
Cannabinoid Receptor Type 1 Regulates Drug Reward Behavior via Glutamate Decarboxylase 67 Transcription

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. JWH-210 Binds to CB1 In Vivo
2.2. JWH-210 Exhibits CB1 Agonist Activity
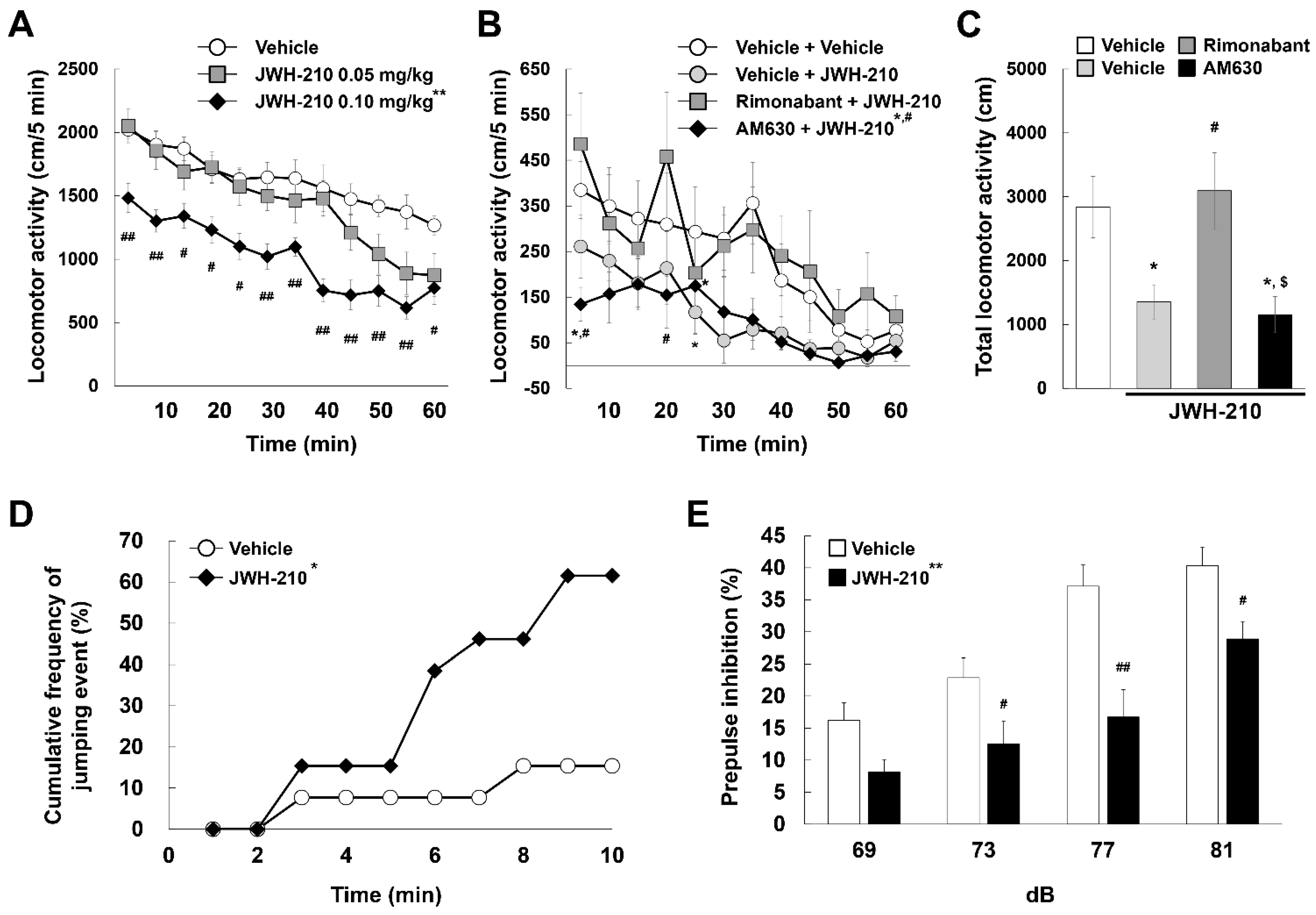
2.3. Five-Day Administration of JWH-210 Affects Impulsivity and Sensorimotor Gating
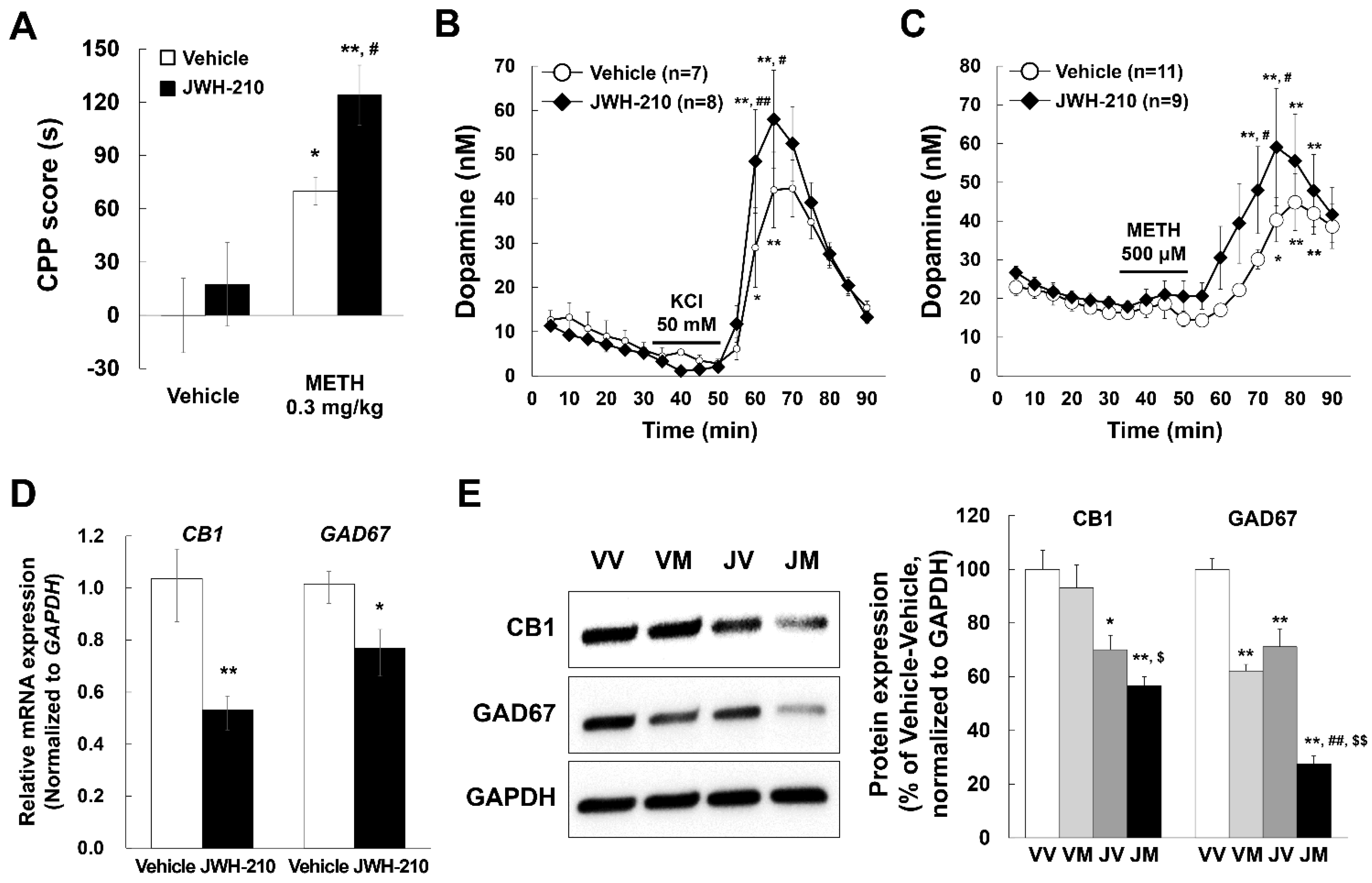
2.4. Pretreatment of JWH-210 Increases METH-Induced CPP
2.5. JWH-210 Treatment Increases KCl-Induced Dopamine Release and METH-Induced Dopamine Efflux
2.6. Administration of JWH-210 Reduced CB1 Expression in Mice Striatum
2.7. CB1 Regulates GAD67 Expression in Cultured Primary Neurons
2.8. CB1 Knockdown Increases METH-Induced CPP
2.9. Apomorphine-Induced Climbing Behavior and METH-Induced CPP Increase with GAD67 Knockdown
2.10. GAD67 Levels in the Human Frontal Cortex Are Significantly Correlated with Drug Use Severity Ratings
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. In Vivo Imaging
4.4. In Vivo CB1 and GAD67 Knockdown
4.5. Locomotor Activity
4.6. Cliff Avoidance Reaction Test
4.7. Prepulse Inhibition Test
4.8. Conditioned Place Preference Test
4.9. Acute Brain Slices and Dopamine Measurement
4.10. Mouse Primary Neuronal Culture
4.11. In Vitro CB1 Knockdown/Overexpression
4.12. Immunocytochemistry/Immunofluorescence
4.13. Quantitative Real-Time PCR (qPCR)
4.14. Western Blotting
4.15. Climbing Behavior
4.16. Immunohistochemistry/Immunofluorescence
4.17. Ethical Approval of the Human Study
4.18. Human RNA-Seq Data
4.19. Correlation Analysis between RNA-Seq Data and the Severity Ratings of Drug Use
4.20. Functional Annotation
4.21. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Azad, S.C.; Eder, M.; Marsicano, G.; Lutz, B.; Zieglgänsberger, W.; Rammes, G. Activation of the cannabinoid receptor type 1 decreases glutamatergic and GABAergic synaptic transmission in the lateral amygdala of the mouse. Learn. Mem. 2003, 10, 116–128. [Google Scholar] [CrossRef] [Green Version]
- Ledent, C.; Valverde, O.; Cossu, G.; Petitet, F.; Aubert, J.F.; Beslot, F.; Böhme, G.A.; Imperato, A.; Pedrazzini, T.; Roques, B.P.; et al. Unresponsiveness to cannabinoids and reduced addictive effects of opiates in CB1 receptor knockout mice. Science 1999, 283, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Bátkai, S.; Kunos, G. The endocannabinoid system as an emerging target of pharmacotherapy. Pharmacol. Rev. 2006, 58, 389–462. [Google Scholar] [CrossRef] [Green Version]
- Zimmer, A.; Zimmer, A.M.; Hohmann, A.G.; Herkenham, M.; Bonner, T.I. Increased mortality, hypoactivity, and hypoalgesia in cannabinoid CB1 receptor knockout mice. Proc. Natl. Acad. Sci. USA 1999, 96, 5780–5785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wise, L.E.; Thorpe, A.J.; Lichtman, A.H. Hippocampal CB(1) receptors mediate the memory impairing effects of Delta(9)-tetrahydrocannabinol. Neuropsychopharmacology 2009, 34, 2072–2080. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Egerton, A.; Kim, E.; Rosso, L.; Riano Barros, D.; Hammers, A.; Brammer, M.; Turkheimer, F.E.; Howes, O.D.; McGuire, P. Acute induction of anxiety in humans by delta-9-tetrahydrocannabinol related to amygdalar cannabinoid-1 (CB1) receptors. Sci. Rep. 2017, 7, 15025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunderson, E.W.; Haughey, H.M.; Ait-Daoud, N.; Joshi, A.S.; Hart, C.L. “Spice” and “K2” herbal highs: A case series and systematic review of the clinical effects and biopsychosocial implications of synthetic cannabinoid use in humans. Am. J. Addict. 2012, 21, 320–326. [Google Scholar] [CrossRef]
- Cohen, K.; Weinstein, A.M. Synthetic and Non-synthetic Cannabinoid Drugs and Their Adverse Effects-A Review from Public Health Prospective. Front. Public Health 2018, 6, 162. [Google Scholar] [CrossRef]
- Mensen, V.T.; Vreeker, A.; Nordgren, J.; Atkinson, A.; de la Torre, R.; Farré, M.; Ramaekers, J.G.; Brunt, T.M. Psychopathological symptoms associated with synthetic cannabinoid use: A comparison with natural cannabis. Psychopharmacology 2019, 236, 2677–2685. [Google Scholar] [CrossRef] [Green Version]
- Clark, P.A.; Capuzzi, K.; Fick, C. Medical marijuana: Medical necessity versus political agenda. Med. Sci. Monit. 2011, 17, Ra249–Ra261. [Google Scholar] [CrossRef] [Green Version]
- Filbey, F.M.; Dunlop, J.; Myers, U.S. Neural effects of positive and negative incentives during marijuana withdrawal. PLoS ONE 2013, 8, e61470. [Google Scholar] [CrossRef] [Green Version]
- Grant, I.; Atkinson, J.H.; Gouaux, B.; Wilsey, B. Medical marijuana: Clearing away the smoke. Open Neurol. J. 2012, 6, 18–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermanns-Clausen, M.; Kneisel, S.; Hutter, M.; Szabo, B.; Auwärter, V. Acute intoxication by synthetic cannabinoids-four case reports. Drug Test. Anal. 2013, 5, 790–794. [Google Scholar] [CrossRef] [PubMed]
- Julian, M.D.; Martin, A.B.; Cuellar, B.; Rodriguez De Fonseca, F.; Navarro, M.; Moratalla, R.; Garcia-Segura, L.M. Neuroanatomical relationship between type 1 cannabinoid receptors and dopaminergic systems in the rat basal ganglia. Neuroscience 2003, 119, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Cheer, J.F.; Wassum, K.M.; Heien, M.L.; Phillips, P.E.; Wightman, R.M. Cannabinoids enhance subsecond dopamine release in the nucleus accumbens of awake rats. J. Neurosci. 2004, 24, 4393–4400. [Google Scholar] [CrossRef] [PubMed]
- French, E.D.; Dillon, K.; Wu, X. Cannabinoids excite dopamine neurons in the ventral tegmentum and substantia nigra. Neuroreport 1997, 8, 649–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oleson, E.B.; Cheer, J.F. A brain on cannabinoids: The role of dopamine release in reward seeking. Cold Spring Harb. Perspect. Med. 2012, 2, a012229. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Hoffman, A.F.; Peng, X.Q.; Lupica, C.R.; Gardner, E.L.; Xi, Z.X. Attenuation of basal and cocaine-enhanced locomotion and nucleus accumbens dopamine in cannabinoid CB1-receptor-knockout mice. Psychopharmacology 2009, 204, 1. [Google Scholar] [CrossRef] [Green Version]
- Panlilio, L.V.; Zanettini, C.; Barnes, C.; Solinas, M.; Goldberg, S.R. Prior exposure to THC increases the addictive effects of nicotine in rats. Neuropsychopharmacology 2013, 38, 1198–1208. [Google Scholar] [CrossRef] [Green Version]
- Szabo, B.; Schlicker, E. Effects of cannabinoids on neurotransmission. Handb. Exp. Pharmacol. 2005, 168, 327–365. [Google Scholar]
- Engler, B.; Freiman, I.; Urbanski, M.; Szabo, B. Effects of exogenous and endogenous cannabinoids on GABAergic neurotransmission between the caudate-putamen and the globus pallidus in the mouse. J. Pharmacol. Exp. Ther. 2006, 316, 608–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katona, I.; Rancz, E.A.; Acsady, L.; Ledent, C.; Mackie, K.; Hajos, N.; Freund, T.F. Distribution of CB1 cannabinoid receptors in the amygdala and their role in the control of GABAergic transmission. J. Neurosci. 2001, 21, 9506–9518. [Google Scholar] [CrossRef] [PubMed]
- Katona, I.; Sperlagh, B.; Sik, A.; Kafalvi, A.; Vizi, E.S.; Mackie, K.; Freund, T.F. Presynaptically located CB1 cannabinoid receptors regulate GABA release from axon terminals of specific hippocampal interneurons. J. Neurosci. 1999, 19, 4544–4558. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, T.; Lauzon, N.M.; de Jaeger, X.; Laviolette, S.R. Cannabinoid transmission in the prelimbic cortex bidirectionally controls opiate reward and aversion signaling through dissociable kappa versus μ-opiate receptor dependent mechanisms. J. Neurosci. 2013, 33, 15642–15651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Blázquez, P.; Rodríguez-Muñoz, M.; Garzón, J. The cannabinoid receptor 1 associates with NMDA receptors to produce glutamatergic hypofunction: Implications in psychosis and schizophrenia. Front. Pharmacol. 2014, 4, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafenêtre, P.; Chaouloff, F.; Marsicano, G. Bidirectional regulation of novelty-induced behavioral inhibition by the endocannabinoid system. Neuropharmacology 2009, 57, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Higuera-Matas, A.; Miguéns, M.; Coria, S.M.; Assis, M.A.; Borcel, E.; del Olmo, N.; Ambrosio, E. Sex-specific disturbances of the glutamate/GABA balance in the hippocampus of adult rats subjected to adolescent cannabinoid exposure. Neuropharmacology 2012, 62, 1975–1984. [Google Scholar] [CrossRef]
- Sieghart, W. Structure, pharmacology, and function of GABAA receptor subtypes. Adv. Pharmacol. 2006, 54, 231–263. [Google Scholar]
- Beveridge, T.J.; Smith, H.R.; Porrino, L.J. Differential development of tolerance to the functional and behavioral effects of repeated baclofen treatment in rats. Pharmacol. Biochem. Behav. 2013, 106, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Pistis, M.; Ferraro, L.; Pira, L.; Flore, G.; Tanganelli, S.; Gessa, G.L.; Devoto, P. Delta(9)-tetrahydrocannabinol decreases extracellular GABA and increases extracellular glutamate and dopamine levels in the rat prefrontal cortex: An in vivo microdialysis study. Brain Res. 2002, 948, 155–158. [Google Scholar] [CrossRef]
- Fattore, L.; Fratta, W. Beyond THC: The New Generation of Cannabinoid Designer Drugs. Front. Behav. Neurosci. 2011, 5, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spielmann, H.; Muller, L.; Averbeck, D.; Balls, M.; Brendler-Schwaab, S.; Castell, J.V.; Curren, R.; de Silva, O.; Gibbs, N.K.; Liebsch, M.; et al. The second ECVAM workshop on phototoxicity testing. The report and recommendations of ECVAM workshop 42. Altern. Lab. Anim. 2000, 28, 777–814. [Google Scholar] [CrossRef]
- Cha, H.J.; Song, Y.J.; Lee, D.E.; Kim, Y.H.; Shin, J.; Jang, C.G.; Suh, S.K.; Kim, S.J.; Yun, J. Receptor Binding Affinities of Synthetic Cannabinoids Determined by Non-Isotopic Receptor Binding Assay. Toxicol. Res. 2019, 35, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Cha, H.J.; Lee, K.W.; Song, M.J.; Hyeon, Y.J.; Hwang, J.Y.; Jang, C.G.; Ahn, J.I.; Jeon, S.H.; Kim, H.U.; Kim, Y.H.; et al. Dependence Potential of the Synthetic Cannabinoids JWH-073, JWH-081, and JWH-210: In Vivo and In Vitro Approaches. Biomol. Ther. 2014, 22, 363–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruno, A.; Lembo, F.; Novellino, E.; Stornaiuolo, M.; Marinelli, L. Beyond radio-displacement techniques for identification of CB1 ligands: The first application of a fluorescence-quenching assay. Sci. Rep. 2014, 4, 3757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, J.; Nagai, T.; Furukawa-Hibi, Y.; Kuroda, K.; Kaibuchi, K.; Greenberg, M.E.; Yamada, K. Neuronal Per Arnt Sim (PAS) domain protein 4 (NPAS4) regulates neurite outgrowth and phosphorylation of synapsin I. J. Biol. Chem. 2013, 288, 2655–2664. [Google Scholar] [CrossRef] [Green Version]
- Niwa, M.; Nabeshima, T. Roles of a novel molecule ‘shati’ in the development of methamphetamine-induced dependence. Curr. Neuropharmacol. 2011, 9, 104–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yates, J.R.; Marusich, J.A.; Gipson, C.D.; Beckmann, J.S.; Bardo, M.T. High impulsivity in rats predicts amphetamine conditioned place preference. Pharmacol. Biochem. Behav. 2012, 100, 370–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiland, B.J.; Heitzeg, M.M.; Zald, D.; Cummiford, C.; Love, T.; Zucker, R.A.; Zubieta, J.K. Relationship between impulsivity, prefrontal anticipatory activation, and striatal dopamine release during rewarded task performance. Psychiatry Res. 2014, 223, 244–252. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Jang, C.G.; Park, W.K. Inhibition by MK-801 of morphine-induced conditioned place preference and postsynaptic dopamine receptor supersensitivity in mice. Pharmacol. Biochem. Behav. 1996, 55, 11–17. [Google Scholar] [CrossRef]
- Smith, J.W.; Fetsko, L.A.; Xu, R.; Wang, Y. Dopamine D2L receptor knockout mice display deficits in positive and negative reinforcing properties of morphine and in avoidance learning. Neuroscience 2002, 113, 755–765. [Google Scholar] [CrossRef]
- Oh, J.H.; Hwang, J.Y.; Hong, S.I.; Ma, S.X.; Seo, J.Y.; Lee, S.Y.; Kim, H.C.; Jang, C.G. The new designer drug buphedrone produces rewarding properties via dopamine D1 receptor activation. Addict. Biol. 2018, 23, 69–79. [Google Scholar] [CrossRef]
- Baker, D.A.; Fuchs, R.A.; Specio, S.E.; Khroyan, T.V.; Neisewander, J.L. Effects of intraaccumbens administration of SCH-23390 on cocaine-induced locomotion and conditioned place preference. Synapse 1998, 30, 181–193. [Google Scholar] [CrossRef]
- Tzschentke, T.M. Measuring reward with the conditioned place preference paradigm: A comprehensive review of drug effects, recent progress and new issues. Prog. Neurobiol. 1998, 56, 613–672. [Google Scholar] [CrossRef]
- Cadoni, C.; Pisanu, A.; Solinas, M.; Acquas, E.; Di Chiara, G. Behavioural sensitization after repeated exposure to Delta 9-tetrahydrocannabinol and cross-sensitization with morphine. Psychopharmacology 2001, 158, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Grinspoon, L.; Bakalar, J.B.; Zimmer, L.; Morgan, J.P. Marijuana addiction. Science 1997, 277, 749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wise, R.A. Dopamine, learning and motivation. Nat. Rev. Neurosci. 2004, 5, 483–494. [Google Scholar] [CrossRef]
- Hyman, S.E.; Malenka, R.C.; Nestler, E.J. Neural mechanisms of addiction: The role of reward-related learning and memory. Ann. Rev. Neurosci. 2006, 29, 565–598. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Kim, Y.; Do, W.; Shin, J.; Cha, H.; Kim, H.; Lee, K.; Song, M. A Study on Neurotoxicity and Psychological Dependence of Synthetic Cannabinoids; Annual Report; National Institute of Food and Drug Safety Evaluation: Osong, Korea, 2013; pp. 1–128.
- Smith, C.T.; San Juan, M.D.; Dang, L.C.; Katz, D.T.; Perkins, S.F.; Burgess, L.L.; Cowan, R.L.; Manning, H.C.; Nickels, M.L.; Claassen, D.O.; et al. Ventral striatal dopamine transporter availability is associated with lower trait motor impulsivity in healthy adults. Transl. Psychiatry 2018, 8, 269. [Google Scholar] [CrossRef]
- Perry, J.L.; Carroll, M.E. The role of impulsive behavior in drug abuse. Psychopharmacology 2008, 200, 1–26. [Google Scholar] [CrossRef]
- Alexander, S.P.; Mathie, A.; Peters, J.A. Guide to Receptors and Channels (GRAC), 3rd edition. Br. J. Pharmacol. 2008, 153 (Suppl. S2), S1–S209. [Google Scholar] [CrossRef] [Green Version]
- Atwood, B.K.; Lee, D.; Straiker, A.; Widlanski, T.S.; Mackie, K. CP47,497-C8 and JWH073, commonly found in ‘Spice’ herbal blends, are potent and efficacious CB(1) cannabinoid receptor agonists. Eur. J. Pharmacol. 2011, 659, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Zamberletti, E.; Beggiato, S.; Steardo, L., Jr.; Prini, P.; Antonelli, T.; Ferraro, L.; Rubino, T.; Parolaro, D. Alterations of prefrontal cortex GABAergic transmission in the complex psychotic-like phenotype induced by adolescent delta-9-tetrahydrocannabinol exposure in rats. Neurobiol. Dis. 2014, 63, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Eggan, S.M.; Lazarus, M.S.; Stoyak, S.R.; Volk, D.W.; Glausier, J.R.; Huang, Z.J.; Lewis, D.A. Cortical glutamic acid decarboxylase 67 deficiency results in lower cannabinoid 1 receptor messenger RNA expression: Implications for schizophrenia. Biol. Psychiatry 2012, 71, 114–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Zessen, R.; Phillips, J.L.; Budygin, E.A.; Stuber, G.D. Activation of VTA GABA neurons disrupts reward consumption. Neuron 2012, 73, 1184–1194. [Google Scholar] [CrossRef] [Green Version]
- Thompson Ray, M.; Weickert, C.S.; Wyatt, E.; Webster, M.J. Decreased BDNF, trkB-TK+ and GAD67 mRNA expression in the hippocampus of individuals with schizophrenia and mood disorders. J. Psychiatry Neurosci. 2011, 36, 195–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straub, R.E.; Lipska, B.K.; Egan, M.F.; Goldberg, T.E.; Callicott, J.H.; Mayhew, M.B.; Vakkalanka, R.K.; Kolachana, B.S.; Kleinman, J.E.; Weinberger, D.R. Allelic variation in GAD1 (GAD67) is associated with schizophrenia and influences cortical function and gene expression. Mol. Psychiatry 2007, 12, 854–869. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Chen, T.; Zhong, N.; Jiang, H.; Du, J.; Xiao, K.; Xu, D.; Wang, Z.; Zhao, M. Gamma-aminobutyric acid and glutamate/glutamine alterations of the left prefrontal cortex in individuals with methamphetamine use disorder: A combined transcranial magnetic stimulation-magnetic resonance spectroscopy study. Ann. Transl. Med. 2020, 8, 347. [Google Scholar] [CrossRef]
- Haddar, M.; Uno, K.; Azuma, K.; Muramatsu, S.-I.; Nitta, A. Inhibitory effects of Shati/Nat8l overexpression in the medial prefrontal cortex on methamphetamine-induced conditioned place preference in mice. Addict. Biol. 2020, 25, e12749. [Google Scholar] [CrossRef] [Green Version]
- Dupuis, J.P.; Bioulac, B.H.; Baufreton, J. Long-term depression at distinct glutamatergic synapses in the basal ganglia. Rev. Neurosci. 2014, 25, 741–754. [Google Scholar] [CrossRef] [Green Version]
- Yun, J.; Gu, S.M.; Yun, H.M.; Son, D.J.; Park, M.H.; Lee, M.S.; Hong, J.T. Myelin oligodendrocyte glycoprotein (MOG35-55)-induced experimental autoimmune encephalomyelitis is ameliorated in interleukin-32 alpha transgenic mice. Oncotarget 2015, 6, 40452–40463. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.M.; Lee, H.P.; Ham, Y.W.; Son, D.J.; Kim, H.Y.; Oh, K.W.; Han, S.B.; Yun, J.; Hong, J.T. Piperlongumine Improves Lipopolysaccharide-Induced Amyloidogenesis by Suppressing NF-KappaB Pathway. Neuromol. Med. 2018, 20, 312–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, S.M.; Yun, J.; Son, D.J.; Kim, H.Y.; Nam, K.T.; Kim, H.D.; Choi, M.G.; Choi, J.S.; Kim, Y.M.; Han, S.B.; et al. Piperlongumine attenuates experimental autoimmune encephalomyelitis through inhibition of NF-kappaB activity. Free Radic. Biol. Med. 2017, 103, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Jo, Y.; Webster, M.J.; Lee, D. Shared co-expression networks in frontal cortex of the normal aged brain and schizophrenia. Schizophr. Res. 2019, 204, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Torrey, E.F.; Webster, M.; Knable, M.; Johnston, N.; Yolken, R.H. The stanley foundation brain collection and neuropathology consortium. Schizophr. Res. 2000, 44, 151–155. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, S.M.; Seo, S.; Park, D.; Kim, S.; Lamichhane, S.; Han, K.-M.; Kim, Y.-H.; Lee, S.; Hong, J.T.; Cha, H.J.; et al. Cannabinoid Receptor Type 1 Regulates Drug Reward Behavior via Glutamate Decarboxylase 67 Transcription. Int. J. Mol. Sci. 2021, 22, 10486. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910486
Gu SM, Seo S, Park D, Kim S, Lamichhane S, Han K-M, Kim Y-H, Lee S, Hong JT, Cha HJ, et al. Cannabinoid Receptor Type 1 Regulates Drug Reward Behavior via Glutamate Decarboxylase 67 Transcription. International Journal of Molecular Sciences. 2021; 22(19):10486. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910486
Chicago/Turabian StyleGu, Sun Mi, Sowoon Seo, Daejin Park, Sanghyeon Kim, Santosh Lamichhane, Kyoung-Moon Han, Young-Hoon Kim, Sangmin Lee, Jin Tae Hong, Hye Jin Cha, and et al. 2021. "Cannabinoid Receptor Type 1 Regulates Drug Reward Behavior via Glutamate Decarboxylase 67 Transcription" International Journal of Molecular Sciences 22, no. 19: 10486. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910486