
Survivin’ Acute Myeloid Leukaemia—A Personalised Target for inv(16) Patients

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
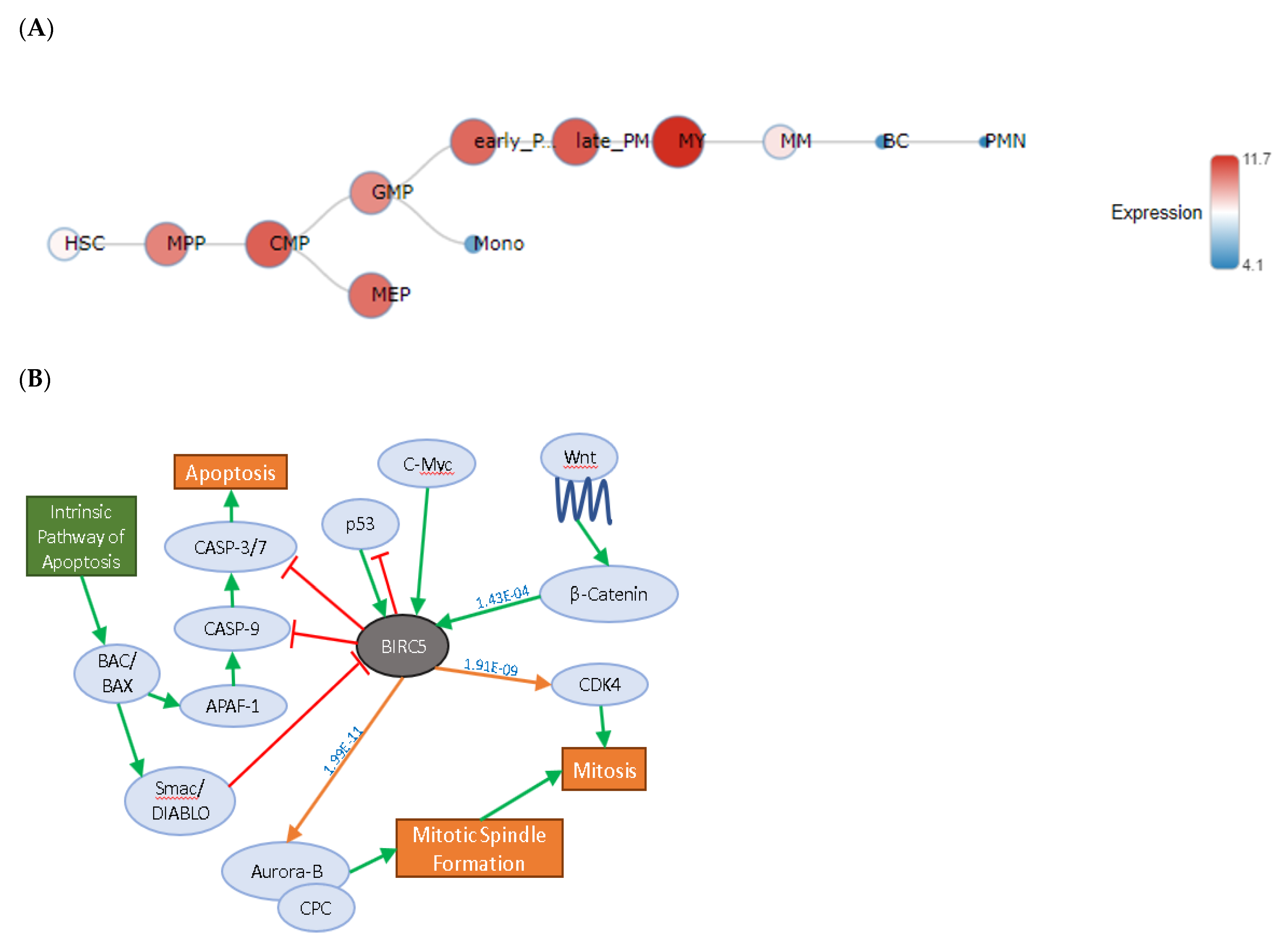
2.1. BIRC5 Expression in Healthy Blood Cells
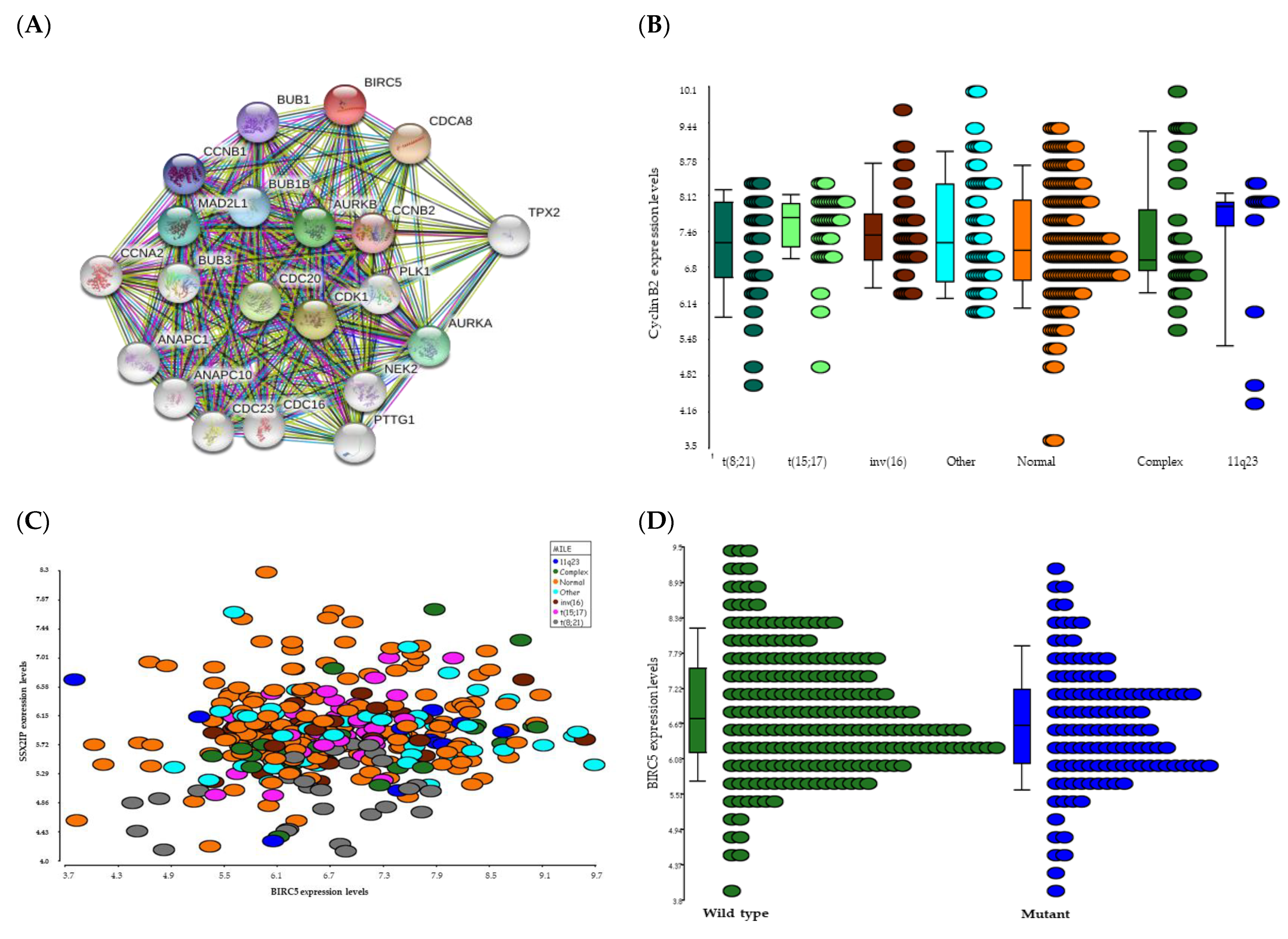
2.2. Pathway Analysis
2.3. Gene Expression Analysis
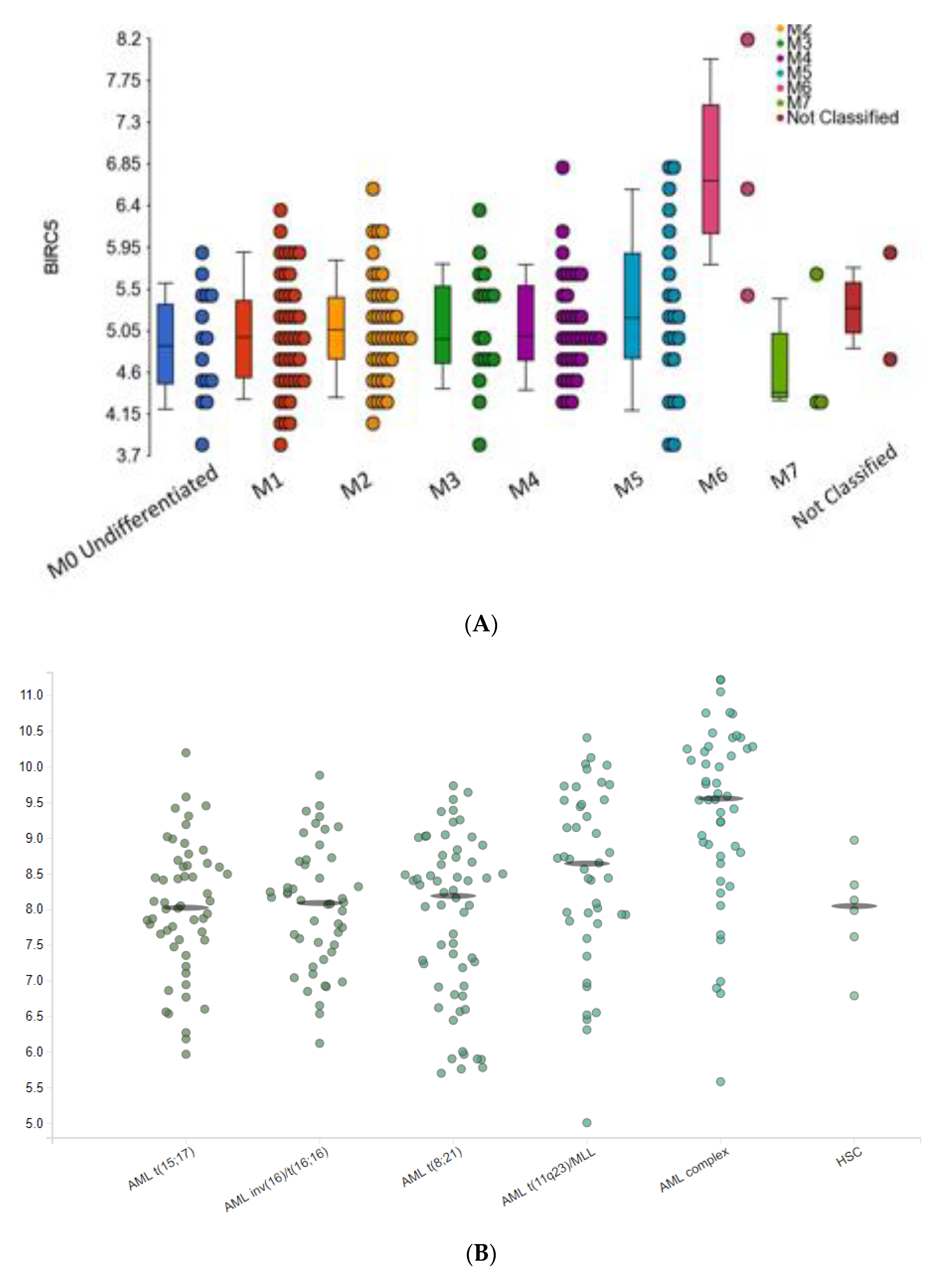
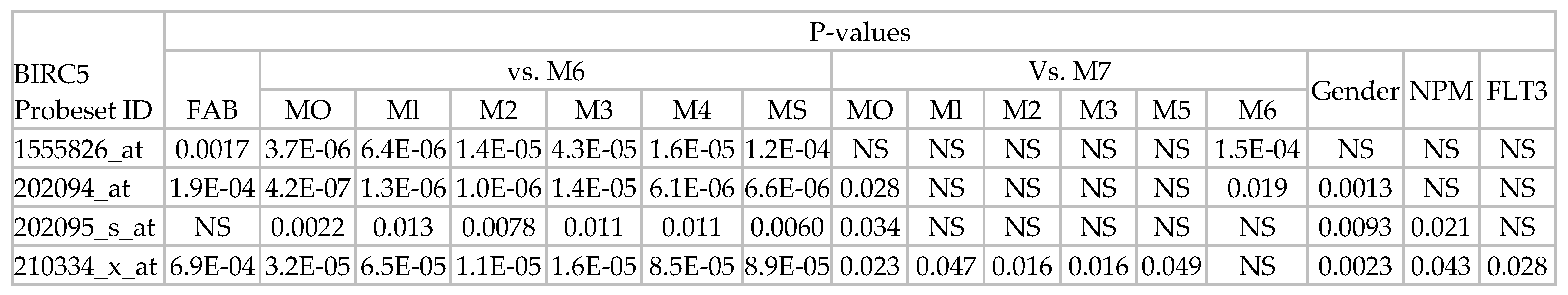
2.3.1. BIRC5 Expression and Clinical Features of AML
2.3.2. BIRC5 Expression Correlates with Poor Outcome Cytogenetics
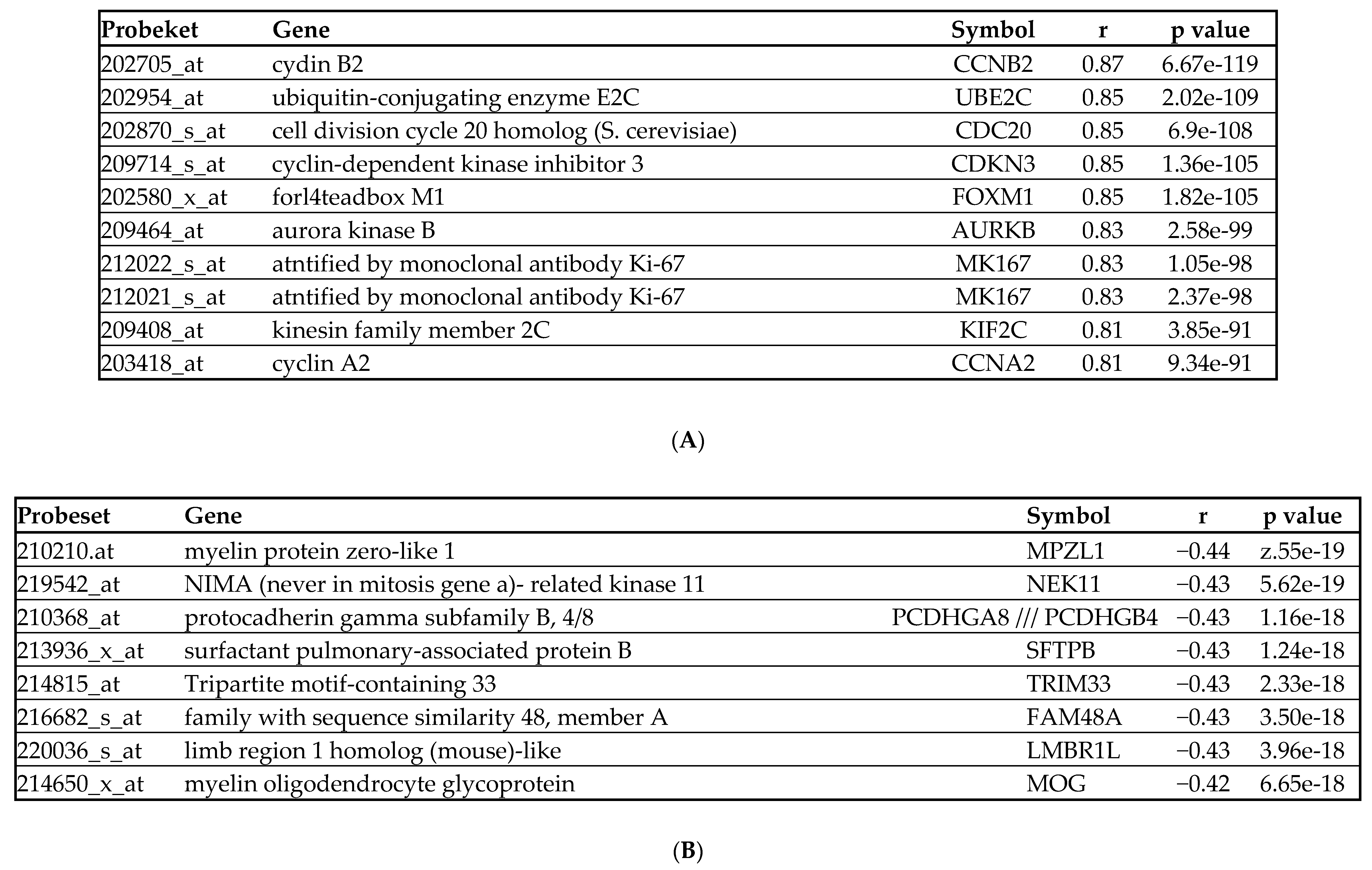
2.3.3. BIRC5 Expression Correlates with Genes Involved in Cell Cycle
2.3.4. BIRC5 Expression Correlates with WT but Not Mutated FLT3
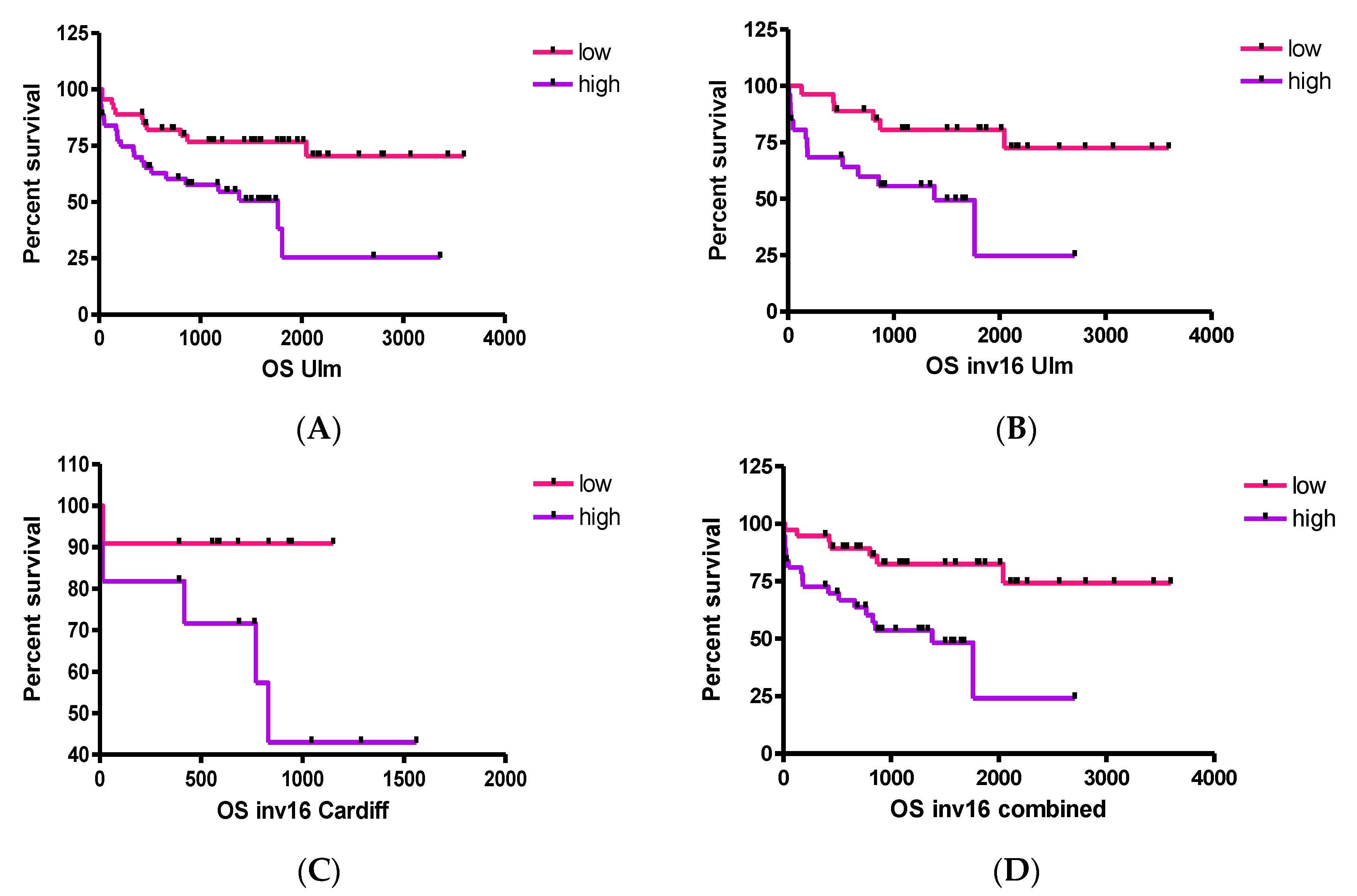
2.3.5. BIRC5 Is Associated with Disease/Relapse Free Survival, but Not OS, in Adults with AML
3. Discussion
4. Materials and Methods
4.1. BIRC5 Expression in Healthy Haematopoietic Cells
4.2. BIRC5 Protein Interaction Analyses
4.3. Association between Genes and Clinical Features
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AML | acute myeloid leukaemia |
| BIRC5 | Baculoviral IAP Repeat Containing 5BM bone marrow |
| CBF | Core Binding Factor |
| CN | Cytogenetically normal |
| CR | Complete remission |
| FAB | French American British |
| FLT3 | FMS-like tyrosine kinase 3 |
| HR | Hazard Ratio |
| IAP | Inhibition of Apoptosis |
| ITD | internal tandem duplication |
| KEGG | Kyoto Encyclopedia of Genes and Genome |
| MILE | Microarray Innovations In Leukemia |
| NPM1 | Nucleophosmin 1 |
| OR | overall remission |
| OS | overall survival |
| RUNX1 | RUNX Family Transcription Factor 1 |
| STRING | search tool for recurring instances of neighbouring genes |
| TCGA | The Cancer Genome Atlas |
| WT | wild type |
| WT1 | Wilm’s Tumour gene 1 |
References
- Shallis, R.M.; Wang, R.; Davidoff, A.; Ma, X.; Zeidan, A.M. Epidemiology of acute myeloid leukemia: Recent progress and enduring challenges. Blood Rev. 2019, 36, 70–87. [Google Scholar] [CrossRef]
- Arber, D.A.; Orazi, A.; Hasserjian, R.; Thiele, J.; Borowitz, M.J.; Le Beau, M.M.; Bloomfield, C.D.; Cazzola, M.; Vardiman, J.W. The 2016 revision to the World Health Organization classification of myeloid neoplasms and acute leukemia. Blood 2016, 127, 2391–2405. [Google Scholar] [CrossRef]
- Nguyen, S.; Leblanc, T.; Fenaux, P.; Witz, F.; Blaise, D.; Pigneux, A.; Thomas, X.; Rigal-Huguet, F.; Lioure, B.; Auvrignon, A.; et al. A white blood cell index as the main prognostic factor in t(8;21) acute myeloid leukemia (AML): A survey of 161 cases from the French AML Intergroup. Blood 2002, 99, 3517–3523. [Google Scholar] [CrossRef] [Green Version]
- Delaunay, J.; Vey, N.; Leblanc, T.; Fenaux, P.; Rigal-Huguet, F.; Witz, F.; Lamy, T.; Auvrignon, A.; Blaise, D.; Pigneux, A.; et al. Prognosis of inv(16)/t(16;16) acute myeloid leukemia (AML): A survey of 110 cases from the French AML Intergroup. Blood 2003, 102, 462–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prada-Arismendy, J.; Arroyave, J.C.; Rothlisberger, S. Molecular biomarkers in acute myeloid leukemia. Blood Rev. 2017, 31, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Stone, R.M.; Mandrekar, S.J.; Sanford, B.L.; Laumann, K.; Geyer, S.; Bloomfield, C.D.; Thiede, C.; Prior, T.W.; Dohner, K.; Marcucci, G.; et al. Midostaurin plus Chemotherapy for Acute Myeloid Leukemia with a FLT3 Mutation. N. Engl. J. Med. 2017, 377, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Daver, N.; Wei, A.H.; Pollyea, D.A.; Fathi, A.T.; Vyas, P.; DiNardo, C.D. New directions for emerging therapies in acute myeloid leukemia: The next chapter. Blood Cancer J. 2020, 10, 107. [Google Scholar] [CrossRef]
- Lagunas-Rangel, F.A.; Chavez-Valencia, V.; Gomez-Guijosa, M.A.; Cortes-Penagos, C. Acute Myeloid Leukemia-Genetic Alterations and Their Clinical Prognosis. Int. J. Hematol. Oncol. Stem Cell Res. 2017, 11, 328–339. [Google Scholar]
- Deol, A.; Sengsayadeth, S.; Ahn, K.W.; Wang, H.L.; Aljurf, M.; Antin, J.H.; Battiwalla, M.; Bornhauser, M.; Cahn, J.Y.; Camitta, B.; et al. Does FLT3 mutation impact survival after hematopoietic stem cell transplantation for acute myeloid leukemia? A Center for International Blood and Marrow Transplant Research (CIBMTR) analysis. Cancer 2016, 122, 3005–3014. [Google Scholar] [CrossRef]
- Atanackovic, D.; Luetkens, T.; Kloth, B.; Fuchs, G.; Cao, Y.; Hildebrandt, Y.; Meyer, S.; Bartels, K.; Reinhard, H.; Lajmi, N.; et al. Cancer-testis antigen expression and its epigenetic modulation in acute myeloid leukemia. Am. J. Hematol. 2011, 86, 918–922. [Google Scholar] [CrossRef]
- Owen, C.; Fitzgibbon, J.; Paschka, P. The clinical relevance of Wilms Tumour 1 (WT1) gene mutations in acute leukaemia. Hematol. Oncol. 2010, 28, 13–19. [Google Scholar] [CrossRef]
- Wheatley, S.P.; Altieri, D.C. Survivin at a glance. J. Cell Sci. 2019, 132, jcs223826. [Google Scholar] [CrossRef] [Green Version]
- Garg, H.; Suri, P.; Gupta, J.C.; Talwar, G.P.; Dubey, S. Survivin: A unique target for tumor therapy. Cancer Cell Int. 2016, 16, 49. [Google Scholar] [CrossRef] [Green Version]
- Oparina, N.; Erlandsson, M.C.; Faldt Beding, A.; Parris, T.; Helou, K.; Karlsson, P.; Einbeigi, Z.; Bokarewa, M.I. Prognostic Significance of BIRC5/Survivin in Breast Cancer: Results from Three Independent Cohorts. Cancers 2021, 13, 2209. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.Q.; Hu, Y.; Xiao, H.J. The prognostic significance of survivin expression in patients with HNSCC: A systematic review and meta-analysis. BMC Cancer 2021, 21, 424. [Google Scholar] [CrossRef] [PubMed]
- Hennigs, J.K.; Minner, S.; Tennstedt, P.; Loser, R.; Huland, H.; Klose, H.; Graefen, M.; Schlomm, T.; Sauter, G.; Bokemeyer, C.; et al. Subcellular Compartmentalization of Survivin is Associated with Biological Aggressiveness and Prognosis in Prostate Cancer. Sci. Rep. 2020, 10, 3250. [Google Scholar] [CrossRef] [PubMed]
- Conde, M.; Michen, S.; Wiedemuth, R.; Klink, B.; Schrock, E.; Schackert, G.; Temme, A. Chromosomal instability induced by increased BIRC5/Survivin levels affects tumorigenicity of glioma cells. BMC Cancer 2017, 17, 889. [Google Scholar] [CrossRef]
- Hoffman, W.H.; Biade, S.; Zilfou, J.T.; Chen, J.; Murphy, M. Transcriptional repression of the anti-apoptotic survivin gene by wild type p53. J. Biol. Chem. 2002, 277, 3247–3257. [Google Scholar] [CrossRef] [Green Version]
- Davis, L.; Mills, K.I.; Orchard, K.H.; Guinn, B.-A. Identification of Genes Whose Expression Overlaps Age Boundaries and Correlates with Risk Groups in Paediatric and Adult Acute Myeloid Leukaemia. Cancers 2020, 12, 2769. [Google Scholar] [CrossRef]
- Bagger, F.O.; Kinalis, S.; Rapin, N. BloodSpot: A database of healthy and malignant haematopoiesis updated with purified and single cell mRNA sequencing profiles. Nucleic Acids Res. 2019, 47, D881–D885. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Mirza, A.; McGuirk, M.; Hockenberry, T.N.; Wu, Q.; Ashar, H.; Black, S.; Wen, S.F.; Wang, L.; Kirschmeier, P.; Bishop, W.R.; et al. Human survivin is negatively regulated by wild-type p53 and participates in p53-dependent apoptotic pathway. Oncogene 2002, 21, 2613–2622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Ambrosini, G.; Chu, E.Y.; Plescia, J.; Tognin, S.; Marchisio, P.C.; Altieri, D.C. Control of apoptosis and mitotic spindle checkpoint by survivin. Nature 1998, 396, 580–584. [Google Scholar] [CrossRef]
- Tamm, I.; Wang, Y.; Sausville, E.; Scudiero, D.A.; Vigna, N.; Oltersdorf, T.; Reed, J.C. IAP-Family Protein Survivin Inhibits Caspase Activity and Apoptosis Induced by Fas (CD95), Bax, Caspases, and Anticancer Drugs. Cancer Res. 1998, 58, 5315–5320. [Google Scholar] [PubMed]
- Shin, S.; Sung, B.-J.; Cho, Y.-S.; Kim, H.-J.; Ha, N.-C.; Hwang, J.-I.; Chung, C.-W.; Jung, Y.-K.; Oh, B.-H. An Anti-apoptotic Protein Human Survivin Is a Direct Inhibitor of Caspase-3 and -7. Biochemistry 2001, 40, 1117–1123. [Google Scholar] [CrossRef]
- Wang, Z.; Fukuda, S.; Pelus, L.M. Survivin regulates the p53 tumor suppressor gene family. Oncogene 2004, 23, 8146–8153. [Google Scholar] [CrossRef] [Green Version]
- Sah, N.K.; Khan, Z.; Khan, G.J.; Bisen, P.S. Structural, functional and therapeutic biology of survivin. Cancer Lett. 2006, 244, 164–171. [Google Scholar] [CrossRef]
- Fang, Z.H.; Dong, C.L.; Chen, Z.; Zhou, B.; Liu, N.; Lan, H.F.; Liang, L.; Liao, W.B.; Zhang, L.; Han, Z.C. Transcriptional regulation of survivin by c-Myc in BCR/ABL-transformed cells: Implications in anti-leukaemic strategy. J. Cell. Mol. Med. 2009, 13, 2039–2052. [Google Scholar] [CrossRef] [PubMed]
- Stewart, D.J. Wnt signaling pathway in non-small cell lung cancer. J. Natl. Cancer Inst. 2014, 106, djt356. [Google Scholar] [CrossRef]
- Suzuki, A.; Hayashida, M.; Ito, T.; Kawano, H.; Nakano, T.; Miura, M.; Akahane, K.; Shiraki, K. Survivin initiates cell cycle entry by the competitive interaction with Cdk4/p16(INK4a) and Cdk2/cyclin E complex activation. Oncogene 2000, 19, 3225–3234. [Google Scholar] [CrossRef] [Green Version]
- Castedo, M.; Perfettini, J.L.; Roumier, T.; Andreau, K.; Medema, R.; Kroemer, G. Cell death by mitotic catastrophe: A molecular definition. Oncogene 2004, 23, 2825–2837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vader, G.; Medema, R.H.; Lens, S.M. The chromosomal passenger complex: Guiding Aurora-B through mitosis. J. Cell Biol. 2006, 173, 833–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Z.; Yao, X.; Wu, M. Direct interaction between survivin and Smac/DIABLO is essential for the anti-apoptotic activity of survivin during taxol-induced apoptosis. J. Biol. Chem. 2003, 278, 23130–23140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haferlach, T.; Kohlmann, A.; Wieczorek, L.; Basso, G.; Kronnie, G.T.; Bene, M.C.; De Vos, J.; Hernandez, J.M.; Hofmann, W.K.; Mills, K.I.; et al. Clinical utility of microarray-based gene expression profiling in the diagnosis and subclassification of leukemia: Report from the International Microarray Innovations in Leukemia Study Group. J. Clin. Oncol. 2010, 28, 2529–2537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinn, B.; Greiner, J.; Schmitt, M.; Mills, K.I. Elevated expression of the leukemia-associated antigen SSX2IP predicts survival in acute myeloid leukemia patients who lack detectable cytogenetic rearrangements. Blood 2009, 113, 1203–1204. [Google Scholar] [CrossRef] [PubMed]
- Guinn, B.A.; Bullinger, L.; Thomas, N.S.; Mills, K.I.; Greiner, J. SSX2IP expression in acute myeloid leukaemia: An association with mitotic spindle failure in t(8;21), and cell cycle in t(15;17) patients. Br. J. Haematol. 2008, 140, 250–251. [Google Scholar] [CrossRef]
- Fukuda, S.; Singh, P.; Moh, A.; Abe, M.; Conway, E.M.; Boswell, H.S.; Yamaguchi, S.; Fu, X.Y.; Pelus, L.M. Survivin mediates aberrant hematopoietic progenitor cell proliferation and acute leukemia in mice induced by internal tandem duplication of Flt3. Blood 2009, 114, 394–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bullinger, L.; Dohner, K.; Bair, E.; Frohling, S.; Schlenk, R.F.; Tibshirani, R.; Dohner, H.; Pollack, J.R. Use of gene-expression profiling to identify prognostic subclasses in adult acute myeloid leukemia. N. Engl. J. Med. 2004, 350, 1605–1616. [Google Scholar] [CrossRef] [Green Version]
- Bullinger, L.; Dohner, K.; Kranz, R.; Stirner, C.; Frohling, S.; Scholl, C.; Kim, Y.H.; Schlenk, R.F.; Tibshirani, R.; Dohner, H.; et al. An FLT3 gene-expression signature predicts clinical outcome in normal karyotype AML. Blood 2008, 111, 4490–4495. [Google Scholar] [CrossRef] [Green Version]
- Bullinger, L.; Rucker, F.G.; Kurz, S.; Du, J.; Scholl, C.; Sander, S.; Corbacioglu, A.; Lottaz, C.; Krauter, J.; Frohling, S.; et al. Gene-expression profiling identifies distinct subclasses of core binding factor acute myeloid leukemia. Blood 2007, 110, 1291–1300. [Google Scholar] [CrossRef] [Green Version]
- Balkhi, M.Y.; Christopeit, M.; Chen, Y.; Geletu, M.; Behre, G. AML1/ETO-induced survivin expression inhibits transcriptional regulation of myeloid differentiation. Exp. Hematol. 2008, 36, 1449–1460. [Google Scholar] [CrossRef]
- Boullosa, L.F.; Savaliya, P.; Bonney, S.; Orchard, L.; Wickenden, H.; Lee, C.; Smits, E.; Banham, A.H.; Mills, K.I.; Orchard, K.; et al. Identification of survivin as a promising target for the immunotherapy of adult B-cell acute lymphoblastic leukemia. Oncotarget 2018, 9, 3853–3866. [Google Scholar] [CrossRef] [Green Version]
- De Luca, M.; Lavia, P.; Guarguaglini, G. A functional interplay between Aurora-A, Plk1 and TPX2 at spindle poles: Plk1 controls centrosomal localization of Aurora-A and TPX2 spindle association. Cell Cycle 2006, 5, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Joukov, V.; Groen, A.C.; Prokhorova, T.; Gerson, R.; White, E.; Rodriguez, A.; Walter, J.C.; Livingston, D.M. The BRCA1/BARD1 heterodimer modulates ran-dependent mitotic spindle assembly. Cell 2006, 127, 539–552. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, C.A.; McCarthy, J.; Turley, E. Cell-surface and mitotic-spindle RHAMM: Moonlighting or dual oncogenic functions? J. Cell Sci. 2008, 121, 925–932. [Google Scholar] [CrossRef] [Green Version]
- Altieri, D.C. Survivin, cancer networks and pathway-directed drug discovery. Nat. Rev. Cancer 2008, 8, 61–70. [Google Scholar] [CrossRef]
- Wu, S.; Su, R.; Jia, H. Cyclin B2 (CCNB2) Stimulates the Proliferation of Triple-Negative Breast Cancer (TNBC) Cells In Vitro and In Vivo. Dis. Markers 2021, 2021, 5511041. [Google Scholar] [CrossRef]
- Sarafan-Vasseur, N.; Lamy, A.; Bourguignon, J.; Le Pessot, F.; Hieter, P.; Sesboue, R.; Bastard, C.; Frebourg, T.; Flaman, J.M. Overexpression of B-type cyclins alters chromosomal segregation. Oncogene 2002, 21, 2051–2057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shubbar, E.; Kovacs, A.; Hajizadeh, S.; Parris, T.Z.; Nemes, S.; Gunnarsdottir, K.; Einbeigi, Z.; Karlsson, P.; Helou, K. Elevated cyclin B2 expression in invasive breast carcinoma is associated with unfavorable clinical outcome. BMC Cancer 2013, 13, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siffroi-Fernandez, S.; Dulong, S.; Li, X.M.; Filipski, E.; Grechez-Cassiau, A.; Peteri-Brunback, B.; Meijer, L.; Levi, F.; Teboul, M.; Delaunay, F. Functional genomics identify Birc5/survivin as a candidate gene involved in the chronotoxicity of cyclin-dependent kinase inhibitors. Cell Cycle 2014, 13, 984–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Bi, C.; Janakakumara, J.V.; Liu, S.C.; Chng, W.J.; Tay, K.G.; Poon, L.F.; Xie, Z.; Palaniyandi, S.; Yu, H.; et al. Enhanced activation of STAT pathways and overexpression of survivin confer resistance to FLT3 inhibitors and could be therapeutic targets in AML. Blood 2009, 113, 4052–4062. [Google Scholar] [CrossRef]
- Yoshida, A.; Ookura, M.; Zokumasu, K.; Ueda, T. Go6976, a FLT3 kinase inhibitor, exerts potent cytotoxic activity against acute leukemia via inhibition of survivin and MCL-1. Biochem. Pharmacol. 2014, 90, 16–24. [Google Scholar] [CrossRef]
- Greiner, J.; Bullinger, L.; Guinn, B.A.; Dohner, H.; Schmitt, M. Leukemia-associated antigens are critical for the proliferation of acute myeloid leukemia cells. Clin. Cancer Res. 2008, 14, 7161–7166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greiner, J.; Li, L.; Ringhoffer, M.; Barth, T.F.; Giannopoulos, K.; Guillaume, P.; Ritter, G.; Wiesneth, M.; Dohner, H.; Schmitt, M. Identification and characterization of epitopes of the receptor for hyaluronic acid-mediated motility (RHAMM/CD168) recognized by CD8+ T cells of HLA-A2-positive patients with acute myeloid leukemia. Blood 2005, 106, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, M.; Schmitt, A.; Rojewski, M.T.; Chen, J.; Giannopoulos, K.; Fei, F.; Yu, Y.; Gotz, M.; Heyduk, M.; Ritter, G.; et al. RHAMM-R3 peptide vaccination in patients with acute myeloid leukemia, myelodysplastic syndrome, and multiple myeloma elicits immunologic and clinical responses. Blood 2008, 111, 1357–1365. [Google Scholar] [CrossRef] [PubMed]
- Greiner, J.; Schmitt, M.; Li, L.; Giannopoulos, K.; Bosch, K.; Schmitt, A.; Dohner, K.; Schlenk, R.F.; Pollack, J.R.; Dohner, H.; et al. Expression of tumor-associated antigens in acute myeloid leukemia: Implications for specific immunotherapeutic approaches. Blood 2006, 108, 4109–4117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Aljahdali, I.; Ling, X. Cancer therapeutics using survivin BIRC5 as a target: What can we do after over two decades of study? J. Exp. Clin. Cancer Res. 2019, 38, 368. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Lyu, H.; Wang, J.; Liu, B. Influence of survivin-targeted therapy on chemosensitivity in the treatment of acute myeloid leukemia. Cancer Lett. 2015, 366, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Shima, H.; Tsurita, G.; Wada, S.; Hirohashi, Y.; Yasui, H.; Hayashi, H.; Miyakoshi, T.; Watanabe, K.; Murai, A.; Asanuma, H.; et al. Randomized phase II trial of survivin 2B peptide vaccination for patients with HLA-A24-positive pancreatic adenocarcinoma. Cancer Sci. 2019, 110, 2378–2385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arber, C.; Feng, X.; Abhyankar, H.; Romero, E.; Wu, M.F.; Heslop, H.E.; Barth, P.; Dotti, G.; Savoldo, B. Survivin-specific T cell receptor targets tumor but not T cells. J. Clin. Investig. 2015, 125, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greiner, J.; Brown, E.; Bullinger, L.; Hills, R.K.; Morris, V.; Döhner, H.; Mills, K.I.; Guinn, B.-a. Survivin’ Acute Myeloid Leukaemia—A Personalised Target for inv(16) Patients. Int. J. Mol. Sci. 2021, 22, 10482. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910482
Greiner J, Brown E, Bullinger L, Hills RK, Morris V, Döhner H, Mills KI, Guinn B-a. Survivin’ Acute Myeloid Leukaemia—A Personalised Target for inv(16) Patients. International Journal of Molecular Sciences. 2021; 22(19):10482. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910482
Chicago/Turabian StyleGreiner, Jochen, Elliott Brown, Lars Bullinger, Robert K. Hills, Vanessa Morris, Hartmut Döhner, Ken I. Mills, and Barbara-ann Guinn. 2021. "Survivin’ Acute Myeloid Leukaemia—A Personalised Target for inv(16) Patients" International Journal of Molecular Sciences 22, no. 19: 10482. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910482