Steroidogenesis in Peripheral and Transition Zones of Human Prostate Cancer Tissue

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Quantification of Steroids by LC/MS Analyses in Human Prostate Steroidogenesis
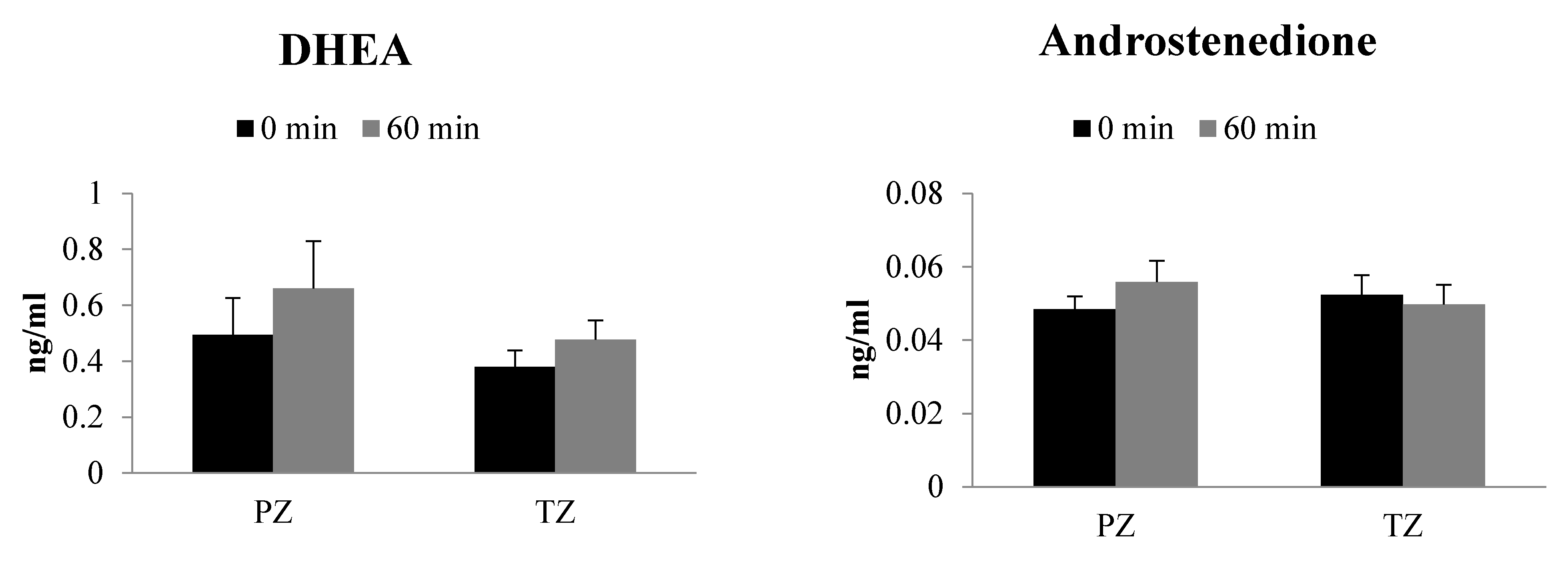
2.2. Basal Steroid Levels in PZ and TZ Prostate Tissues
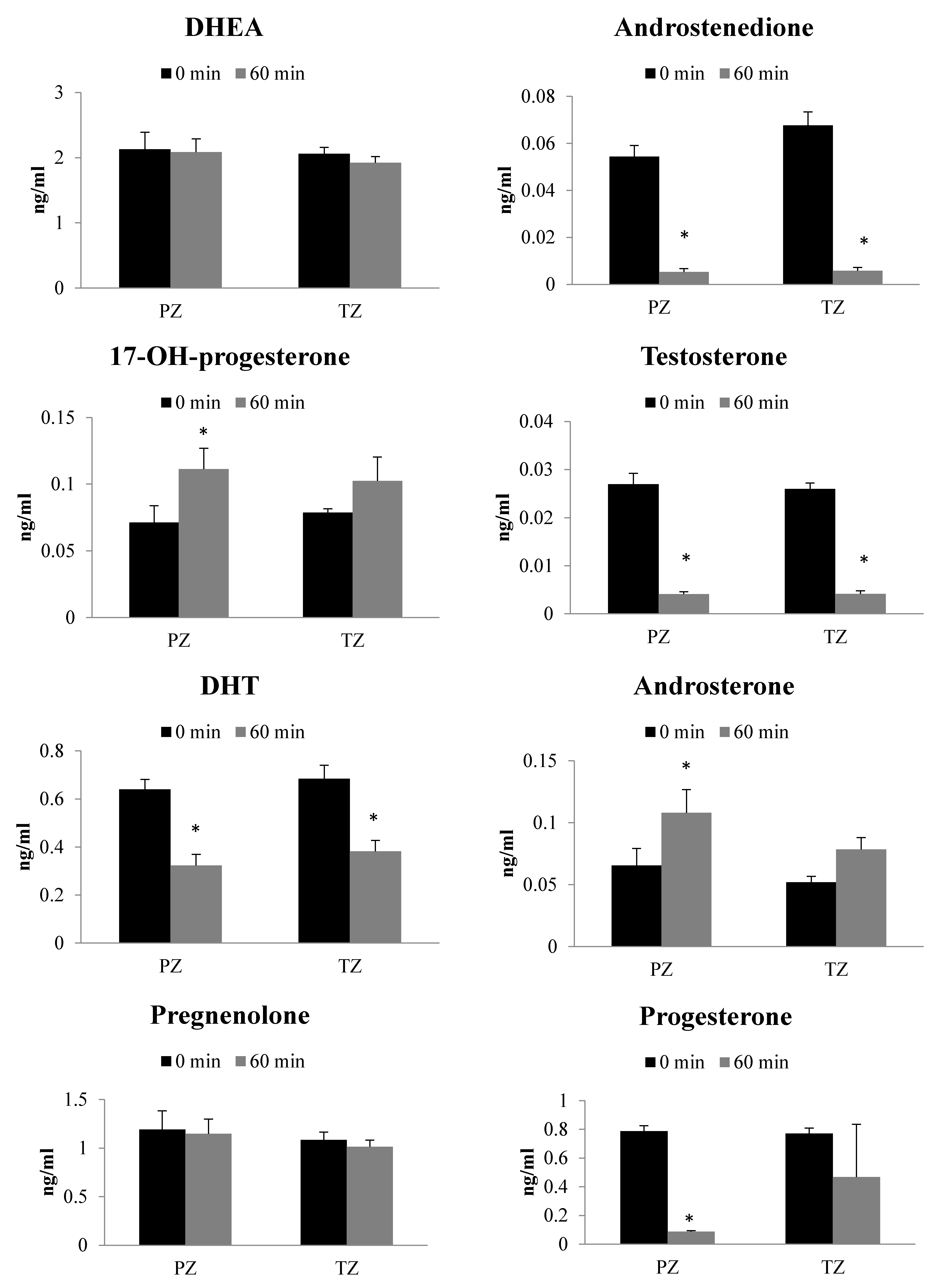
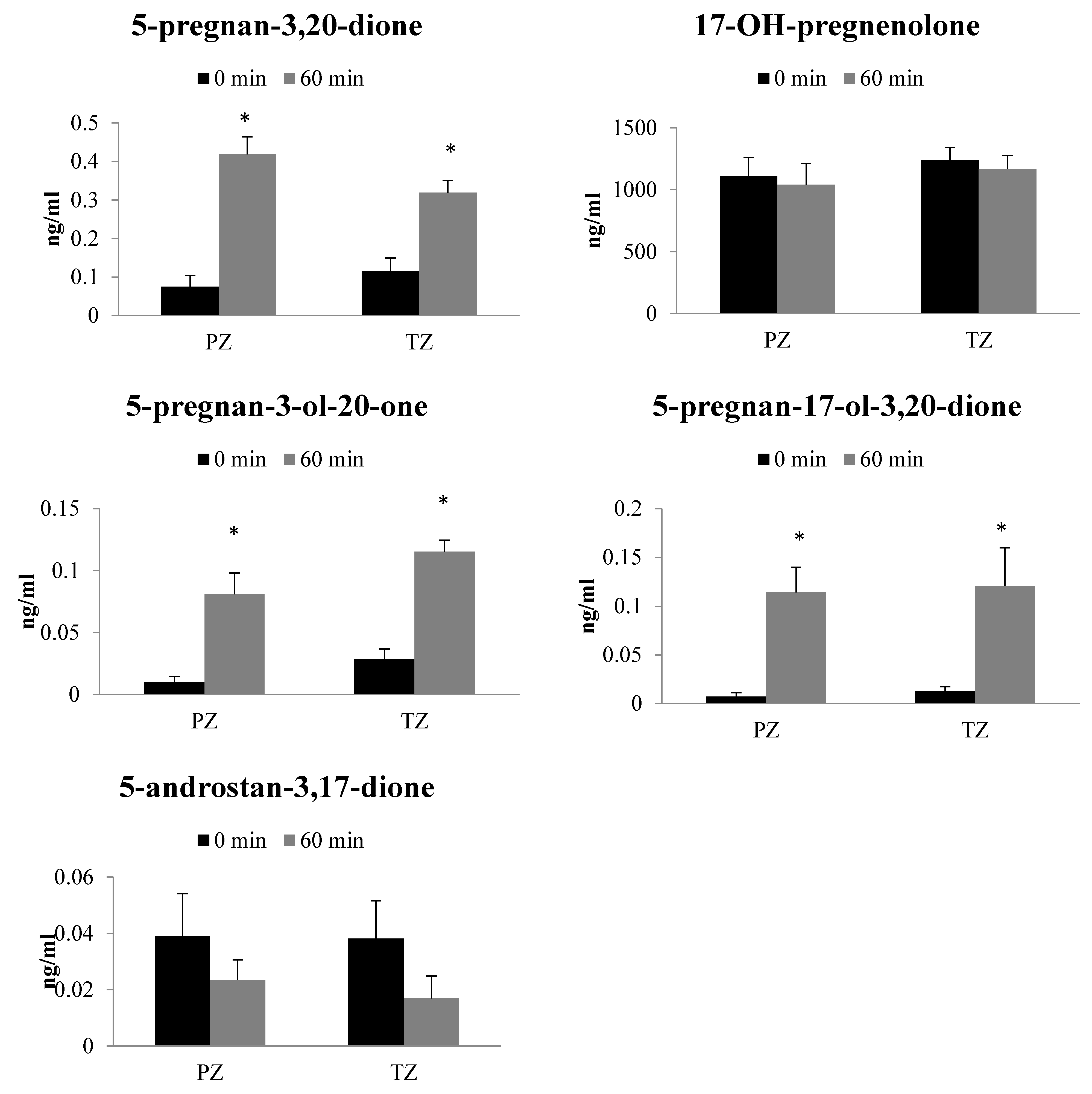
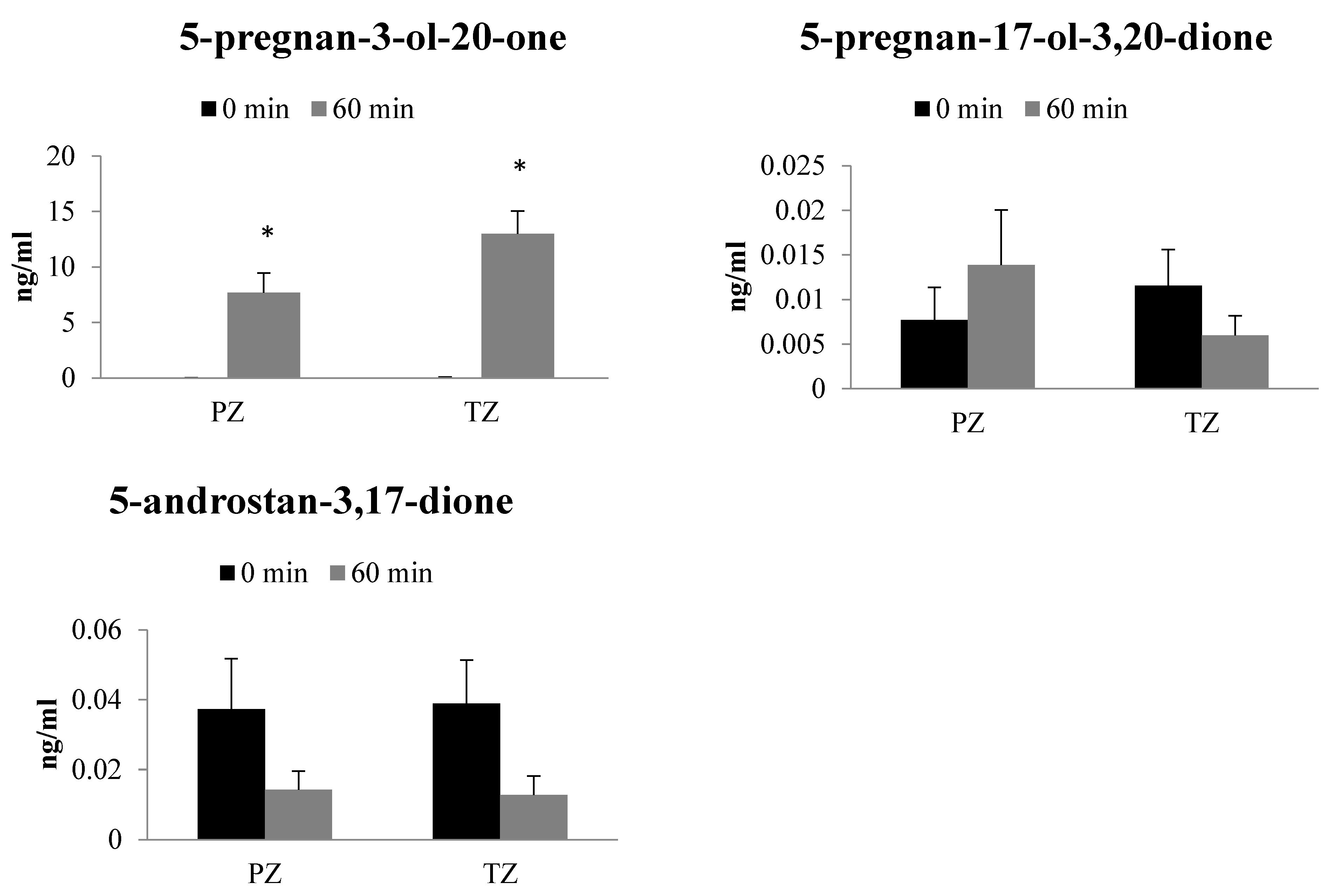
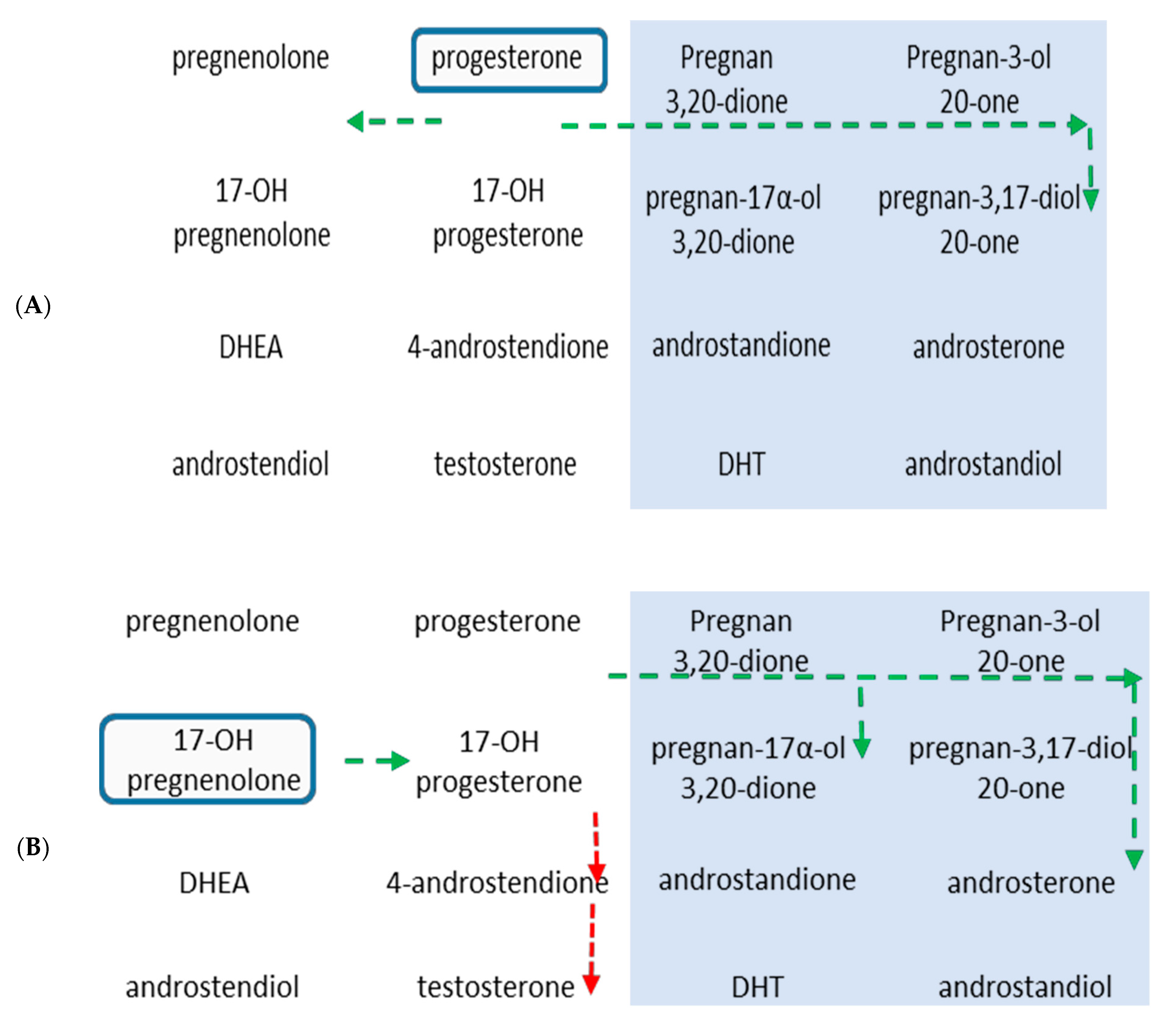
2.3. Metabolism of 17-OH-Pregnenolone in TZ and PZ Prostate Tissues
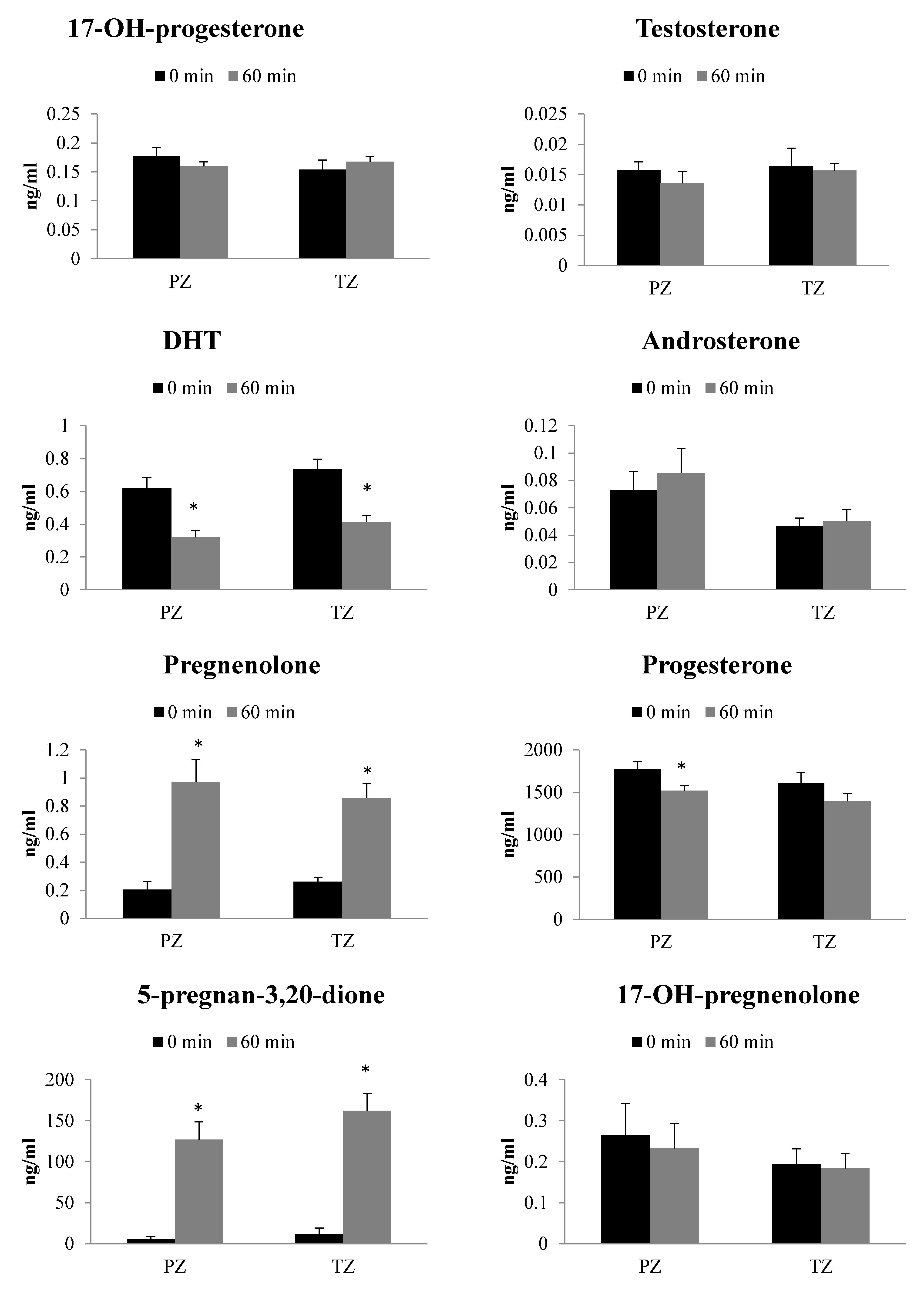
2.4. Metabolism of Progesterone in TZ and PZ Prostate Tissues
3. Discussion
4. Materials and Methods
4.1. Steroid Standards and Chemicals
4.2. Human Prostate Samples
4.3. Preparation of Human Prostate Homogenates
4.4. Steroid Biotransformation Assay with Human Prostate Homogenates
4.5. Analysis of Steroids by LC/MS
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| PZ | Peripheral zone |
| TZ | Transition zone |
| CRPC | Castration-resistant prostate cancer |
| PCa | Prostate cancer |
| LC/MS | Liquid chromatography-mass spectrometry |
| DHT | Dihydrotestosterone |
| HSD3B | 3β-hydroxysteroid dehydrogenase |
References
- Dunn, M.W.; Kazer, M.W. Prostate cancer overview. Semin. Oncol. Nurs. 2011, 27, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Akin-Olugbade, O.; Kirschenbaum, A. Overview of prostate anatomy, histology, and pathology. Endocrinol. Metab. Clin. N. Am. 2011, 40, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Thomas, I.C.; Nolley, R.; Ferrari, M.; Brooks, J.D.; Leppert, J.T. Biologic differences between peripheral and transition zone prostate cancer. Prostate 2015, 75, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Sinnott, J.A.; Rider, J.R.; Carlsson, J.; Gerke, T.; Tyekucheva, S.; Penney, K.L.; Sesso, H.D.; Loda, M.; Fall, K.; Stampfer, M.J.; et al. Molecular differences in transition zone and peripheral zone prostate tumors. Carcinogenesis 2015, 36, 632–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlsson, J.; Helenius, G.; Karlsson, M.G.; Andrén, O.; Klinga-Levan, K.; Olsson, B. Differences in microRNA expression during tumor development in the transition and peripheral zones of the prostate. BMC Cancer 2013, 13, 362. [Google Scholar] [CrossRef]
- Kumar, V.L.; Majumder, P.K. Prostate gland: Structure, functions and regulation. Int. Urol. Nephrol. 1995, 27, 231–243. [Google Scholar] [CrossRef]
- Heinlein, C.A.; Chang, C. Androgen receptor in prostate cancer. Endocr. Rev. 2004, 25, 276–308. [Google Scholar] [CrossRef] [Green Version]
- Crawford, E.D. Understanding the epidemiology, natural history, and key pathways involved in prostate cancer. Urology 2009, 73, S4–S10. [Google Scholar] [CrossRef]
- Zhu, M.-L.; Kyprianou, N. Role of androgens and the androgen receptor in epithelial-mesenchymal transition and invasion of prostate cancer cells. FASEB J. 2010, 24, 769–777. [Google Scholar] [CrossRef] [Green Version]
- Miyoshi, Y.; Uemura, H.; Umemoto, S.; Sakamaki, K.; Morita, S.; Suzuki, K.; Shibata, Y.; Masumori, N.; Ichikawa, T.; Mizokami, A.; et al. High testosterone levels in prostate tissue obtained by needle biopsy correlate with poor-prognosis factors in prostate cancer patients. BMC Cancer 2014, 14, 717. [Google Scholar] [CrossRef] [Green Version]
- Huggins, C.; Hodges, C.V. Studies on prostatic cancer: I. The effect of castration, of estrogen and of androgen injection on serum phosphatases in metastatic carcinoma of the prostate. J. Urol. 2002, 168, 9–12. [Google Scholar] [CrossRef]
- Mostaghel, E.A.; Nelson, P.S. Intracrine androgen metabolism in prostate cancer progression: Mechanisms of castration resistance and therapeutic implications. Best Pr. Res. Clin. Endocrinol. Metab. 2008, 22, 243–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirschenbaum, A.; Liu, X.-H.; Yao, S.; Narla, G.; Friedman, S.L.; Martignetti, J.A.; Levine, A.C. Sex steroids have differential effects on growth and gene expression in primary human prostatic epithelial cell cultures derived from the peripheral versus transition zones. Carcinogenesis 2006, 27, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekar, T.; Yang, J.C.; Gao, A.C.; Evans, C.P. Targeting molecular resistance in castration-resistant prostate cancer. BMC Med. 2015, 13, 206. [Google Scholar] [CrossRef] [Green Version]
- Mostaghel, E.A. Steroid hormone synthetic pathways in prostate cancer. Transl. Urol. 2013, 2, 212–227. [Google Scholar] [CrossRef]
- Bennett, N.C.; Hooper, J.D.; Lambie, D.; Lee, C.S.; Yang, T.; Vesey, D.A.; Samaratunga, H.; Johnson, D.W.; Gobe, G.C. Evidence for steroidogenic potential in human prostate cell lines and tissues. Am. J. Pathol. 2012, 181, 1078–1087. [Google Scholar] [CrossRef]
- Mitsiades, N.; Sung, C.C.; Schultz, N.; Danila, D.C.; He, B.; Eedunuri, V.K.; Fleisher, M.; Sander, C.; Sawyers, C.L.; Scher, H.I. Distinct patterns of dysregulated expression of enzymes involved in androgen synthesis and metabolism in metastatic prostate cancer tumors. Cancer Res. 2012, 72, 6142–6152. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, R.B.; Mostaghel, E.A.; Vessella, R.; Hess, D.L.; Kalhorn, T.F.; Higano, C.S.; True, L.D.; Nelson, P.S. Maintenance of intratumoral androgens in metastatic prostate cancer: A mechanism for castration-resistant tumor growth. Cancer Res. 2008, 68, 4447–4454. [Google Scholar] [CrossRef] [Green Version]
- Sharifi, N.; Auchus, R.J. Steroid biosynthesis and prostate cancer. Steroids 2012, 77, 719–726. [Google Scholar] [CrossRef]
- Locke, J.A.; Guns, E.S.; Lubik, A.A.; Adomat, H.H.; Hendy, S.C.; Wood, C.A.; Ettinger, S.L.; Gleave, M.E.; Nelson, C.C. Androgen levels increase by intratumoral de novo steroidogenesis during progression of castration-resistant prostate cancer. Cancer Res. 2008, 68, 6407–6415. [Google Scholar] [CrossRef] [Green Version]
- Lewis, S.R.; Hedman, C.J.; Ziegler, T.; Ricke, W.A.; Jorgensen, J.S. Steroidogenic factor 1 promotes aggressive growth of castration-resistant prostate cancer cells by stimulating steroid synthesis and cell proliferation. Endocrinology 2014, 155, 358–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, C.; Chen, S.; Ng, P.; Bubley, G.J.; Nelson, P.S.; Mostaghel, E.A.; Marck, B.; Matsumoto, A.M.; Simon, N.I.; Wang, H.; et al. Intratumoral de novo steroid synthesis activates androgen receptor in castration-resistant prostate cancer and is upregulated by treatment with CYP17A1 inhibitors. Cancer Res. 2011, 71, 6503–6513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferraldeschi, R.; Sharifi, N.; Auchus, R.J.; Attard, G. Molecular pathways: Inhibiting steroid biosynthesis in prostate cancer. Clin. Cancer Res. 2013, 19, 3353–3359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, S.M.; Mostaghel, E.A.; Nelson, P.S. Androgen action and metabolism in prostate cancer. Mol. Cell Endocrinol. 2012, 360, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luu-The, V. Assessment of steroidogenesis and steroidogenic enzyme functions. J. Steroid Biochem. Mol. Biol. 2013, 137, 176–182. [Google Scholar] [CrossRef]
- Chang, K.-H.; Li, R.; Papari-Zareei, M.; Watumull, L.; Zhao, Y.D.; Auchus, R.J.; Sharifi, N. Dihydrotestosterone synthesis bypasses testosterone to drive castration-resistant prostate cancer. Proc. Natl. Acad. Sci. USA 2011, 108, 13728–13733. [Google Scholar] [CrossRef] [Green Version]
- Hofland, J.; van Weerden, W.M.; Dits, N.F.J.; Steenbergen, J.; van Leenders, G.J.L.H.; Jenster, G.; Schröder, F.H.; de Jong, F.H. Evidence of limited contributions for intratumoral steroidogenesis in prostate cancer. Cancer Res. 2010, 70, 1256–1264. [Google Scholar] [CrossRef] [Green Version]
- Jeong, C.W.; Yoon, C.Y.; Jeong, S.J.; Hong, S.K.; Byun, S.-S.; Lee, S.E. Limited expression of cytochrome p450 17α-hydroxylase/17,20-lyase in prostate cancer cell lines. Korean J. Urol. 2011, 52, 494–497. [Google Scholar] [CrossRef] [Green Version]
- Deb, S.; Pham, S.; Ming, D.S.; Chin, M.Y.; Adomat, H.; Hurtado-Coll, A.; Gleave, M.E.; Guns, E.S.T. Characterization of precursor-dependent steroidogenesis in human prostate cancer models. Cancers 2018, 10, 343. [Google Scholar] [CrossRef] [Green Version]
- Yun, J.W.; Lee, S.; Ryu, D.; Park, S.; Park, W.Y.; Joung, J.G.; Jeong, J. Biomarkers associated with tumor heterogeneity in prostate cancer. Transl. Oncol. 2019, 12, 43–48. [Google Scholar] [CrossRef]
- Le Goff, J.M.; Martin, P.M.; Ojasoo, T.; Raynaud, J.P. Non-michaelian behavior of 5 alpha-reductase in human prostate. J. Steroid Biochem. 1989, 33, 155–163. [Google Scholar] [CrossRef]
- Sakai, M.; Martinez-Arguelles, D.B.; Aprikian, A.G.; Magliocco, A.M.; Papadopoulos, V. De novo steroid biosynthesis in human prostate cell lines and biopsies. Prostate 2016, 76, 575–587. [Google Scholar] [CrossRef]
- Fankhauser, M.; Tan, Y.; Macintyre, G.; Haviv, I.; Hong, M.K.H.; Nguyen, A.; Pedersen, J.S.; Costello, A.J.; Hovens, C.M.; Corcoran, N.M. Canonical androstenedione reduction is the predominant source of signaling androgens in hormone-refractory prostate cancer. Clin. Cancer Res. 2014, 20, 5547–5557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishizaki, F.; Nishiyama, T.; Kawasaki, T.; Miyashiro, Y.; Hara, N.; Takizawa, I.; Naito, M.; Takahashi, K. Androgen deprivation promotes intratumoral synthesis of dihydrotestosterone from androgen metabolites in prostate cancer. Sci. Rep. 2013, 3, 1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohler, J.L.; Titus, M.A.; Wilson, E.M. Potential prostate cancer drug target: Bioactivation of androstanediol by conversion to dihydrotestosterone. Clin. Cancer Res. 2011, 17, 5844–5849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamae, D.; Mostaghel, E.; Montgomery, B.; Nelson, P.S.; Balk, S.P.; Kantoff, P.W.; Taplin, M.-E.; Penning, T.M. The DHEA-sulfate depot following P450c17 inhibition supports the case for AKR1C3 inhibition in high risk localized and advanced castration resistant prostate cancer. Chem. Biol. Interact. 2015, 234, 332–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.-H.; Li, R.; Kuri, B.; Lotan, Y.; Roehrborn, C.G.; Liu, J.; Vessella, R.; Nelson, P.S.; Kapur, P.; Guo, X.; et al. A gain-of-function mutation in DHT synthesis in castration-resistant prostate cancer. Cell 2013, 154, 1074–1084. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, O.L.; Koff, W.J.; Muraro, F.; Santos, E.B.; Gomes Soares, D.F.; Trindade, V.M. Steroid 5-alpha reductase type 2 activity in biopsies from malignant and normal prostatic tissues. Clin. Chim. Acta 2008, 391, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Mizokami, A.; Koh, E.; Izumi, K.; Narimoto, K.; Takeda, M.; Honma, S.; Dai, J.; Keller, E.T.; Namiki, M. Prostate cancer stromal cells and LNCaP cells coordinately activate the androgen receptor through synthesis of testosterone and dihydrotestosterone from dehydroepiandrosterone. Endocr. Relat. Cancer 2009, 16, 1139–1155. [Google Scholar] [CrossRef] [Green Version]
- Mostaghel, E.A.; Marck, B.T.; Plymate, S.R.; Vessella, R.L.; Balk, S.; Matsumoto, A.M.; Nelson, P.S.; Montgomery, R.B. Resistance to CYP17A1 inhibition with abiraterone in castration-resistant prostate cancer: Induction of steroidogenesis and androgen receptor splice variants. Clin. Cancer Res. 2011, 17, 5913–5925. [Google Scholar] [CrossRef] [Green Version]
- Pham, S.; Deb, S.; Ming, D.S.; Adomat, H.; Hosseini-Beheshti, E.; Zoubeidi, A.; Gleave, M.; Guns, E.S.T. Next-generation steroidogenesis inhibitors, dutasteride and abiraterone, attenuate but still do not eliminate androgen biosynthesis in 22RV1 cells in vitro. J. Steroid Biochem. Mol. Biol. 2014, 144, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, N. The 5α-androstanedione pathway to dihydrotestosterone in castration-resistant prostate cancer. J. Investig. Med. 2012, 60, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Titus, M.A.; Li, Y.; Kozyreva, O.G.; Maher, V.; Godoy, A.; Smith, G.J.; Mohler, J.L. 5α-reductase type 3 enzyme in benign and malignant prostate. Prostate 2014, 74, 235–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adomat, H.H.; Bains, O.S.; Lubieniecka, J.M.; Gleave, M.E.; Guns, E.S.; Grigliatti, T.A.; Reid, R.E.; Riggs, K.W. Validation of a sequential extraction and liquid chromatography-tandem mass spectrometric method for determination of dihydrotestosterone, androstanediol and androstanediol-glucuronide in prostate tissues. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2012, 902, 84–95. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PZ Tissue | TZ Tissue | |||
|---|---|---|---|---|
| Avg (ng/mL) | SEM | Avg (ng/mL) | SEM | |
| Dehydroepiandrosterone | 0.514 | 0.221 | 0.342 | 0.092 |
| Androstenedione | 0.009 | 0.003 | 0.010 | 0.003 |
| 17-OH-progesterone | 0.002 | 0.001 | 0.007 | 0.003 |
| Testosterone | 0.010 | 0.002 | 0.013 | 0.005 |
| Dihydrotestosterone | 2.086 | 1.611 | 0.688 | 0.068 |
| Androsterone | 0.055 | 0.019 | 0.044 | 0.011 |
| Pregnenolone | 0.467 | 0.142 | 0.383 | 0.111 |
| Progesterone | 0.021 | 0.006 | 0.022 | 0.009 |
| 5-Pregnan-3,20-dione | 0.014 | 0.007 | 0.022 | 0.014 |
| 17-OH-pregnenolone | 0.228 | 0.085 | 0.175 | 0.065 |
| 5-pregnan-3-ol-20-one | 0.008 | 0.004 | 0.008 | 0.005 |
| 5-pregnan-17-ol-3,20-dione | 0.008 | 0.007 | 0.010 | 0.006 |
| 5-androstan-3,17-dione | 0.025 | 0.015 | 0.005 | 0.003 |
| 17-OH-Pregnenolone | Progesterone | |||
|---|---|---|---|---|
| PZ | TZ | PZ | TZ | |
| Dehydroepiandrosterone | 1.00 | 3.28 | 1.00 | 0.59 |
| Androstenedione | 1.00 | 1.26 | 1.00 | 0.34 |
| 17-OH-progesterone | 1.00 | 0.59 | 1.00 | 0.77 |
| Testosterone | 1.00 | 0.96 | 1.00 | 0.33 |
| Dihydrotestosterone | 1.00 | 0.95 | 1.00 | 1.08 |
| Androsterone | 1.00 | 0.62 | 1.00 | 0.30 |
| Pregnenolone | 1.00 | 1.60 | 1.00 | 0.78 |
| Progesterone | 1.00 | 0.43 | 1.00 | 0.84 |
| 5-Pregnan-3,20-dione | 1.00 | 0.59 | 1.00 | 1.25 |
| 17-OH-pregnenolone | 1.00 | 1.07 | 1.00 | 0.36 |
| 5-pregnan-3-ol-20-one | 1.00 | 1.22 | 1.00 | 1.69 |
| 5-pregnan-17-ol-3,20-dione | 1.00 | 1.01 | 1.00 | 0.91 |
| 5-androstan-3,17-dione | 1.00 | 1.36 | 1.00 | 1.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deb, S.; Chin, M.Y.; Pham, S.; Adomat, H.; Hurtado-Coll, A.; Gleave, M.E.; Tomlinson Guns, E.S. Steroidogenesis in Peripheral and Transition Zones of Human Prostate Cancer Tissue. Int. J. Mol. Sci. 2021, 22, 487. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020487
Deb S, Chin MY, Pham S, Adomat H, Hurtado-Coll A, Gleave ME, Tomlinson Guns ES. Steroidogenesis in Peripheral and Transition Zones of Human Prostate Cancer Tissue. International Journal of Molecular Sciences. 2021; 22(2):487. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020487
Chicago/Turabian StyleDeb, Subrata, Mei Yieng Chin, Steven Pham, Hans Adomat, Antonio Hurtado-Coll, Martin E. Gleave, and Emma S. Tomlinson Guns. 2021. "Steroidogenesis in Peripheral and Transition Zones of Human Prostate Cancer Tissue" International Journal of Molecular Sciences 22, no. 2: 487. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020487