
Enteric Microbiota-Mediated Serotonergic Signaling in Pathogenesis of Irritable Bowel Syndrome


Department of Internal Medicine II, Shimane University Faculty of Medicine, Izumo 693-8501, Japan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(19), 10235; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910235
Submission received: 1 September 2021
/
Revised: 18 September 2021
/
Accepted: 19 September 2021
/
Published: 23 September 2021
(This article belongs to the Special Issue Pathophysiology of the Serotonin System in the Nervous System and Beyond)
Abstract
:Irritable bowel syndrome (IBS) is a chronic functional disorder that affects the gastrointestinal tract. Details regarding the pathogenesis of IBS remain largely unknown, though the dysfunction of the brain-gut-microbiome (BGM) axis is a major etiological factor, in which neurotransmitters serve as a key communication tool between enteric microbiota and the brain. One of the most important neurotransmitters in the pathology of IBS is serotonin (5-HT), as it influences gastrointestinal motility, pain sensation, mucosal inflammation, immune responses, and brain activity, all of which shape IBS features. Genome-wide association studies discovered susceptible genes for IBS in serotonergic signaling pathways. In clinical practice, treatment strategies targeting 5-HT were effective for a certain portion of IBS cases. The synthesis of 5-HT in intestinal enterochromaffin cells and host serotonergic signaling is regulated by enteric resident microbiota. Dysbiosis can trigger IBS development, potentially through aberrant 5-HT signaling in the BGM axis; thus, the manipulation of the gut microbiota may be an alternative treatment strategy. However, precise information regarding the mechanisms underlying the microbiota-mediated intestinal serotonergic pathway related to the pathogenesis of IBS remains unclear. The present review summarizes current knowledge and recent progress in understanding microbiome–serotonin interaction in IBS cases.
1. Background
Irritable bowel syndrome (IBS), a gastrointestinal (GI) disorder characterized by chronic abdominal pain with aberrant bowel movements in the absence of nondetectable causes [1,2], is the most common GI disease, with global prevalence estimated to be 10–15% [3]. Rome IV criteria are commonly used to diagnose this disorder in clinical practice, which classify cases into four different subtypes on the basis of bowel habits and fecal condition: constipation-predominant (IBS-C), diarrhea-predominant (IBS-D), mixed (IBS-M), and unclassified (IBS-U) [4,5].
Details regarding the pathogenic mechanisms remain largely unclear; thus, current treatments for IBS are mostly focused on symptoms, with limited efficacy, and not as radical care [6,7,8]. Although this disorder is not associated with increased risk of mortality [9], a number of IBS patients show markedly decreased quality of life, including poor socioeconomical activities as a result of IBS [2,10]; thus, the high prevalence of chronic features in affected patients places a financial burden on global healthcare systems [11,12,13]. As a result, the introduction of an effective treatment strategy based on mechanistic studies is highly anticipated.
Numerous basic and clinical studies were conducted to clarify the cause of IBS from multiple aspects, including genetic factors, low-grade mucosal inflammation and immune activation following a severe GI infection, increased gut mucosal permeability, alterations in gut microbiota, aberrant bile salt metabolism, hypersensitivity to particular diet components, abnormal neurotransmitter pathways, and altered central nervous system (CNS) processing [14,15,16,17,18]. Although each factor contributes to forming a certain portion of the IBS etiology, clinical features of the disorder are heterogeneous, and likely created by a mixture of genetic and environmental factors [15,16]. Therefore, the determination of and focus on a specific target molecule or pathway related to IBS pathogenies is challenging, though recent progress in omics technologies, including epigenomics, metabolomics, transcriptomics, and proteomics, provided efficient methods for identification of new pathways and potential targets [19,20,21]. These novel approaches provided results suggesting the importance of interactions between enteric microbiota and neurotransmitters, and their pathways in the context of IBS pathogenesis [15,18,22,23].
Neurotransmitters are chemical substances that transmit signals between neurons and target cells, such as muscles, glands, and other neurons, throughout the body [24]. Enteric microbiota play a critical role in regulating a variety of neurotransmitters, including histamine, serotonin (5-hydroxytryptamine, 5-HT), glutamate, γ-aminobutyric acid (GABA), dopamine, acetylcholine, and catecholamines [18,22,24], and each biological substance influences the activity of the enteric nervous system (ENS) independently or corporately as part of the pathology of IBS [22,25]. Among those, 5-HT is one of the most well-studied neurotransmitters in IBS research investigations [26,27,28]. Enteric 5-HT boosts visceral hypersensitivity, increases mucosal permeability, alters gut motility, activates the immune system, and induces inflammation, which synergistically contribute to forming IBS symptoms [28]. Indeed, pharmacological interventions regarding 5-HT receptors are commonly used for treating IBS patients in current clinical practice [7,29]. Overall, 5-HT in association with enteric microbiota is a promising target for both medical care and IBS research.
Nevertheless, unanswered questions remain before fully revealing the etiology of IBS because of insufficient evidence, likely due to abundant limitations in IBS research [22,30] because it is often difficult to acquire quantified reproducible data when investigating functional disorders such as IBS for clinical and preclinical studies. Clinical studies of IBS patients tend to show huge placebo effects, which make the obtained results difficult to interpret [1,31]. Unlike other GI disorders, such as inflammatory bowel disease (IBD), there are neither reliable tests nor universally accepted biomarkers available to diagnose and evaluate IBS-related disease activity [5,30,32]. In preclinical studies, several animal IBS models that show abnormal intestinal motility and/or visceral hypersensitivity have been established. However, those are completely different from human IBS in terms of dietary contents and habits, social behavior, mental status, the intestinal immune system, and resident enteric microbiota [30,33,34]. Additional technology and knowledge for dealing with these limitations are needed.
This review summarizes existing evidence and recent progress in the involvement of gut microbiota-mediated 5-HT in IBS. Additional research targeting the interactions between enteric microbiota and serotonergic signaling could provide deeper understanding and mechanistic insight regarding the pathology of IBS.
2. Brain–Gut–Microbiome Interactions in IBS
Trillions of gut microbes coexist in humans and supply a variety of beneficial functions to the host, such as creating essential nutrients and vitamins from indigestible or poorly absorbable dietary contents, training the immune system, and limiting the settlement or growth of harmful microorganisms [35,36,37]. An alternation of the gut microbial population, termed dysbiosis, is potentially associated with both GI and non-GI disorders, such as diabetes, obesity, chronic kidney disease, and several psychiatric and neurologic disorders [38,39,40]. Unfortunately, the underlying causes and mechanisms of dysbiosis in related diseases are largely unknown. In fact, it is yet to be shown whether dysbiosis is a cause or consequence of a target disease, while even a “healthy microbiome” is poorly defined, which is a million-dollar question [41,42,43].
Numerous research studies were conducted to investigate mechanisms related to the influence of the gut microbiota on health and disorders beyond the GI tract, particularly in the brain [44]. Some preclinical and clinical studies showed that gut microbes communicate with the CNS through multiple channels, including nervous, endocrine, and immune signaling pathways [23,45,46]. Additionally, the brain influences the intestinal microenvironment by modulating gut motility, secretion, and permeability through the neuron–glia–epithelium axis and visceral nerves [23,44,45,46]. On the basis of these findings, the concept of the brain–gut–microbiome (BGM) axis was proposed to elucidate bidirectional communication between gut microbiota and the CNS [23,45,46].
The dysfunction of BGM interactions is a central pathological factor in the context of IBS [22,23,45,46], and dysbiosis was observed in patients with IBS in most related clinical studies [47,48,49]. Traditional fecal analysis results demonstrated a certain bacterial population in IBS patients that is distinctively different from that in healthy individuals, with lower bacterial diversity [40,50,51,52,53]. Fecal samples from IBS patients show a higher Firmicutes/Bacteroidetes ratio, lower abundance of Lactobacillus and Bifidobacterium, and higher levels of Escherichia coli and Enterobacter [30,54,55,56,57]. In addition, Clostridiales I, Faecalibacterium, and Bifidobacterium genera were reported to be decreased in IBS patients [58]. Recently, the presence of mucosal biofilm consisting of an overgrowth of E. coli and Ruminococcus gnavus was proposed to be an endoscopic feature in a subgroup of IBS patients [59]. That study noted that such biofilms contain high concentrations of bile acids that can induce bile acid-dependent abnormalities in bowel movements [59]. These observations suggest that reversing the altered bacterial composition, along with normalizing the BGM axis, may be an ideal treatment strategy for IBS, though it is unknown if dysbiosis has a causal effect for IBS development. Moreover, though bacteria are the richest and most well-investigated intestinal microorganisms, dysbiosis is also found in viral or fungal compositions in IBS patients. Multiomics analyses indicated specific IBS subset-related changes in phage populations [19], while mycobiome analysis findings revealed that intestinal fungi play an important role in the pathogenesis of IBS [60]. Observations from these different viewpoints profoundly indicate the complexity of enteric microbiology and encountered difficulties in microbiology research.
Epidemiological evidence showed that psychiatric and GI functional disorders are frequently complicated [61,62]. Individuals suffering from anxiety or depression have a significantly high prevalence of IBS, while IBS patients have a threefold increased risk of anxiety or depression as compared with that of healthy controls [61,62,63]. Some drugs developed primarily for psychiatric disorders can also be effective in IBS patients, and change the gut microbial composition [64,65,66]. In addition, microbial dysbiosis with aberrant microbial metabolites and ENS dysfunction are related to a variety of neurological and psychiatric disorders, including autism spectrum, Parkinson’s disease, Alzheimer’s disease, anxiety, and depression [23,67,68,69].
The microbiota also plays a significant role in CNS and ENS development [68,69]. Germ-free (GF) animals and rodent models treated with broad-spectrum antibiotics in early life had abnormal neurodevelopment [70]. GF mice showed increased permeability of the blood–brain barrier (BBB) with an immature phenotype of CNS microglia, which was reversed by fecal transplantation from specific pathogen-free (SPF) mice or the administration of bacteria producing short-chain fatty acids (SCFAs) [71]. Additionally, GF mice had disrupted motor activity with reduced anxiety-like behavior due to the altered expression of anxiety and synaptic-plasticity-related genes in the brain as compared to mice with a normal intestinal microbiota [72]. On the other hand, microbiota-derived molecules or metabolites can induce host-derived cytokines and inflammation in the CNS, which contribute to development of brain disorders by changing BBB permeability, brain vascular physiology, and brain structure [44]. Increased numbers of Akkermansia muciniphila and Acinetobacter calcoaceticus, and a decrease in Parabacteroides distasonis were observed in patients with multiple sclerosis [73,74], while several dysbiosis-related conditions in Parkinson’s disease were also reported [75]. These results indicate the importance of the quality of resident microbiota to maintain homeostasis in the gut and brain.
Together, these findings show that, although it remains unclear why and how a dysfunction of the BGM axis emerges, breaking the vicious cycle of aberrant BGM interactions by modulating gut microbiota may be an effective treatment option for patients with IBS and psychiatric disorders [55,76,77]. Indeed, antibiotics and probiotics were effective for some IBS patients, with fecal microbial transplantation (FMT) currently being investigated in clinical studies [50,78,79,80]. Reports of clinical trials based on evidence obtained from systematic microbial research results are anticipated.
3. Microbiota-Mediated Serotonergic Signaling in Intestines
Enteric 5-HT plays a significant role in the BGM axis under both homeostatic and pathogenic conditions [81]. Over 95% of total body 5-HT exists in the GI tract, where it is biosynthesized with L-tryptophan, mainly in enterochromaffin (EC) cells [82,83]. GF and antibiotics-treated mice displayed significantly low levels of peripheral 5-HT [84,85], while colonization with normal gut microbes increased colonic 5-HT production [81]. Specific resident bacteria, including Streptococcus spp., Enterococcus spp., and Corynebacterium spp., directly produce 5-HT [86], while a specific enteric resident microbiota modulates host peripheral serotonin levels. Indigenous spore-forming bacteria and Clostridium ramosum promote 5-HT biosynthesis from colonic EC cells [85,87]. A. muciniphila and its extracellular vesicles promote intestinal 5-HT biosynthesis and extracellular availability through TLR2 signaling [88,89], while SadA-expressing Staphylococci promote peripheral 5-HT synthesis [90]. Another study showed that probiotic strain Escherichia coli Nissle 1917 enhanced 5-HT bioavailability in ileal tissue [91], while Lactobacillus rhamnosus exerted antidepressant effects and decreased colonic 5-HT levels in a mouse model of depression [92]. In contrast, luminal 5-HT alters the gut bacterial population. Exposure to 5-HT reduces the expression of sporulation factors and membrane transporters in Turicibacter sanguinis [93], and increased enteric 5-HT results in gut dysbiosis characterized by increased Bacilli species, and decreases in Bifidobacterium species and A. muciniphila populations [94]. Low availability of 5-HT alters the gut bacterial composition, while 5-HT both stimulated and inhibited the growth of commensal bacteria in vitro in concentration-dependent and species-specific manners [95]. These results indicate that a certain enteric resident microbiota has bidirectional communication with the host serotonergic system to promote habitation in the intestines. Together, gut resident microbiota and microbial metabolites regulate the host serotonergic pathway that includes EC cells, tryptophan hydroxylase (TPH), serotonin reuptake transporter (SERT), 5-HT receptors, and microbial quorum sensing, which is detailed below (Figure 1, Table 1 and Table 2).
- (1)
- EC cells
The density of EC cells in the gut is modulated by the microbiota or microbiome-derived products including SCFAs, with specificity related to bacterial species [85]. Indeed, the numbers of intestinal EC cells were reduced in GF animal models [96,104], while the administration of FMT with SPF feces increased 5-HT-producing EC cells and M2 macrophages in the GI tract [105]. Bacteriodes thetaiotaomicron restores 5-HT+ EC cells and shapes EC networks in the GI tract of GF mice by producing acetate, propionate, and succinate [96], and those metabolites stimulate EC cell activity and increase the availability of tryptophan [81,85]. Mucosal inflammation induced by Trichinella spiralis increases the number of EC cells along with development of postinfectious (PI)-IBS [101]. Clostridium ramosum stimulates host 5-HT secretion and programs the differentiation of colonic intestinal stem progenitors toward a secretory 5-HT-producing lineage [87]. In contrast, Bifidobacterium pseudolongum reduces the content of 5-HT in colonic mucosa by reducing EC cells [103]. These observations indicate that EC cells are regulated by specific bacteria and induction of aberrant serotonergic signaling by dysbiosis.
- (2)
- TPH
Two isoforms, TPH1 and TPH2, comprise TPH, a rate-limiting enzyme involved in the biosynthesis of 5-HT that converts L-tryptophan into 5-hydroxytryptophan (5-HTP), a direct precursor of 5-HT [106,107]. TPH1 is expressed in the peripheral tissue such as in the lungs, heart, and kidneys, and intestines, while TPH2 is primarily expressed in the serotonergic neurons of the brain and ENS [106,107,108,109]. In the intestines, TPH1 is mostly located in EC cells, and regulated by resident microbiota and their metabolites [106,107]. Genomewide association study (GWAS) results demonstrated that polymorphisms of the TPH gene are associated with the development of IBS in humans [110,111,112,113], though contradictory clinical results regarding intestinal TPH1 expression in IBS patients were also reported [113,114,115]. In a preclinical study, colonic Tph1 expression and 5-HT biosynthesis were increased by microbiota-derived SCFAs, such as butyrate and acetate [81]. On the other hand, host genetics related to the serotonergic pathway alter the gut microbial composition, as Tph1–/– mice have a different gut microbiota than that of Tph1+/+ mice, which is related to colitis susceptibility [95]. These findings indicate bilateral communication in microbiota-serotonergic pathways related to TPH regulation.
- (3)
- SERT
A member of the neurotransmitter-sodium symporter family termed SERT regulates the extracellular availability of 5-HT in the gut and brain by 5-HT uptake [116]. Gut mucosal SERT expression is regulated by multiple stimulation factors, including glucagon-like peptides, transforming growth factor beta, immune response, inflammation, growth factors, and microbiota members [116]. A genetic or environmental abnormality in SERT expression is associated with aberrant mucosal 5-HT levels, and can cause a variety of GI functional diseases including IBS [116,117]. T. spiralis and Campylobacter jejuni reduce SERT expression in the gut [100,101], while L. rhamnosus upregulated gene expression and protein levels of SERT in a rat model of PI-IBS [100], and Lactobacillus acidophilus and Bifidobacterium longum supernatants to upregulate SERT expression in HT-29 and Caco-2 cells [102]. SERT deficiency is related to dysbiosis and changes in the metabolic function of the mouse enteric microbiome [94], and female SERT–/– rats showed visceral hypersensitivity and accelerated GI motility [118]. In humans, reduced SERT expression was observed in the rectal tissue of patients with IBS [119]. GWAS results also indicated that polymorphisms of SERT are susceptible to IBS development [14,120]. On the other hand, an increase in gut luminal 5-HT level by oral supplementation with 5-HT or SERT deficiency in the host increases the relative abundance of spore-forming T. sanguinis organisms that uptake 5-HT, which can be reversed by exposure to fluoxetine, a selective serotonin reuptake inhibitor (SSRI) [93].
- (4)
- 5-HT receptors
The 5-HT receptors have a variety of biological functions in the host, such as increasing visceral hypersensitivity and mucosal permeability, inducing inflammation along with activation of immune cells, and changing gut motility [28]. These wide-ranging effects may be because of the vast localization and diversity of 5-HT receptors. So far, 14 different 5-HT receptors in 7 families (5-HT1–7) were identified [121], with 5-HT3 and 5-HT4 being the most investigated serotonin receptors in the intestine. The 5-HT3 receptors are present in sensory and myenteric neurons, while 5-HT4 receptors are located in presynaptic sites [122]. Agents targeting these receptors are commonly utilized in clinical practice for treating IBS patients [7,123]. The function and expression of 5-HT receptors are also regulated by gut resident microbiota. Normal gut microbes increase colonic 5-HT production and activate the 5-HT4 receptor [124,125], while A. muciniphila increases the gene expression of Htr4, and decreases that of the Htr2B, Htr3B, and Htr7 genes in the colon [89]. The colonization of GF mice with spore-forming bacteria also increased colonic 5-HT by the upregulation of Htr4 [85].
- (5)
- Quorum sensing (QS)
5-HT is critical for both host physiological functions and communication among gut microbiota members. Additionally, it plays a significant role in QS, as it enables bacteria to detect and control cell population density by releasing small molecules and altering their gene expression [126]. Under homeostatic conditions, 5-HT-dependent QS can be helpful in forming a steady microbial flora, and prevent pathobiont invasion and settlement, while an abnormal QS process based on the dysregulation of 5-HT signaling potentially induces a dysbiosis condition in patients with IBS. QS-regulated mediators produced by Staphylococcus aureus interact with intrinsic intestinal neurons and smooth muscle cells, and cause dysmotility in the host GI system [127], a process potentially involved in the pathology of PI-IBS [128]. Furthermore, 5-HT activates virulence factors in and biofilm formation by pathogenic bacteria through QS [129].
4. Microbiota-Mediated Serotonergic Signaling in IBS Pathology
The enteric-microbiota-mediated serotonergic pathway plays a homeostatic role in GI functions [28], while dysfunction of the pathway may contribute to shaping IBS features. We now discuss the molecular mechanisms of the microbe-mediated serotonergic system related to IBS features, including GI motility, visceral pain sensation, and mucosal inflammation with activated immune response (Figure 2).
- (1)
- Role of Gut Microbe-Mediated 5-HT Signaling in GI Motility
The role of 5-HT in GI motility was extensively investigated [130]; 5-HT stimulates peristaltic reflexes in the GI tract, resulting in ascending contractile and descending relaxant limbs [83], while it regulates segmentation motor patterns in the small intestine of guinea pigs [131]. Serotonergic neurons appear to be more important than EC cells are for the regulation of constitutive GI motility, as Tph2–/– but not Tph1–/– mice showed delayed GI motility, which is due to reductions in contractile complexes and excitatory synaptic transmission associated with low 5-HT availability [132]. In addition, Tph2–/– mice have an immature ENS, particularly dopaminergic neurons, responsible for homeostatic GI movement [132]. Mice with the SERT Ala56 mutation have hyperactive SERT function and low 5-HT availability, resulting in decreased intestinal motility shown in both in vivo and in vitro findings, which can be reversed by 5-HT4 receptor antagonists [133]. The use of the SERT antagonist fluoxetine enhances GI motility and SERT–/– mice with hyperavailability of 5-HT also show such enhanced motility [133]. Furthermore, endogenous 5-HT can function as a modulator of GI motility via activation of the 5-HT3 and 5-HT4 receptors in the ENS [134], while antagonists of those receptors was reported to cause a delay in intestinal transit and reverse corticotrophin-releasing hormone-induced defecation in rats [135,136].
Enteric resident microbiota play a key role in serotonergic pathway-mediated GI motility. Indeed, GF animals show significantly slower GI transit as compared to control animals with normal gut microbiota, while treatments with antibiotics delay GI motility along with decreased peripheral 5-HT biosynthesis [137]. Other reports showed that colonization with gut microbiota increases gut motility in ex-GF animals in conjunction with an elevated luminal 5-HT level, while administration of pharmacologic antagonists of 5-HT4 receptors resulted in recovery GI transit in GF rodents [105,124,138]. In addition, gut bacteria-derived 5-hydroxyindole, a major 5-HT metabolite, translocates through intestinal smooth muscle cells and directly accelerates colonic motility via the activation of L-type calcium channels [139]. Moreover, BTBR mice, a mouse model of ASD, showed an impaired serotonergic pathway with the downregulation of Tph1 and upregulation of Sert in the gut, which was associated with a reduction in 5-HT-producing Blautia bacteria [140]. BTBR mice also demonstrated impaired bile acid synthesis due to a decrease in bile-metabolizing Bifidobacterium and Blautia bacterial species in the gut, which causes increased mucosal permeability, delayed GI transit, and an autism-like behavioral phenotype [140]. Furthermore, 5-HT with gut microbial stimulation can increase the number of M2 macrophages adjacent to the ENS in the colonic muscular layer, known to be associated with acceleration of GI motility [105]. SCFAs, microbial metabolites, stimulate colonic transit via intraluminal 5-HT release, potentially through the GPR43 receptor on mucosal mast cells [141,142]. Together, these findings show that enteric microbial members regulate GI motility through 5-HT signaling by multiple mechanisms, while dysregulation of this system causes aberrant GI movement, potentially related to an IBS symptom.
- (2)
- Role of Gut Microbe-Mediated 5-HT Signaling in Visceral Pain Sensation
Commensal microbiota activities are critical for developing homeostatic pain sensitivity by inducing normal excitability in the gut sensory neurons, which are absent in GF animals. Indeed, while GF mice show limited mucosal inflammation, visceral hypersensitivity due to altered pain processing in the brain is evident, which can be normalized by FMT with feces from conventional mice [143,144,145]. Gut microbe components, such as certain TLR ligands, formyl peptide receptor 1 agonists, and SCFAs, can directly enhance visceral pain sensitivity by stimulating primary nociceptive neurons in dorsal root ganglia (DRG) or indirectly by activating inflammatory immune response in the gut [146,147]. On the other hand, microbe-mediated kynurenic acid, serine proteases, and bile acids directly reduce pain by inactivating DRG neurons or indirectly by releasing opioid-like factors from mucosal immune cells [146].
The gut microbiota also plays a significant role in development of chronic abdominal pain, an essential feature of IBS [1,2,147,148]. Fecal microbiota samples obtained from IBS patients transmitted hypersensitivity to colonic distension in rats, indicating that gut microbial components are responsible for abnormal pain sensation associated with IBS [149]. In addition, gut-microbiota-mediated neurotransmitters play a significant role in visceral pain sensation [18,22]. Among those neurotransmitters, peripheral 5-HT appears to preferentially act to induce pain by stimulating mesenteric sensory fibers, and vagal and spinal afferent fibers [136]. The release of enteric 5-HT is associated with the severity of abdominal pain in IBS patients, though the effect of 5-HT is dependent on type of 5-HT receptors activated [82,121]. The 5-HT3 receptor located in vagal afferent nerve endings in the gut and peripheral endings in spinal afferent nerves plays an important role in visceral hypersensitivity and the nociceptive process as part of the pathogenesis of IBS [29,150]. The central terminals of vagal afferents also exhibit the 5-HT3 receptor, which modulates other neurotransmitters in the brain [147,150]. As a result, the activation of that receptor promotes the excitability and activity of GI vagal afferents, while ramosetron, an antagonist of the 5-HT3 receptor, effectively reduces visceral hypersensitivity and modulates GI transit in IBS-D patients [7,150,151]. Moreover, microbiota-mediated mucosal inflammation and increased permeability in the gut is a trigger of pain through the 5-HT pathway, while 5-HT3 receptor antagonists have an anti-inflammatory role [152]. Increased mucosal permeability in PI-IBS promotes intestinal mechanosensitivity, which has an effect on visceral sensitivity. Indeed, 5-HT metabolism dysfunction in IBS patients is associated with deterioration of intestinal barrier function [152,153,154]. Together, these results indicate that the gut microbiota is deeply involved in 5-HT-mediated pain sensations, while a dysfunction of the serotonergic pathway associated with enteric dysbiosis in IBS cases can cause visceral hypersensitivity.
- (3)
- Role of Gut-Microbe-Mediated 5-HT Signaling in Mucosal Inflammation and Immune Response
The 5-HT promotes intestinal inflammation through various 5-HT receptors on a variety of mucosal immune cells [155,156]. Indeed, increased mucosal 5-HT and EC cell numbers have been observed in association with several inflammatory conditions including IBD. In animal models of colitis, 5-HT activates peritoneal macrophages and also splenic dendritic cells to produce proinflammatory cytokines in a nuclear factor kappa B-dependent manner, along with sequential T cell activation [157,158]. Tph1–/– mice with low 5-HT availability are tolerant to experimental colitis [95], while SERT-deficient animals with elevated 5-HT availability were susceptible to gut mucosal inflammation [159]. The pharmacological inhibition of enteric 5-HT by peripheral TPH inhibitors alleviates inflammation [160]. These findings indicate that enteric 5-HT plays a proinflammatory role in intestinal inflammation.
Persistent low-grade mucosal inflammation with aberrant immune cell activation after development of severe enterocolitis is one of the mechanisms underlying the pathogenesis of IBS, especially PI-IBS [161,162,163]. A massive infiltration of mast cells in colonic mucosa associated with the elevated availability of mucosal 5-HT was observed in IBS patients and is correlated with abdominal pain [164]. Although it remains unknown why mucosal inflammation persists even after elimination of pathobionts, the dysbiosis of gut resident microbiota and altered serotonergic signaling are key factors for PI-IBS development. For example, helminth T. spiralis induced PI-IBS in mice, along with increased luminal 5-HT level and numbers of 5-HT+ EC cells, and reduced SERT expression in the gut [165]. Chemically induced colitis promotes visceral hypersensitivity by increasing the number of 5-HT3 receptors expressing nerve fibers in the GI tract [166]. Hypersensitivity to 5-HT in colonic serosal and mesenteric endings remains after recovery from colitis because of persistent alterations in the dynamics of enteric 5-HT3 receptors and mast cells [167]. Obtained findings with a postinflammatory IBS rat model showed visceral hypersensitivity accompanied by fecal microbial dysbiosis, elevated serum 5-HT level, the upregulated expression of colonic 5-HT3A/5-HT2B receptors, and impaired tight-junction protein expression, while the administration of a 5-HT3A receptor antagonist or FMT from the feces of normal healthy rats alleviated the IBS-like symptoms [168]. In addition, microbiota-derived SCFAs play an important role in maintaining gut homeostasis, and appear to serve a dual role in GI mucosal immunity and inflammation [169]. They play an anti-inflammatory role by strengthening epithelial-barrier integrity through the upregulation of G-protein coupled receptors in the gut, and inducing and maintaining regulatory T cells [170,171]. On the other hand, SCFAs can induce mucosal inflammation through the upregulation of TPH1 transcription and the promotion of mucosal 5-HT production in the serotonergic pathway [81]. Together, these findings show that gut microbial dysbiosis is associated with persistent low-grade mucosal inflammation accompanied by aberrant serotonergic signaling in IBS.
5. Microbiota Mediation of Serotonergic Signaling Outside GI Tract
The role of 5-HT was initially investigated in regard to brain homeostasis and psychiatric disorders [172,173,174]. Central 5-HT influences various CNS-related activities, such as nociception, sleep, sexual behavior, cognition, reward, learning, memory, emesis, motor tone, and body temperature in a homeostatic condition [172,173,174]. Furthermore, certain populations of patients with psychiatric diseases, including depression and anxiety, demonstrate aberrant serotonergic signaling in the brain and are treated by drugs targeting 5-HT pathways [175,176]. On the other hand, accumulating evidence indicates that enteric microbiota and its metabolites are essential in maintaining brain homeostasis through the serotonergic pathway, while gut dysbiosis is deeply associated with CNS disorders through the dysregulation of 5-HT signaling in the BGM axis [44,177,178]. Manipulation of the gut microbiota can be effective in treating CNS diseases that regulate 5-HT level as one of the mechanisms [179]. Since IBS and neurological diseases are often complicated, and treatment targeting 5-HT signaling can be effective for both, a dysfunction of the same pathway related to serotonergic signaling could be present in affected patients [180,181].
In the most recent decade, multiple preclinical and clinical studies showed new functions of gut-derived 5-HT in metabolic conditions, such as regarding glucose homeostasis, lipid metabolism, and bone density, and metabolic diseases including obesity and Type 2 diabetes [134]. T. sanguinis with a SERT-like structure decreases serum triglyceride level and alters gene expression regarding lipid metabolism, which can be blocked by the use of the SSRI fluoxetine [93]. Clostridium ramosum also altered gene expressions related to lipid transport and storage function in mice fed a high-fat diet by increasing Tph1 expression, 5-HT level, and EC cell number in the gut [87]. Studies that employed pharmacological inhibition or genetic depletion of TPH1 also demonstrated control of glucose homeostasis by enteric microbiota members through regulation of EC cell 5-HT synthesis [182]. A population-based cross-sectional study also indicated that IBS is positively related to metabolic syndrome [183], while integrated and longitudinal multiomics analysis results revealed purine metabolism as a specific host-microbial metabolic pathway in IBS patents [20]. These findings show that an aberrant metabolic pathway is involved in the pathology of IBS, in which certain gut-microbiota components influence host metabolism in a 5-HT-dependant manner. The details of 5-HT-mediated metabolic changes in the pathogenesis of IBS require clarification in future investigations.
6. Treatment of IBS by Modulating Microbiota-Mediated Serotonergic Pathways
Pharmacological agents targeting serotonergic pathways, including 5-HT3 agonists, 5-HT4 antagonists, SSRI, tricyclic antidepressants, and serotonin-norepinephrine reuptake inhibitors, are commonly used to treat IBS patients in clinical practice [7,8,123,181]. Additionally, intervention of the gut microbiota by use of probiotics, prebiotics, synbiotics, elimination diets, and nonsystemic antibiotics, and FMT can be effective in certain IBS cases [6,49,50,51,55,58,184]. However, details regarding microbe-based serotonergic targeting treatment remain largely unknown. An important question is whether specific antibiotics or phage therapy targeting select microbiota can be used to regulate 5-HT and/or its downstream pathway. In terms of FMT, which fecal components can be used to modulate the 5-HT pathway in IBS patients? Is it possible to develop new biomarkers that can efficiently reveal an aberrant 5-HT pathway in IBS patients? Answers to these key questions could shed light on new directions for preclinical research and clinical practice.
7. Conclusions
Accumulating evidence indicates that aberrant 5-HT pathways associated with an altered enteric microbiota are deeply involved in the pathogenesis of IBS. However, results remain insufficient to elucidate the whole features of IBS. Particularly important in regard to IBS pathology, it has not been determined whether abnormal serotonergic signaling provides a causal effect related to dysbiosis or is a consequence of an alternating microbial community. Novel findings based on mechanistic studies and large-scale clinical trials regarding microbe-mediated serotonergic pathway are required to fully uncover the pathogenesis of IBS, which could be helpful to provide better treatment for IBS patients.
Author Contributions
Conceptualization, Y.M.; writing—original draft preparation, Y.M.; supervision, S.I. Both authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Enck, P.; Aziz, Q.; Barbara, G.; Farmer, A.D.; Fukudo, S.; Mayer, E.A.; Niesler, B.; Quigley, E.M.M.; Rajilić-Stojanović, M.; Schemann, M.; et al. Irritable bowel syndrome. Nat. Rev. Dis. Prim. 2016, 2, 16014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, A.C.; Lacy, B.E.; Talley, N.J. Irritable bowel syndrome. N. Engl. J. Med. 2017, 376, 2566–2578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperber, A.D.; Dumitrascu, D.; Fukudo, S.; Gerson, C.; Ghoshal, U.C.; Gwee, K.A.; Hungin, A.P.S.; Kang, J.-Y.; Minhu, C.; Schmulson, M.; et al. The global prevalence of IBS in adults remains elusive due to the heterogeneity of studies: A Rome foundation working team literature review. Gut 2017, 66, 1075–1082. [Google Scholar] [CrossRef] [Green Version]
- Schmulson, M.J.; Drossman, D.A. What is new in Rome IV. J. Neurogastroenterol. Motil. 2017, 23, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Drossman, D.A.; Hasler, W.L. Rome IV-functional GI disorders: Disorders of gut-brain interaction. Gastroenterology 2016, 150, 1257–1261. [Google Scholar] [CrossRef] [PubMed]
- Ford, A.C.; Harris, L.A.; Lacy, B.E.; Quigley, E.M.M.; Moayyedi, P. Systematic review with meta-analysis: The efficacy of prebiotics, probiotics, synbiotics and antibiotics in irritable bowel syndrome. Aliment. Pharmacol. Ther. 2018, 48, 1044–1060. [Google Scholar] [CrossRef] [Green Version]
- Camilleri, M.; Boeckxstaens, G. Dietary and pharmacological treatment of abdominal pain in IBS. Gut 2017, 66, 966–974. [Google Scholar] [CrossRef] [PubMed]
- Simrén, M.; Tack, J. New treatments and therapeutic targets for IBS and other functional bowel disorders. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 589–605. [Google Scholar] [CrossRef]
- Staller, K.; Olén, O.; Söderling, J.; Roelstraete, B.; Törnblom, H.; Khalili, H.; Joshi, A.D.; Nguyen, L.H.; Song, M.; Kuo, B.; et al. Mortality risk in irritable bowel syndrome: Results from a nationwide prospective cohort study. Am. J. Gastroenterol. 2020, 115, 746–755. [Google Scholar] [CrossRef]
- Mönnikes, H. Quality of life in patients with irritable bowel syndrome. J. Clin. Gastroenterol. 2011, 45, S98–S101. [Google Scholar] [CrossRef]
- Levy, R.L.; Von Korff, M.; Whitehead, W.E.; Stang, P.; Saunders, K.; Jhingran, P.; Barghout, V.; Feld, A.D. Costs of care for irritable bowel syndrome patients in a health maintenance organization. Am. J. Gastroenterol. 2001, 96, 3122–3129. [Google Scholar] [CrossRef]
- Doshi, J.A.; Cai, Q.; Buono, J.L.; Spalding, W.M.; Sarocco, P.; Tan, H.; Stephenson, J.J.; Carson, R.T. Economic burden of irritable bowel syndrome with constipation: A retrospective analysis of health care costs in a commercially insured population. J. Manag. Care Spec. Pharm. 2014, 20, 382–390. [Google Scholar] [CrossRef]
- Black, C.J.; Ford, A.C. Global burden of irritable bowel syndrome: Trends, predictions and risk factors. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.-Y.; Fang, X.-C.; Li, X.-Q.; Fei, G.-J. Ethnic differences in genetic polymorphism associated with irritable bowel syndrome. World J. Gastroenterol. 2020, 26, 2049–2063. [Google Scholar] [CrossRef]
- Ford, A.C.; Sperber, A.D.; Corsetti, M.; Camilleri, M. Irritable bowel syndrome. Lancet 2020, 396, 1675–1688. [Google Scholar] [CrossRef]
- Holtmann, G.J.; Ford, A.C.; Talley, N.J. Pathophysiology of irritable bowel syndrome. Lancet Gastroenterol. Hepatol. 2016, 1, 133–146. [Google Scholar] [CrossRef]
- Barbara, G.; Grover, M.; Bercik, P.; Corsetti, M.; Ghoshal, U.C.; Ohman, L.; Rajilić-Stojanović, M. Rome foundation working team report on post-infection irritable bowel syndrome. Gastroenterology 2019, 156, 46–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gros, M.; Gros, B.; Mesonero, J.E.; Latorre, E. Neurotransmitter dysfunction in irritable bowel syndrome: Emerging approaches for management. J. Clin. Med. 2021, 10, 3429. [Google Scholar] [CrossRef] [PubMed]
- Mihindukulasuriya, K.A.; Mars, R.A.T.; Johnson, A.J.; Ward, T.; Priya, S.; Lekatz, H.R.; Kalari, K.R.; Droit, L.; Zheng, T.; Blekhman, R.; et al. Multi-omics analyses show disease, diet, and transcriptome interactions with the virome. Gastroenterology 2021. [Google Scholar] [CrossRef]
- Mars, R.A.T.; Yang, Y.; Ward, T.; Houtti, M.; Priya, S.; Lekatz, H.R.; Tang, X.; Sun, Z.; Kalari, K.R.; Korem, T.; et al. Longitudinal multi-omics reveals subset-specific mechanisms underlying irritable bowel syndrome. Cell 2020, 182, 1460–1473. [Google Scholar] [CrossRef] [PubMed]
- Putignani, L.; Del Chierico, F.; Vernocchi, P.; Cicala, M.; Cucchiara, S.; Dallapiccola, B. Gut Microbiota Dysbiosis as risk and premorbid factors of IBD and IBS along the childhood-adulthood transition. Inflamm. Bowel Dis. 2016, 22, 487–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishima, Y.; Ishihara, S. Molecular mechanisms of microbiota-mediated pathology in irritable bowel syndrome. Int. J. Mol. Sci. 2020, 21, 8664. [Google Scholar] [CrossRef]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The microbiota-gut-brain axis: From motility to mood. Gastroenterology 2021, 160, 1486–1501. [Google Scholar] [CrossRef] [PubMed]
- Strandwitz, P. Neurotransmitter modulation by the gut microbiota. Brain Res. 2018, 1693, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Hellström, P.M. Pathophysiology of the irritable bowel syndrome—Reflections of today. Best Pract. Res. Clin. Gastroenterol. 2019, 40, 101620. [Google Scholar] [CrossRef] [PubMed]
- Mohammad-Zadeh, L.F.; Moses, L.; Gwaltney-Brant, S.M. Serotonin: A review. J. Vet. Pharmacol. Ther. 2008, 31, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Bruta, K.; Vanshika; Bhasin, K.; Bhawana. The role of serotonin and diet in the prevalence of irritable bowel syndrome: A systematic review. Transl. Med. Commun. 2021, 6, 1. [Google Scholar] [CrossRef]
- Crowell, M.D. Role of serotonin in the pathophysiology of the irritable bowel syndrome. Br. J. Pharmacol. 2004, 141, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Ford, A.C.; Brandt, L.J.; Young, C.; Chey, W.D.; Foxx-Orenstein, A.E.; Moayyedi, P. Efficacy of 5-HT3 antagonists and 5-HT4 agonists in irritable bowel syndrome: Systematic review and meta-analysis. Am. J. Gastroenterol. 2009, 104, 1831–1843. [Google Scholar] [CrossRef]
- Chang, L.; Di Lorenzo, C.; Farrugia, G.; Hamilton, F.A.; Mawe, G.M.; Pasricha, P.J.; Wiley, J.W. Functional bowel disorders: A roadmap to guide the next generation of research. Gastroenterology 2018, 154, 723–735. [Google Scholar] [CrossRef]
- Qin, H.-Y.; Cheng, C.-W.; Tang, X.-D.; Bian, Z.-X. Impact of psychological stress on irritable bowel syndrome. World J. Gastroenterol. 2014, 20, 14126–14131. [Google Scholar] [CrossRef]
- Barbara, G. IBS: Biomarkers for IBS: Ready for prime time? Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 9–10. [Google Scholar] [CrossRef]
- Vannucchi, M.G.; Evangelista, S. Experimental models of irritable bowel syndrome and the role of the enteric neurotransmission. J. Clin. Med. 2018, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Qin, H.-Y.; Wu, J.C.Y.; Tong, X.-D.; Sung, J.J.Y.; Xu, H.-X.; Bian, Z.-X. Systematic review of animal models of post-infectious/post-inflammatory irritable bowel syndrome. J. Gastroenterol. 2011, 46, 164–174. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The human intestinal microbiome in health and disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominguez-Bello, M.G.; Godoy-Vitorino, F.; Knight, R.; Blaser, M.J. Role of the microbiome in human development. Gut 2019, 68, 1108–1114. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.A.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.K.; Lynch, S.V.; Knight, R. Current understanding of the human microbiome. Nat. Med. 2018, 24, 392–400. [Google Scholar] [CrossRef]
- Chang, C.; Lin, H. Dysbiosis in gastrointestinal disorders. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 3–15. [Google Scholar] [CrossRef]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef] [PubMed]
- Simpson, C.A.; Mu, A.; Haslam, N.; Schwartz, O.S.; Simmons, J.G. Feeling down? A systematic review of the gut microbiota in anxiety/depression and irritable bowel syndrome. J. Affect. Disord. 2020, 266, 429–446. [Google Scholar] [CrossRef]
- Amato, K.R.; Arrieta, M.-C.; Azad, M.B.; Bailey, M.T.; Broussard, J.L.; Bruggeling, C.E.; Claud, E.C.; Costello, E.K.; Davenport, E.R.; Dutilh, B.E.; et al. The human gut microbiome and health inequities. Proc. Natl. Acad. Sci. USA 2021, 118, e2017947118. [Google Scholar] [CrossRef]
- Integrative HMP (iHMP); Proctor, L.M.; Creasy, H.H.; Fettweis, J.M.; Lloyd-Price, J.; Mahurkar, A.; Zhou, W.; Buck, G.A.; Snyder, M.P.; Strauss, J.F., III; et al. Research network consortium the integrative human microbiome project. Nature 2019, 569, 641–648. [Google Scholar] [CrossRef] [Green Version]
- Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The healthy human microbiome. Genome Med. 2016, 8, 51. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Jiang, Y.; Xu, K.; Cui, M.; Ye, W.; Zhao, G.; Jin, L.; Chen, X. The progress of gut microbiome research related to brain disorders. J. Neuroinflamm. 2020, 17, 25. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.R.; Osadchiy, V.; Kalani, A.; Mayer, E.A. The brain-gut-microbiome axis. Cell. Mol. Gastroenterol. Hepatol. 2018, 6, 133–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osadchiy, V.; Martin, C.R.; Mayer, E.A. The gut-brain axis and the microbiome: Mechanisms and clinical implications. Clin. Gastroenterol. Hepatol. 2019, 17, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Chey, W.; Menees, S. The gut microbiome and irritable bowel syndrome. F1000Research 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Chong, P.P.; Chin, V.K.; Looi, C.Y.; Wong, W.F.; Madhavan, P.; Yong, V.C. The microbiome and irritable bowel syndrome—A review on the pathophysiology, current research and future therapy. Front. Microbiol. 2019, 10, 1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salem, A.E.; Singh, R.; Ayoub, Y.K.; Khairy, A.M.; Mullin, G.E. The gut microbiome and irritable bowel syndrome: State of art review. Arab J. Gastroenterol. 2018, 19, 136–141. [Google Scholar] [CrossRef]
- Staudacher, H.M.; Whelan, K. Altered gastrointestinal microbiota in irritable bowel syndrome and its modification by diet: Probiotics, prebiotics and the low FODMAP diet. In Proceedings of the Nutrition Society; Cambridge University Press: Cambridge, UK, 2016; Volume 75, pp. 306–318. [Google Scholar]
- Enck, P.; Mazurak, N. Microbiota and irritable bowel syndrome: A critical inventory. Z. Gastroenterol. 2019, 57, 859–870. [Google Scholar] [CrossRef]
- Collins, S.M. A role for the gut microbiota in IBS. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Drago, L.; Valentina, C.; Fabio, P. Gut microbiota, dysbiosis and colon lavage. Dig. Liver Dis. 2019, 51, 1209–1213. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Xiong, L.; Li, L.; Li, M.; Chen, M. Alterations of gut microbiota in patients with irritable bowel syndrome: A systematic review and meta-analysis. J. Gastroenterol. Hepatol. 2017, 32, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Rodiño-Janeiro, B.K.; Vicario, M.; Alonso-Cotoner, C.; Pascua-García, R.; Santos, J. A review of microbiota and irritable bowel syndrome: Future in therapies. Adv. Ther. 2018, 35, 289–310. [Google Scholar] [CrossRef] [Green Version]
- Duan, R.; Zhu, S.; Wang, B.; Duan, L. Alterations of gut microbiota in patients with irritable bowel syndrome based on 16S rRNA-targeted sequencing. Clin. Transl. Gastroenterol. 2019, 10, e00012. [Google Scholar] [CrossRef]
- Wang, L.; Alammar, N.; Singh, R.; Nanavati, J.; Song, Y.; Chaudhary, R.; Mullin, G.E. Gut microbial dysbiosis in the irritable bowel syndrome: A systematic review and meta-analysis of case-control studies. J. Acad. Nutr. Diet. 2020, 120, 565–586. [Google Scholar] [CrossRef] [Green Version]
- Pittayanon, R.; Lau, J.T.; Yuan, Y.; Leontiadis, G.I.; Tse, F.; Surette, M.; Moayyedi, P. Gut microbiota in patients with irritable bowel syndrome-A systematic review. Gastroenterology 2019, 157, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Baumgartner, M.; Lang, M.; Holley, H.; Crepaz, D.; Hausmann, B.; Pjevac, P.; Moser, D.; Haller, F.; Hof, F.; Beer, A.; et al. Mucosal biofilms are an endoscopic feature of irritable bowel syndrome and ulcerative colitis. Gastroenterology 2021. [Google Scholar] [CrossRef]
- Gu, Y.; Zhou, G.; Qin, X.; Huang, S.; Wang, B.; Cao, H. The potential role of gut mycobiome in irritable bowel syndrome. Front. Microbiol. 2019, 10, 1894. [Google Scholar] [CrossRef]
- Zamani, M.; Alizadeh-Tabari, S.; Zamani, V. Systematic review with meta-analysis: The prevalence of anxiety and depression in patients with irritable bowel syndrome. Aliment. Pharmacol. Ther. 2019, 50, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Koloski, N.A.; Jones, M.; Talley, N.J. Evidence that independent gut-to-brain and brain-to-gut pathways operate in the irritable bowel syndrome and functional dyspepsia: A 1-year population-based prospective study. Aliment. Pharmacol. Ther. 2016, 44, 592–600. [Google Scholar] [CrossRef] [PubMed]
- Sibelli, A.; Chalder, T.; Everitt, H.; Workman, P.; Windgassen, S.; Moss-Morris, R. A systematic review with meta-analysis of the role of anxiety and depression in irritable bowel syndrome onset. Psychol. Med. 2016, 46, 3065–3080. [Google Scholar] [CrossRef] [PubMed]
- Vich Vila, A.; Collij, V.; Sanna, S.; Sinha, T.; Imhann, F.; Bourgonje, A.R.; Mujagic, Z.; Jonkers, D.M.A.E.; Masclee, A.A.M.; Fu, J.; et al. Impact of commonly used drugs on the composition and metabolic function of the gut microbiota. Nat. Commun. 2020, 11, 362. [Google Scholar] [CrossRef]
- Lukić, I.; Getselter, D.; Ziv, O.; Oron, O.; Reuveni, E.; Koren, O.; Elliott, E. Antidepressants affect gut microbiota and Ruminococcus flavefaciens is able to abolish their effects on depressive-like behavior. Transl. Psychiatry 2019, 9, 133. [Google Scholar] [CrossRef] [PubMed]
- Kułak-Bejda, A.; Bejda, G.; Waszkiewicz, N. Antidepressants for irritable bowel syndrome—A systematic review. Pharmacol. Rep. 2017, 69, 1366–1379. [Google Scholar] [CrossRef] [PubMed]
- Obata, Y.; Pachnis, V. The effect of microbiota and the immune system on the development and Organization of the Enteric Nervous System. Gastroenterology 2016, 151, 836–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.; Xing, C.; Long, W.; Wang, H.Y.; Liu, Q.; Wang, R.-F. Impact of microbiota on central nervous system and neurological diseases: The gut-brain axis. J. Neuroinflamm. 2019, 16, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiss, C.N.; Olofsson, L.E. The role of the gut microbiota in development, function and disorders of the central nervous system and the enteric nervous system. J. Neuroendocrinol. 2019, 31, e12684. [Google Scholar] [CrossRef]
- Spichak, S.; Guzzetta, K.E.; O’Leary, O.F.; Clarke, G.; Dinan, T.G.; Cryan, J.F. Without a bug’s life: Germ-free rodents to interrogate microbiota-gut-neuroimmune interactions. Drug Discov. Today Dis. Model. 2018, 28, 79–93. [Google Scholar] [CrossRef]
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Toth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Kundu, P.; et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 2014, 6, 263ra158. [Google Scholar] [CrossRef] [Green Version]
- Heijtz, R.D.; Wang, S.; Anuar, F.; Qian, Y.; Bjorkholm, B.; Samuelsson, A.; Hibberd, M.L.; Forssberg, H.; Pettersson, S. Normal gut microbiota modulates brain development and behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 3047–3052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boziki, M.K.; Kesidou, E.; Theotokis, P.; Mentis, A.-F.A.; Karafoulidou, E.; Melnikov, M.; Sviridova, A.; Rogovski, V.; Boyko, A.; Grigoriadis, N. Microbiome in multiple sclerosis: Where are we, what we know and do not know. Brain Sci. 2020, 10, 234. [Google Scholar] [CrossRef] [Green Version]
- Cekanaviciute, E.; Yoo, B.B.; Runia, T.F.; Debelius, J.W.; Singh, S.; Nelson, C.A.; Kanner, R.; Bencosme, Y.; Lee, Y.K.; Hauser, S.L.; et al. Gut bacteria from multiple sclerosis patients modulate human T cells and exacerbate symptoms in mouse models. Proc. Natl. Acad. Sci. USA 2017, 114, 10713–10718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Liao, J.; Liu, X.; Zhong, Y.; Cai, X.; Long, L. Review: The role of intestinal dysbiosis in Parkinson’s disease. Front. Cell. Infect. Microbiol. 2021, 11, 615075. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Kazmi, S.A.; Jameson, K.G.; Hsiao, E.Y. The microbiome as a modifier of neurodegenerative disease risk. Cell Host Microbe 2020, 28, 201–222. [Google Scholar] [CrossRef]
- Sasmita, A.O. Modification of the gut microbiome to combat neurodegeneration. Rev. Neurosci. 2019, 30, 795–805. [Google Scholar] [CrossRef]
- König, J.; Brummer, R.J. Faecal microbiota transplantation in IBS—New evidence for success? Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 199–200. [Google Scholar] [CrossRef] [PubMed]
- Lacy, B.E.; Pimentel, M.; Brenner, D.M.; Chey, W.D.; Keefer, L.A.; Long, M.D.; Moshiree, B. ACG Clinical guideline: Management of irritable bowel syndrome. Am. J. Gastroenterol. 2021, 116, 17–44. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, M.; Lembo, A.; Chey, W.D.; Zakko, S.; Ringel, Y.; Yu, J.; Mareya, S.M.; Shaw, A.L.; Bortey, E.; Forbes, W.P.; et al. Rifaximin therapy for patients with irritable bowel syndrome without constipation. N. Engl. J. Med. 2011, 364, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Reigstad, C.S.; Salmonson, C.E.; Rainey, J.F., III; Szurszewski, J.H.; Linden, D.R.; Sonnenburg, J.L.; Farrugia, G.; Kashyap, P.C.; Rainey, J.F.; Szurszewski, J.H.; et al. Gut microbes promote colonic serotonin production through an effect of short-chain fatty acids on enterochromaffin cells. FASEB J. 2015, 29, 1395–1403. [Google Scholar] [CrossRef] [Green Version]
- Cremon, C.; Carini, G.; Wang, B.; Vasina, V.; Cogliandro, R.F.; De Giorgio, R.; Stanghellini, V.; Grundy, D.; Tonini, M.; De Ponti, F.; et al. Intestinal serotonin release, sensory neuron activation, and abdominal pain in irritable bowel syndrome. Am. J. Gastroenterol. 2011, 106, 1290–1298. [Google Scholar] [CrossRef]
- Gershon, M.D. 5-Hydroxytryptamine (serotonin) in the gastrointestinal tract. Curr. Opin. Endocrinol. Diabetes Obes. 2013, 20, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Sjögren, K.; Engdahl, C.; Henning, P.; Lerner, U.H.; Tremaroli, V.; Lagerquist, M.K.; Bäckhed, F.; Ohlsson, C. The gut microbiota regulates bone mass in mice. J. Bone Miner. Res. 2012, 27, 1357–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roshchina, V.V. Evolutionary considerations of neurotransmitters in microbial, plant, and animal cells. In Microbial Endocrinology; Springer: New York, NY, USA, 2010; pp. 17–52. [Google Scholar]
- Mandić, A.D.; Woting, A.; Jaenicke, T.; Sander, A.; Sabrowski, W.; Rolle-Kampcyk, U.; von Bergen, M.; Blaut, M. Clostridium ramosum regulates enterochromaffin cell development and serotonin release. Sci. Rep. 2019, 9, 1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Xu, W.; Wang, R.; Cheng, R.; Tang, Z.; Zhang, M. The outer membrane protein Amuc_1100 of Akkermansia muciniphila promotes intestinal 5-HT biosynthesis and extracellular availability through TLR2 signalling. Food Funct. 2021, 12, 3597–3610. [Google Scholar] [CrossRef] [PubMed]
- Yaghoubfar, R.; Behrouzi, A.; Ashrafian, F.; Shahryari, A.; Moradi, H.R.; Choopani, S.; Hadifar, S.; Vaziri, F.; Nojoumi, S.A.; Fateh, A.; et al. Modulation of serotonin signaling/metabolism by Akkermansia muciniphila and its extracellular vesicles through the gut-brain axis in mice. Sci. Rep. 2020, 10, 22119. [Google Scholar] [CrossRef]
- Luqman, A.; Nega, M.; Nguyen, M.-T.; Ebner, P.; Götz, F. SadA-Expressing Staphylococci in the human gut show increased cell adherence and internalization. Cell Rep. 2018, 22, 535–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nzakizwanayo, J.; Dedi, C.; Standen, G.; Macfarlane, W.M.; Patel, B.A.; Jones, B.V. Escherichia coli Nissle 1917 enhances bioavailability of serotonin in gut tissues through modulation of synthesis and clearance. Sci. Rep. 2015, 5, 17324. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, P.; Huang, L.; Li, P.; Zhang, D. Effects of regulating gut microbiota on the serotonin metabolism in the chronic unpredictable mild stress rat model. Neurogastroenterol. Motil. 2019, 31, e13677. [Google Scholar] [CrossRef] [Green Version]
- Fung, T.C.; Vuong, H.E.; Luna, C.D.G.; Pronovost, G.N.; Aleksandrova, A.A.; Riley, N.G.; Vavilina, A.; McGinn, J.; Rendon, T.; Forrest, L.R.; et al. Intestinal serotonin and fluoxetine exposure modulate bacterial colonization in the gut. Nat. Microbiol. 2019, 4, 2064–2073. [Google Scholar] [CrossRef]
- Singhal, M.; Turturice, B.A.; Manzella, C.R.; Ranjan, R.; Metwally, A.A.; Theorell, J.; Huang, Y.; Alrefai, W.A.; Dudeja, P.K.; Finn, P.W.; et al. Serotonin transporter deficiency is associated with dysbiosis and changes in metabolic function of the mouse intestinal microbiome. Sci. Rep. 2019, 9, 2138. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.H.; Wang, H.; Denou, E.; Ghia, J.-E.; Rossi, L.; Fontes, M.E.; Bernier, S.P.; Shajib, M.S.; Banskota, S.; Collins, S.M.; et al. Modulation of gut microbiota composition by serotonin signaling influences intestinal immune response and susceptibility to colitis. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 709–728. [Google Scholar] [CrossRef] [Green Version]
- Modasia, A.; Parker, A.; Jones, E.; Stentz, R.; Brion, A.; Goldson, A.; Defernez, M.; Wileman, T.; Ashley Blackshaw, L.; Carding, S.R. Regulation of enteroendocrine cell networks by the major human gut symbiont Bacteroides thetaiotaomicron. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Engevik, M.A.; Luck, B.; Visuthranukul, C.; Ihekweazu, F.D.; Engevik, A.C.; Shi, Z.; Danhof, H.A.; Chang-Graham, A.L.; Hall, A.; Endres, B.T.; et al. Human-derived Bifidobacterium dentium modulates the mammalian serotonergic system and gut–brain axis. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 221–248. [Google Scholar] [CrossRef] [PubMed]
- Ranuh, R.; Athiyyah, A.F.; Darma, A.; Risky, V.P.; Riawan, W.; Surono, I.S.; Sudarmo, S.M. Effect of the probiotic Lactobacillus plantarum IS-10506 on BDNF and 5HT stimulation: Role of intestinal microbiota on the gut-brain axis. Iran J. Microbiol. 2019, 11, 145–150. [Google Scholar] [CrossRef]
- Chen, C.-M.; Wu, C.-C.; Huang, C.-L.; Chang, M.-Y.; Cheng, S.-H.; Lin, C.-T.; Tsai, Y.-C. Lactobacillus plantarum PS128 promotes intestinal motility, mucin production, and serotonin signaling in mice. Probiotics Antimicrob. Proteins 2021, 1–11. [Google Scholar] [CrossRef]
- Cao, Y.-N.; Feng, L.-J.; Liu, Y.-Y.; Jiang, K.; Zhang, M.-J.; Gu, Y.-X.; Wang, B.-M.; Gao, J.; Wang, Z.-L.; Wang, Y.-M. Effect of Lactobacillus rhamnosus GG supernatant on serotonin transporter expression in rats with post-infectious irritable bowel syndrome. World J. Gastroenterol. 2018, 24, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Wheatcroft, J.; Wakelin, D.; Smith, A.; Mahoney, C.R.; Mawe, G.; Spiller, R. Enterochromaffin cell hyperplasia and decreased serotonin transporter in a mouse model of postinfectious bowel dysfunction. Neurogastroenterol. Motil. 2005, 17, 863–870. [Google Scholar] [CrossRef]
- Cao, Y.-N.; Feng, L.-J.; Wang, B.-M.; Jiang, K.; Li, S.; Xu, X.; Wang, W.-Q.; Zhao, J.-W.; Wang, Y.-M. Lactobacillus acidophilus and Bifidobacterium longum supernatants upregulate the serotonin transporter expression in intestinal epithelial cells. Saudi J. Gastroenterol. 2018, 24, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Tatsuoka, M.; Osaki, Y.; Ohsaka, F.; Tsuruta, T.; Kadota, Y.; Tochio, T.; Hino, S.; Morita, T.; Sonoyama, K. Consumption of indigestible saccharides and administration of Bifidobacterium pseudolongum reduce mucosal serotonin in murine colonic mucosa. Br. J. Nutr. 2021, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Uribe, A.; Alam, M.; Johansson, O.; Midtvedt, T.; Theodorsson, E. Microflora modulates endocrine cells in the gastrointestinal mucosa of the rat. Gastroenterology 1994, 107, 1259–1269. [Google Scholar] [CrossRef]
- Yang, M.; Fukui, H.; Eda, H.; Kitayama, Y.; Hara, K.; Kodani, M.; Tomita, T.; Oshima, T.; Watari, J.; Miwa, H. Involvement of gut microbiota in the association between gastrointestinal motility and 5-HT expression/M2 macrophage abundance in the gastrointestinal tract. Mol. Med. Rep. 2017, 16, 3482–3488. [Google Scholar] [CrossRef] [Green Version]
- Swami, T.; Weber, H.C. Updates on the biology of serotonin and tryptophan hydroxylase. Curr. Opin. Endocrinol. Diabetes Obes. 2018, 25, 12–21. [Google Scholar] [CrossRef]
- Jones, L.A.; Sun, E.W.; Martin, A.M.; Keating, D.J. The ever-changing roles of serotonin. Int. J. Biochem. Cell Biol. 2020, 125, 105776. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Beaulieu, J.-M.; Sotnikova, T.D.; Gainetdinov, R.R.; Caron, M.G. Tryptophan hydroxylase-2 controls brain serotonin synthesis. Science 2004, 305, 217. [Google Scholar] [CrossRef] [PubMed]
- McKinney, J.; Knappskog, P.M.; Haavik, J. Different properties of the central and peripheral forms of human tryptophan hydroxylase. J. Neurochem. 2005, 92, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.; Kohen, R.; Cain, K.C.; Jarrett, M.E.; Heitkemper, M.M. Associations of tryptophan hydroxylase gene polymorphisms with irritable bowel syndrome. Neurogastroenterol. Motil. 2011, 23, 233-e116. [Google Scholar] [CrossRef] [Green Version]
- Jun, S.-E.; Kohen, R.; Cain, K.C.; Jarrett, M.E.; Heitkemper, M.M. TPH gene polymorphisms are associated with disease perception and quality of life in women with irritable bowel syndrome. Biol. Res. Nurs. 2014, 16, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Katsumata, R.; Shiotani, A.; Murao, T.; Ishii, M.; Fujita, M.; Matsumoto, H.; Haruma, K. The TPH1 rs211105 gene polymorphism affects abdominal symptoms and quality of life of diarrhea-predominant irritable bowel syndrome. J. Clin. Biochem. Nutr. 2018, 62, 270–276. [Google Scholar] [CrossRef]
- Grasberger, H.; Chang, L.; Shih, W.; Presson, A.P.; Sayuk, G.S.; Newberry, R.D.; Karagiannides, I.; Pothoulakis, C.; Mayer, E.; Merchant, J.L. Identification of a functional TPH1 polymorphism associated with irritable bowel syndrome bowel habit subtypes. Am. J. Gastroenterol. 2013, 108, 1766–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerckhoffs, A.P.M.; ter Linde, J.J.M.; Akkermans, L.M.A.; Samsom, M. SERT and TPH-1 mRNA expression are reduced in irritable bowel syndrome patients regardless of visceral sensitivity state in large intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G1053–G1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiotani, A.; Kusunoki, H.; Ishii, M.; Imamura, H.; Manabe, N.; Kamada, T.; Hata, J.; Merchant, J.L.; Haruma, K. Pilot study of Biomarkers for predicting effectiveness of ramosetron in diarrhea-predominant irritable bowel syndrome: Expression of S100A10 and polymorphisms of TPH1. Neurogastroenterol. Motil. 2015, 27, 82–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, D.-C.; Cao, H.-L.; Xu, M.-Q.; Wang, S.-N.; Wang, Y.-M.; Yan, F.; Wang, B.-M. Regulation of the serotonin transporter in the pathogenesis of irritable bowel syndrome. World J. Gastroenterol. 2016, 22, 8137–8148. [Google Scholar] [CrossRef] [PubMed]
- Tada, Y.; Ishihara, S.; Kawashima, K.; Fukuba, N.; Sonoyama, H.; Kusunoki, R.; Oka, A.; Mishima, Y.; Oshima, N.; Moriyama, I.; et al. Downregulation of serotonin reuptake transporter gene expression in healing colonic mucosa in presence of remaining low-grade inflammation in ulcerative colitis. J. Gastroenterol. Hepatol. 2016, 31, 1443–1452. [Google Scholar] [CrossRef]
- Bi, Z.; Zhang, S.; Meng, Y.; Feng, Y.; Wang, Y.; Wang, E.; Pan, X.; Zhu, R.; Fan, H.; Pang, S.; et al. Female serotonin transporter-knockout rat: A potential model of irritable bowel syndrome. FASEB J. 2021, 35, e21701. [Google Scholar] [CrossRef]
- Coates, M.D.; Mahoney, C.R.; Linden, D.R.; Sampson, J.E.; Chen, J.; Blaszyk, H.; Crowell, M.D.; Sharkey, K.A.; Gershon, M.D.; Mawe, G.M. Molecular defects in mucosal serotonin content and decreased serotonin reuptake transporter in ulcerative colitis and irritable bowel syndrome. Gastroenterology 2004, 126, 1657–1664. [Google Scholar] [CrossRef]
- Del Colle, A.; Israelyan, N.; Gross Margolis, K. Novel aspects of enteric serotonergic signaling in health and brain-gut disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G130–G143. [Google Scholar] [CrossRef] [Green Version]
- Hoyer, D.; Martin, G. 5-HT receptor classification and nomenclature: Towards a harmonization with the human genome. Neuropharmacology 1997, 36, 419–428. [Google Scholar] [CrossRef]
- Gershon, M.D. Review article: Serotonin receptors and transporters—Roles in normal and abnormal gastrointestinal motility. Aliment. Pharmacol. Ther. 2004, 20, 3–14. [Google Scholar] [CrossRef]
- Binienda, A.; Storr, M.; Fichna, J.; Salaga, M. Efficacy and safety of serotonin receptor ligands in the treatment of irritable bowel syndrome: A review. Curr. Drug Targets 2018, 19, 1774–1781. [Google Scholar] [CrossRef]
- De Vadder, F.; Grasset, E.; Mannerås Holm, L.; Karsenty, G.; Macpherson, A.J.; Olofsson, L.E.; Bäckhed, F. Gut microbiota regulates maturation of the adult enteric nervous system via enteric serotonin networks. Proc. Natl. Acad. Sci. USA 2018, 115, 6458–6463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattarai, Y.; Williams, B.B.; Battaglioli, E.J.; Whitaker, W.R.; Till, L.; Grover, M.; Linden, D.R.; Akiba, Y.; Kandimalla, K.K.; Zachos, N.C.; et al. Gut microbiota-produced tryptamine activates an epithelial G-protein-coupled receptor to increase colonic secretion. Cell Host Microbe 2018, 23, 775–785.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, C.M.; Bassler, B.L. QUORUM SENSING: Cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlig, F.; Grundy, L.; Garcia-Caraballo, S.; Brierley, S.M.; Foster, S.J.; Grundy, D. Identification of a quorum sensing-dependent communication pathway mediating bacteria-gut-brain cross talk. iScience 2020, 23, 101695. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.-S.; He, S.-H.; Zheng, P.-Y.; Wu, L.; Yang, P.-C. Mast cells play a crucial role in Staphylococcus aureus peptidoglycan-induced diarrhea. Am. J. Pathol. 2007, 171, 537–547. [Google Scholar] [CrossRef] [Green Version]
- Knecht, L.D.; O’Connor, G.; Mittal, R.; Liu, X.Z.; Daftarian, P.; Deo, S.K.; Daunert, S. Serotonin activates bacterial quorum sensing and enhances the virulence of Pseudomonas aeruginosa in the host. EBioMedicine 2016, 9, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Terry, N.; Margolis, K.G. Serotonergic mechanisms regulating the GI tract: Experimental evidence and therapeutic relevance. Handb. Exp. Pharmacol. 2017, 239, 319–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, M.; Chambers, J.D.; Gwynne, R.M.; Bornstein, J.C. Serotonin and cholecystokinin mediate nutrient-induced segmentation in guinea pig small intestine. Am. J. Physiol. Liver Physiol. 2013, 304, G749–G761. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Chalazonitis, A.; Huang, Y.-Y.; Mann, J.J.; Margolis, K.G.; Yang, Q.M.; Kim, D.O.; Cote, F.; Mallet, J.; Gershon, M.D. Essential roles of enteric neuronal serotonin in gastrointestinal motility and the development/survival of enteric dopaminergic neurons. J. Neurosci. 2011, 31, 8998–9009. [Google Scholar] [CrossRef]
- Margolis, K.G.; Li, Z.; Stevanovic, K.; Saurman, V.; Israelyan, N.; Anderson, G.M.; Snyder, I.; Veenstra-VanderWeele, J.; Blakely, R.D.; Gershon, M.D. Serotonin transporter variant drives preventable gastrointestinal abnormalities in development and function. J. Clin. Invest. 2016, 126, 2221–2235. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.M.; Young, R.L.; Leong, L.; Rogers, G.B.; Spencer, N.J.; Jessup, C.F.; Keating, D.J. The diverse metabolic roles of peripheral serotonin. Endocrinology 2017, 158, 1049–1063. [Google Scholar] [CrossRef]
- Miyata, K.; Ito, H.; Fukudo, S. Involvement of the 5-HT3 receptor in CRH-induce defecation in rats. Am. J. Physiol. 1998, 274, G827–G831. [Google Scholar] [CrossRef]
- Mawe, G.M.; Hoffman, J.M. Serotonin signalling in the gut--functions, dysfunctions and therapeutic targets. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 473–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, X.; Ding, C.; Zhao, W.; Xu, L.; Tian, H.; Gong, J.; Zhu, M.; Li, J.; Li, N. Antibiotics-induced depletion of mice microbiota induces changes in host serotonin biosynthesis and intestinal motility. J. Transl. Med. 2017, 15, 13. [Google Scholar] [CrossRef] [Green Version]
- Kashyap, P.C.; Marcobal, A.; Ursell, L.K.; Larauche, M.; Duboc, H.; Earle, K.A.; Sonnenburg, E.D.; Ferreyra, J.A.; Higginbottom, S.K.; Million, M.; et al. Complex interactions among diet, gastrointestinal transit, and gut microbiota in humanized mice. Gastroenterology 2013, 144, 967–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waclawiková, B.; Bullock, A.; Schwalbe, M.; Aranzamendi, C.; Nelemans, S.A.; van Dijk, G.; El Aidy, S. Gut bacteria-derived 5-hydroxyindole is a potent stimulant of intestinal motility via its action on L-type calcium channels. PLoS Biol. 2021, 19, e3001070. [Google Scholar] [CrossRef] [PubMed]
- Golubeva, A.V.; Joyce, S.A.; Moloney, G.; Burokas, A.; Sherwin, E.; Arboleya, S.; Flynn, I.; Khochanskiy, D.; Moya-Pérez, A.; Peterson, V.; et al. Microbiota-related changes in bile acid & tryptophan metabolism are associated with gastrointestinal dysfunction in a mouse model of autism. EBioMedicine 2017, 24, 166–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukumoto, S.; Tatewaki, M.; Yamada, T.; Fujimiya, M.; Mantyh, C.; Voss, M.; Eubanks, S.; Harris, M.; Pappas, T.N.; Takahashi, T. Short-chain fatty acids stimulate colonic transit via intraluminal 5-HT release in rats. Am. J. Physiol. Integr. Comp. Physiol. 2003, 284, R1269–R1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaki, S.; Mitsui, R.; Hayashi, H.; Kato, I.; Sugiya, H.; Iwanaga, T.; Furness, J.B.; Kuwahara, A. Short-chain fatty acid receptor, GPR43, is expressed by enteroendocrine cells and mucosal mast cells in rat intestine. Cell Tissue Res. 2006, 324, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Amaral, F.A.; Sachs, D.; Costa, V.V.; Fagundes, C.T.; Cisalpino, D.; Cunha, T.M.; Ferreira, S.H.; Cunha, F.Q.; Silva, T.A.; Nicoli, J.R.; et al. Commensal microbiota is fundamental for the development of inflammatory pain. Proc. Natl. Acad. Sci. USA 2008, 105, 2193–2197. [Google Scholar] [CrossRef] [Green Version]
- McVey Neufeld, K.A.; Mao, Y.K.; Bienenstock, J.; Foster, J.A.; Kunze, W.A. The microbiome is essential for normal gut intrinsic primary afferent neuron excitability in the mouse. Neurogastroenterol. Motil. 2013, 25, 183-e88. [Google Scholar] [CrossRef]
- Luczynski, P.; Tramullas, M.; Viola, M.; Shanahan, F.; Clarke, G.; O’Mahony, S.; Dinan, T.G.; Cryan, J.F. Microbiota regulates visceral pain in the mouse. eLife 2017, 6, e25887. [Google Scholar] [CrossRef]
- Guo, R.; Chen, L.-H.; Xing, C.; Liu, T. Pain regulation by gut microbiota: Molecular mechanisms and therapeutic potential. Br. J. Anaesth. 2019, 123, 637–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Thiel, I.A.M.; Botschuijver, S.; de Jonge, W.J.; Seppen, J. Painful interactions: Microbial compounds and visceral pain. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165534. [Google Scholar] [CrossRef]
- Lacy, B.E.; Mearin, F.; Chang, L.; Chey, W.D.; Lembo, A.J.; Simren, M.; Spiller, R. Bowel disorders. Gastroenterology 2016, 150, 1393–1407. [Google Scholar] [CrossRef] [Green Version]
- Crouzet, L.; Gaultier, E.; Del’Homme, C.; Cartier, C.; Delmas, E.; Dapoigny, M.; Fioramonti, J.; Bernalier-Donadille, A. The hypersensitivity to colonic distension of IBS patients can be transferred to rats through their fecal microbiota. Neurogastroenterol. Motil. 2013, 25, e272–e282. [Google Scholar] [CrossRef]
- Browning, K.N. Role of central vagal 5-HT3 receptors in gastrointestinal physiology and pathophysiology. Front. Neurosci. 2015, 9, 413. [Google Scholar] [CrossRef] [Green Version]
- Cangemi, D.J.; Lacy, B.E. Management of irritable bowel syndrome with diarrhea: A review of nonpharmacological and pharmacological interventions. Therap. Adv. Gastroenterol. 2019, 12, 1756284819878950. [Google Scholar] [CrossRef] [PubMed]
- Kato, S. Role of serotonin 5-HT3 receptors in intestinal inflammation. Biol. Pharm. Bull. 2013, 36, 1406–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camilleri, M.; Lasch, K.; Zhou, W. Irritable bowel syndrome: Methods, mechanisms, and pathophysiology. The confluence of increased permeability, inflammation, and pain in irritable bowel syndrome. AJP Gastrointest. Liver Physiol. 2012, 303, G775–G785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keszthelyi, D.; Troost, F.J.; Jonkers, D.M.; van Eijk, H.M.; Lindsey, P.J.; Dekker, J.; Buurman, W.A.; Masclee, A.A.M. Serotonergic reinforcement of intestinal barrier function is impaired in irritable bowel syndrome. Aliment. Pharmacol. Ther. 2014, 40, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Shajib, M.S.; Khan, W.I. The role of serotonin and its receptors in activation of immune responses and inflammation. Acta Physiol. 2015, 213, 561–574. [Google Scholar] [CrossRef] [PubMed]
- Herr, N.; Bode, C.; Duerschmied, D. The effects of serotonin in immune cells. Front. Cardiovasc. Med. 2017, 4, 48. [Google Scholar] [CrossRef] [Green Version]
- Ghia, J.; Li, N.; Wang, H.; Collins, M.; Deng, Y.; El–Sharkawy, R.T.; Côté, F.; Mallet, J.; Khan, W.I. Serotonin has a key role in pathogenesis of experimental colitis. Gastroenterology 2009, 137, 1649–1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Ghia, J.-E.; Wang, H.; McClemens, J.; Cote, F.; Suehiro, Y.; Mallet, J.; Khan, W.I. Serotonin activates dendritic cell function in the context of gut inflammation. Am. J. Pathol. 2011, 178, 662–671. [Google Scholar] [CrossRef] [Green Version]
- Bischoff, S.C.; Mailer, R.; Pabst, O.; Weier, G.; Sedlik, W.; Li, Z.; Chen, J.J.; Murphy, D.L.; Gershon, M.D. Role of serotonin in intestinal inflammation: Knockout of serotonin reuptake transporter exacerbates 2,4,6-trinitrobenzene sulfonic acid colitis in mice. Am. J. Physiol. Liver Physiol. 2009, 296, G685–G695. [Google Scholar] [CrossRef]
- Margolis, K.G.; Stevanovic, K.; Li, Z.; Yang, Q.M.; Oravecz, T.; Zambrowicz, B.; Jhaver, K.G.; Diacou, A.; Gershon, M.D. Pharmacological reduction of mucosal but not neuronal serotonin opposes inflammation in mouse intestine. Gut 2014, 63, 928–937. [Google Scholar] [CrossRef]
- El-Salhy, M.; Hausken, T.; Gilja, O.H.; Hatlebakk, J.G. The possible role of gastrointestinal endocrine cells in the pathophysiology of irritable bowel syndrome. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 139–148. [Google Scholar] [CrossRef]
- Ng, Q.X.; Soh, A.Y.S.; Loke, W.; Lim, D.Y.; Yeo, W.-S. The role of inflammation in irritable bowel syndrome (IBS). J. Inflamm. Res. 2018, 11, 345–349. [Google Scholar] [CrossRef] [Green Version]
- Ishihara, S.; Tada, Y.; Fukuba, N.; Oka, A.; Kusunoki, R.; Mishima, Y.; Oshima, N.; Moriyama, I.; Yuki, T.; Kawashima, K.; et al. Pathogenesis of irritable bowel syndrome—Review regarding associated infection and immune activation. Digestion 2013, 87, 204–211. [Google Scholar] [CrossRef]
- Zhang, L.; Song, J.; Hou, X. Mast cells and irritable bowel syndrome: From the bench to the bedside. J. Neurogastroenterol. Motil. 2016, 22, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.J.; Sharkey, K.A.; McKay, D.M. Modulation of the immune response by helminths: A role for serotonin? Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, K.; Lo, M.W.; Hosoya, T.; Tashima, K.; Takayama, H.; Murayama, T.; Horie, S. Experimental colitis alters expression of 5-HT receptors and transient receptor potential vanilloid 1 leading to visceral hypersensitivity in mice. Lab. Investig. 2012, 92, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Coldwell, J.R.; Phillis, B.D.; Sutherland, K.; Howarth, G.S.; Blackshaw, L.A. Increased responsiveness of rat colonic splanchnic afferents to 5-HT after inflammation and recovery. J. Physiol. 2007, 579, 203–213. [Google Scholar] [CrossRef]
- Ma, J.; Li, J.; Qian, M.; He, N.; Cao, Y.; Liu, Y.; Wu, K.; He, S. The comprehensive pathophysiological changes in a novel rat model of postinflammatory visceral hypersensitivity. FASEB J. 2019, 33, 13560–13571. [Google Scholar] [CrossRef] [Green Version]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; de Roos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- Charnay, Y.; Léger, L. Brain serotonergic circuitries. Dialogues Clin. Neurosci. 2010, 12, 471–487. [Google Scholar] [CrossRef]
- Carhart-Harris, R.; Nutt, D. Serotonin and brain function: A tale of two receptors. J. Psychopharmacol. 2017, 31, 1091–1120. [Google Scholar] [CrossRef] [Green Version]
- Barnes, N.M.; Sharp, T. A review of central 5-HT receptors and their function. Neuropharmacology 1999, 38, 1083–1152. [Google Scholar] [CrossRef]
- Franke, L.; Schewe, H.-J.; Uebelhack, R.; Berghöfer, A.; Müller-Oerlinghausen, B. Platelet-5HT uptake and gastrointestinal symptoms in patients suffering from major depression. Life Sci. 2003, 74, 521–531. [Google Scholar] [CrossRef]
- Martin, C.B.; Martin, V.S.; Trigo, J.M.; Chevarin, C.; Maldonado, R.; Fink, L.H.; Cunningham, K.A.; Hamon, M.; Lanfumey, L.; Mongeau, R. 5-HT2C Receptor desensitization moderates anxiety in 5-HTT deficient mice: From behavioral to cellular evidence. Int. J. Neuropsychopharmacol. 2015, 18, pyu056. [Google Scholar] [CrossRef] [Green Version]
- Rogers, G.B.; Keating, D.J.; Young, R.L.; Wong, M.-L.; Licinio, J.; Wesselingh, S. From gut dysbiosis to altered brain function and mental illness: Mechanisms and pathways. Mol. Psychiatry 2016, 21, 738–748. [Google Scholar] [CrossRef] [Green Version]
- Mathee, K.; Cickovski, T.; Deoraj, A.; Stollstorff, M.; Narasimhan, G. The gut microbiome and neuropsychiatric disorders: Implications for attention deficit hyperactivity disorder (ADHD). J. Med. Microbiol. 2020, 69, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.-F.; Zhu, Y.-L.; Zhou, Z.-L.; Jia, X.-B.; Xu, Y.-D.; Yang, Q.; Cui, C.; Shen, Y.-Q. Neuroprotective effects of fecal microbiota transplantation on MPTP-induced Parkinson’s disease mice: Gut microbiota, glial reaction and TLR4/TNF-α signaling pathway. Brain Behav. Immun. 2018, 70, 48–60. [Google Scholar] [CrossRef]
- Dekel, R.; Drossman, D.A.; Sperber, A.D. The use of psychotropic drugs in irritable bowel syndrome. Expert Opin. Investig. Drugs 2013, 22, 329–339. [Google Scholar] [CrossRef] [PubMed]
- North, C.S.; Hong, B.A.; Alpers, D.H. Relationship of functional gastrointestinal disorders and psychiatric disorders: Implications for treatment. World J. Gastroenterol. 2007, 13, 2020–2027. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.M.; Yabut, J.M.; Choo, J.M.; Page, A.J.; Sun, E.W.; Jessup, C.F.; Wesselingh, S.L.; Khan, W.I.; Rogers, G.B.; Steinberg, G.R.; et al. The gut microbiome regulates host glucose homeostasis via peripheral serotonin. Proc. Natl. Acad. Sci. USA 2019, 116, 19802–19804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Niu, K.; Momma, H.; Kobayashi, Y.; Chujo, M.; Otomo, A.; Fukudo, S.; Nagatomi, R. Irritable bowel syndrome is positively related to metabolic syndrome: A population-based cross-sectional study. PLoS ONE 2014, 9, e112289. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.A.; Baffy, N. Modulation of the gut microbiota: A focus on treatments for irritable bowel syndrome. Postgrad. Med. 2017, 129, 872–888. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Overview of serotonergic pathway modulated by gut commensal microbiota components. Gut 5-HT is biosynthesized with L-tryptophan (Trp) derived from dietary contents in enterochromaffin (EC) cells, in which tryptophan hydroxylase 1 (TPH1) converts Trp into 5-hydroxytryptophan (5-HTP), a direct precursor of 5-HT. Activation of EC cells and TPH1 is dependent on resident microbiota and their metabolites. Released 5-HT binds to various 5-HT receptors (5-HTRs) on immune cells, such as mast cells and macrophages, which potentially induces mucosal inflammation in the gut. Enteric 5-HT can modulate GI motility by stimulating the enteric nervous system (ENS), while it also acts to induce pain by activating afferent fibers as well as inducing mucosal inflammation, mainly through 5-HT3 and 5-HT4 receptors. Released 5-HT is taken up by serotonin reuptake transport (SERT) and degraded by monoamine oxidase (MAO) into 5-hydroxyindole acetic acid (5-HIAA) in the cells to optimize the luminal 5-HT level. 5-HT can also influence bacterial composition by quorum sensing (QS).
Figure 1.
Overview of serotonergic pathway modulated by gut commensal microbiota components. Gut 5-HT is biosynthesized with L-tryptophan (Trp) derived from dietary contents in enterochromaffin (EC) cells, in which tryptophan hydroxylase 1 (TPH1) converts Trp into 5-hydroxytryptophan (5-HTP), a direct precursor of 5-HT. Activation of EC cells and TPH1 is dependent on resident microbiota and their metabolites. Released 5-HT binds to various 5-HT receptors (5-HTRs) on immune cells, such as mast cells and macrophages, which potentially induces mucosal inflammation in the gut. Enteric 5-HT can modulate GI motility by stimulating the enteric nervous system (ENS), while it also acts to induce pain by activating afferent fibers as well as inducing mucosal inflammation, mainly through 5-HT3 and 5-HT4 receptors. Released 5-HT is taken up by serotonin reuptake transport (SERT) and degraded by monoamine oxidase (MAO) into 5-hydroxyindole acetic acid (5-HIAA) in the cells to optimize the luminal 5-HT level. 5-HT can also influence bacterial composition by quorum sensing (QS).

Figure 2.
Influence of gut dysbiosis on serotonergic signaling in pathology of IBS. A dysbiotic condition with abundant microbial species that upregulate 5-HT pathway and/or diminished microbial groups that downregulate 5-HT signaling results in massive synthesis and hyperavailability of 5-HT through activating enterochromaffin (EC) cells and tryptophan hydroxylase 1 (TPH1), and decreasing expression of serotonin reuptake transport (SERT). Excessive 5-HT can induce low-grade mucosal inflammation, enhance visceral pain sensation, and promote GI motility, potentially leading to IBS-D development. In contrast, the other dysbiotic condition with decreased microbial species that upregulate 5-HT pathway and/or increased microbial groups that downregulate 5-HT signaling causes limited production of 5-HT, resulting in impaired GI motility. This condition may be associated with IBS-C or chronic constipation. Trp, L-tryptophan; IBS-D, diarrhea-predominant irritable bowel syndrome; IBS-C, constipation-predominant irritable bowel syndrome.
Figure 2.
Influence of gut dysbiosis on serotonergic signaling in pathology of IBS. A dysbiotic condition with abundant microbial species that upregulate 5-HT pathway and/or diminished microbial groups that downregulate 5-HT signaling results in massive synthesis and hyperavailability of 5-HT through activating enterochromaffin (EC) cells and tryptophan hydroxylase 1 (TPH1), and decreasing expression of serotonin reuptake transport (SERT). Excessive 5-HT can induce low-grade mucosal inflammation, enhance visceral pain sensation, and promote GI motility, potentially leading to IBS-D development. In contrast, the other dysbiotic condition with decreased microbial species that upregulate 5-HT pathway and/or increased microbial groups that downregulate 5-HT signaling causes limited production of 5-HT, resulting in impaired GI motility. This condition may be associated with IBS-C or chronic constipation. Trp, L-tryptophan; IBS-D, diarrhea-predominant irritable bowel syndrome; IBS-C, constipation-predominant irritable bowel syndrome.


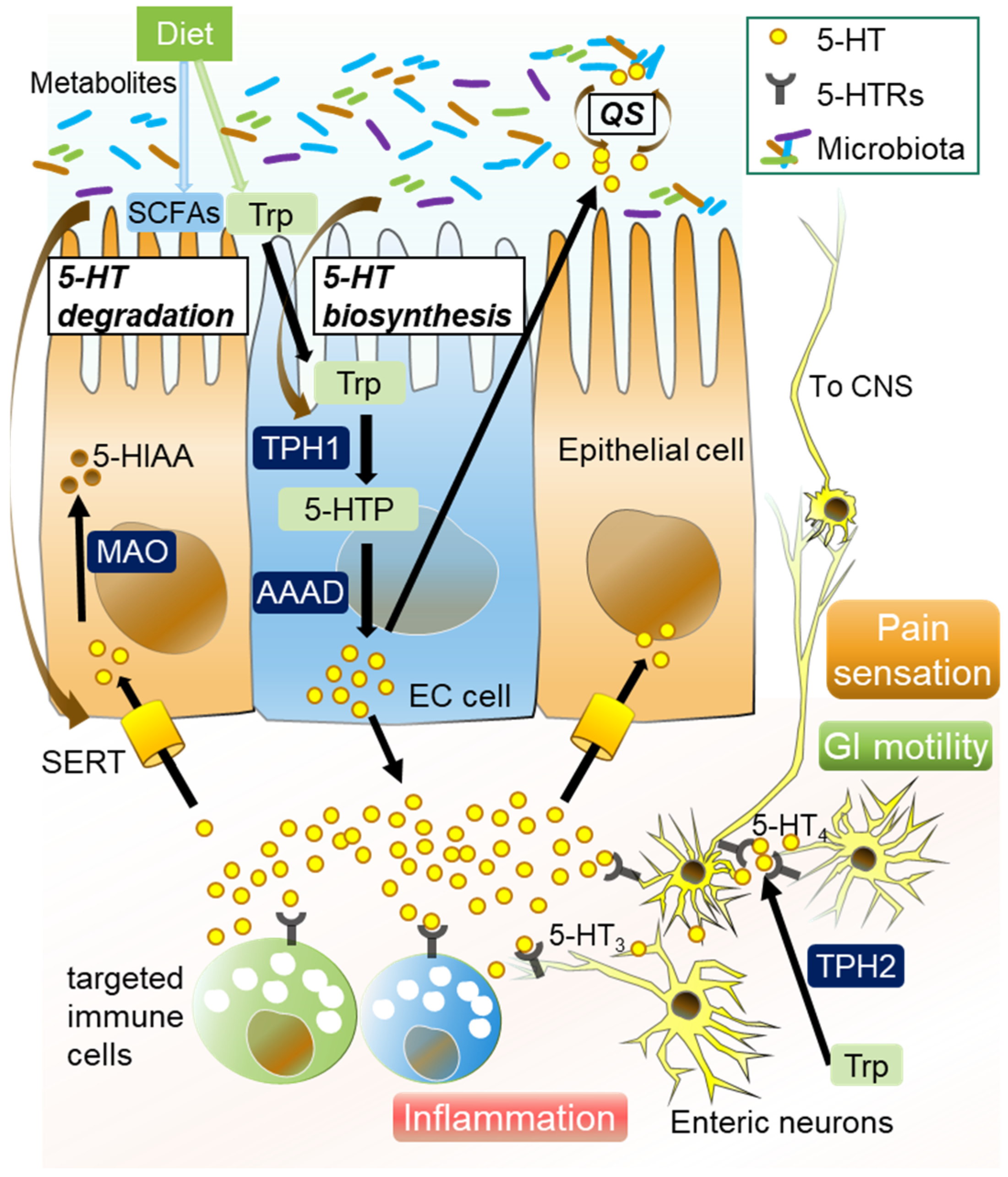{kind=link}
{kind=link}
Table 1.
Specific enteric microbiota members that regulate serotonergic pathway in GI.
| Upregulation of 5-HT in Microbiota | Mechanisms of Action and Observations | Ref. |
| Akkermansia muciniphila (Amuc_1100) | Promote intestinal 5-HT biosynthesis and extracellular availability through TLR2 signaling. | [88] |
| Akkermansiamuciniphila (extracellular vesicles) | Increase expression of the Htr4 gene, and decreases that of the Htr2B, Htr3B, and Htr7 genes. | [89] |
| Bacteriodes thetaiotaomicron | Restore 5-HT+ EC cells and shape EC networks in the GI tract of GF mice by producing SCFAs. | [96] |
| Bifidobacterium dentium | Increase intestinal 5-HT level, expressions of 5-HTra receptors 2a and 4, and SERT by producing acetate. | [97] |
| Clostridium ramosum | Promote 5-HT synthesis in colonic EC cells and program differentiation of intestinal stem progenitors toward a secretory 5-HT-producing lineage. | [85,87] |
| Corynebacterium spp., Enterococcus spp., Streptococcus spp. | Enable the direct production of 5-HT. | [86] |
| Escherichia coli Nissle 1917 | Enhance 5-HT bioavailability in ileal tissue through interaction with compounds secreted from host tissue. | [91] |
| Indigenous spore-forming bacteria | Enhance colonic 5-HT pathway by upregulation of Htr4. | [87] |
| Lactobacillus plantarum IS-10506 | Increase gut 5-HT production along with brain 5-HTT, neurotrophin, and brain-derived neurotrophic factor. | [98] |
| Lactobacillus plantarum PS128 | Increase 5-HT+ cells in the gut and alter expression levels of Tph1, Chga, Slc6a4, and Htr4. | [99] |
| SadA-expressing Staphylococci | Promote converting 5-HTP into 5-HT. | [90] |
| Trichinella spiralis and Campylobacter jejuni (pathogens) | Increase EC cell number and reduce SERT expression. | [100,101] |
| Downregulation of 5-HT in Microbiota | Mechanisms of action and observations | Ref. |
| Bifidobacterium longum and Lactobacillus acidophilus | Upregulate SERT expression. | [102] |
| Bifidobacterium pseudolongum | Diminish EC cells. | [103] |
| Lactobacillus rhamnosus | Upregulate gene and protein level of SERT. | [92,100] |
Table 2.
Specific enteric microbiota influenced by aberrant serotonergic pathway in GI.
| 5-HT Pathway-Induced Specific Dysbiosis | Mechanisms of Action and Observations | Ref. |
|---|---|---|
| Turicibacter sanguinis | Reduce sporulation factors and membrane transporters by 5-HT supplementation and in SERT−/− mice. | [93] |
| Bacilli spp. (Lactobacillus, Streptococcus, Enterococcus, and Listeria) | Increase in SERT−/− mice. | [94] |
| Bifidobacterium spp. | Decrease in SERT−/− mice. | [94] |
| Akkermansia muciniphila | Increase in Tph1–/– mice and decrease in SERT−/− mice. | [94,95] |
| Two distinct Bacteroidales OTUs | Decrease in Tph1–/– mice. | [95] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mishima, Y.; Ishihara, S. Enteric Microbiota-Mediated Serotonergic Signaling in Pathogenesis of Irritable Bowel Syndrome. Int. J. Mol. Sci. 2021, 22, 10235. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910235
AMA Style
Mishima Y, Ishihara S. Enteric Microbiota-Mediated Serotonergic Signaling in Pathogenesis of Irritable Bowel Syndrome. International Journal of Molecular Sciences. 2021; 22(19):10235. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910235
Chicago/Turabian StyleMishima, Yoshiyuki, and Shunji Ishihara. 2021. "Enteric Microbiota-Mediated Serotonergic Signaling in Pathogenesis of Irritable Bowel Syndrome" International Journal of Molecular Sciences 22, no. 19: 10235. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910235
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.