
Melatonin-Activated Receptor Signaling Pathways Mediate Protective Effects on Surfactant-Induced Increase in Jejunal Mucosal Permeability in Rats


Abstract
:1. Introduction
2. Results
3. Discussion
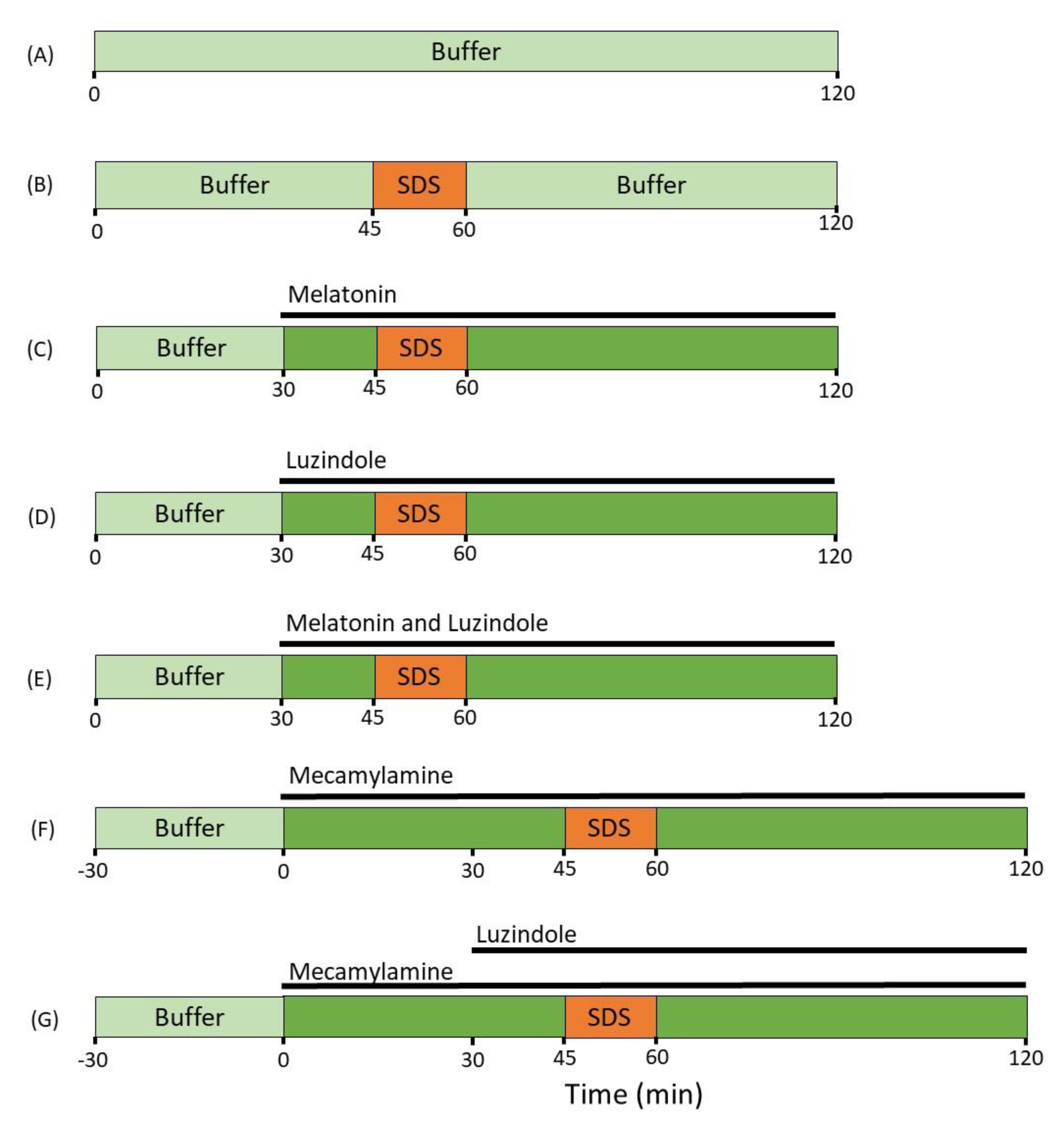
4. Materials and Methods
4.1. Chemicals and Drugs
4.2. Study Formulations
4.3. Animals, Anesthesia and Surgery
4.4. Perfusion Study
4.5. Determination of Blood-to-Lumen Jejunal Mucosal 51Cr-EDTA Clearance
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schoultz, I.; Keita, Å.V. The Intestinal Barrier and Current Techniques for the Assessment of Gut Permeability. Cells 2020, 9, 1909. [Google Scholar] [CrossRef] [PubMed]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms Regulating Intestinal Barrier Integrity and Its Pathological Implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Barker, N. Adult Intestinal Stem Cells: Critical Drivers of Epithelial Homeostasis and Regeneration. Nat. Rev. Mol. Cell Biol. 2014, 15, 19–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, I.; Bradbury, N.A. Chapter Ten-Physiology of the Gut: Experimental Models for Investigating Intestinal Fluid and Electrolyte Transport. In Current Topics in Membranes; Levitane, I., Delpire, E., Rasgado-Flores, H., Eds.; Cell Volume Regulation; Academic Press: Cambridge, MA, USA, 2018; Volume 81, pp. 337–381. [Google Scholar]
- Buckley, A.; Turner, J.R. Cell Biology of Tight Junction Barrier Regulation and Mucosal Disease. Cold Spring Harb Perspect Biol 2018, 10, a029314. [Google Scholar] [CrossRef]
- Odenwald, M.A.; Turner, J.R. The Intestinal Epithelial Barrier: A Therapeutic Target? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, M. Leaky Gut: Mechanisms, Measurement and Clinical Implications in Humans. Gut 2019, 68, 1516–1526. [Google Scholar] [CrossRef]
- Camilleri, M.; Gorman, H. Intestinal Permeability and Irritable Bowel Syndrome. Neurogastroenterol. Motil. 2007, 19, 545–552. [Google Scholar] [CrossRef]
- Sangild, P.T.; Shen, R.L.; Pontoppidan, P.; Rathe, M. Animal Models of Chemotherapy-Induced Mucositis: Translational Relevance and Challenges. Am. J. Physiol.-Gastrointest. Liver Physiol. 2018, 314, G231–G246. [Google Scholar] [CrossRef]
- Zhou, Q.; Verne, G.N. Intestinal Hyperpermeability: A Gateway to Multi-Organ Failure? Available online: https://www.jci.org/articles/view/124366/pdf (accessed on 29 August 2021).
- Odenwald, M.A.; Turner, J.R. Intestinal Permeability Defects: Is It Time to Treat? Clin. Gastroenterol. Hepatol. 2013, 11, 1075–1083. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, M.; Vitetta, L. Adjunctive Treatments for the Prevention of Chemotherapy- and Radiotherapy-Induced Mucositis. Integr. Cancer 2018, 17, 1027–1047. [Google Scholar] [CrossRef] [Green Version]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.-D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal Permeability–a New Target for Disease Prevention and Therapy. BMC Gastroenterol. 2014, 14, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Zisapel, N.; Tarrasch, R.; Laudon, M. The Relationship between Melatonin and Cortisol Rhythms: Clinical Implications of Melatonin Therapy. Drug Dev. Res. 2005, 65, 119–125. [Google Scholar] [CrossRef]
- Bubenik, G.A. REVIEW: Gastrointestinal Melatonin: Localization, Function, and Clinical Relevance. Dig. Dis. Sci. 2002, 47, 2336–2348. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Mayo, J.C.; Tan, D.-X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an Antioxidant: Under Promises but over Delivers. J. Pineal. Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Pieri, C.; Marra, M.; Moroni, F.; Recchioni, R.; Marcheselli, F. Melatonin: A Peroxyl Radical Scavenger More Effective than Vitamin E. Life Sci. 1994, 55, PL271–PL276. [Google Scholar] [CrossRef]
- Slominski, R.M.; Reiter, R.J.; Schlabritz-Loutsevitch, N.; Ostrom, R.S.; Slominski, A.T. Melatonin Membrane Receptors in Peripheral Tissues: Distribution and Functions. Mol. Cell Endocrinol 2012, 351, 152–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommansson, A.; Saudi, W.S.W.; Nylander, O.; Sjöblom, M. Melatonin Inhibits Alcohol-Induced Increases in Duodenal Mucosal Permeability in Rats in Vivo. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, 95–105. [Google Scholar] [CrossRef]
- Monobe, M.; Hino, M.; Sumi, M.; Uzawa, A.; Hirayama, R.; Ando, K.; Kojima, S. Protective Effects of Melatonin on γ-Ray Induced Intestinal Damage. Int. J. Radiat. Biol. 2005, 81, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Sommansson, A.; Nylander, O.; Sjöblom, M. Melatonin Decreases Duodenal Epithelial Paracellular Permeability via a Nicotinic Receptor-Dependent Pathway in Rats in Vivo. J. Pineal Res. 2013, 54, 282–291. [Google Scholar] [CrossRef]
- Maher, S.; Brayden, D.J. Formulation Strategies to Improve the Efficacy of Intestinal Permeation Enhancers. Adv. Drug Deliv. Rev. 2021, 113925. [Google Scholar] [CrossRef] [PubMed]
- Dahlgren, D.; Roos, C.; Lundqvist, A.; Tannergren, C.; Sjöblom, M.; Sjögren, E.; Lennernäs, H. Effect of Absorption-Modifying Excipients, Hypotonicity, and Enteric Neural Activity in an in Vivo Model for Small Intestinal Transport. Int. J. Pharm. 2018, 549, 239–248. [Google Scholar] [CrossRef]
- Dahlgren, D.; Cano-Cebrián, M.J.; Hellström, P.M.; Wanders, A.; Sjöblom, M.; Lennernäs, H. Prevention of Rat Intestinal Injury with a Drug Combination of Melatonin and Misoprostol. Int. J. Mol. Sci. 2020, 21, 6771. [Google Scholar] [CrossRef] [PubMed]
- Galligan, J.J.; North, R.A. Pharmacology and Function of Nicotinic Acetylcholine and P2X Receptors in the Enteric Nervous System. Neurogastroenterol. Motil. 2004, 16, 64–70. [Google Scholar] [CrossRef]
- Furness, J.B. The Enteric Nervous System and Neurogastroenterology. Nat. Rev. Gastroenterol Hepatol 2012, 9, 286–294. [Google Scholar] [CrossRef]
- Fihn, B.-M.; Sjöqvist, A.; Jodal, M. Involvement of Enteric Nerves in Permeability Changes Due to Deoxycholic Acid in Rat Jejunum in Vivo. Acta Physiol. Scand. 2003, 178, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Nylander, O.; Pihl, L.; Perry, M. Hypotonicity-Induced Increases in Duodenal Mucosal Permeability Facilitates Adjustment of Luminal Osmolality. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G360–G370. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Liang, H.; Li, Z.-F.; Xiang, H.; Liu, W.; Li, J.-G. Vagus Nerve Stimulation Attenuates Intestinal Epithelial Tight Junctions Disruption in Endotoxemic Mice Through A7 Nicotinic Acetylcholine Receptors. Shock 2013, 40, 144–151. [Google Scholar] [CrossRef]
- Nylander, O.; Sababí, M.; Bark, J. Characterization of 51Cr-EDTA as a Marker of Duodenal Mucosal Permeability. Acta Physiol. Scand. 1991, 143, 117–126. [Google Scholar] [CrossRef]
- Dahlgren, D.; Sjöblom, M.; Hedeland, M.; Lennernäs, H. The In Vivo Effect of Transcellular Permeation Enhancers on the Intestinal Permeability of Two Peptide Drugs Enalaprilat and Hexarelin. Pharmaceutics 2020, 12, 99. [Google Scholar] [CrossRef] [Green Version]
- Anosov, A.A.; Smirnova, E.Y.; Korepanova, E.A.; Shogenov, I.M. The Effects of SDS at Subsolubilizing Concentrations on the Planar Lipid Bilayer Permeability: Two Kinds of Current Fluctuations. Chem. Phys. Lipids 2019, 218, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Danielsen, E.M.; Hansen, G.H. Intestinal Surfactant Permeation Enhancers and Their Interaction with Enterocyte Cell Membranes in a Mucosal Explant System. Tissue Barriers 2017, 5, e1361900. [Google Scholar] [CrossRef]
- Anderberg, E.K.; Artursson, P. Epithelial Transport of Drugs in Cell Culture. VIII: Effects of Sodium Dodecyl Sulfate on Cell Membrane and Tight Junction Permeability in Human Intestinal Epithelial (Caco-2) Cells. J. Pharm. Sci. 1993, 82, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Swenson, E.S.; Milisen, W.B.; Curatolo, W. Intestinal Permeability Enhancement: Efficacy, Acute Local Toxicity, and Reversibility. Pharm Res. 1994, 11, 1132–1142. [Google Scholar] [CrossRef] [PubMed]
- Dahlgren, D.; Roos, C.; Lundqvist, A.; Tannergren, C.; Sjöblom, M.; Sjögren, E.; Lennernäs, H. Time-Dependent Effects on Small Intestinal Transport by Absorption-Modifying Excipients. Eur. J. Pharm. Biopharm. 2018, 132, 19–28. [Google Scholar] [CrossRef]
- Sjöblom, M.; Jedstedt, G.; Flemström, G. Peripheral Melatonin Mediates Neural Stimulation of Duodenal Mucosal Bicarbonate Secretion. J. Clin. Invest. 2001, 108, 625–633. [Google Scholar] [CrossRef]
- Sjöblom, M. The Duodenal Mucosal Bicarbonate Secretion. Ups J. Med. Sci. 2005, 110, 115–149. [Google Scholar] [CrossRef] [PubMed]
- Drago, F.; Macauda, S.; Salehi, S. Small Doses of Melatonin Increase Intestinal Motility in Rats. Dig. Dis. Sci. 2002, 47, 1969–1974. [Google Scholar] [CrossRef]
- Bacher, I.; Wu, B.; Shytle, D.R.; George, T.P. Mecamylamine–a Nicotinic Acetylcholine Receptor Antagonist with Potential for the Treatment of Neuropsychiatric Disorders. Expert Opin. Pharmacother. 2009, 10, 2709–2721. [Google Scholar] [CrossRef]
- Hällgren, A.; Wilander, E.; Nylander, O. Acid-Induced Increase in Duodenal Mucosal Permeability Is Augmented by Nitric Oxide Inhibition and Vasopressin. Acta Physiol. Scand. 1997, 160, 363–370. [Google Scholar] [CrossRef]
- Pihl, L.; Wilander, E.; Nylander, O. Comparative Study of the Effect of Luminal Hypotonicity on Mucosal Permeability in Rat Upper Gastrointestinal Tract. Acta Physiol. 2008, 193, 67–78. [Google Scholar] [CrossRef]
- Lambert, G.P.; Chang, R.T.; Xia, T.; Summers, R.W.; Gisolfi, C.V. Absorption from Different Intestinal Segments during Exercise. J. Appl. Physiol. 1997, 83, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Mora, M.J.; Petiti, J.P.; Longhi, M.R.; Torres, A.I.; Granero, G.E. Intestinal Uptake and Toxicity Evaluation of Acetazolamide and Its Multicomponent Complexes with Hidroxypropyl-β-Cyclodextrin in Rats. Int. J. Pharm. 2015, 478, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Fihn, B.-M.; Sjöqvist, A.; Jodal, M. Effect of Cholera Toxin on Passive Transepithelial Transport of 51Cr-Ethylenediaminetetraacetic Acid and 14C-Mannitol in Rat Jejunum. Acta Physiol. Scand. 2001, 171, 153–160. [Google Scholar] [CrossRef]
- Zakeri-Milani, P.; Valizadeh, H.; Azarmi, Y.; Islambolchilar, Z.; Barzegar, S.; Barzegar-Jalali, M. Predicting Human Intestinal Permeability Using Single-Pass Intestinal Perfusion in Rat. J. Pharm. Pharm. Sci. 2007, 10, 368–379. [Google Scholar]
- Cao, X.; Gibbs, S.T.; Fang, L.; Miller, H.A.; Landowski, C.P.; Shin, H.-C.; Lennernäs, H.; Zhong, Y.; Amidon, G.L.; Yu, L.X.; et al. Why Is It Challenging to Predict Intestinal Drug Absorption and Oral Bioavailability in Human Using Rat Model. Pharm. Res. 2006, 23, 1675–1686. [Google Scholar] [CrossRef] [PubMed]
- Sedin, J.; Sjöblom, M.; Nylander, O. Prevention of Duodenal Ileus Reveals Functional Differences in the Duodenal Response to Luminal Hypertonicity in Sprague-Dawley and Dark Agouti Rats. Acta Physiol. 2014, 210, 573–589. [Google Scholar] [CrossRef]
- Sedin, J.; Sjöblom, M.; Nylander, O. The Selective Cyclooxygenase-2 Inhibitor Parecoxib Markedly Improves the Ability of the Duodenum to Regulate Luminal Hypertonicity in Anaesthetized Rats. Acta Physiol. 2012, 205, 433–451. [Google Scholar] [CrossRef]
- Dahlgren, D.; Roos, C.; Peters, K.; Lundqvist, A.; Tannergren, C.; Sjögren, E.; Sjöblom, M.; Lennernäs, H. Evaluation of Drug Permeability Calculation Based on Luminal Disappearance and Plasma Appearance in the Rat Single-Pass Intestinal Perfusion Model. Eur. J. Pharm. Biopharm. 2019, 142, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Dahlgren, D.; Sjöblom, M.; Hellström, P.M.; Lennernäs, H. Chemotherapeutics-Induced Intestinal Mucositis: Pathophysiology and Potential Treatment Strategies. Front. Pharm. 2021, 12, 681417. [Google Scholar] [CrossRef]
- Esteban-Zubero, E.; López-Pingarrón, L.; Alatorre-Jiménez, M.A.; Ochoa-Moneo, P.; Buisac-Ramón, C.; Rivas-Jiménez, M.; Castán-Ruiz, S.; Antoñanzas-Lombarte, Á.; Tan, D.X.; García, J.J.; et al. Melatonin’s Role as a Co-Adjuvant Treatment in Colonic Diseases: A Review. Life Sci. 2017, 170, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Dahlgren, D.; Roos, C.; Lundqvist, A.; Tannergren, C.; Langguth, P.; Sjöblom, M.; Sjögren, E.; Lennernäs, H. Preclinical Effect of Absorption Modifying Excipients on Rat Intestinal Transport of Model Compounds and the Mucosal Barrier Marker 51Cr-EDTA. Mol. Pharm. 2017, 14, 4243–4251. [Google Scholar] [CrossRef] [PubMed]
- Pihl, L.; Nylander, O. Products of Cyclooxygenase-2 Depress Duodenal Function in Rats Subjected to Abdominal Surgery. Acta Physiol. 2006, 186, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Nylander, O.; Kvietys, P.; Granger, D.N. Effects of Hydrochloric Acid on Duodenal and Jejunal Mucosal Permeability in the Rat. Am. J. Physiol. Gastrointest. Liver Physiol. 1989, 257, G653–G660. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Groups | MABP (mmHg) (0–120 min) | MABP (mmHg) (0–15 min) | MABP (mmHg) (105–120 min) |
|---|---|---|---|
| Control | 111 ± 5 | 113 ± 2 | 113 ± 7 |
| SDS | 100 ± 4 | 104 ± 4 | 97 ± 5 |
| SDS and melatonin | 104 ± 5 | 109 ± 8 | 99 ± 3 |
| SDS and luzindole | 100 ± 5 | 100 ± 6 | 105 ± 6 |
| SDS, melatonin and luzindole | 111 ± 8 | 112 ± 10 | 112 ± 9 |
| SDS and mecamylamine | 79 ± 6 | 95 ± 3 | 77 ± 7 * |
| SDS, mecamylamine and luzindole | 93 ± 4 | 91 ± 7 | 93 ± 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peters, K.; Dahlgren, D.; Lennernäs, H.; Sjöblom, M. Melatonin-Activated Receptor Signaling Pathways Mediate Protective Effects on Surfactant-Induced Increase in Jejunal Mucosal Permeability in Rats. Int. J. Mol. Sci. 2021, 22, 10762. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910762
Peters K, Dahlgren D, Lennernäs H, Sjöblom M. Melatonin-Activated Receptor Signaling Pathways Mediate Protective Effects on Surfactant-Induced Increase in Jejunal Mucosal Permeability in Rats. International Journal of Molecular Sciences. 2021; 22(19):10762. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910762
Chicago/Turabian StylePeters, Karsten, David Dahlgren, Hans Lennernäs, and Markus Sjöblom. 2021. "Melatonin-Activated Receptor Signaling Pathways Mediate Protective Effects on Surfactant-Induced Increase in Jejunal Mucosal Permeability in Rats" International Journal of Molecular Sciences 22, no. 19: 10762. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910762