
Chrysin Ameliorates Cyclosporine-A-Induced Renal Fibrosis by Inhibiting TGF-β1-Induced Epithelial–Mesenchymal Transition


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effect of Chrysin on CsA-Induced Renal Dysfunction
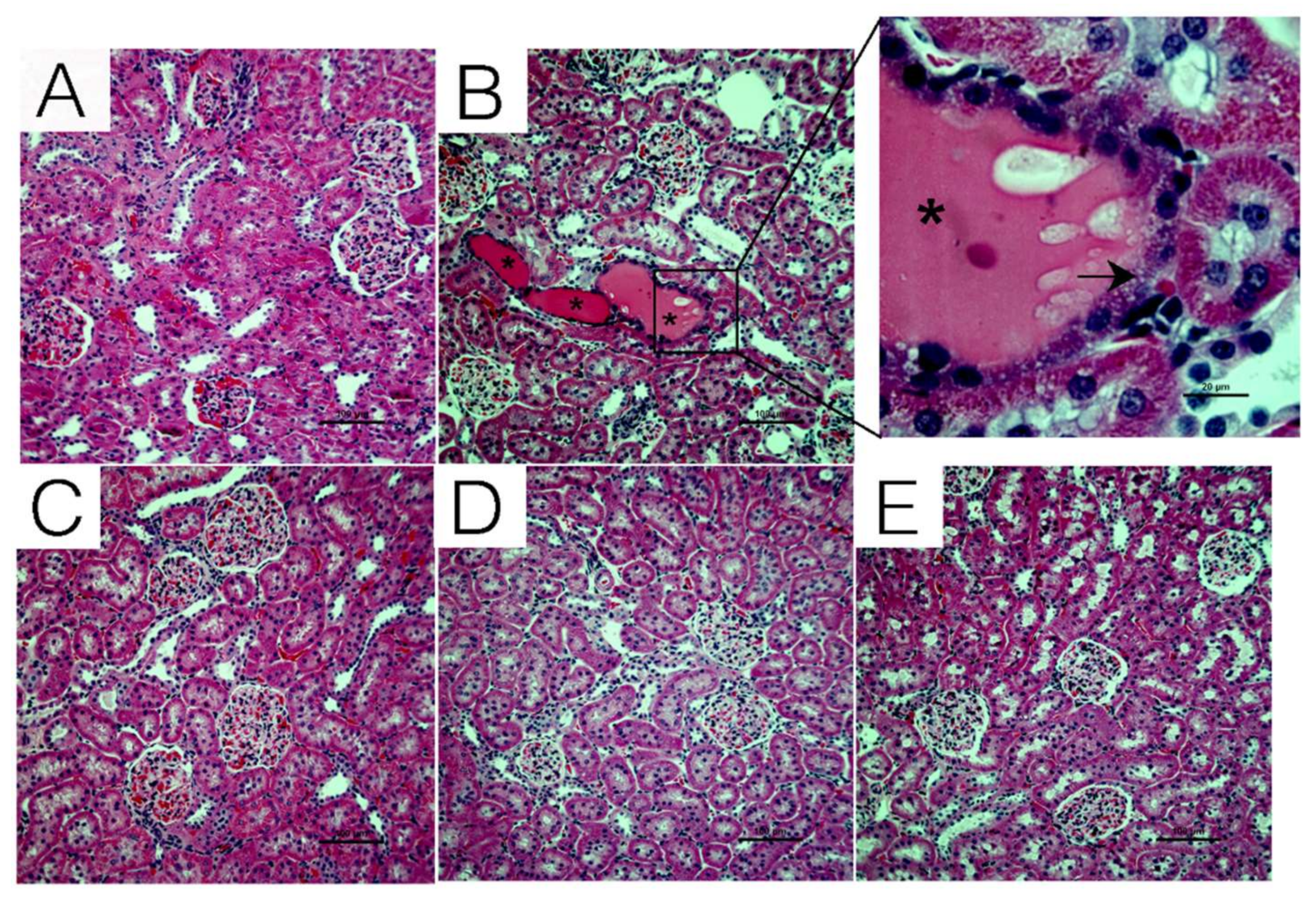
2.2. Effect of Chrysin on Histopathological Damage Induced by CsA
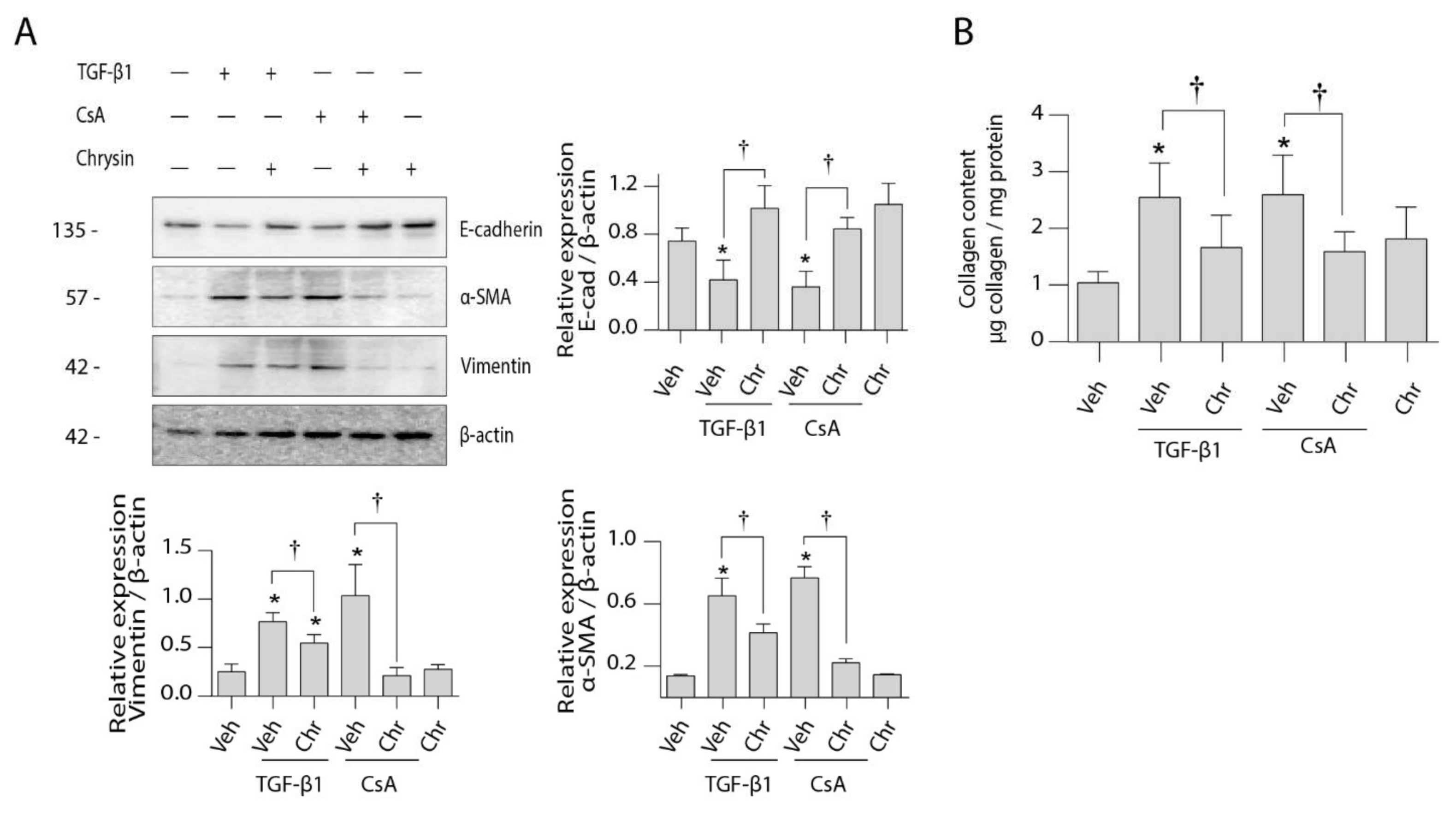
2.3. Effect of Chrysin on CsA-Induced Collagen Deposition
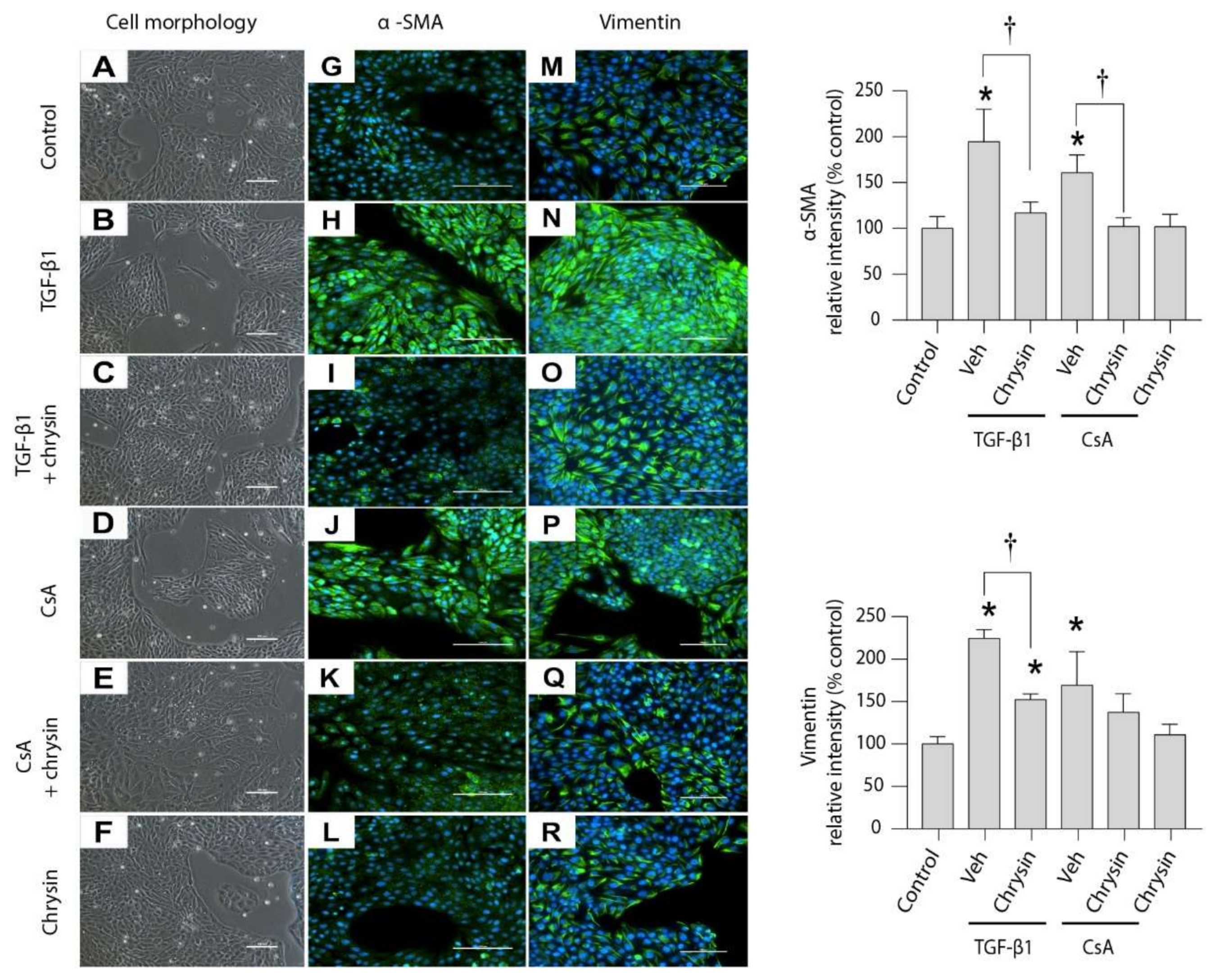
2.4. Effect of Chrysin on TGF-β1- or CsA-Induced Changes in LLC-PK1 Cells
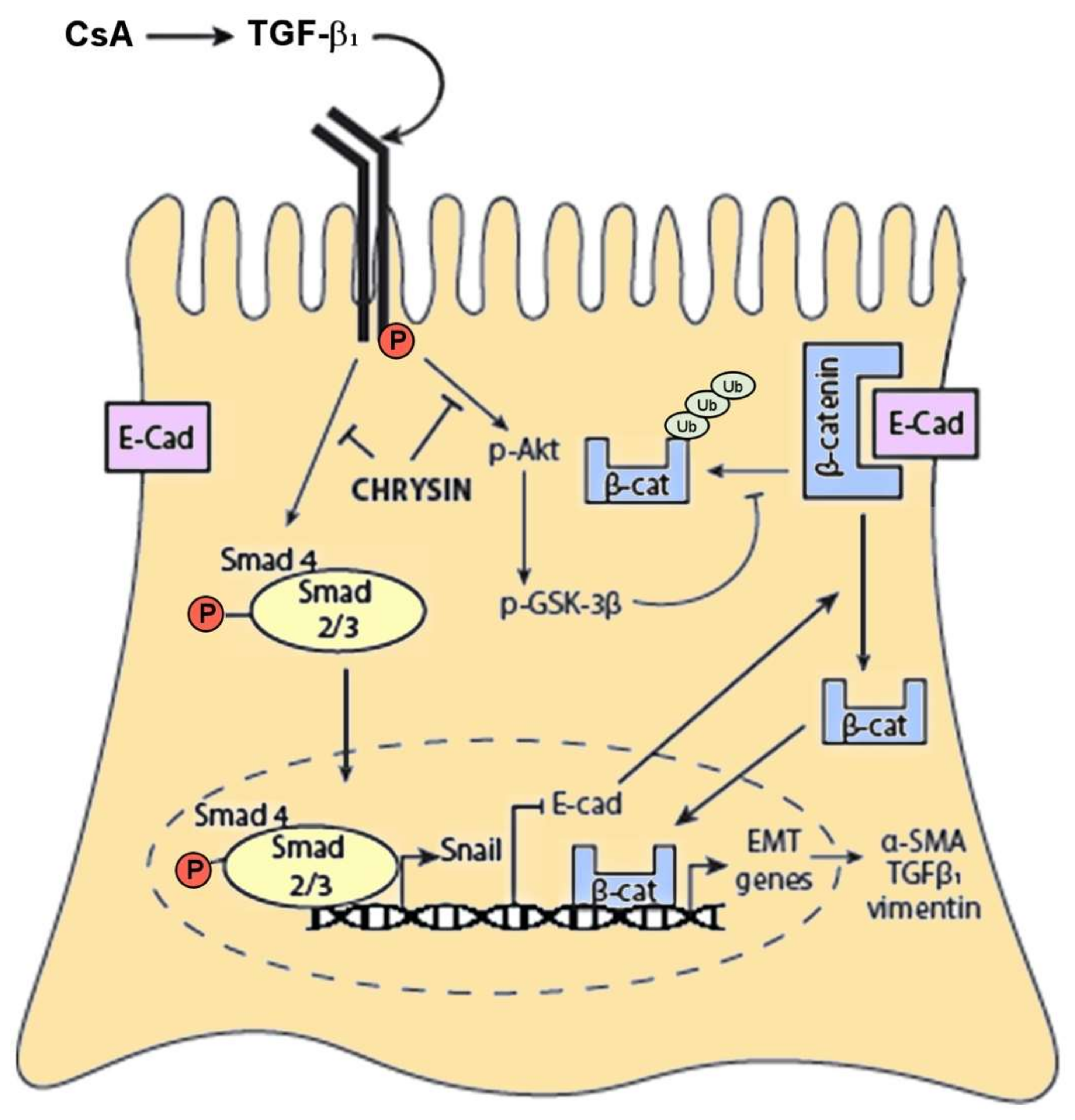
2.5. Effect of Chrysin on CsA-Induced TGF-β1 Signaling Cascade
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animal Studies
4.3. Analysis of Urine and Serum Chemistry
4.4. Histology
4.5. Cell Culture
4.6. Treatment of Cells
4.7. Assessment of EMT in Vitro by Morphological Examination and Immunocytochemistry
4.8. Cell Migration Assay
4.9. Collagen Determination
4.10. Determination of Protein Expression by Western Blot Analysis
4.11. Determination of Protein Expression by ELISA
4.12. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dalrymple, L.S.; Katz, R.; Kestenbaum, B.; Shlipak, M.G.; Sarnak, M.J.; Stehman-Breen, C.; Seliger, S.; Siscovick, D.; Newman, A.B.; Fried, L. Chronic Kidney Disease and the Risk of End-Stage Renal Disease versus Death. J. Gen. Intern. Med. 2011, 26, 379–385. [Google Scholar] [CrossRef] [Green Version]
- Grande, M.T.; Sánchez-Laorden, B.; López-Blau, C.; De Frutos, C.A.; Boutet, A.; Arévalo, M.; Rowe, R.G.; Weiss, S.J.; López-Novoa, J.M.; Nieto, M.A. Snail1-Induced Partial Epithelial-to-Mesenchymal Transition Drives Renal Fibrosis in Mice and Can Be Targeted to Reverse Established Disease. Nat. Med. 2015, 21, 989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphreys, B.D.; Duffield, J.D.; Bonventre, J.V. Renal Stem Cells in Recovery from Acute Kidney Injury. Minerva Urol. Nefrol. Ital. J. Urol. Nephrol. 2006, 58, 13–21. [Google Scholar]
- Ishibe, S.; Cantley, L.G. Epithelial–Mesenchymal–Epithelial Cycling in Kidney Repair. Curr. Opin. Nephrol. Hypertens. 2008, 17, 379–385. [Google Scholar] [CrossRef]
- Kalluri, R.; Weinberg, R.A. The Basics of Epithelial-Mesenchymal Transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christiansen, J.J.; Rajasekaran, A.K. Reassessing Epithelial to Mesenchymal Transition as a Prerequisite for Carcinoma Invasion and Metastasis. Cancer Res. 2006, 66, 8319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, T.; Noble, N.A.; Miller, D.E.; Border, W.A. Sustained Expression of TGF-Β1 Underlies Development of Progressive Kidney Fibrosis. Kidney Int. 1994, 45, 916–927. [Google Scholar] [CrossRef] [Green Version]
- Verrecchia, F.; Mauviel, A. Transforming Growth Factor-β Signaling Through the Smad Pathway: Role in Extracellular Matrix Gene Expression and Regulation. J. Investig. Dermatol. 2002, 118, 211–215. [Google Scholar] [CrossRef] [Green Version]
- Derynck, R.; Zhang, Y.E. Smad-Dependent and Smad-Independent Pathways in TGF-β Family Signalling. Nature 2003, 425, 577–584. [Google Scholar] [CrossRef]
- Rahimi, R.A.; Leof, E.B. TGF-β Signaling: A Tale of Two Responses. J. Cell. Biochem. 2007, 102, 593–608. [Google Scholar] [CrossRef]
- Medici, D.; Hay, E.D.; Goodenough, D.A. Cooperation between Snail and LEF-1 Transcription Factors Is Essential for TGF-Β1-Induced Epithelial-Mesenchymal Transition. Mol. Biol. Cell 2006, 17, 1871–1879. [Google Scholar] [CrossRef] [Green Version]
- Kattla, J.J.; Carew, R.M.; Heljić, M.; Godson, C.; Brazil, D.P. Protein Kinase B/Akt Activity Is Involved in Renal TGF-Β1-Driven Epithelial-Mesenchymal Transition in Vitro and in Vivo. Am. J. Physiol. Ren. Physiol. 2008, 295, F215–F225. [Google Scholar] [CrossRef] [Green Version]
- Curci, C.; Castellano, G.; Stasi, A.; Divella, C.; Loverre, A.; Gigante, M.; Simone, S.; Cariello, M.; Montinaro, V.; Lucarelli, G.; et al. Endothelial-to-Mesenchymal Transition and Renal Fibrosis in Ischaemia/Reperfusion Injury Are Mediated by Complement Anaphylatoxins and Akt Pathway. Nephrol. Dial. Transplant. 2014, 29, 799–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, R.; Xia, L.; Ning, X.; Liu, L.; Sun, W.; Huang, C.; Wang, H.; Sun, S.; Chernoff, J. Hypoxia-Induced Bmi1 Promotes Renal Tubular Epithelial Cell–Mesenchymal Transition and Renal Fibrosis via PI3K/Akt Signal. Mol. Biol. Cell 2014, 25, 2650–2659. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Jing, Z.; Deng, H.; Li, Z.; Zhuang, Z.; Wang, S.; Wang, Y. Soluble Epoxide Hydrolase Inhibition Ameliorates Proteinuria-Induced Epithelial-Mesenchymal Transition by Regulating the PI3K-Akt-GSK-3β Signaling Pathway. Biochem. Biophys. Res. Commun. 2015, 463, 70–75. [Google Scholar] [CrossRef]
- Cho, H.J.; Baek, K.E.; Saika, S.; Jeong, M.-J.; Yoo, J. Snail Is Required for Transforming Growth Factor-Beta-Induced Epithelial-Mesenchymal Transition by Activating PI3 Kinase/Akt Signal Pathway. Biochem. Biophys. Res. Commun. 2007, 353, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.; Lou, L.; Gupta, S.; Ribeiro-Neto, F.; Altschuler, D.L. A Novel Epac-Rap-PP2A Signaling Module Controls CAMP-Dependent Akt Regulation. J. Biol. Chem. 2008, 283, 23129–23138. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Han, H.J. Troglitazone Ameliorates High Glucose-Induced EMT and Dysfunction of SGLTs through PI3K/Akt, GSK-3β, Snail1, and β-Catenin in Renal Proximal Tubule Cells. Am. J. Physiol. Ren. Physiol. 2009, 298, F1263–F1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bienz, M. β-Catenin: A Pivot between Cell Adhesion and Wnt Signalling. Curr. Biol. 2005, 15, R64–R67. [Google Scholar] [CrossRef] [Green Version]
- Yook, J.I.; Li, X.-Y.; Ota, I.; Fearon, E.R.; Weiss, S.J. Wnt-Dependent Regulation of the E-Cadherin Repressor Snail. J. Biol. Chem. 2005, 280, 11740–11748. [Google Scholar] [CrossRef] [Green Version]
- Prokop, J.W.; Liu, Y.; Milsted, A.; Peng, H.; Rauscher, F.J. A Method for in Silico Identification of SNAIL/SLUG DNA Binding Potentials to the E-Box Sequence Using Molecular Dynamics and Evolutionary Conserved Amino Acids. J. Mol. Model. 2013, 19, 3463–3469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.; Qin, H.; Shi, Q.; Zhang, Y.; Zhou, F.; Wu, H.; Ding, S.; Niu, Z.; Lu, Y.; Shen, P. Chrysin Attenuates Inflammation by Regulating M1/M2 Status via Activating PPARgamma. Biochem. Pharmacol. 2014, 89, 503–514. [Google Scholar] [CrossRef]
- Jiang, Y.; Gong, F.-L.; Zhao, G.-B.; Li, J. Chrysin Suppressed Inflammatory Responses and the Inducible Nitric Oxide Synthase Pathway after Spinal Cord Injury in Rats. Int. J. Mol. Sci. 2014, 15, 12270–12279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, K.V.; Mohamed Jaabir, M.S.; Thomas, P.A.; Geraldine, P. Protective Role of Chrysin against Oxidative Stress in D-Galactose-Induced Aging in an Experimental Rat Model. Geriatr. Gerontol. Int. 2012, 12, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Rashid, S.; Ali, N.; Nafees, S.; Ahmad, S.T.; Arjumand, W.; Hasan, S.K.; Sultana, S. Alleviation of Doxorubicin-Induced Nephrotoxicity and Hepatotoxicity by Chrysin in Wistar Rats. Toxicol. Mech. Methods 2013, 23, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Pingili, R.B.; Pawar, A.K.; Challa, S.R.; Kodali, T.; Koppula, S.; Toleti, V. A Comprehensive Review on Hepatoprotective and Nephroprotective Activities of Chrysin against Various Drugs and Toxic Agents. Chem. Biol. Interact. 2019, 308, 51–60. [Google Scholar] [CrossRef]
- Kang, M.-K.; Park, S.-H.; Choi, Y.-J.; Shin, D.; Kang, Y.-H. Chrysin Inhibits Diabetic Renal Tubulointerstitial Fibrosis through Blocking Epithelial to Mesenchymal Transition. J. Mol. Med. 2015, 93, 759–772. [Google Scholar] [CrossRef]
- Lee, E.-J.; Kang, M.-K.; Kim, D.Y.; Kim, Y.-H.; Oh, H.; Kang, Y.-H. Chrysin Inhibits Advanced Glycation End Products-Induced Kidney Fibrosis in Renal Mesangial Cells and Diabetic Kidneys. Nutrients 2018, 10, 882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, D.J.; Loertcher, R.; Rubin, M.F.; Tilney, N.L.; Carpenter, C.B.; Strom, T.B. Cyclosporine: A New Immunosuppressive Agent for Organ Transplantation. Ann. Intern. Med. 1984, 101, 667–682. [Google Scholar] [CrossRef] [PubMed]
- Shih, W.; Hines, W.H.; Neilson, E.G. Effects of Cyclosporin A on the Development of Immune-Mediated Interstitial Nephritis. Kidney Int. 1988, 33, 1113–1118. [Google Scholar] [CrossRef] [Green Version]
- Bennett, W.M.; DeMattos, A.; Meyer, M.M.; Andoh, T.; Barry, J.M. Chronic Cyclosporine Nephropathy: The Achilles’ Heel of Immunosuppressive Therapy. Kidney Int. 1996, 50, 1089–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klintmalm, G.; Sundelin, B.; Bohman, S.-O.; Wilczek, H. Interstitial Fibrosis in Renal Allografts after 12 to 46 Months of Cyclosporin Treatment: Beneficial Effect of Low Doses in Early Post-Transplantation Period. Lancet 1984, 324, 950–954. [Google Scholar] [CrossRef]
- Ruiz, P.; Kolbeck, P.C.; Scroggs, M.W.; Sanfilippo, F. Associations between Cyclosporine Therapy and Interstitial Fibrosis in Renal Allograft Biopsies. Transplantation 1988, 45, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, S.H.; Jaremko, G.; Duraj, F.F.; Wilczek, H.E. Renal Fibrosis in Cyclosporin A-Treated Renal Allograft Recipients: Morphological Findings in Relation to Renal Hemodynamics. Transpl. Int. 1996, 9, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Shehata, M.; Cope, G.H.; Johnson, T.S.; Raftery, A.T.; El Nahas, A.M. Cyclosporine Enhances the Expression of TGF-β in the Juxtaglomerular Cells of the Rat Kidney. Kidney Int. 1995, 48, 1487–1496. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.; Burke, J.F.; McGowan, T.A.; Zhu, Y.; Dunn, S.R.; McCue, P.; Kanalas, J.; Sharma, K. Effect of Anti-Transforming Growth Factor-Βbgr; Antibodies in Cyclosporine-Induced Renal Dysfunction. Kidney Int. 2001, 59, 498–506. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Ye, J.; Yu, L.; Dong, X.; Feng, J.; Xiong, Y.; Gu, X.; Li, S. Klotho Mitigates Cyclosporine A (CsA)-Induced Epithelial–Mesenchymal Transition (EMT) and Renal Fibrosis in Rats. Int. Urol. Nephrol. 2017, 49, 345–352. [Google Scholar] [CrossRef]
- McMorrow, T.; Gaffney, M.M.; Slattery, C.; Campbell, E.; Ryan, M.P. Cyclosporine A Induced Epithelial–Mesenchymal Transition in Human Renal Proximal Tubular Epithelial Cells. Nephrol. Dial. Transplant. 2005, 20, 2215–2225. [Google Scholar] [CrossRef] [Green Version]
- Slattery, C.; Campbell, E.; McMorrow, T.; Ryan, M.P. Cyclosporine A-Induced Renal Fibrosis: A Role for Epithelial-Mesenchymal Transition. Am. J. Pathol. 2005, 167, 395–407. [Google Scholar] [CrossRef]
- Kuruş, M.; Eşrefoğlu, M.; Bay, A.; Öztürk, F. Protective Effect of Oral L-Arginine Supplementation on Cyclosporine Induced Nephropathy in Rats. Int. Urol. Nephrol. 2005, 37, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Ateşşahin, A.; Çeribaşı, A.O.; Yılmaz, S. Lycopene, a Carotenoid, Attenuates Cyclosporine-Induced Renal Dysfunction and Oxidative Stress in Rats. Basic Clin. Pharmacol. Toxicol. 2007, 100, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Diegelmann, R.F.; Evans, M.C. Wound Healing: An Overview of Acute, Fibrotic and Delayed Healing. Front. Biosci. J. Virtual Libr. 2004, 9, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Jiang, M.; Zhang, Y.; Liu, X.; Du, Q.; Feng, G. Chrysin Alleviates Allergic Inflammation and Airway Remodeling in a Murine Model of Chronic Asthma. Int. Immunopharmacol. 2016, 32, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Balta, C.; Herman, H.; Boldura, O.M.; Gasca, I.; Rosu, M.; Ardelean, A.; Hermenean, A. Chrysin Attenuates Liver Fibrosis and Hepatic Stellate Cell Activation through TGF-β/Smad Signaling Pathway. Chem. Biol. Interact. 2015, 240, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.H.; Al Za, M.; Adham, S.A.; Yasin, J.; Nemmar, A.; Schupp, N. Therapeutic Effect of Chrysin on Adenine-Induced Chronic Kidney Disease in Rats. Cell. Physiol. Biochem. 2016, 38, 248–257. [Google Scholar] [CrossRef]
- Sereno, J.; Rodrigues-Santos, P.; Vala, H.; Rocha-Pereira, P.; Alves, R.; Fernandes, J.; Santos-Silva, A.; Carvalho, E.; Teixeira, F.; Reis, F. Transition from Cyclosporine-Induced Renal Dysfunction to Nephrotoxicity in an in Vivo Rat Model. Int. J. Mol. Sci. 2014, 15, 8979–8997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, C.; Fornoni, A.; Cornacchia, F.; Bellotti, N.; Fasoli, G.; Foschi, A.; Mazzucchelli, I.; Mazzullo, T.; Semeraro, L.; Dal Canton, A. Cyclosporine Induces Different Responses in Human Epithelial, Endothelial and Fibroblast Cell Cultures. Kidney Int. 2000, 58, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Marconi, G.D.; Fonticoli, L.; Rajan, T.S.; Pierdomenico, S.D.; Trubiani, O.; Pizzicannella, J.; Diomede, F. Epithelial-Mesenchymal Transition (EMT): The Type-2 EMT in Wound Healing, Tissue Regeneration and Organ Fibrosis. Cells 2021, 10, 1587. [Google Scholar] [CrossRef]
- Kalluri, R.; Neilson, E.G. Epithelial-Mesenchymal Transition and Its Implications for Fibrosis. J. Clin. Investig. 2003, 112, 1776–1784. [Google Scholar] [CrossRef]
- Kasai, H.; Allen, J.T.; Mason, R.M.; Kamimura, T.; Zhang, Z. TGF-Β1 Induces Human Alveolar Epithelial to Mesenchymal Cell Transition (EMT). Respir. Res. 2005, 6, 56. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Huang, J.; Xiang, T.; Yin, X.; Luo, X.; Huang, J.; Luo, F.; Li, H.; Li, H.; Ren, G. Chrysin Inhibits Metastatic Potential of Human Triple-Negative Breast Cancer Cells by Modulating Matrix Metalloproteinase-10, Epithelial to Mesenchymal Transition, and PI3K/Akt Signaling Pathway. J. Appl. Toxicol. 2014, 34, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Katsuno, Y.; Lamouille, S.; Derynck, R. TGF-β Signaling and Epithelial–Mesenchymal Transition in Cancer Progression. Curr. Opin. Oncol. 2013, 25, 76–84. [Google Scholar] [CrossRef]
- Cabezas, F.; Farfán, P.; Marzolo, M.-P. Participation of the SMAD2/3 Signalling Pathway in the down Regulation of Megalin/LRP2 by Transforming Growth Factor Beta (TGF-SS1). PLoS ONE 2019, 14, e0213127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akool, E.-S.; Doller, A.; Babelova, A.; Tsalastra, W.; Moreth, K.; Schaefer, L.; Pfeilschifter, J.; Eberhardt, W. Molecular Mechanisms of TGFβ Receptor-Triggered Signaling Cascades Rapidly Induced by the Calcineurin Inhibitors Cyclosporin A and FK506. J. Immunol. 2008, 181, 2831. [Google Scholar] [CrossRef] [Green Version]
- Cho, W.; Kim, Y.; Kim, J.; Park, S.; Park, D.; Kim, B.-C.; Jeoung, D.; Kim, Y.-M.; Choe, J. Suppressor of Cytokine Signaling 1 Is a Positive Regulator of TGF-β–Induced Prostaglandin Production in Human Follicular Dendritic Cell–like Cells. J. Immunol. 2015, 194, 4287. [Google Scholar] [CrossRef]
- Cano, A.; Perez-Moreno, M.A.; Rodrigo, I.; Locascio, A.; Blanco, M.J.; del Barrio, M.G.; Portillo, F.; Nieto, M.A. The Transcription Factor Snail Controls Epithelial-Mesenchymal Transitions by Repressing E-Cadherin Expression. Nat. Cell Biol. 2000, 2, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Carver, E.A.; Jiang, R.; Lan, Y.; Oram, K.F.; Gridley, T. The Mouse Snail Gene Encodes a Key Regulator of the Epithelial-Mesenchymal Transition. Mol. Cell. Biol. 2001, 21, 8184–8188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batlle, E.; Sancho, E.; Franci, C.; Dominguez, D.; Monfar, M.; Baulida, J.; Garcia De Herreros, A. The Transcription Factor Snail Is a Repressor of E-Cadherin Gene Expression in Epithelial Tumour Cells. Nat. Cell Biol. 2000, 2, 84–89. [Google Scholar] [CrossRef]
- Medici, D.; Hay, E.D.; Olsen, B.R.; Bronner-Fraser, M. Snail and Slug Promote Epithelial-Mesenchymal Transition through β-Catenin–T-Cell Factor-4-Dependent Expression of Transforming Growth Factor-Β3. Mol. Biol. Cell 2008, 19, 4875–4887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masszi, A.; Fan, L.; Rosivall, L.; McCulloch, C.A.; Rotstein, O.D.; Mucsi, I.; Kapus, A. Integrity of Cell-Cell Contacts Is a Critical Regulator of TGF-Β1-Induced Epithelial-to-Myofibroblast Transition: Role for β-Catenin. Am. J. Pathol. 2004, 165, 1955–1967. [Google Scholar] [CrossRef]
- Kang, H.R.; Lee, C.G.; Homer, R.J.; Elias, J.A. Semaphorin 7A Plays a Critical Role in TGF-β1–Induced Pulmonary Fibrosis. J. Exp. Med. 2007, 204, 1083. [Google Scholar] [CrossRef]
- Caraci, F.; Gili, E.; Calafiore, M.; Failla, M.; La Rosa, C.; Crimi, N.; Sortino, M.A.; Nicoletti, F.; Copani, A.; Vancheri, C. TGF-Β1 Targets the GSK-3β/β-Catenin Pathway via ERK Activation in the Transition of Human Lung Fibroblasts into Myofibroblasts. Pharmacol. Res. 2008, 57, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Ramesh, G.; Sun, L.; Dong, Z. Impaired Wound Healing in Hypoxic Renal Tubular Cells: Roles of Hypoxia-Inducible Factor-1 and Glycogen Synthase Kinase 3β/β-Catenin Signaling. J. Pharmacol. Exp. Ther. 2012, 340, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Luo, Q.; Xiong, Z.; Liang, W.; Chen, L.; Xiong, Z. Telmisartan Counteracts TGF-Β1 Induced Epithelial-to-Mesenchymal Transition via PPAR-γ in Human Proximal Tubule Epithelial Cells. Int. J. Clin. Exp. Pathol. 2012, 5, 522–529. [Google Scholar] [PubMed]
- Hatanaka, H.; Koizumi, N.; Okumura, N.; Kay, E.P.; Mizuhara, E.; Hamuro, J.; Kinoshita, S. Epithelial-Mesenchymal Transition-Like Phenotypic Changes of Retinal Pigment Epithelium Induced by TGF-β Are Prevented by PPAR-γ Agonists. Investig. Ophthalmol. Vis. Sci. 2012, 53, 6955–6963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rani, N.; Bharti, S.; Bhatia, J.; Tomar, A.; Nag, T.C.; Ray, R.; Arya, D.S. Inhibition of TGF-β by a Novel PPAR-γ Agonist, Chrysin, Salvages β-Receptor Stimulated Myocardial Injury in Rats through MAPKs-Dependent Mechanism. Nutr. Metab. 2015, 12, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Pang, L.; Zhang, Y.; Lin, J.; Zhou, H. Fenofibrate Improved Interstitial Fibrosis of Renal Allograft through Inhibited Epithelial-Mesenchymal Transition Induced by Oxidative Stress. Oxid. Med. Cell. Longev. 2019, 2019, 8936856. [Google Scholar] [CrossRef]
- Lu, Q.; Wang, W.-W.; Zhang, M.-Z.; Ma, Z.-X.; Qiu, X.-R.; Shen, M.; Yin, X.-X. ROS Induces Epithelial-Mesenchymal Transition via the TGF-Β1/PI3K/Akt/MTOR Pathway in Diabetic Nephropathy. Exp. Ther. Med. 2019, 17, 835–846. [Google Scholar] [CrossRef] [Green Version]
- Zha, D.; Wu, S.; Gao, P.; Wu, X. Telmisartan Attenuates Uric Acid-Induced Epithelial-Mesenchymal Transition in Renal Tubular Cells. BioMed. Res. Int. 2019, 2019, 3851718. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Li, Y.; Sarkar, F.H. Signaling Mechanism(s) of Reactive Oxygen Species in Epithelial-Mesenchymal Transition Reminiscent of Cancer Stem Cells in Tumor Progression. Curr. Stem Cell Res. Ther. 2010, 5, 74–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samarghandian, S.; Farkhondeh, T.; Azimi-Nezhad, M. Protective Effects of Chrysin Against Drugs and Toxic Agents. Dose Response 2017, 15, 1559325817711782. [Google Scholar] [CrossRef] [PubMed]
- Kucukler, S.; Benzer, F.; Yildirim, S.; Gur, C.; Kandemir, F.M.; Bengu, A.S.; Ayna, A.; Caglayan, C.; Dortbudak, M.B. Protective Effects of Chrysin Against Oxidative Stress and Inflammation Induced by Lead Acetate in Rat Kidneys: A Biochemical and Histopathological Approach. Biol. Trace Elem. Res. 2021, 199, 1501–1514. [Google Scholar] [CrossRef] [PubMed]
- Sassi, A.; Boubaker, J.; Loussaief, A.; Jomaa, K.; Ghedira, K.; Chekir-Ghedira, L. Protective Effect of Chrysin, a Dietary Flavone against Genotoxic and Oxidative Damage Induced by Mitomycin C in Balb/C Mice. Nutr. Cancer 2021, 73, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.F.; Kimura, K.; Bernhardt, W.M.; Shrimanker, N.; Akai, Y.; Hohenstein, B.; Saito, Y.; Johnson, R.S.; Kretzler, M.; Cohen, C.D.; et al. Hypoxia Promotes Fibrogenesis in Vivo via HIF-1 Stimulation of Epithelial-to-Mesenchymal Transition. J. Clin. Investig. 2007, 117, 3810–3820. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Ning, X.; Zhang, Y.; Lu, Y.; Nie, Y.; Han, S.; Liu, L.; Du, R.; Xia, L.; He, L.; et al. Hypoxia-Inducible Factor-1α Induces Twist Expression in Tubular Epithelial Cells Subjected to Hypoxia, Leading to Epithelial-to-Mesenchymal Transition. Kidney Int. 2009, 75, 1278–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, B.; Xue, J.; Li, Z.; Shi, X.; Jiang, B.-H.; Fang, J. Chrysin Inhibits Expression of Hypoxia-Inducible Factor-1α through Reducing Hypoxia-Inducible Factor-1α Stability and Inhibiting Its Protein Synthesis. Mol. Cancer Ther. 2007, 6, 220. [Google Scholar] [CrossRef] [Green Version]
- Dieperink, H.; Leyssac, P.P.; Starklint, H.; Kemp, E. Long-Term Cyclosporin Nephrotoxicity in the Rat: Effects on Renal Function and Morphology. Nephrol. Dial. Transplant. 1988, 3, 317–326. [Google Scholar]
- Wang, X.; Morris, M.E. Pharmacokinetics and Bioavailability of the Flavonoid 7,8-Benzoflavone in Rats. J. Pharm. Sci. 2008, 97, 4546–4556. [Google Scholar] [CrossRef]
- Sunilkumar, S.; Ford, S.M. Elevated Glucose Concentration in Culture Media Decreases Membrane Trafficking of SGLT2 in LLC-PK1 Cells via a CAMP/PKA-Dependent Pathway. Am. J. Physiol. Cell Physiol. 2019, 316, C913–C924. [Google Scholar] [CrossRef]
- Denoon, T.; Sunilkumar, S.; Ford, S.M. Acetoacetate Enhances Oxidative Metabolism and Response to Toxicants of Cultured Kidney Cells. Toxicol. Lett. 2020, 323, 19–24. [Google Scholar] [CrossRef]
- Vittal, R.; Horowitz, J.C.; Moore, B.B.; Zhang, H.; Martinez, F.J.; Toews, G.B.; Standiford, T.J.; Thannickal, V.J. Modulation of Prosurvival Signaling in Fibroblasts by a Protein Kinase Inhibitor Protects against Fibrotic Tissue Injury. Am. J. Pathol. 2005, 166, 367–375. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Catalog # | Dilution |
|---|---|---|
| Cell signaling technology | ||
| E-cadherin | 3195 | 1:1000 |
| Smad 3 | 9523 | 1:1000 |
| Phospho-Smad 3 | 9520 | 1:1000 |
| Snail | 3879 | 1:500 |
| TGF-β1 | 3711 | 1:500 |
| Santa Cruz Biotechnology | ||
| β-catenin | sc-7963 | 1:1000 |
| Vimentin | sc-6260 | 1:1000 |
| Goat anti-rabbit IgG-HRP | sc-2004 | 1:10,000 |
| Goat anti-mouse IgG-HRP | sc-2354 | 1:10,000 |
| Invitrogen | ||
| α-SMA | 14-9760-80 | 1:1000 |
| Anti-mouse IgG FITC | F2761 | 1:1000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagavally, R.R.; Sunilkumar, S.; Akhtar, M.; Trombetta, L.D.; Ford, S.M. Chrysin Ameliorates Cyclosporine-A-Induced Renal Fibrosis by Inhibiting TGF-β1-Induced Epithelial–Mesenchymal Transition. Int. J. Mol. Sci. 2021, 22, 10252. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910252
Nagavally RR, Sunilkumar S, Akhtar M, Trombetta LD, Ford SM. Chrysin Ameliorates Cyclosporine-A-Induced Renal Fibrosis by Inhibiting TGF-β1-Induced Epithelial–Mesenchymal Transition. International Journal of Molecular Sciences. 2021; 22(19):10252. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910252
Chicago/Turabian StyleNagavally, Rohan Reddy, Siddharth Sunilkumar, Mumtaz Akhtar, Louis D. Trombetta, and Sue M. Ford. 2021. "Chrysin Ameliorates Cyclosporine-A-Induced Renal Fibrosis by Inhibiting TGF-β1-Induced Epithelial–Mesenchymal Transition" International Journal of Molecular Sciences 22, no. 19: 10252. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910252