Human Primary Dermal Fibroblasts Interacting with 3-Dimensional Matrices for Surgical Application Show Specific Growth and Gene Expression Programs

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Growth of Fibroblasts on the Different 3D Matrices
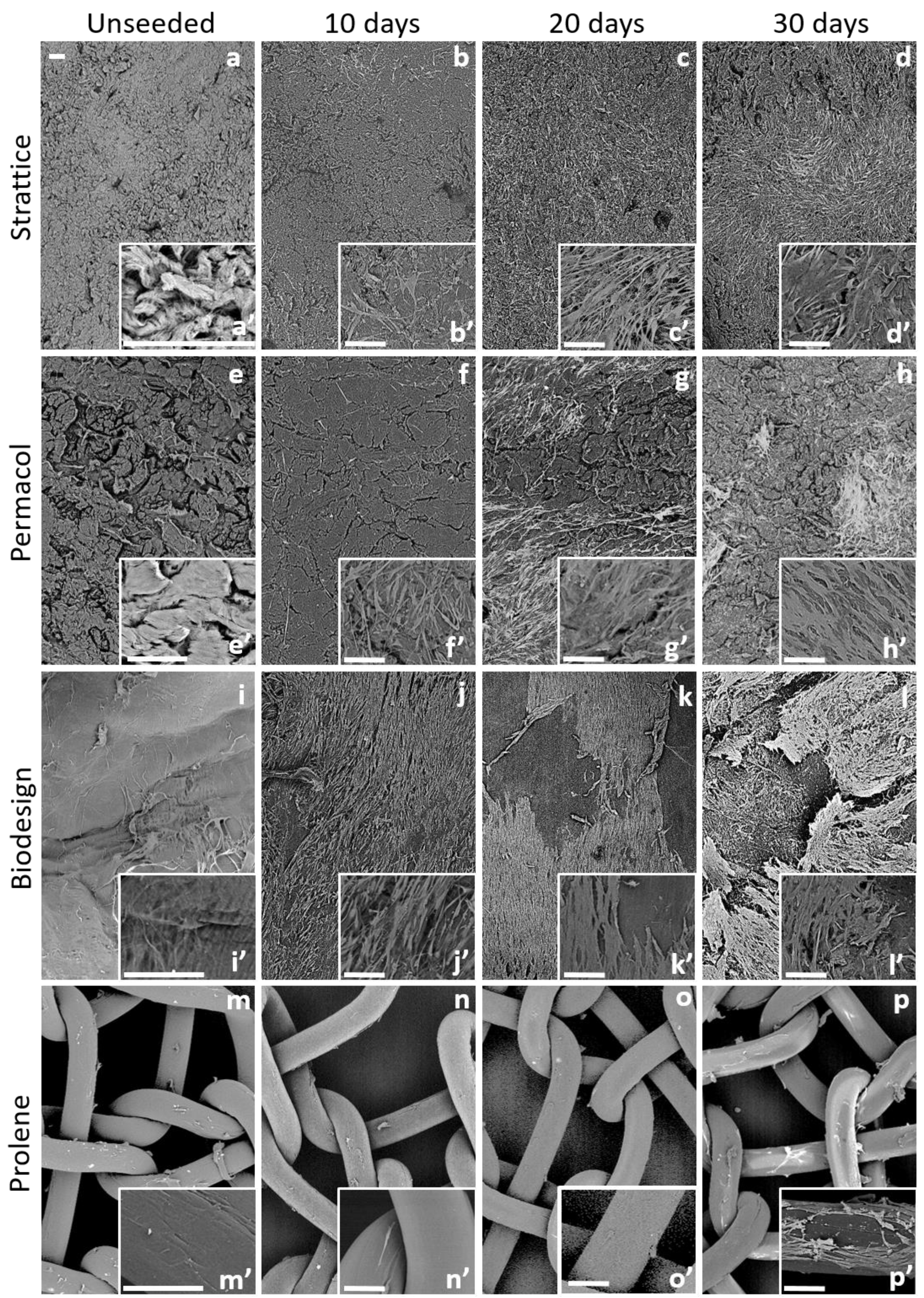
2.1.1. Morphological Examination by SEM
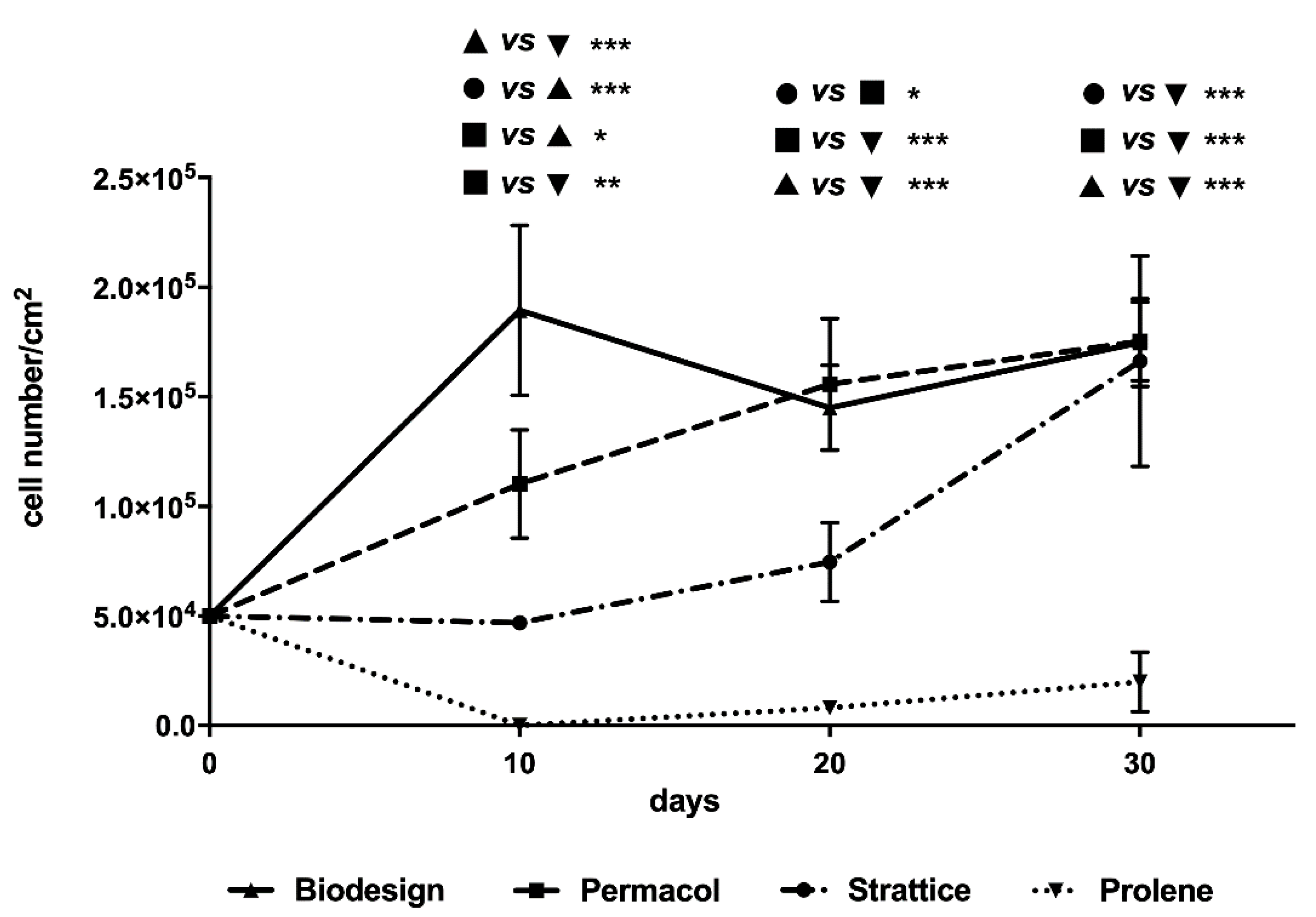
2.1.2. Cell Growth Curves
2.2. Gene and Protein Expression
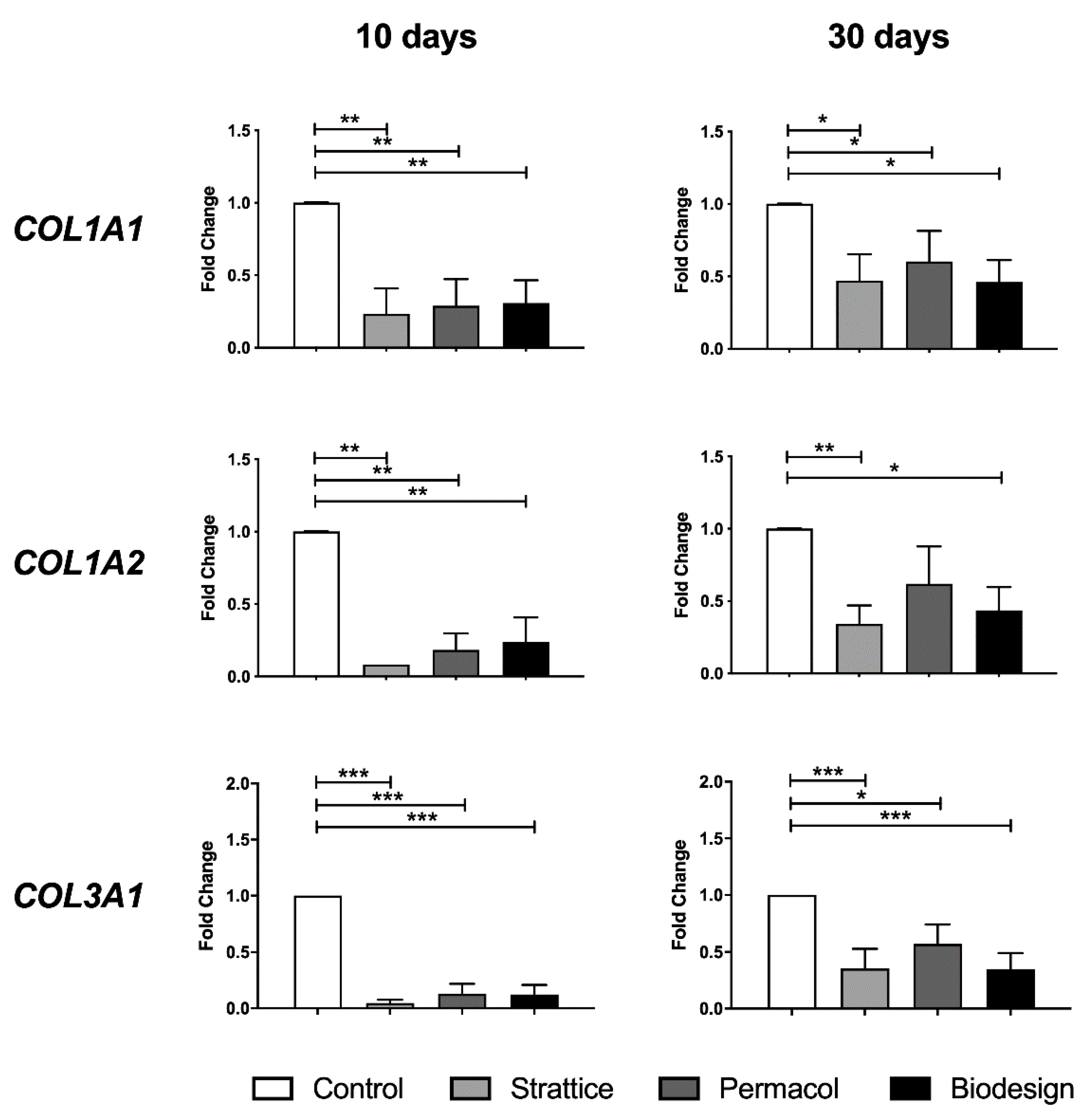
2.2.1. Expression of Collagen Genes
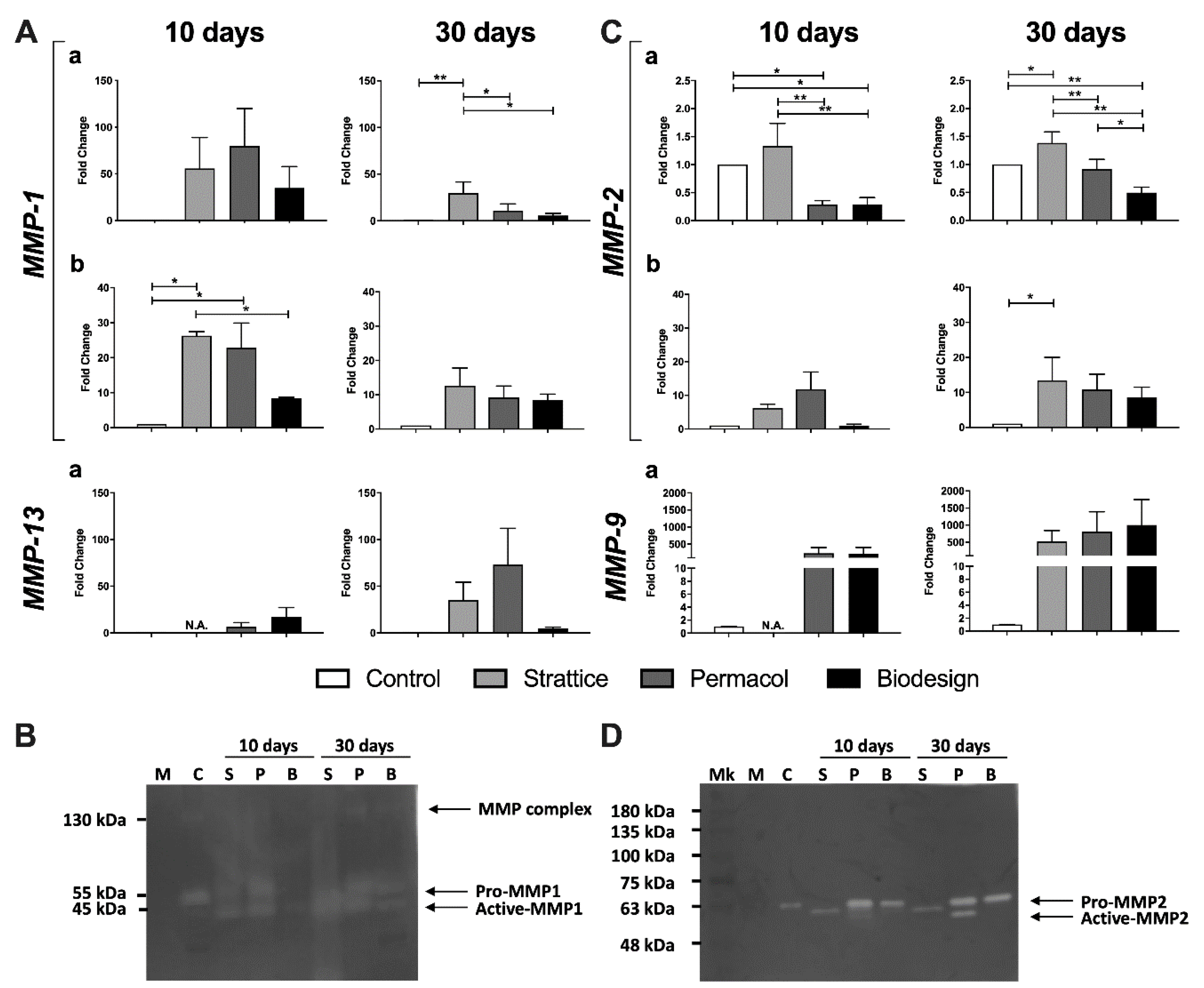
2.2.2. Expression and Activity of Extracellular Matrix Degrading Enzymes
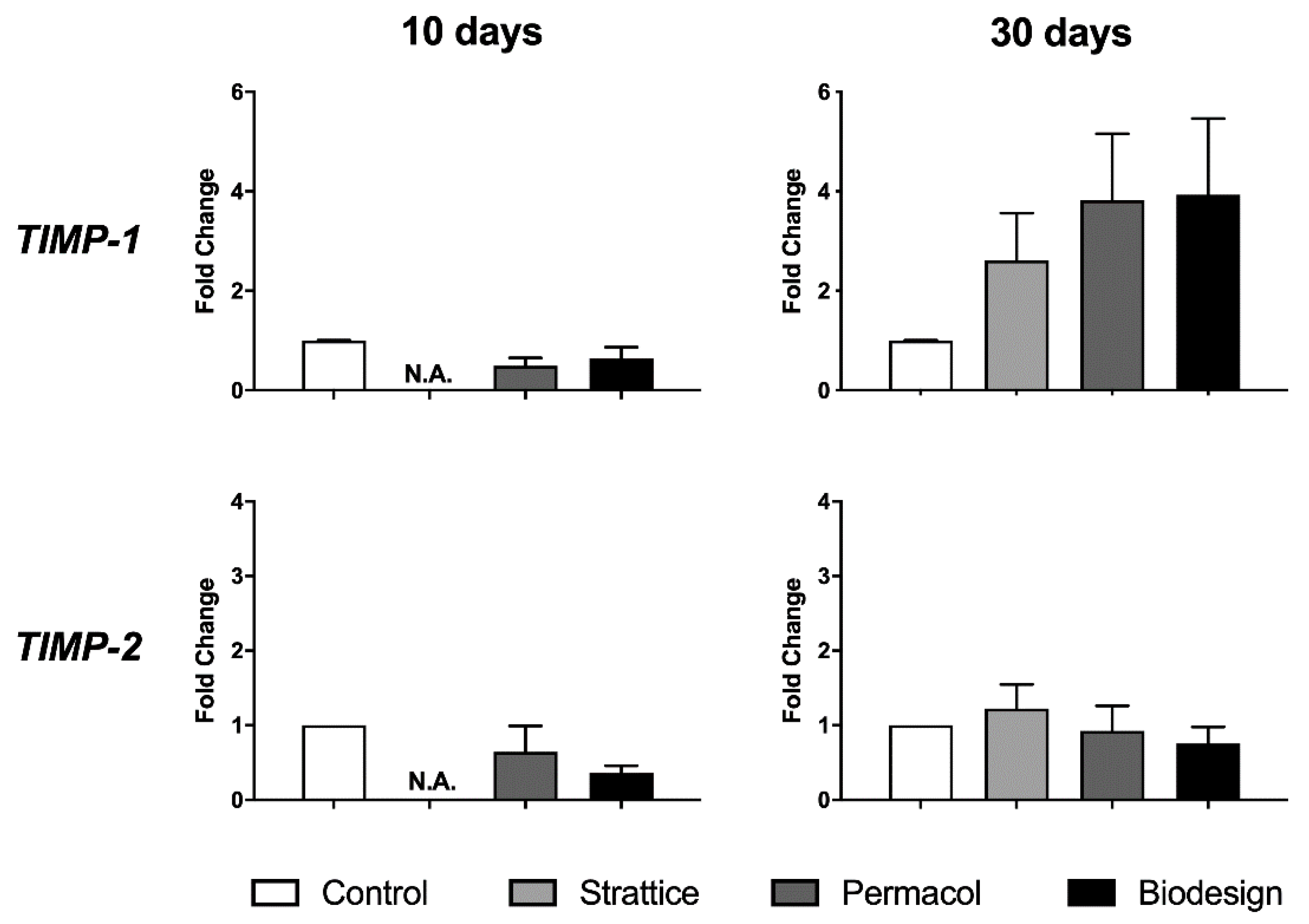
2.2.3. Expression of Metalloproteinase Inhibitors
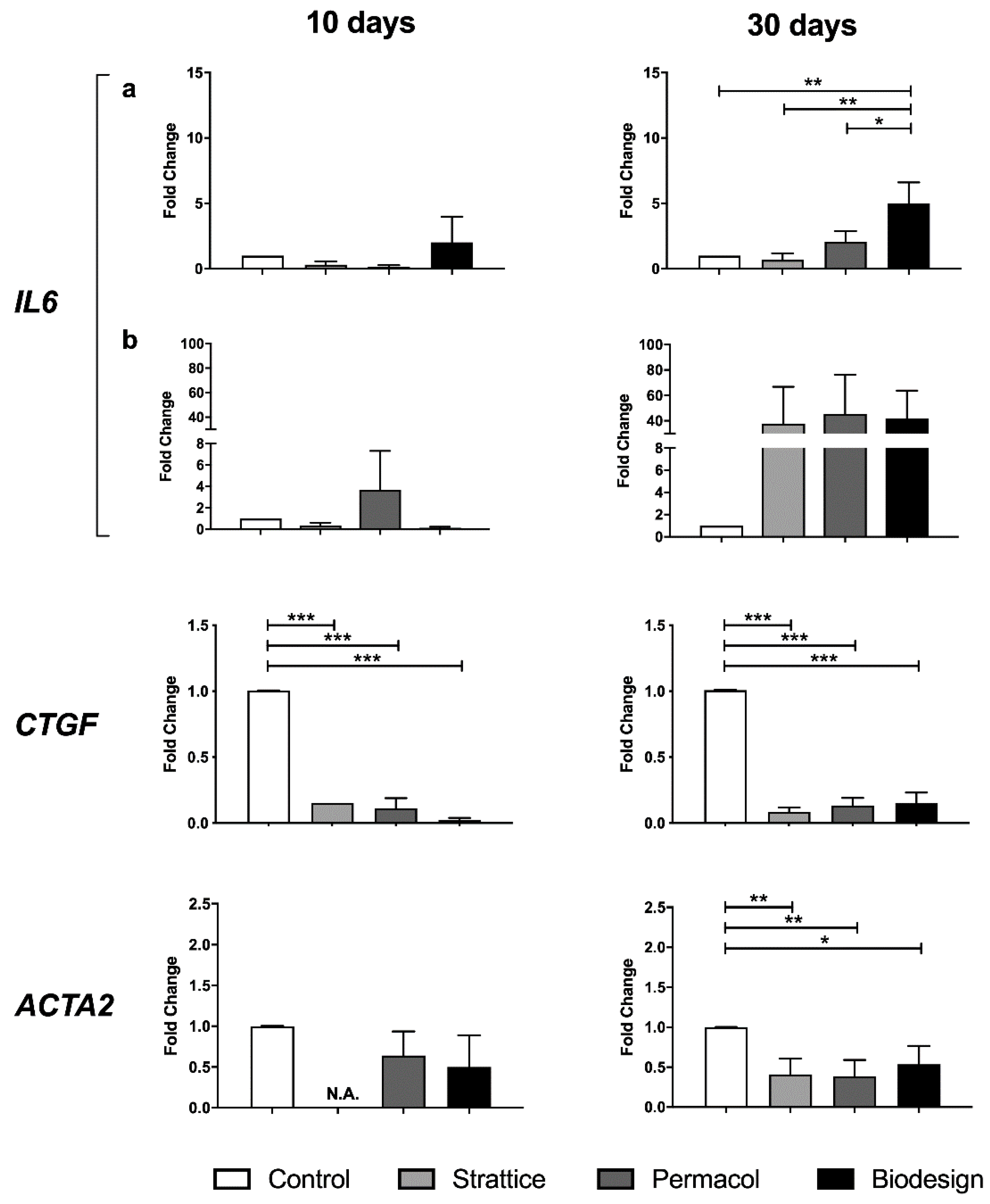
2.2.4. Expression of Cytokines and Alpha-Smooth Muscle Actin (α-SMA)
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Type and Preparation of Matrices
4.3. Experimental Setup
4.4. Scanning Electron Microscopy (SEM)
4.5. Cell Counting
4.6. RNA Extraction and qPCR Analysis
4.7. Quantification of Secreted Proteins by ELISA
4.8. Zymography Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 2D | Two dimensional |
| 3D | Three dimensional |
| α-SMA | Actin, alpha 2, smooth muscle protein |
| ACTA2 | Actin, alpha 2, smooth muscle gene |
| ANOVA | Analysis of variance |
| B2M | Beta-2-microglobulin |
| COL1A1 | CollCollagen type I alpha 1 chain |
| COL1A2 | CollCollagen type I alpha 2 chain |
| COL3A1 | Collagen type III alpha 1 chain |
| Cq | Cycle quantitative (PCR cycle number at which the amplification curve intersects the threshold line) |
| CTGF | Connective tissue growth factor |
| ECM | Extracellular matrix |
| ELISA | Enzyme-linked immunosorbent assay |
| FBR | Foreign body response |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| IL-6 | Interleukin 6 |
| MMP | Metalloproteinase |
| MSC | Mesenchymal stem cell |
| SEM | Scanning Electron Microscopy |
| TIMP | Metallopeptidase inhibitor |
References
- Ansaloni, L.; Catena, F.; Coccolini, F.; Gazzotti, F.; D’Alessandro, L.; Pinna, A.D. Inguinal hernia repair with porcine small intestine submucosa: 3-year follow-up results of a randomized controlled trial of Lichtenstein’s repair with polypropylene mesh versus Surgisis Inguinal Hernia Matrix. Am. J. Surg. 2009, 198, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Ellis, H.L.; Asaolu, O.; Nebo, V.; Kasem, A. Biological and synthetic mesh use in breast reconstructive surgery: A literature review. World J. Surg. Oncol. 2016, 14, 1–9. [Google Scholar]
- Junge, K.; Binnebosel, M.; von Trotha, K.T.; Rosch, R.; Klinge, U.; Neumann, U.P.; Lynen Jansen, P. Mesh biocompatibility: Effects of cellular inflammation and tissue remodelling. Langenbeck’s Arch. Surg. 2012, 397, 255–270. [Google Scholar] [CrossRef] [PubMed]
- FitzGerald, J.F.; Kumar, A.S. Biologic versus Synthetic Mesh Reinforcement: What are the Pros and Cons? Clin. Colon Rectal Surg. 2014, 27, 140–148. [Google Scholar]
- Deerenberg, E.B.; Mulder, I.M.; Grotenhuis, N.; Ditzel, M.; Jeekel, J.; Lange, J.F. Experimental study on synthetic and biological mesh implantation in a contaminated environment. Br. J. Surg. 2012, 99, 1734–1741. [Google Scholar] [CrossRef]
- Sandor, M.; Xu, H.; Connor, J.; Lombardi, J.; Harper, J.R.; Silverman, R.P.; McQuillan, D.J. Host response to implanted porcine-derived biologic materials in a primate model of abdominal wall repair. Tissue Eng. Part A 2008, 14, 2021–2031. [Google Scholar] [CrossRef]
- Shankaran, V.; Weber, D.J.; Reed, R.L., 2nd; Luchette, F.A. A review of available prosthetics for ventral hernia repair. Ann. Surg. 2011, 253, 16–26. [Google Scholar] [CrossRef]
- Cevasco, M.; Itani, K.M. Ventral hernia repair with synthetic, composite, and biologic mesh: Characteristics, indications, and infection profile. Surg. Infect. (Larchmt) 2012, 13, 209–215. [Google Scholar] [CrossRef]
- El-Hayek, K.M.; Chand, B. Biologic prosthetic materials for hernia repairs. J. Long Term Eff. Med. Implants 2010, 20, 159–169. [Google Scholar] [CrossRef]
- Sadava, E.E.; Krpata, D.M.; Gao, Y.; Rosen, M.J.; Novitsky, Y.W. Wound healing process and mediators: Implications for modulations for hernia repair and mesh integration. J. Biomed. Mater. Res. A 2014, 102, 295–302. [Google Scholar] [CrossRef]
- Rastegarpour, A.; Cheung, M.; Vardhan, M.; Ibrahim, M.M.; Butler, C.E.; Levinson, H. Surgical mesh for ventral incisional hernia repairs: Understanding mesh design. Plast Surg. (Oakv) 2016, 24, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Baylon, K.; Rodriguez-Camarillo, P.; Elias-Zuniga, A.; Diaz-Elizondo, J.A.; Gilkerson, R.; Lozano, K. Past, Present and Future of Surgical Meshes: A Review. Membranes 2017, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Ventral Hernia Working, G.; Breuing, K.; Butler, C.E.; Ferzoco, S.; Franz, M.; Hultman, C.S.; Kilbridge, J.F.; Rosen, M.; Silverman, R.P.; Vargo, D. Incisional ventral hernias: Review of the literature and recommendations regarding the grading and technique of repair. Surgery 2010, 148, 544–558. [Google Scholar]
- Novitsky, Y.W.; Rosen, M.J. The biology of biologics: Basic science and clinical concepts. Plast. Reconstr. Surg. 2012, 130 (Suppl. 2), 9S–17S. [Google Scholar] [CrossRef] [PubMed]
- Nilsen, T.J.; Dasgupta, A.; Huang, Y.C.; Wilson, H.; Chnari, E. Do Processing Methods Make a Difference in Acellular Dermal Matrix Properties? Aesthet. Surg. J. 2016, 36 (Suppl. 2), S7–S22. [Google Scholar] [CrossRef]
- Sandor, M.; Leamy, P.; Assan, P.; Hoonjan, A.; Huang, L.T.; Edwards, M.; Zuo, W.; Li, H.; Xu, H. Relevant In Vitro Predictors of Human Acellular Dermal Matrix-Associated Inflammation and Capsule Formation in a Nonhuman Primate Subcutaneous Tissue Expander Model. Eplasty 2017, 17, e1. [Google Scholar]
- Deeken, C.R.; Eliason, B.J.; Pichert, M.D.; Grant, S.A.; Frisella, M.M.; Matthews, B.D. Differentiation of biologic scaffold materials through physicomechanical, thermal, and enzymatic degradation techniques. Ann. Surg. 2012, 255, 595–604. [Google Scholar] [CrossRef]
- Dubay, D.A.; Wang, X.; Kirk, S.; Adamson, B.; Robson, M.C.; Franz, M.G. Fascial fibroblast kinetic activity is increased during abdominal wall repair compared to dermal fibroblasts. Wound Repair Regen. 2004, 12, 539–545. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, L.J.; Blatnik, J.A.; Krpata, D.M.; Anderson, J.M.; Criss, C.N.; Posielski, N.; Novitsky, Y.W. Methodology of fibroblast and mesenchymal stem cell coating of surgical meshes: A pilot analysis. J. Biomed. Mater. Res. B Appl. Biomater. 2014, 102, 797–805. [Google Scholar] [CrossRef]
- Gigante, A.; Cesari, E.; Busilacchi, A.; Manzotti, S.; Kyriakidou, K.; Greco, F.; Di Primio, R.; Mattioli-Belmonte, M. Collagen I membranes for tendon repair: Effect of collagen fiber orientation on cell behavior. J. Orthop. Res. 2009, 27, 826–832. [Google Scholar] [CrossRef]
- Zhong, S.; Teo, W.E.; Zhu, X.; Beuerman, R.W.; Ramakrishna, S.; Yung, L.Y. An aligned nanofibrous collagen scaffold by electrospinning and its effects on in vitro fibroblast culture. J. Biomed. Mater. Res. A 2006, 79, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Bashur, C.A.; Dahlgren, L.A.; Goldstein, A.S. Effect of fiber diameter and orientation on fibroblast morphology and proliferation on electrospun poly(D,L-lactic-co-glycolic acid) meshes. Biomaterials 2006, 27, 5681–5688. [Google Scholar] [CrossRef]
- Delaine-Smith, R.M.; Green, N.H.; Matcher, S.J.; MacNeil, S.; Reilly, G.C. Monitoring fibrous scaffold guidance of three-dimensional collagen organisation using minimally-invasive second harmonic generation. PLoS ONE 2014, 9, e89761. [Google Scholar] [CrossRef]
- Novitsky, Y.W.; Orenstein, S.B.; Kreutzer, D.L. Comparative analysis of histopathologic responses to implanted porcine biologic meshes. Hernia 2014, 18, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Lipson, K.E.; Wong, C.; Teng, Y.; Spong, S. CTGF is a central mediator of tissue remodeling and fibrosis and its inhibition can reverse the process of fibrosis. Fibrogenesis Tissue Repair. 2012, 5 (Suppl. 1), S24. [Google Scholar] [CrossRef] [Green Version]
- Helary, C.; Foucault-Bertaud, A.; Godeau, G.; Coulomb, B.; Guille, M.M. Fibroblast populated dense collagen matrices: Cell migration, cell density and metalloproteinases expression. Biomaterials 2005, 26, 1533–1543. [Google Scholar] [CrossRef] [PubMed]
- Kramann, R.; DiRocco, D.P.; Humphreys, B.D. Understanding the origin, activation and regulation of matrix-producing myofibroblasts for treatment of fibrotic disease. J. Pathol. 2013, 231, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Chen, B.; Liu, T.; Chen, X. Reversal of myofibroblast differentiation: A review. Eur. J. Pharmacol. 2014, 734, 83–90. [Google Scholar] [CrossRef]
- Grotendorst, G.R.; Duncan, M.R. Individual domains of connective tissue growth factor regulate fibroblast proliferation and myofibroblast differentiation. FASEB J. 2005, 19, 729–738. [Google Scholar] [CrossRef]
- Brigstock, D.R. Connective tissue growth factor (CCN2, CTGF) and organ fibrosis: Lessons from transgenic animals. J. Cell Commun. Signal. 2010, 4, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.N.; Finch, J.G. Which mesh for hernia repair? Ann. R. Coll. Surg. Engl. 2010, 92, 272–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bornstein, P.; Sage, E.H. Matricellular proteins: Extracellular modulators of cell function. Curr. Opin. Cell Biol. 2002, 14, 608–616. [Google Scholar] [CrossRef]
- Dasu, M.R.; Barrow, R.E.; Spies, M.; Herndon, D.N. Matrix metalloproteinase expression in cytokine stimulated human dermal fibroblasts. Burns 2003, 29, 527–531. [Google Scholar] [CrossRef]
- Mauviel, A. Cytokine regulation of metalloproteinase gene expression. J. Cell Biochem. 1993, 53, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A. Common and unique mechanisms regulate fibrosis in various fibroproliferative diseases. J. Clin. Investig. 2007, 117, 524–529. [Google Scholar] [CrossRef] [Green Version]
- Orenstein, S.B.; Qiao, Y.; Kaur, M.; Klueh, U.; Kreutzer, D.L.; Novitsky, Y.W. Human monocyte activation by biologic and biodegradable meshes in vitro. Surg. Endosc. 2010, 24, 805–811. [Google Scholar] [CrossRef]
- Orenstein, S.B.; Qiao, Y.; Klueh, U.; Kreutzer, D.L.; Novitsky, Y.W. Activation of human mononuclear cells by porcine biologic meshes in vitro. Hernia 2010, 14, 401–407. [Google Scholar] [CrossRef]
- Fries, K.M.; Felch, M.E.; Phipps, R.P. Interleukin-6 is an autocrine growth factor for murine lung fibroblast subsets. Am. J. Respir. Cell Mol. Biol. 1994, 11, 552–560. [Google Scholar] [CrossRef]
- Nguyen, H.N.; Noss, E.H.; Mizoguchi, F.; Huppertz, C.; Wei, K.S.; Watts, G.F.M.; Brenner, M.B. Autocrine Loop Involving IL-6 Family Member LIF, LIF Receptor, and STAT4 Drives Sustained Fibroblast Production of Inflammatory Mediators. Immunity 2017, 46, 220–232. [Google Scholar] [CrossRef] [Green Version]
- Butler, C.E.; Burns, N.K.; Campbell, K.T.; Mathur, A.B.; Jaffari, M.V.; Rios, C.N. Comparison of cross-linked and non-cross-linked porcine acellular dermal matrices for ventral hernia repair. J. Am. Coll. Surg. 2010, 211, 368–376. [Google Scholar] [CrossRef]
- Deeken, C.R.; Melman, L.; Jenkins, E.D.; Greco, S.C.; Frisella, M.M.; Matthews, B.D. Histologic and biomechanical evaluation of crosslinked and non-crosslinked biologic meshes in a porcine model of ventral incisional hernia repair. J. Am. Coll. Surg. 2011, 212, 880–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Silva, G.S.; Krpata, D.M.; Gao, Y.; Criss, C.N.; Anderson, J.M.; Soltanian, H.T.; Rosen, M.J.; Novitsky, Y.W. Lack of identifiable biologic behavior in a series of porcine mesh explants. Surgery 2014, 156, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Filocamo, M.M.R.; Corsolini, F.; Stroppiano, M.; Stroppiana, G.; Grossi, S.; Lualdi, S.; Tappino, B.; Lanza, F.; Galotto, S.; Biancheri, R. Cell Line and DNA Biobank From Patients Affected by Genetic Diseases. Open J. Bioresour. 2014, 1, e2. [Google Scholar]
- Tang, J.; Hu, M.; Lee, S.; Roblin, R. A polymerase chain reaction based method for detecting Mycoplasma/Acholeplasma contaminants in cell culture. J. Microbiol. Methods 2000, 39, 121–126. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trade Name | Manufacturer | Species | Tissue Type | Crosslinked | Sterilization |
|---|---|---|---|---|---|
| Strattice | LifeCell-Acelity | Porcine | Dermis | No | E-beam |
| Permacol | Covidien | Porcine | Dermis | Yes (hexamethylene diisocyanate) | Gamma irradiation |
| Surgisis-Biodesign | Cook-Medical | Porcine | Small intestine submucosa | No | Ethylene oxide |
| Prolene Polypropylene Mesh | Ethicon | Synthetic | Polypropylene high weight, monofilament | N.A. (a) | Ethylene oxide |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grossi, S.; Grimaldi, A.; Congiu, T.; Parnigoni, A.; Campanelli, G.; Campomenosi, P. Human Primary Dermal Fibroblasts Interacting with 3-Dimensional Matrices for Surgical Application Show Specific Growth and Gene Expression Programs. Int. J. Mol. Sci. 2021, 22, 526. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020526
Grossi S, Grimaldi A, Congiu T, Parnigoni A, Campanelli G, Campomenosi P. Human Primary Dermal Fibroblasts Interacting with 3-Dimensional Matrices for Surgical Application Show Specific Growth and Gene Expression Programs. International Journal of Molecular Sciences. 2021; 22(2):526. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020526
Chicago/Turabian StyleGrossi, Sarah, Annalisa Grimaldi, Terenzio Congiu, Arianna Parnigoni, Giampiero Campanelli, and Paola Campomenosi. 2021. "Human Primary Dermal Fibroblasts Interacting with 3-Dimensional Matrices for Surgical Application Show Specific Growth and Gene Expression Programs" International Journal of Molecular Sciences 22, no. 2: 526. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020526