.96 ± 4.96 | 121.50 ± 4.79 | 5.011 * | 1.115 |
| Plasma corticosterone (ng/mL) | 239.43 ± 22.42 | 241.72 ± 18.74 | 303.83 ± 27.73 | 263.02 ± 28.59 | 3.009 | 0.608 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-J.; Lin, H.-T.; Chaudhary, M.A.; Lee, Y.-C.; Wang, D.-C. Effects of Prenatal Phthalate Exposure and Childhood Exercise on Maternal Behaviors in Female Rats at Postpartum: A Role of Oxtr Methylation in the Hypothalamus. Int. J. Mol. Sci. 2021, 22, 9847. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22189847
Lee Y-J, Lin H-T, Chaudhary MA, Lee Y-C, Wang D-C. Effects of Prenatal Phthalate Exposure and Childhood Exercise on Maternal Behaviors in Female Rats at Postpartum: A Role of Oxtr Methylation in the Hypothalamus. International Journal of Molecular Sciences. 2021; 22(18):9847. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22189847
Chicago/Turabian StyleLee, Yi-Ju, Hwai-Ting Lin, Muhammad Asad Chaudhary, Yi-Ching Lee, and Dean-Chuan Wang. 2021. "Effects of Prenatal Phthalate Exposure and Childhood Exercise on Maternal Behaviors in Female Rats at Postpartum: A Role of Oxtr Methylation in the Hypothalamus" International Journal of Molecular Sciences 22, no. 18: 9847. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22189847