
Age-Related Changes in the Matrisome of the Mouse Skeletal Muscle

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Identification of Proteins in the Gastrocnemius Muscle
2.2. Matrisome Quantification: Old SM vs. Adult SM
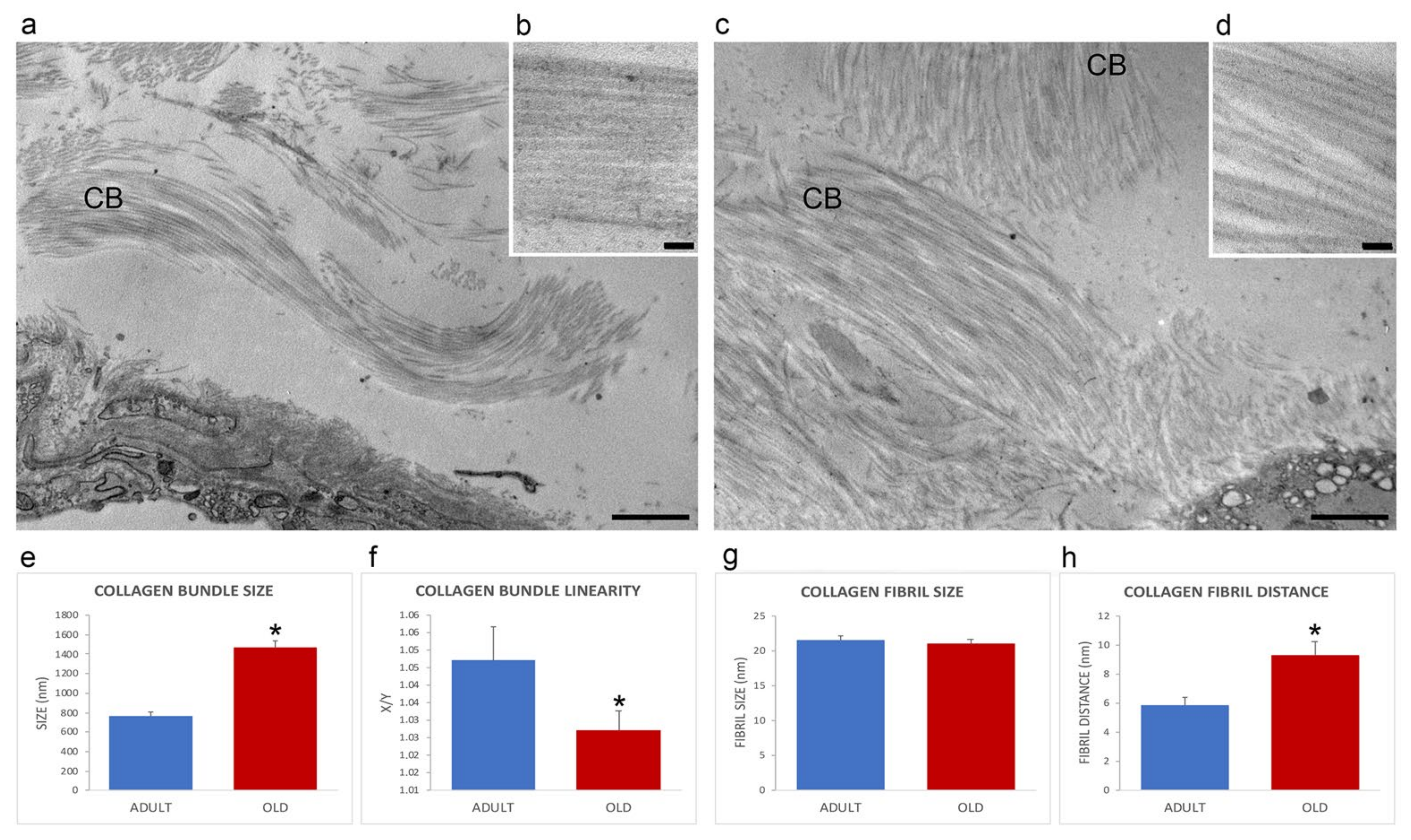
2.3. Ultrastructural Morphology and Morphometrical Evaluation
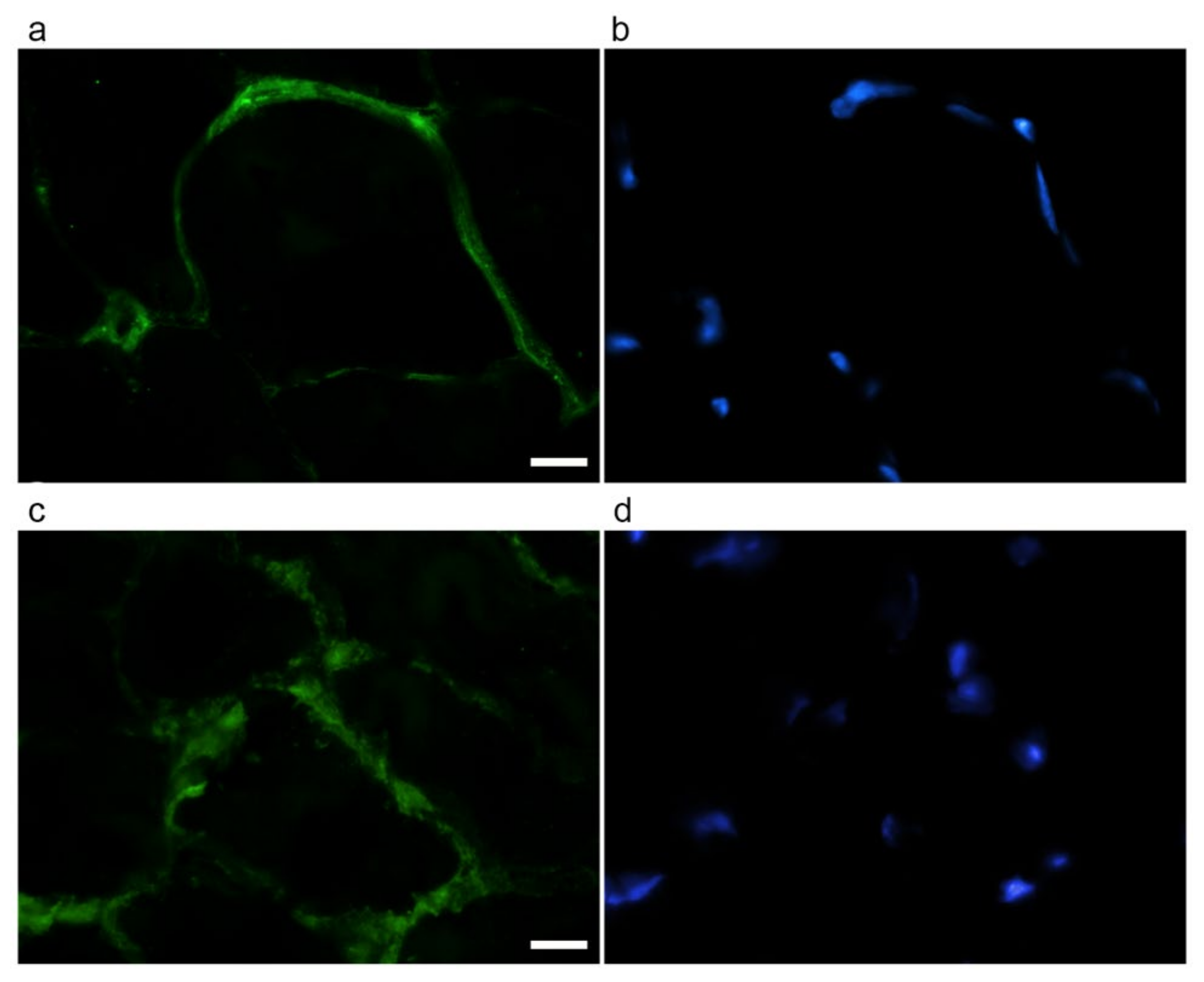
2.4. Evaluation of ECM Components by Immunofluorescence
3. Discussion
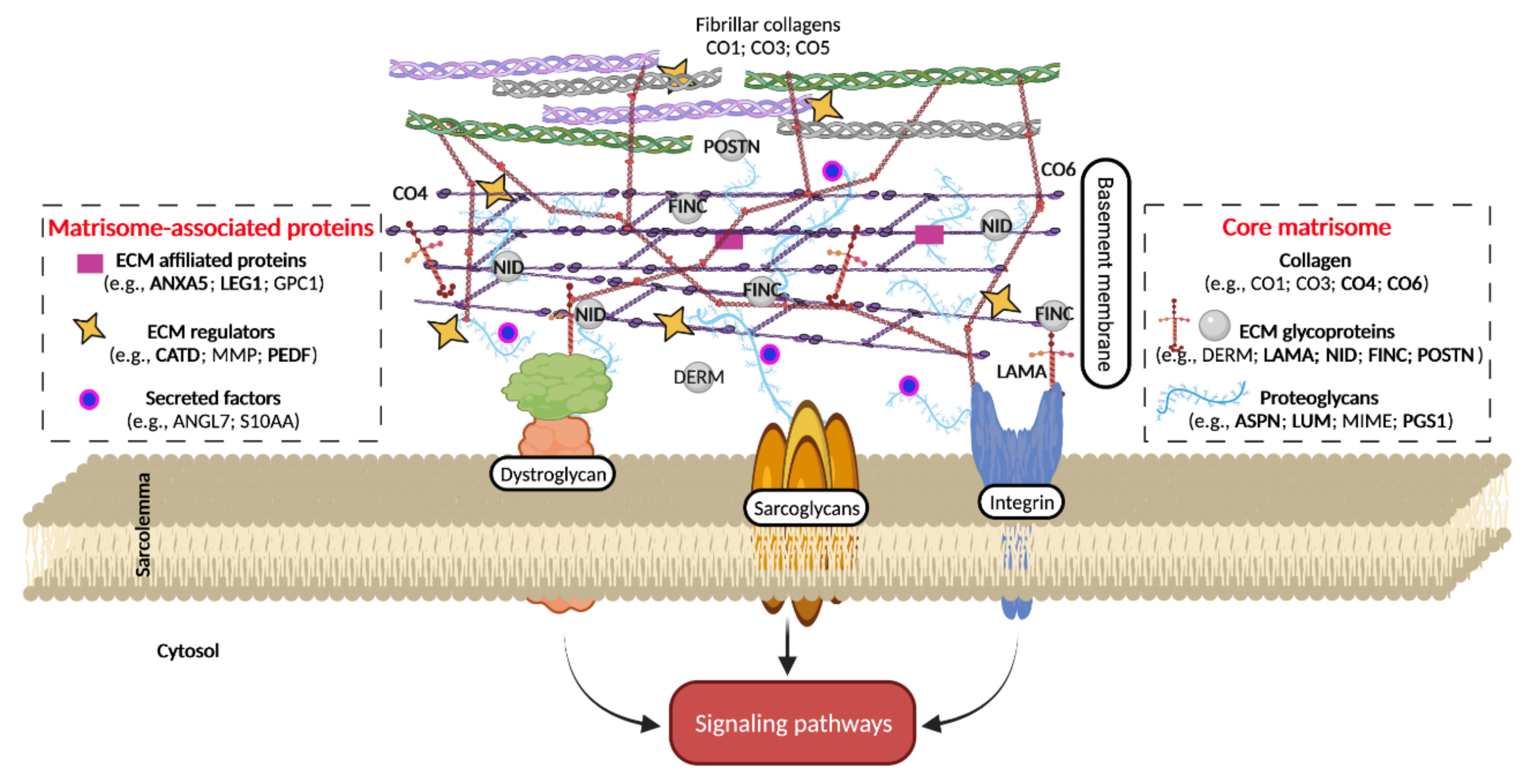
3.1. Alteration of Core Matrisome in Aged vs. Adult Mice
3.2. Alteration of Matrisome-Associated Proteins in Aged vs. Adult Mice
4. Materials and Methods
4.1. Reagents
4.2. Mice
4.3. Proteomic Analysis
4.3.1. Preparation of Protein Samples for Proteomic Analysis
4.3.2. Liquid Chromatography with Tandem Mass Spectrometry (LC-MS/MS)
4.3.3. Data Processing for Protein Identification and Quantification
4.4. Bioinformatic Analysis
4.5. ECM Component Evaluation by Immunofluorescence
4.6. Ultrastructural Morphological and Morphometrical Evaluation
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rowland, L.A.; Bal, N.C.; Periasamy, M. The Role of Skeletal-Muscle-Based Thermogenic Mechanisms in Vertebrate Endothermy. Biol. Rev. Camb. Philos. Soc. 2015, 90, 1279–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskin, K.K.; Winders, B.R.; Olson, E.N. Muscle as a “Mediator” of Systemic Metabolism. Cell Metab. 2015, 21, 237–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz-Jentoft, A.J.; Sayer, A.A. Sarcopenia. Lancet 2019, 393, 2636–2646. [Google Scholar] [CrossRef]
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyère, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A.; et al. Sarcopenia: Revised European Consensus on Definition and Diagnosis. Age Ageing 2019, 48, 16–31. [Google Scholar] [CrossRef] [Green Version]
- Marty, E.; Liu, Y.; Samuel, A.; Or, O.; Lane, J. A Review of Sarcopenia: Enhancing Awareness of an Increasingly Prevalent Disease. Bone 2017, 105, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Walston, J.D. Sarcopenia in Older Adults. Curr. Opin. Rheumatol. 2012, 24, 623–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etienne, J.; Liu, C.; Skinner, C.M.; Conboy, M.J.; Conboy, I.M. Skeletal Muscle as an Experimental Model of Choice to Study Tissue Aging and Rejuvenation. Skelet. Muscle 2020, 10, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domingues-Faria, C.; Vasson, M.-P.; Goncalves-Mendes, N.; Boirie, Y.; Walrand, S. Skeletal Muscle Regeneration and Impact of Aging and Nutrition. Ageing Res. Rev. 2016, 26, 22–36. [Google Scholar] [CrossRef]
- Melouane, A.; Yoshioka, M.; St-Amand, J. Extracellular Matrix/Mitochondria Pathway: A Novel Potential Target for Sarcopenia. Mitochondrion 2020, 50, 63–70. [Google Scholar] [CrossRef]
- Cade, W.T.; Yarasheski, K.E. Metabolic and Molecular Aspects of Sarcopenia. In Principles of Molecular Medicine; Runge, M.S., Patterson, C., Eds.; Humana Press: Totowa, NJ, USA, 2006; pp. 529–534. ISBN 978-1-59259-963-9. [Google Scholar]
- McCormick, R.; Vasilaki, A. Age-Related Changes in Skeletal Muscle: Changes to Life-Style as a Therapy. Biogerontology 2018, 19, 519–536. [Google Scholar] [CrossRef] [Green Version]
- Thomas, K.; Engler, A.J.; Meyer, G.A. Extracellular Matrix Regulation in the Muscle Satellite Cell Niche. Connect. Tissue Res. 2015, 56, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Liu, Y.; Zhang, H. Extracellular Matrix: An Important Regulator of Cell Functions and Skeletal Muscle Development. Cell Biosci. 2021, 11, 65. [Google Scholar] [CrossRef] [PubMed]
- Street, S.F. Lateral Transmission of Tension in Frog Myofibers: A Myofibrillar Network and Transverse Cytoskeletal Connections Are Possible Transmitters. J. Cell Physiol. 1983, 114, 346–364. [Google Scholar] [CrossRef] [PubMed]
- Thorsteinsdóttir, S.; Deries, M.; Cachaço, A.S.; Bajanca, F. The Extracellular Matrix Dimension of Skeletal Muscle Development. Dev. Biol. 2011, 354, 191–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calve, S.; Odelberg, S.J.; Simon, H.-G. A Transitional Extracellular Matrix Instructs Cell Behavior during Muscle Regeneration. Dev. Biol. 2010, 344, 259–271. [Google Scholar] [CrossRef] [Green Version]
- Kragstrup, T.W.; Kjaer, M.; Mackey, A.L. Structural, Biochemical, Cellular, and Functional Changes in Skeletal Muscle Extracellular Matrix with Aging. Scand. J. Med. Sci. Sports 2011, 21, 749–757. [Google Scholar] [CrossRef] [Green Version]
- Trensz, F.; Lucien, F.; Couture, V.; Söllrald, T.; Drouin, G.; Rouleau, A.-J.; Grandbois, M.; Lacraz, G.; Grenier, G. Increased Microenvironment Stiffness in Damaged Myofibers Promotes Myogenic Progenitor Cell Proliferation. Skelet. Muscle 2015, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Stearns-Reider, K.M.; D’Amore, A.; Beezhold, K.; Rothrauff, B.; Cavalli, L.; Wagner, W.R.; Vorp, D.A.; Tsamis, A.; Shinde, S.; Zhang, C.; et al. Aging of the Skeletal Muscle Extracellular Matrix Drives a Stem Cell Fibrogenic Conversion. Aging Cell 2017, 16, 518–528. [Google Scholar] [CrossRef] [Green Version]
- Urciuolo, A.; Quarta, M.; Morbidoni, V.; Gattazzo, F.; Molon, S.; Grumati, P.; Montemurro, F.; Tedesco, F.S.; Blaauw, B.; Cossu, G.; et al. Collagen VI Regulates Satellite Cell Self-Renewal and Muscle Regeneration. Nat. Commun. 2013, 4, 1964. [Google Scholar] [CrossRef] [Green Version]
- Højlund, K.; Yi, Z.; Hwang, H.; Bowen, B.; Lefort, N.; Flynn, C.R.; Langlais, P.; Weintraub, S.T.; Mandarino, L.J. Characterization of the Human Skeletal Muscle Proteome by One-Dimensional Gel Electrophoresis and HPLC-ESI-MS/MS. Mol. Cell Proteom. 2008, 7, 257–267. [Google Scholar] [CrossRef] [Green Version]
- Lang, F.; Aravamudhan, S.; Nolte, H.; Türk, C.; Hölper, S.; Müller, S.; Günther, S.; Blaauw, B.; Braun, T.; Krüger, M. Dynamic Changes in the Mouse Skeletal Muscle Proteome during Denervation-Induced Atrophy. Dis. Model. Mech. 2017, 10, 881–896. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, A.S.; Murgia, M.; Nagaraj, N.; Treebak, J.T.; Cox, J.; Mann, M. Deep Proteomics of Mouse Skeletal Muscle Enables Quantitation of Protein Isoforms, Metabolic Pathways, and Transcription Factors. Mol. Cell Proteom. 2015, 14, 841–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ubaida-Mohien, C.; Lyashkov, A.; Gonzalez-Freire, M.; Tharakan, R.; Shardell, M.; Moaddel, R.; Semba, R.D.; Chia, C.W.; Gorospe, M.; Sen, R.; et al. Discovery Proteomics in Aging Human Skeletal Muscle Finds Change in Spliceosome, Immunity, Proteostasis and Mitochondria. eLife 2019, 8, e49874. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, K.R.; Lipp, S.; Acuna, A.; Leng, Y.; Bu, Y.; Calve, S. Comparative Analysis of the Extracellular Matrix Proteome across the Myotendinous Junction. J. Proteome Res. 2020, 19, 3955–3967. [Google Scholar] [CrossRef] [PubMed]
- Malatesta, M.; Fattoretti, P.; Giagnacovo, M.; Pellicciari, C.; Zancanaro, C. Physical Training Modulates Structural and Functional Features of Cell Nuclei in Type II Myofibers of Old Mice. Rejuvenation Res. 2011, 14, 543–552. [Google Scholar] [CrossRef]
- Zancanaro, C.; Mariotti, R.; Perdoni, F.; Nicolato, E.; Malatesta, M. Physical Training Is Associated with Changes in Nuclear Magnetic Resonance and Morphometrical Parameters of the Skeletal Muscle in Senescent Mice. Eur. J. Histochem. 2007, 51, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Costanzo, M.; Cisterna, B.; Malatesta, M. Effect of Physical Exercise on the Ultrastructural Features of Skeletal Muscle Mitochondria in Old Mice. Microscopie 2013, 20, 37–43. [Google Scholar] [CrossRef]
- Cisterna, B.; Giagnacovo, M.; Costanzo, M.; Fattoretti, P.; Zancanaro, C.; Pellicciari, C.; Malatesta, M. Adapted Physical Exercise Enhances Activation and Differentiation Potential of Satellite Cells in the Skeletal Muscle of Old Mice. J. Anat. 2016, 228, 771–783. [Google Scholar] [CrossRef]
- Malatesta, M.; Giagnacovo, M.; Cardani, R.; Meola, G.; Pellicciari, C. RNA Processing Is Altered in Skeletal Muscle Nuclei of Patients Affected by Myotonic Dystrophy. Histochem. Cell Biol. 2011, 135, 419–425. [Google Scholar] [CrossRef]
- Malva, A.D.; Albenzio, M.; Santillo, A.; Russo, D.; Figliola, L.; Caroprese, M.; Marino, R. Methods for Extraction of Muscle Proteins from Meat and Fish Using Denaturing and Nondenaturing Solutions. J. Food Qual. 2018, 2018, e8478471. [Google Scholar] [CrossRef]
- Wilson, R.; Diseberg, A.F.; Gordon, L.; Zivkovic, S.; Tatarczuch, L.; Mackie, E.J.; Gorman, J.J.; Bateman, J.F. Comprehensive Profiling of Cartilage Extracellular Matrix Formation and Maturation Using Sequential Extraction and Label-Free Quantitative Proteomics. Mol. Cell Proteom. 2010, 9, 1296–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, N.; Taniguchi, T.; Goda, Y.; Kosaka, H.; Higashino, K.; Sakai, T.; Katoh, S.; Yasui, N.; Sairyo, K.; Taniguchi, H. Proteomic Analysis of Human Tendon and Ligament: Solubilization and Analysis of Insoluble Extracellular Matrix in Connective Tissues. J. Proteome Res. 2016, 15, 4709–4721. [Google Scholar] [CrossRef] [PubMed]
- Naba, A.; Clauser, K.R.; Ding, H.; Whittaker, C.A.; Carr, S.A.; Hynes, R.O. The Extracellular Matrix: Tools and Insights for the “Omics” Era. Matrix Biol. 2016, 49, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Taha, I.N.; Clauser, K.R.; Gao, Y.T.; Naba, A. MatrisomeDB: The ECM-Protein Knowledge Database. Nucleic Acids Res. 2020, 48, D1136–D1144. [Google Scholar] [CrossRef] [PubMed]
- Théron, L.; Gueugneau, M.; Coudy, C.; Viala, D.; Bijlsma, A.; Butler-Browne, G.; Maier, A.; Béchet, D.; Chambon, C. Label-Free Quantitative Protein Profiling of Vastus Lateralis Muscle during Human Aging. Mol. Cell Proteom. 2014, 13, 283–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, T.-H.; Choi, M.; Banfai, B.; Liu, Y.; MacLean, B.X.; Dunkley, T.; Vitek, O. Selection of Features with Consistent Profiles Improves Relative Protein Quantification in Mass Spectrometry Experiments. Mol. Cell Proteom. 2020, 19, 944–959. [Google Scholar] [CrossRef] [Green Version]
- Malatesta, M.; Perdoni, F.; Muller, S.; Zancanaro, C.; Pellicciari, C. Nuclei of Aged Myofibres Undergo Structural and Functional Changes Suggesting Impairment in RNA Processing. Eur. J. Histochem. 2009, 53, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Zahn, J.M.; Sonu, R.; Vogel, H.; Crane, E.; Mazan-Mamczarz, K.; Rabkin, R.; Davis, R.W.; Becker, K.G.; Owen, A.B.; Kim, S.K. Transcriptional Profiling of Aging in Human Muscle Reveals a Common Aging Signature. PLoS Genet. 2006, 2, e115. [Google Scholar] [CrossRef]
- Ohlendieck, K. Proteomics of Skeletal Muscle Differentiation, Neuromuscular Disorders and Fiber Aging. Expert Rev. Proteom. 2010, 7, 283–296. [Google Scholar] [CrossRef]
- Doran, P.; Donoghue, P.; O’Connell, K.; Gannon, J.; Ohlendieck, K. Proteomics of Skeletal Muscle Aging. Proteomics 2009, 9, 989–1003. [Google Scholar] [CrossRef]
- Ahmad, K.; Shaikh, S.; Ahmad, S.S.; Lee, E.J.; Choi, I. Cross-Talk Between Extracellular Matrix and Skeletal Muscle: Implications for Myopathies. Front. Pharmacol. 2020, 11, 142. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, K.; Lee, E.J.; Moon, J.S.; Park, S.-Y.; Choi, I. Multifaceted Interweaving Between Extracellular Matrix, Insulin Resistance, and Skeletal Muscle. Cells 2018, 7, 148. [Google Scholar] [CrossRef] [Green Version]
- Hynes, R.O. The Extracellular Matrix: Not Just Pretty Fibrils. Science 2009, 326, 1216–1219. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Cánoves, P.; Neves, J.; Sousa-Victor, P. Understanding Muscle Regenerative Decline with Aging: New Approaches to Bring Back Youthfulness to Aged Stem Cells. FEBS J. 2020, 287, 406–416. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, J.; Holden, P.; Hansen, U. The Expanded Collagen VI Family: New Chains and New Questions. Connect. Tissue Res. 2013, 54, 345–350. [Google Scholar] [CrossRef] [Green Version]
- Kovanen, V.; Suominen, H.; Risteli, J.; Risteli, L. Type IV Collagen and Laminin in Slow and Fast Skeletal Muscle in Rats--Effects of Age and Life-Time Endurance Training. Coll. Relat. Res. 1988, 8, 145–153. [Google Scholar] [CrossRef]
- Snow, M.H. The Effects of Aging on Satellite Cells in Skeletal Muscles of Mice and Rats. Cell Tissue Res. 1977, 185, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Csapo, R.; Gumpenberger, M.; Wessner, B. Skeletal Muscle Extracellular Matrix—What Do We Know About Its Composition, Regulation, and Physiological Roles? A Narrative Review. Front. Physiol. 2020, 11, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, L.K.; Kayupov, E.; Gumucio, J.P.; Mendias, C.L.; Claflin, D.R.; Brooks, S.V. Intrinsic Stiffness of Extracellular Matrix Increases with Age in Skeletal Muscles of Mice. J. Appl. Physiol. 2014, 117, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Alnaqeeb, M.A.; Al Zaid, N.S.; Goldspink, G. Connective Tissue Changes and Physical Properties of Developing and Ageing Skeletal Muscle. J. Anat. 1984, 139 Pt 4, 677–689. [Google Scholar] [PubMed]
- Lacraz, G.; Rouleau, A.-J.; Couture, V.; Söllrald, T.; Drouin, G.; Veillette, N.; Grandbois, M.; Grenier, G. Increased Stiffness in Aged Skeletal Muscle Impairs Muscle Progenitor Cell Proliferative Activity. PLoS ONE 2015, 10, e0136217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovanen, V.; Suominen, H. Age- and Training-Related Changes in the Collagen Metabolism of Rat Skeletal Muscle. Eur. J. Appl. Physiol. Occup. Physiol. 1989, 58, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Haus, J.M.; Carrithers, J.A.; Trappe, S.W.; Trappe, T.A. Collagen, Cross-Linking, and Advanced Glycation End Products in Aging Human Skeletal Muscle. J. Appl. Physiol. 2007, 103, 2068–2076. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-J.; Lin, I.-H.; Lee, C.-W.; Chen, Y.-F. Aged Skeletal Muscle Retains the Ability to Remodel Extracellular Matrix for Degradation of Collagen Deposition after Muscle Injury. Int. J. Mol. Sci. 2021, 22, 2123. [Google Scholar] [CrossRef]
- Gillies, A.R.; Lieber, R.L. Structure and Function of the Skeletal Muscle Extracellular Matrix. Muscle Nerve 2011, 44, 318–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelse, K.; Pöschl, E.; Aigner, T. Collagens—Structure, Function, and Biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar] [CrossRef] [Green Version]
- Pavan, P.; Monti, E.; Bondí, M.; Fan, C.; Stecco, C.; Narici, M.; Reggiani, C.; Marcucci, L. Alterations of Extracellular Matrix Mechanical Properties Contribute to Age-Related Functional Impairment of Human Skeletal Muscles. Int. J. Mol. Sci. 2020, 21, 3992. [Google Scholar] [CrossRef]
- Gosselin, L.E.; Martinez, D.A.; Vailas, A.C.; Sieck, G.C. Passive Length-Force Properties of Senescent Diaphragm: Relationship with Collagen Characteristics. J. Appl. Physiol. 1994, 76, 2680–2685. [Google Scholar] [CrossRef]
- Hohenester, E.; Yurchenco, P.D. Laminins in Basement Membrane Assembly. Cell Adhes. Migr. 2013, 7, 56–63. [Google Scholar] [CrossRef] [Green Version]
- Grzelkowska-Kowalczyk, K. The Importance of Extracellular Matrix in Skeletal Muscle Development and Function; IntechOpen: London, UK, 2016; ISBN 978-953-51-2416-0. [Google Scholar]
- Sosa, P.; Alcalde-Estévez, E.; Asenjo-Bueno, A.; Plaza, P.; Carrillo-López, N.; Olmos, G.; López-Ongil, S.; Ruiz-Torres, M.P. Aging-Related Hyperphosphatemia Impairs Myogenic Differentiation and Enhances Fibrosis in Skeletal Muscle. J. Cachexia Sarcopenia Muscle 2021. [Google Scholar] [CrossRef] [PubMed]
- Conway, S.J.; Izuhara, K.; Kudo, Y.; Litvin, J.; Markwald, R.; Ouyang, G.; Arron, J.R.; Holweg, C.T.J.; Kudo, A. The Role of Periostin in Tissue Remodeling across Health and Disease. Cell Mol. Life Sci. 2014, 71, 1279–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horiuchi, K.; Amizuka, N.; Takeshita, S.; Takamatsu, H.; Katsuura, M.; Ozawa, H.; Toyama, Y.; Bonewald, L.F.; Kudo, A. Identification and Characterization of a Novel Protein, Periostin, with Restricted Expression to Periosteum and Periodontal Ligament and Increased Expression by Transforming Growth Factor Beta. J. Bone Miner. Res. 1999, 14, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Lorts, A.; Schwanekamp, J.A.; Baudino, T.A.; McNally, E.M.; Molkentin, J.D. Deletion of Periostin Reduces Muscular Dystrophy and Fibrosis in Mice by Modulating the Transforming Growth Factor-β Pathway. Proc. Natl. Acad. Sci. USA 2012, 109, 10978–10983. [Google Scholar] [CrossRef] [Green Version]
- Järvinen, T.A.; Kannus, P.; Järvinen, T.L.; Jozsa, L.; Kalimo, H.; Järvinen, M. Tenascin-C in the Pathobiology and Healing Process of Musculoskeletal Tissue Injury. Scand. J. Med. Sci. Sports 2000, 10, 376–382. [Google Scholar] [CrossRef]
- Settles, D.L.; Cihak, R.A.; Erickson, H.P. Tenascin-C Expression in Dystrophin-Related Muscular Dystrophy. Muscle Nerve 1996, 19, 147–154. [Google Scholar] [CrossRef]
- Schoser, B.G.; Faissner, A.; Goebel, H.H. Immunolocalization of Tenascin-C in Human Type II Fiber Atrophy. J. Mol. Neurosci. 1999, 13, 167–175. [Google Scholar] [CrossRef]
- Flück, M.; Mund, S.I.; Schittny, J.C.; Klossner, S.; Durieux, A.-C.; Giraud, M.-N. Mechano-Regulated Tenascin-C Orchestrates Muscle Repair. PNAS 2008, 105, 13662–13667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isenberg, J.S.; Roberts, D.D. Thrombospondin-1 in Maladaptive Aging Responses: A Concept Whose Time Has Come. Am. J. Physiol. Cell Physiol. 2020, 319, C45–C63. [Google Scholar] [CrossRef]
- Frazier, E.P.; Isenberg, J.S.; Shiva, S.; Zhao, L.; Schlesinger, P.; Dimitry, J.; Abu-Asab, M.S.; Tsokos, M.; Roberts, D.D.; Frazier, W.A. Age-Dependent Regulation of Skeletal Muscle Mitochondria by the Thrombospondin-1 Receptor CD47. Matrix Biol. 2011, 30, 154–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, B.; Serrano, A.L.; Tjwa, M.; Suelves, M.; Ardite, E.; De Mori, R.; Baeza-Raja, B.; Martínez de Lagrán, M.; Lafuste, P.; Ruiz-Bonilla, V.; et al. Fibrinogen Drives Dystrophic Muscle Fibrosis via a TGFbeta/Alternative Macrophage Activation Pathway. Genes Dev. 2008, 22, 1747–1752. [Google Scholar] [CrossRef] [Green Version]
- Kanapuru, B.; Ershler, W.B. Inflammation, Coagulation, and the Pathway to Frailty. Am. J. Med. 2009, 122, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Ameye, L.; Young, M.F. Mice Deficient in Small Leucine-Rich Proteoglycans: Novel in Vivo Models for Osteoporosis, Osteoarthritis, Ehlers-Danlos Syndrome, Muscular Dystrophy, and Corneal Diseases. Glycobiology 2002, 12, 107R–116R. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, M.S. Proteoglycans and Pathophysiology. J. Appl. Physiol. 2007, 103, 735–736. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xu, X.; Ma, H.; Jiang, J. Integrative Analysis of Transcriptomics and Proteomics of Skeletal Muscles of the Chinese Indigenous Shaziling Pig Compared with the Yorkshire Breed. BMC Genet. 2016, 17, 80. [Google Scholar] [CrossRef] [Green Version]
- Davey, J.R.; Watt, K.I.; Parker, B.L.; Chaudhuri, R.; Ryall, J.G.; Cunningham, L.; Qian, H.; Sartorelli, V.; Sandri, M.; Chamberlain, J.; et al. Integrated Expression Analysis of Muscle Hypertrophy Identifies Asb2 as a Negative Regulator of Muscle Mass. JCI Insight 2016, 1. [Google Scholar] [CrossRef] [Green Version]
- Kühl, I.; Miranda, M.; Atanassov, I.; Kuznetsova, I.; Hinze, Y.; Mourier, A.; Filipovska, A.; Larsson, N.-G. Transcriptomic and Proteomic Landscape of Mitochondrial Dysfunction Reveals Secondary Coenzyme Q Deficiency in Mammals. eLife 2017, 6, e30952. [Google Scholar] [CrossRef]
- Croissant, C.; Gounou, C.; Bouvet, F.; Tan, S.; Bouter, A. Annexin-A6 in Membrane Repair of Human Skeletal Muscle Cell: A Role in the Cap Subdomain. Cells 2020, 9, 1742. [Google Scholar] [CrossRef]
- Croissant, C.; Carmeille, R.; Brévart, C.; Bouter, A. Annexins and Membrane Repair Dysfunctions in Muscular Dystrophies. Int. J. Mol. Sci. 2021, 22, 5276. [Google Scholar] [CrossRef]
- Cooper, D.N.; Barondes, S.H. Evidence for Export of a Muscle Lectin from Cytosol to Extracellular Matrix and for a Novel Secretory Mechanism. J. Cell Biol. 1990, 110, 1681–1691. [Google Scholar] [CrossRef] [Green Version]
- Giresi, P.G.; Stevenson, E.J.; Theilhaber, J.; Koncarevic, A.; Parkington, J.; Fielding, R.A.; Kandarian, S.C. Identification of a Molecular Signature of Sarcopenia. Physiol. Genom. 2005, 21, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Watt, D.J.; Jones, G.E.; Goldring, K. The Involvement of Galectin-1 in Skeletal Muscle Determination, Differentiation and Regeneration. Glycoconj. J. 2002, 19, 615–619. [Google Scholar] [CrossRef]
- L’hôte, C.; Cordier, B.; Labasse, A.; Boileau, C.; Costes, B.; Henrotin, Y. Identification of New Biomarkers for Sarcopenia and Characterization of Cathepsin D Biomarker. JCSM Rapid Commun. 2021, 4, 122–132. [Google Scholar] [CrossRef]
- Alameddine, H.S. Matrix Metalloproteinases in Skeletal Muscles: Friends or Foes? Neurobiol. Dis. 2012, 48, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Chandler, K.B.; Brnakova, Z.; Sanda, M.; Wang, S.; Stalnaker, S.H.; Bridger, R.; Zhao, P.; Wells, L.; Edwards, N.J.; Goldman, R. Site-Specific Glycan Microheterogeneity of Inter-Alpha-Trypsin Inhibitor Heavy Chain H4. J. Proteome Res. 2014, 13, 3314–3329. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Hou, H.; Yi, W.; Yang, G.; Gu, C.; Lau, W.B.; Gao, E.; Ma, X.; Lu, Z.; Wei, X.; et al. Increased Expression of Pigment Epithelium-Derived Factor in Aged Mesenchymal Stem Cells Impairs Their Therapeutic Efficacy for Attenuating Myocardial Infarction Injury. Eur. Heart J. 2013, 34, 1681–1690. [Google Scholar] [CrossRef] [PubMed]
- Boraldi, F.; Lofaro, F.D.; Accorsi, A.; Ross, E.; Malagoli, D. Toward the Molecular Deciphering of Pomacea Canaliculata Immunity: First Proteomic Analysis of Circulating Hemocytes. Proteomics 2019, 19, e1800314. [Google Scholar] [CrossRef] [PubMed]
- Boraldi, F.; Moscarelli, P.; Lofaro, F.D.; Sabia, C.; Quaglino, D. The Mineralization Process of Insoluble Elastin Fibrillar Structures: Ionic Environment vs Degradation. Int. J. Biol. Macromol. 2020, 149, 693–706. [Google Scholar] [CrossRef]
- Singhto, N.; Thongboonkerd, V. Exosomes Derived from Calcium Oxalate-Exposed Macrophages Enhance IL-8 Production from Renal Cells, Neutrophil Migration and Crystal Invasion through Extracellular Matrix. J. Proteom. 2018, 185, 64–76. [Google Scholar] [CrossRef]
- Azizi, E.; Deslauriers, A.R.; Holt, N.C.; Eaton, C.E. Resistance to Radial Expansion Limits Muscle Strain and Work. Biomech. Model. Mechanobiol. 2017, 16, 1633–1643. [Google Scholar] [CrossRef] [Green Version]
- Sharafi, B.; Blemker, S.S. A Mathematical Model of Force Transmission from Intrafascicularly Terminating Muscle Fibers. J. Biomech. 2011, 44, 2031–2039. [Google Scholar] [CrossRef] [Green Version]
- Gillies, A.R.; Chapman, M.A.; Bushong, E.A.; Deerinck, T.J.; Ellisman, M.H.; Lieber, R.L. High Resolution Three-Dimensional Reconstruction of Fibrotic Skeletal Muscle Extracellular Matrix. J. Physiol. 2017, 595, 1159–1171. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Symbol | Protein Name | Protein Symbol | Protein Name |
|---|---|---|---|
| MATRISOME DIVISION: CORE MATRISOME | |||
| Category: collagen | |||
| CO1A1 | Collagen alpha-1(I) chain | CO6A1 | Collagen alpha-1(VI) chain |
| CO1A2 | Collagen alpha-2(I) chain | CO6A2 | Collagen alpha-2(VI) chain |
| CO3A1 | Collagen alpha-1(III) chain | CO6A6 | Collagen alpha-6(VI) chain |
| CO4A1 | Collagen alpha-1(IV) chain | COBA1 | Collagen alpha-1(XI) chain |
| CO4A2 | Collagen alpha-2(IV) chain | COEA1 | Collagen alpha-1(XIV) chain |
| CO5A1 | Collagen alpha-1(V) chain | COFA1 | Collagen alpha-1(XV) chain |
| CO5A2 | Collagen alpha-2(V) chain | COIA1 | Collagen alpha-1(XVIII) chain |
| Category: ECM-glycoproteins | |||
| ADIPO | Adiponectin | LAMB1 | Laminin subunit beta-1 |
| AGRIN | Agrin | LAMB2 | Laminin subunit beta-2 |
| BGH3 | Transforming growth factor-beta-induced protein ig-h3 | LAMC1 | Laminin subunit gamma-1 |
| CILP1 | Cartilage intermediate layer protein 1 | MFAP2 | Microfibrillar-associated protein 2 |
| CILP2 | Cartilage intermediate layer protein 2 | MFAP4 | Microfibril-associated glycoprotein 4 |
| COMP | Cartilage oligomeric matrix protein | MFAP5 | Microfibrillar-associated protein 5 |
| CREL1 | Cysteine-rich with EGF-like domain protein 1 | MFGM | Lactadherin |
| DERM | Dermatopontin | NID1 | Nidogen-1 |
| FBN1 | Fibrillin-1 | NID2 | Nidogen-2 |
| FBN2 | Fibrillin-2 | POSTN | Periostin |
| FIBA | Fibrinogen alpha chain | SLIT2 | Slit homolog 2 protein |
| FIBB | Fibrinogen beta chain | SRPX | Sushi-repeat-containing protein SRPX |
| FIBG | Fibrinogen gamma chain | SSPO | SCO-spondin |
| FINC | Fibronectin | TENA | Tenascin |
| IGS10 | Immunoglobulin superfamily member 10 | TINAL | Tubulointerstitial nephritis antigen-like |
| LAMA2 | Laminin subunit alpha-2 | TSP1 | Thrombospondin-1 |
| LAMA3 | Laminin subunit alpha-3 | TSP4 | Thrombospondin-4 |
| LAMA4 | Laminin subunit alpha-4 | VMA5A | von Willebrand factor A domain-containing protein 5A |
| LAMA5 | Laminin subunit alpha-5 | VWA1 | von Willebrand factor A domain-containing protein 1 |
| Category: Proteoglycans | |||
| ASPN | Asporin | PGBM | Basement membrane-specific heparan sulfate proteoglycan core protein |
| CHADL | Chondroadherin-like protein | PGS1 | Biglycan |
| FMOD | Fibromodulin | PGS2 | Decorin |
| LUM | Lumican | PRELP | Prolargin |
| MIME | Mimecan | PRG2 | Bone marrow proteoglycan |
| MATRISOME DIVISION: MATRISOME-ASSOCIATED PROTEINS | |||
| Category: ECM affiliated proteins | |||
| ANXA1 | Annexin A1 | GPC1 | Glypican-1 |
| ANXA2 | Annexin A2 | HEMO | Hemopexin |
| ANXA3 | Annexin A3 | LEG1 | Galectin-1 |
| ANXA4 | Annexin A4 | LEGL | Galectin-related protein |
| ANXA5 | Annexin A5 | LMAN1 | Protein ERGIC-53 |
| ANXA6 | Annexin A6 | PLXA4 | Plexin-A4 |
| ANXA7 | Annexin A7 | PLXB3 | Plexin-B3 |
| ANX11 | Annexin A11 | ||
| Category: ECM regulators | |||
| A1AT1 | Alpha-1-antitrypsin 1-1 | ITIH1 | Inter-alpha-trypsin inhibitor heavy chain H1 |
| A1AT2 | Alpha-1-antitrypsin 1-2 | ITIH2 | Inter-alpha-trypsin inhibitor heavy chain H2 |
| A1AT3 | Alpha-1-antitrypsin 1-3 | ITIH3 | Inter-alpha-trypsin inhibitor heavy chain H3 |
| A1AT4 | Alpha-1-antitrypsin 1-4 | ITIH4 | Inter alpha-trypsin inhibitor, heavy chain 4 |
| A2AP | Alpha-2-antiplasmin | ITIH5 | Inter-alpha-trypsin inhibitor heavy chain H5 |
| AMBP | Protein AMBP | KNG1 | Kininogen-1 |
| ANGT | Angiotensinogen | MMP17 | Matrix metalloproteinase-17 |
| ANT3 | Antithrombin-III | PEDF | Pigment epithelium-derived factor |
| CATB | Cathepsin B | PLMN | Plasminogen |
| CATD | Cathepsin D | PZP | Pregnancy zone protein |
| CBG | Corticosteroid-binding globulin | SERPH | Serpin H1 |
| CPN2 | Carboxypeptidase N subunit 2 | SPA3K | Serine protease inhibitor A3K |
| CYTB | Cystatin-B | SPA3M | Serine protease inhibitor A3M |
| FA12 | Coagulation factor XII | SPA3N | Serine protease inhibitor A3N |
| HEP2 | Heparin cofactor 2 | SPI2 | Serpin I2 |
| HRG | Histidine-rich glycoprotein | SULF2 | Extracellular sulfatase Sulf-2 |
| HYAL2 | Hyaluronidase-2 | TGM2 | Protein-glutamine gamma-glutamyltransferase 2 |
| IC1 | Plasma protease C1 inhibitor | THRB | Prothrombin |
| ILEUA | Leukocyte elastase inhibitor A | ||
| Category: secreted factors | |||
| ANGL7 | Angiopoietin-related protein 7 | S10A1 | Protein S100-A1 |
| FILA2 | Filaggrin-2 | S10A4 | Protein S100-A4 |
| HGFA | Hepatocyte growth factor activator | S10A6 | Protein S100-A6 |
| INHBA | Inhibin beta A chain | S10AA | Protein S100-A10 |
| MEG11 | Multiple epidermal growth factor-like domains protein 11 | WN10A | Protein Wnt-10a |
| PBS Extract | ||||
|---|---|---|---|---|
| Category | Protein Symbol | Protein Name | Ratio (Old/Adult) Log2 Fold Change | p-Value |
| ECM regulators | A1AT2 | Alpha-1-antitrypsin 1-2 | 0.69 | 0.006 |
| A2AP | Alpha-2-antiplasmin | 1.04 | 0.038 | |
| CATD | Cathepsin D | 0.58 | 0.001 | |
| CBG | Corticosteroid-binding globulin | 1.82 | 0.049 | |
| ILEUA | Leukocyte elastase inhibitor A | 1.11 | 0.008 | |
| ITIH2 | Inter-alpha-trypsin inhibitor heavy chain H2 | 2.00 | 0.038 | |
| ITIH4 | Inter alpha-trypsin inhibitor, heavy chain 4 | 1.42 | 0.018 | |
| KNG1 | Kininogen-1 | 0.97 | 0.043 | |
| PEDF | Pigment epithelium-derived factor | 1.11 | 0.040 | |
| PZP | Pregnancy zone protein | 0.86 | 0.012 | |
| SPA3N | Serine protease inhibitor A3N | 1.77 | 0.028 | |
| ECM glycoproteins | FIBA | Fibrinogen alpha chain | 1.19 | 0.001 |
| FIBB | Fibrinogen beta chain | 1.19 | 0.001 | |
| FIBG | Fibrinogen gamma chain | 1.14 | 0.001 | |
| TSP4 | Thrombospondin-4 | 1.44 | 0.000 | |
| VMA5A | von Willebrand factor A domain-containing protein 5A | 0.81 | 0.000 | |
| Proteoglycans | ASPN | Asporin | 0.53 | 0.004 |
| LUM | Lumican | 0.48 | 0.030 | |
| PRELP | Prolargin | 0.37 | 0.001 | |
| ECM affiliated proteins | ANXA4 | Annexin A4 | 0.63 | 0.002 |
| ANXA5 | Annexin A5 | 0.51 | 0.009 | |
| ANXA6 | Annexin A6 | −0.39 | 0.008 | |
| ANX11 | Annexin A11 | 0.28 | 0.021 | |
| HEMO | Hemopexin | 0.74 | 0.018 | |
| U/T Extract | ||||
| Collagen | CO4A1 | Collagen alpha-1(IV) chain | 1.30 | 0.016 |
| CO6A1 | Collagen alpha-1(VI) chain | 0.61 | 0.000 | |
| CO6A2 | Collagen alpha-2(VI) chain | 0.59 | 0.001 | |
| ECM regulators | A1AT2 | Alpha-1-antitrypsin 1-2 | 0.87 | 0.022 |
| CATD | Cathepsin D | 1.04 | 0.000 | |
| TGM2 | Protein-glutamine gamma-glutamyltransferase 2 | 0.80 | 0.013 | |
| ECM glycoproteins | BGH3 | Transforming growth factor-beta-induced protein ig-h3 | 0.78 | 0.016 |
| CILP1 | Cartilage intermediate layer protein 1 | 1.47 | 0.000 | |
| DERM | Dermatopontin | 0.51 | 0.004 | |
| FINC | Fibronectin | 1.35 | 0.036 | |
| LAMA2 | Laminin subunit alpha-2 | 0.45 | 0.000 | |
| LAMA5 | Laminin subunit alpha-5 | 0.79 | 0.002 | |
| LAMB1 | Laminin subunit beta-1 | 0.47 | 0.010 | |
| LAMB2 | Laminin subunit beta-2 | 0.53 | 0.000 | |
| LAMC1 | Laminin subunit gamma-1 | 0.56 | 0.000 | |
| NID1 | Nidogen-1 | 0.54 | 0.000 | |
| NID2 | Nidogen-2 | 1.09 | 0.000 | |
| POSTN | Periostin | 1.28 | 0.003 | |
| TENA | Tenascin | 2.55 | 0.022 | |
| TSP4 | Thrombospondin-4 | 1.34 | 0.048 | |
| Proteoglycans | ASPN | Asporin | 0.79 | 0.008 |
| FMOD | Fibromodulin | 0.73 | 0.005 | |
| LUM | Lumican | 0.81 | 0.001 | |
| MIME | Mimecan | 0.54 | 0.008 | |
| PGS1 | Biglycan | 1.05 | 0.002 | |
| PRELP | Prolargin | 0.87 | 0.007 | |
| ECM affiliated proteins | ANXA2 | Annexin A2 | 0.42 | 0.005 |
| ANXA5 | Annexin A5 | 2.28 | 0.002 | |
| HEMO | Hemopexin | 1.59 | 0.007 | |
| ANX11 | Annexin A11 | 0.90 | 0.000 | |
| LEG1 | Galectin-1 | 1.19 | 0.021 | |
| GuHCl Extract | ||||
| Collagens | COFA1 | Collagen alpha-1(XV) chain | 0.67 | 0.019 |
| ECM glycoproteins | LAMA5 | Laminin subunit alpha-5 | 0.81 | 0.006 |
| ECM affiliated proteins | ANXA2 | Annexin A2 | 0.67 | 0.046 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lofaro, F.D.; Cisterna, B.; Lacavalla, M.A.; Boschi, F.; Malatesta, M.; Quaglino, D.; Zancanaro, C.; Boraldi, F. Age-Related Changes in the Matrisome of the Mouse Skeletal Muscle. Int. J. Mol. Sci. 2021, 22, 10564. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910564
Lofaro FD, Cisterna B, Lacavalla MA, Boschi F, Malatesta M, Quaglino D, Zancanaro C, Boraldi F. Age-Related Changes in the Matrisome of the Mouse Skeletal Muscle. International Journal of Molecular Sciences. 2021; 22(19):10564. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910564
Chicago/Turabian StyleLofaro, Francesco Demetrio, Barbara Cisterna, Maria Assunta Lacavalla, Federico Boschi, Manuela Malatesta, Daniela Quaglino, Carlo Zancanaro, and Federica Boraldi. 2021. "Age-Related Changes in the Matrisome of the Mouse Skeletal Muscle" International Journal of Molecular Sciences 22, no. 19: 10564. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910564