Comprehensive Analysis of Cardiac Xeno-Graft Unveils Rejection Mechanisms

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Results
2.1. Recipient Monkey Blood Analysis Showed Systemic Inflammatory Responses at Rejection
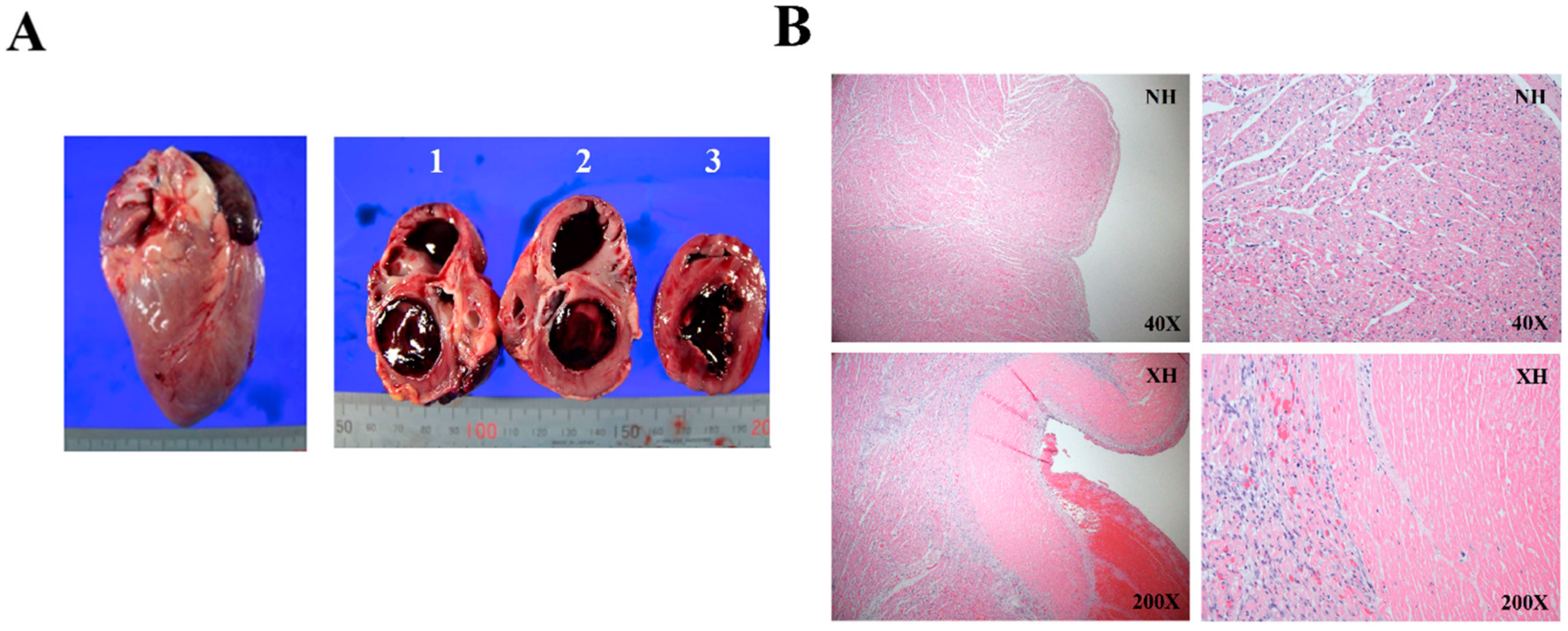
2.2. Histological Assessment of Rejected Xenograft Showed Myocardial Infarction and Fibrosis
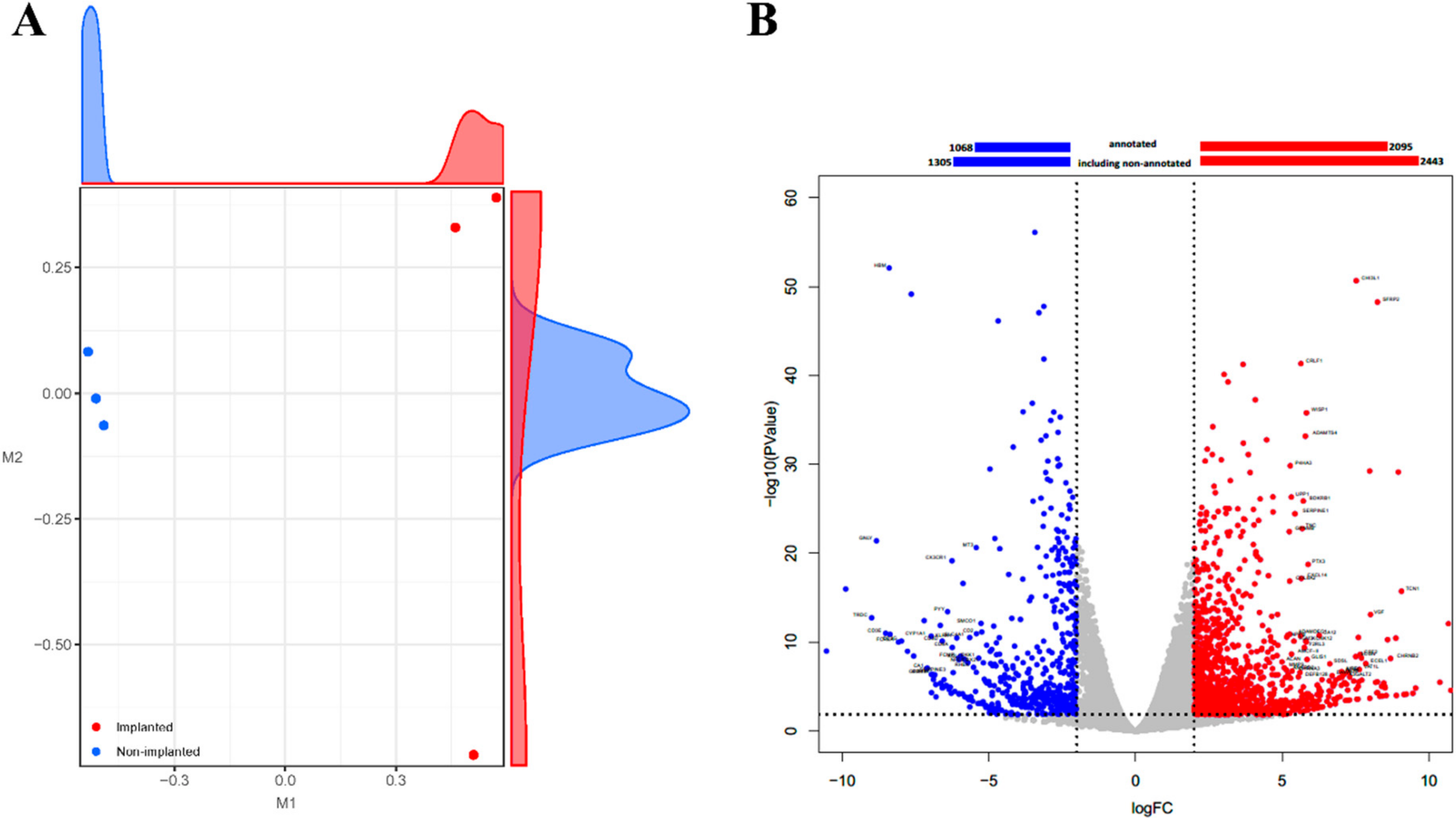
2.3. Transcriptome Analysis Showed Alterations of a Number of Genes in a Rejected Xenograft
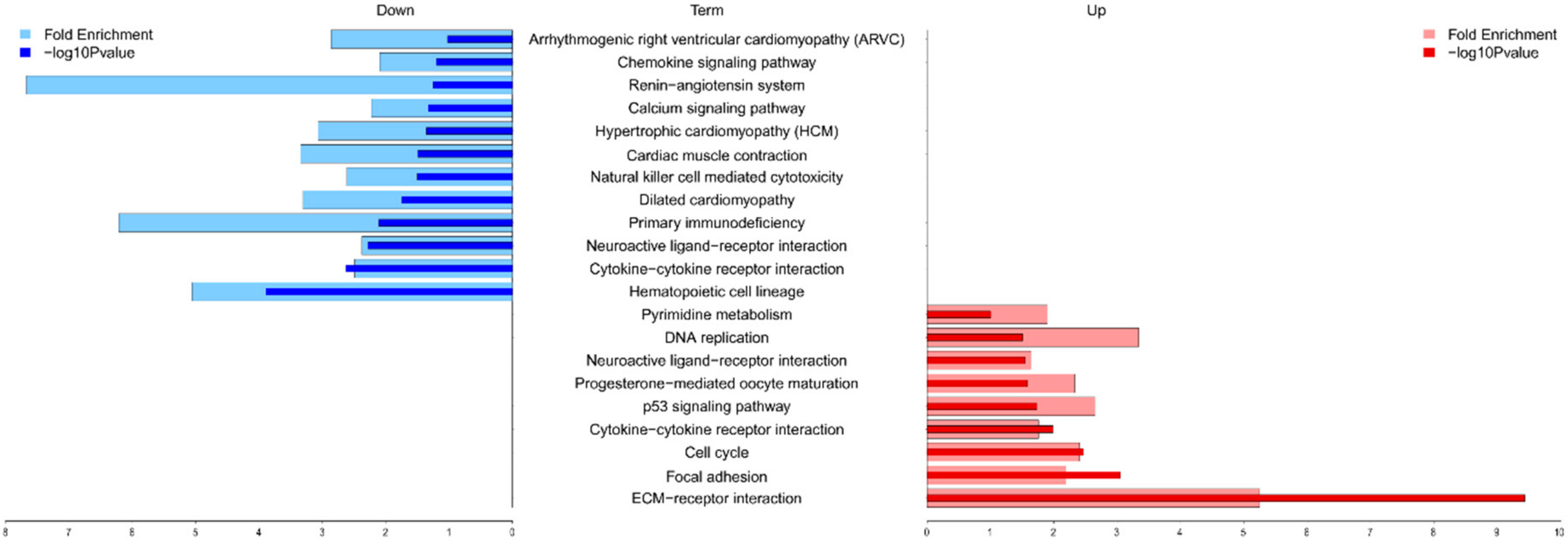
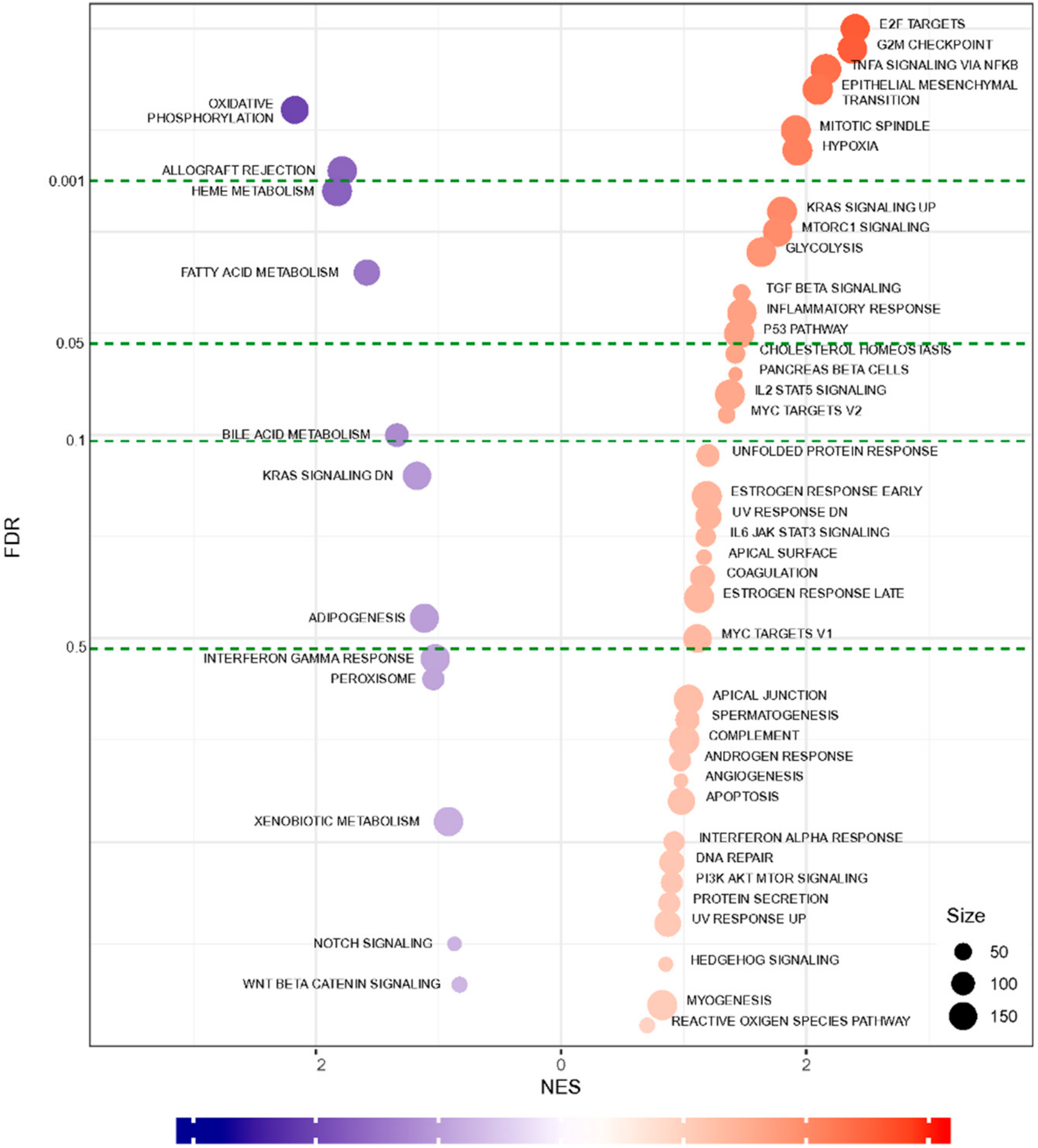
2.4. Functional Annotation and Gene Set Enrichment Analysis (GSEA) Suggested Multiple Molecular Mechanisms Involved in Graft Rejection
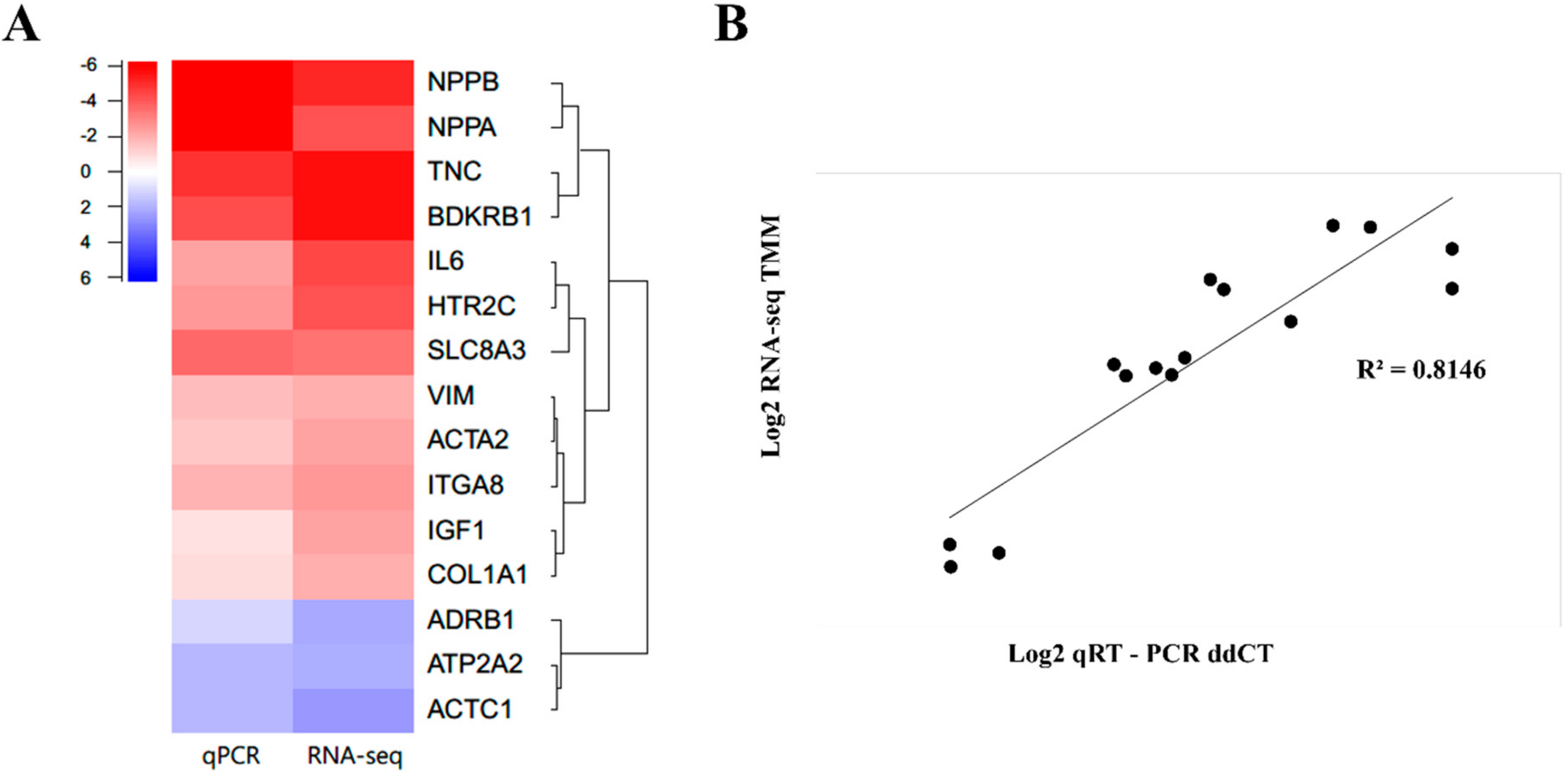
2.5. qPCR Validation of Randomly Selected DEGs Confirmed a Similar Expression Trend
3. Discussion
4. Methods
4.1. Ethics Statements
4.2. Experimental Animals and Heterotopic Transplantation Procedure
4.3. Immunosuppressive Regimen
4.4. Hematological and Biochemical Analysis of a Cynomolgus Monkey
4.5. Histological Analysis
4.6. RNA Sequencing Preparation
4.7. Transcriptome Alignment and Differentially Expressed Genes (DEGs) Analysis of the Porcine Cardiac Tissue
4.8. Gene Ontology and Pathway Enrichment Analysis
4.9. Gene Set Enrichment Analysis (GSEA)
4.10. Quantitative Real-Time PCR (qPCR)
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halloran, K.M.; Parkes, M.D.; Chang, J.; Timofte, I.L.; Snell, G.I.; Westall, G.P.; Hachem, R.; Kreisel, D.; Trulock, E.; Roux, A.; et al. Molecular assessment of rejection and injury in lung transplant biopsies. J. Heart Lung Transpl. 2019, 38, 504–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasr, M.; Sigdel, T.; Sarwal, M. Advances in diagnostics for transplant rejection. Expert Rev. Mol. Diagn. 2016, 16, 1121–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langin, M.; Mayr, T.; Reichart, B.; Michel, S.; Buchholz, S.; Guethoff, S.; Dashkevich, A.; Baehr, A.; Egerer, S.; Bauer, A.; et al. Consistent success in life-supporting porcine cardiac xenotransplantation. Nature 2018, 564, 430–433. [Google Scholar] [CrossRef] [PubMed]
- Mohiuddin, M.M.; Singh, A.K.; Corcoran, P.C.; Thomas, M.L., III; Clark, T.; Lewis, B.G.; Hoyt, R.F.; Eckhaus, M.; Pierson, R.N., III; Belli, A.J. Chimeric 2C10R4 anti-CD40 antibody therapy is critical for long-term survival of GTKO. hCD46. hTBM pig-to-primate cardiac xenograft. Nat. Commun. 2016, 7, 11138. [Google Scholar] [CrossRef] [PubMed]
- Van den Hoogen, P.; Huibers, M.M.; Sluijter, J.P.; de Weger, R.A. Cardiac allograft vasculopathy: A donor or recipient induced pathology? J. Cardiovasc. Transl. Res. 2015, 8, 106–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Fueyo, A.; Strom, T.B. Immunologic basis of graft rejection and tolerance following transplantation of liver or other solid organs. Gastroenterology 2011, 140, 51–64. [Google Scholar] [CrossRef] [Green Version]
- Herrer, I.; Rosello-Lleti, E.; Ortega, A.; Tarazon, E.; Molina-Navarro, M.M.; Trivino, J.C.; Martinez-Dolz, L.; Almenar, L.; Lago, F.; Sanchez-Lazaro, I.; et al. Gene expression network analysis reveals new transcriptional regulators as novel factors in human ischemic cardiomyopathy. BMC Med. Genom. 2015, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Sweet, M.E.; Cocciolo, A.; Slavov, D.; Jones, K.L.; Sweet, J.R.; Graw, S.L.; Reece, T.B.; Ambardekar, A.V.; Bristow, M.R.; Mestroni, L.; et al. Transcriptome analysis of human heart failure reveals dysregulated cell adhesion in dilated cardiomyopathy and activated immune pathways in ischemic heart failure. BMC Genom. 2018, 19, 812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, K.S.; Kim, Y.J.; Kim, M.; Lee, B.H.; Heo, S.Y.; Kang, M.J.; Kang, Y.K.; Lee, J.W.; Lee, K.K.; Kim, J.H.; et al. Resurrection of an alpha-1,3-galactosyltransferase gene-targeted miniature pig by recloning using postmortem ear skin fibroblasts. Theriogenology 2011, 75, 933–939. [Google Scholar] [CrossRef]
- Van Gelder, T.; Balk, A.H.M.M.; Zondervan, P.E.; Maat, A.W.P.M.; Mochtar, B.; van der Meer, P.; Weimar, W. C-reactive protein in the monitoring of acute rejection after heart transplantation. Transpl. Int. 1998, 11, 361–364. [Google Scholar] [CrossRef]
- Scozzi, D.; Ibrahim, M.; Menna, C.; Krupnick, A.S.; Kreisel, D.; Gelman, A.E. The Role of Neutrophils in Transplanted Organs. Am. J. Transpl. 2017, 17, 328–335. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Soriano, R.M.; Almenar, L.; Martínez-Dolz, L.; Reganon, E.; Martínez-Sales, V.; Chamorro, C.I.; Vila, V.; Martín-Pastor, J.; Villa, P.; Salvador, A. Diagnostic usefulness of inflammatory markers in acute cellular rejection after heart transplantation. Transpl. Proc. 2006, 38, 2569–2571. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Morley, M.; Brandimarto, J.; Hannenhalli, S.; Hu, Y.; Ashley, E.A.; Tang, W.H.; Moravec, C.S.; Margulies, K.B.; Cappola, T.P.; et al. RNA-Seq identifies novel myocardial gene expression signatures of heart failure. Genomics 2015, 105, 83–89. [Google Scholar] [CrossRef]
- Li, X.; Liu, C.Y.; Li, Y.S.; Xu, J.; Li, D.G.; Li, X.; Han, D. Deep RNA sequencing elucidates microRNA-regulated molecular pathways in ischemic cardiomyopathy and nonischemic cardiomyopathy. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Spivey, T.L.; Uccellini, L.; Ascierto, M.L.; Zoppoli, G.; De Giorgi, V.; Delogu, L.G.; Engle, A.M.; Thomas, J.M.; Wang, E.; Marincola, F.M.; et al. Gene expression profiling in acute allograft rejection: Challenging the immunologic constant of rejection hypothesis. J. Transl. Med. 2011, 9, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giancotti, F.G.; Ruoslahti, E. Integrin signaling. Science 1999, 285, 1028–1032. [Google Scholar] [CrossRef]
- Howard, C.M.; Baudino, T.A. Dynamic cell–cell and cell–ECM interactions in the heart. J. Mol. Cell. Cardiol. 2014, 70, 19–26. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. The Extracellular Matrix in Ischemic and Nonischemic Heart Failure. Circ. Res. 2019, 125, 117–146. [Google Scholar] [CrossRef] [PubMed]
- Gil-Cayuela, C.; Rivera, M.; Ortega, A.; Tarazón, E.; Triviño, J.C.; Lago, F.; González-Juanatey, J.R.; Almenar, L.; Martínez-Dolz, L.; Portolés, M. RNA sequencing analysis identifies new human collagen genes involved in cardiac remodeling. J. Am. Coll. Cardiol. 2015, 65, 1265–1267. [Google Scholar] [CrossRef] [Green Version]
- Roberts, D.D. Emerging functions of matricellular proteins. Cell Mol. Life Sci. 2011, 68, 3133–3136. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Dobaczewski, M.; Gonzalez-Quesada, C.; Chen, W.; Biernacka, A.; Li, N.; Lee, D.-W.; Frangogiannis, N.G. Endogenous thrombospondin 1 protects the pressure-overloaded myocardium by modulating fibroblast phenotype and matrix metabolism. Hypertension 2011, 58, 902–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Quesada, C.; Cavalera, M.; Biernacka, A.; Kong, P.; Lee, D.-W.; Saxena, A.; Frunza, O.; Dobaczewski, M.; Shinde, A.; Frangogiannis, N.G. Thrombospondin-1 induction in the diabetic myocardium stabilizes the cardiac matrix in addition to promoting vascular rarefaction through angiopoietin-2 upregulation. Circ. Res. 2013, 113, 1331–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sezaki, S.; Hirohata, S.; Iwabu, A.; Nakamura, K.; Toeda, K.; Miyoshi, T.; Yamawaki, H.; Demircan, K.; Kusachi, S.; Shiratori, Y.; et al. Thrombospondin-1 is induced in rat myocardial infarction and its induction is accelerated by ischemia/reperfusion. Exp. Biol. Med. 2005, 230, 621–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frolova, E.G.; Sopko, N.; Blech, L.; Popović, Z.B.; Li, J.; Vasanji, A.; Drumm, C.; Krukovets, I.; Jain, M.K.; Penn, M.S. Thrombospondin-4 regulates fibrosis and remodeling of the myocardium in response to pressure overload. FASEB J. 2012, 26, 2363–2373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, J.M.; Maillet, M.; Vanhoutte, D.; Schloemer, A.; Sargent, M.A.; Blair, N.S.; Lynch, K.A.; Okada, T.; Aronow, B.J.; Osinska, H. A thrombospondin-dependent pathway for a protective ER stress response. Cell 2012, 149, 1257–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imanaka-Yoshida, K.; Hiroe, M. Tenascin-C Regulates Fibrosis, Inflammation and Immunological Response. J. Card. Fail. 2009, 15, S143. [Google Scholar] [CrossRef]
- Nishioka, T.; Onishi, K.; Shimojo, N.; Nagano, Y.; Matsusaka, H.; Ikeuchi, M.; Ide, T.; Tsutsui, H.; Hiroe, M.; Yoshida, T.; et al. Tenascin-C may aggravate left ventricular remodeling and function after myocardial infarction in mice. Am. J. Physiol. Heart Circ. Physiol. 2010, 298, H1072–H1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imanaka-Yoshida, K.; Aoki, H. Tenascin-C and mechanotransduction in the development and diseases of cardiovascular system. Front. Physiol. 2014, 5, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyon, R.C.; Zanella, F.; Omens, J.H.; Sheikh, F. Mechanotransduction in cardiac hypertrophy and failure. Circ. Res. 2015, 116, 1462–1476. [Google Scholar] [CrossRef] [Green Version]
- Maron, B.J.; Towbin, J.A.; Thiene, G.; Antzelevitch, C.; Corrado, D.; Arnett, D.; Moss, A.J.; Seidman, C.E.; Young, J.B.; American Heart, A.; et al. Contemporary definitions and classification of the cardiomyopathies: An American Heart Association Scientific Statement from the Council on Clinical Cardiology, Heart Failure and Transplantation Committee; Quality of Care and Outcomes Research and Functional Genomics and Translational Biology Interdisciplinary Working Groups; and Council on Epidemiology and Prevention. Circulation 2006, 113, 1807–1816. [Google Scholar] [CrossRef] [Green Version]
- Sisakian, H. Cardiomyopathies: Evolution of pathogenesis concepts and potential for new therapies. World J. Cardiol. 2014, 6, 478–494. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, J.; Gordon, H.; Hollman, A.; Bishop, M. Clinical aspects of cardiomyopathy. Br. Med. J. 1961, 1, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, M.; Anderson, M.E. Mechanisms of altered Ca2⁺ handling in heart failure. Circ. Res. 2013, 113, 690–708. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Gareri, C.; Rockman, H.A. G-Protein-Coupled Receptors in Heart Disease. Circ. Res. 2018, 123, 716–735. [Google Scholar] [CrossRef] [PubMed]
- De Lucia, C.; Eguchi, A.; Koch, W.J. New Insights in Cardiac beta-Adrenergic Signaling During Heart Failure and Aging. Front. Pharm. 2018, 9, 904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, T.D.; Swigart, P.M.; Rodrigo, M.C.; Ishizaka, S.; Joho, S.; Turnbull, L.; Tecott, L.H.; Baker, A.J.; Foster, E.; Grossman, W.; et al. Alpha1-adrenergic receptors prevent a maladaptive cardiac response to pressure overload. J. Clin. Invest. 2006, 116, 1005–1015. [Google Scholar] [CrossRef]
- Eisner, D.A.; Caldwell, J.L.; Kistamás, K.; Trafford, A.W. Calcium and excitation-contraction coupling in the heart. Circ. Res. 2017, 121, 181–195. [Google Scholar] [CrossRef]
- Rahm, A.-K.; Lugenbiel, P.; Schweizer, P.A.; Katus, H.A.; Thomas, D. Role of ion channels in heart failure and channelopathies. Biophys. Rev. 2018, 10, 1097–1106. [Google Scholar] [CrossRef]
- Stewart, S.; Winters, G.L.; Fishbein, M.C.; Tazelaar, H.D.; Kobashigawa, J.; Abrams, J.; Andersen, C.B.; Angelini, A.; Berry, G.J.; Burke, M.M.; et al. Revision of the 1990 working formulation for the standardization of nomenclature in the diagnosis of heart rejection. J. Heart Lung Transpl. 2005, 24, 1710–1720. [Google Scholar] [CrossRef]
- Fenton, M.; Simmonds, J.; Shah, V.; Brogan, P.; Klein, N.; Deanfield, J.; Burch, M. Inflammatory Cytokines, Endothelial Function, and Chronic Allograft Vasculopathy in Children: An Investigation of the Donor and Recipient Vasculature After Heart Transplantation. Am. J. Transpl. 2016, 16, 1559–1568. [Google Scholar] [CrossRef] [Green Version]
- Horwitz, P.A.; Tsai, E.J.; Putt, M.E.; Gilmore, J.M.; Lepore, J.J.; Parmacek, M.S.; Kao, A.C.; Desai, S.S.; Goldberg, L.R.; Brozena, S.C.; et al. Detection of cardiac allograft rejection and response to immunosuppressive therapy with peripheral blood gene expression. Circulation 2004, 110, 3815–3821. [Google Scholar] [CrossRef] [PubMed]
- Chih, S.; Chruscinski, A.; Ross, H.J.; Tinckam, K.; Butany, J.; Rao, V. Antibody-Mediated Rejection: An Evolving Entity in Heart Transplantation. J. Transpl. 2012, 210210. [Google Scholar] [CrossRef]
- Tarazón, E.; Ortega, A.; Gil-Cayuela, C.; Sánchez-Lacuesta, E.; Marín, P.; Lago, F.; González-Juanatey, J.R.; Martínez-Dolz, L.; Portolés, M.; Rivera, M.; et al. SERCA2a: A potential non-invasive biomarker of cardiac allograft rejection. J. Heart Lung Transpl. 2017, 36, 1322–1328. [Google Scholar] [CrossRef] [Green Version]
- Dhar, K.; Moulton, A.M.; Rome, E.; Qiu, F.; Kittrell, J.; Raichlin, E.; Zolty, R.; Um, J.Y.; Moulton, M.J.; Basma, H.; et al. Targeted myocardial gene expression in failing hearts by RNA sequencing. J. Transl. Med. 2016, 14, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.S.; Chee, H.K.; Kim, J.S.; Kim, W.S.; Park, J.H.; Shin, K.C.; Park, K.S.; Lee, S.W.; Cho, K.H.; Park, W.J.; et al. Non-invasive myocardial strain imaging to evaluate graft rejection in the cardiac xenotransplantation. J. Korean Soc. Transpl. 2017, 31, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Bioinformatics, Babraham Institute: Cambridge, UK, 2010. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2013, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Tan, Q.; Kir, J.; Liu, D.; Bryant, D.; Guo, Y.; Stephens, R.; Baseler, M.W.; Lane, H.C. DAVID Bioinformatics Resources: Expanded annotation database and novel algorithms to better extract biology from large gene lists. Nucleic Acids Res. 2007, 35, W169–W175. [Google Scholar] [CrossRef]
- Consortium, G.O. The Gene Ontology (GO) database and informatics resource. Nucleic Acids Res. 2004, 32, D258–D261. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Post-Operation Day | ||||
|---|---|---|---|---|---|
| −7 | −3 | 1 | 4 | 7 | |
| Calcium (mg/dL) | 10.20 | 10.20 | 9.00 | 9.00 | 9.40 |
| Phosphorus (mg/dL) | 6.40 | 5.10 | 5.90 | 5.30 | 4.50 |
| BUN (mg/dL) | 29.00 | 25.00 | 32.00 | 39.00 | 35.00 |
| Uric acid (mg/dL) | 0.30 | 0.30 | <0.10 | 0.20 | 0.10 |
| Creatinine (mg/dL) | 45.00 | 1.08 | 1.09 | 0.92 | 0.83 |
| Na (mEq/L) | 143.00 | 147.00 | 149.00 | 143.00 | 142.00 |
| K (mEq/L) | 4.90 | 5.10 | 5.10 | 5.30 | 5.80 |
| Cl (mEq/L) | 100.00 | 106.00 | 111.00 | 109.00 | 103.00 |
| TCO2 (mmol/L) | 15.00 | 19.00 | 23.00 | 21.00 | 23.00 |
| hs-CRP (mg/dL) | 0.12 | 0.20 | 6.72 | 1.14 | 13.68 |
| AST(GOT) (IU/L) | 53.00 | 51.00 | 1419.00 | 306.00 | 103.00 |
| ALT(GPT) (IU/L) | 85.00 | 79.00 | 255.00 | 157.00 | 94.00 |
| Total protein (g/dL) | 6.90 | 7.30 | 6.10 | 5.60 | 5.70 |
| Albumin (g/dL) | 3.90 | 4.20 | 3.30 | 3.00 | 2.80 |
| TBIL (mg/dL) | 0.40 | 0.40 | 0.40 | 0.40 | 0.30 |
| Alk. Phos (IU/L) | 513.00 | 466.00 | 400.00 | 332.00 | 317.00 |
| Chol (mg/dL) | 114.00 | 124.00 | 103.00 | 109.00 | 112.00 |
| GGT (IU/L) | 45.00 | 51.00 | 40.00 | 35.00 | 46.00 |
| LD(LDH) (IU/L) | 1997.00 | ||||
| Troponin (ng/mL) | 19.10 | ||||
| Tacrolimus (ng/mL) | <2.00 | 3.70 | 11.30 | ||
| WBC (×103 cells/uL) | 10.54 | 9.63 | 11.34 | 10.70 | 12.97 |
| RBC (×107 cells/uL) | 5.18 | 5.00 | 4.33 | 4.10 | 3.82 |
| Hb (g/dL) | 12.10 | 11.80 | 10.00 | 9.40 | 8.90 |
| Hct (%) | 40.50 | 38.90 | 33.20 | 32.70 | 29.30 |
| MCV (fL) | 78.20 | 77.70 | 76.70 | 79.80 | 76.80 |
| MCH (pg) | 23.40 | 23.60 | 23.10 | 22.90 | 23.40 |
| MCHC (g/dL) | 29.90 | 30.40 | 30.10 | 28.70 | 30.50 |
| RDW (%) | 14.60 | 14.20 | 13.90 | 15.10 | 16.50 |
| Platelet (×103 cells/uL) | 457.00 | 488.00 | 439.00 | 516.00 | 258.00 |
| PCT (%) | 0.54 | 0.38 | 0.33 | 0.60 | 0.21 |
| MPV (fL) | 11.90 | 7.80 | 7.50 | 11.60 | 8.00 |
| PDW | 15.00 | 14.10 | |||
| Seg. neut. (%) | 38.00 | 85.70 | 81.00 | 75.00 | 79.00 |
| Lymphocyte (%) | 54.00 | 6.20 | 3.00 | 9.00 | 2.00 |
| Monocyte (%) | 7.00 | 6.20 | 12.00 | 13.00 | 16.00 |
| Eosinophil (%) | 0.40 | 1.00 | |||
| Basophil (%) | 1.00 | 0.20 | |||
| LUC (%) | 1.40 | ||||
| ANC (uL) | 4005.00 | 8245.00 | 9299.00 | 8025.00 | 10,246.00 |
| PT-INR | 0.85 | 0.76 | 0.83 | 0.78 | 0.82 |
| PT (%) | 133.00 | 166.00 | 139.00 | 154.00 | 142.00 |
| PT (sec) | 9.10 | 8.10 | 8.90 | 8.40 | 8.80 |
| aPTT (sec) | 20.70 | 18.90 | 19.40 | 18.10 | 21.30 |
| Fibrinogen (mg/dL) | 114.00 | 241.00 | 354.00 | 270.00 | 314.00 |
| AT III (%) | 108.00 | 128.00 | 98.00 | 94.00 | 99.00 |
| Protein C (%) | 151.00 | 149.00 | 164.00 | 169.00 | 154.00 |
| Term | Category * | Genes | p Value | Fold Enrichment |
|---|---|---|---|---|
| ECM-receptor interaction | Up | IBSP, TNC, ITGA2, ITGA3, CHAD, HMMR, LAMA1, LAMC3, COMP, ITGA8, ITGB6, COL6A3, SV2B, TNN, COL1A1, SV2A, COL11A2, THBS1, SV2C, THBS2, COL11A1, THBS4, FN1, SPP1 | 1.386 × 10−8 | 3.9161 |
| Cytokine-cytokine receptor interaction | Both | TNFRSF6B, CSF2, IL1R2, IL22RA1, CCL2, TNFRSF12A, CCR1, TNFSF14, FASLG, CXCR3, CX3CL1, TNFSF18, IL11, CXCL10, LIF, TNFRSF11B, IL12RB1, CCL20, CXCR5, CLCF1, CXCR6, XCR1, IL1A, IL6, IL18RAP, FLT3, IL25, INHBB, CCL11, OSM, INHBA, PRLR, CXCL14, CCR3, CCR2, CX3CR1, NGFR, XCL1 | 5.430 × 10−5 | 1.9879 |
| Hematopoietic cell lineage | Down | CSF2, IL1R2, IL6, CD3G, CD3D, CD8A, FLT3, CD3E, ITGA2, ANPEP, ITGA3, IL11, CD38, DNTT, MS4A1, CD2, CD5, IL1A | 1.170 × 10−4 | 2.8687 |
| Neuroactive ligand-receptor interaction | Up | F2RL3, MCHR1, CCKAR, DRD1, GABRB3, GABRB2, CYSLTR2, DRD2, GABRB1, F2RL1, GRIK5, BDKRB1, HCRTR1, P2RY6, PTGIR, HRH3, NMUR1, CNR2, GABRD, PTGER3, GABRA1, GABRA4, GRIN1, GRIN2A, NTSR2, GRM4, P2RY10, CHRM4, ADRB1, PRLR, MLNR, ADRA1A, FSHB, HTR2C, HTR2A | 3.664 × 10−4 | 1.8739 |
| Focal adhesion | Up | IBSP, PGF, TNC, CHAD, COMP, ITGB6, COL6A3, TNN, THBS1, COL11A2, COL11A1, THBS2, THBS4, FN1, SPP1, MYLK3, ITGA2, IGF1, ITGA3, MAPK10, LAMA1, RASGRF1, LAMC3, ITGA8, COL1A1 | 1.030 × 10−2 | 1.7048 |
| Hypertrophic cardiomyopathy | Down | ACE, ACTC1, IL6, ATP2A2, ITGA8, CACNG6, ITGB6, CACNG5, CACNB2, IGF1, ITGA2, ITGA3 | 4.275 × 10−2 | 1.935 |
| Calcium signaling pathway | Down | SLC8A3, CCKAR, DRD1, PTGER3, ERBB4, CYSLTR2, MYLK3, GRIN1, CACNA1I, GRIN2A, BDKRB1, CD38, GNAL, ATP2B3, ADRB1, ATP2A2, ADRA1A, HTR2C, HTR2A, CACNA1B | 5.215 × 10−2 | 1.5575 |
| Cell cycle | Up | CDC6, PKMYT1, ESPL1, MCM2, MCM3, MCM4, MCM5, CCNB1, CDKN1A, CCNB3, CCNB2, PLK1, BUB1, BUB1B, MYC | 6.859 × 10−2 | 1.6447 |
| Dilated cardiomyopathy | Down | ACTC1, ADRB1, ATP2A2, ITGA8, CACNG6, ADCY6, ITGB6, CACNG5, CACNB2, IGF1, ITGA2, ITGA3 | 6.876 × 10−2 | 1.7878 |
| Bladder cancer | Both | RPS6KA5, CDKN1A, PGF, MMP9, CDH1, THBS1, MYC | 8.108 × 10−2 | 2.2844 |
| Progesterone-mediated oocyte maturation | Up | CCNB1, CCNB3, CCNB2, PLK1, ADCY6, BUB1, PKMYT1, IGF1, MAPK11, MAPK10, CPEB1 | 9.290 × 10−2 | 1.7531 |
| Cluster | Enrichment Score | Category * | Term | Count | p-Value | Fold Enrichment |
|---|---|---|---|---|---|---|
| 1 | 1.652 | CC | cell-cell adherens junction | 25 | 5.00 × 10−3 | 1.834 |
| MF | cadherin binding involved in cell-cell adhesion | 20 | 4.00 × 10−2 | 1.61 | ||
| BP | cell-cell adhesion | 19 | 5.00 × 10−2 | 1.622 | ||
| 2 | 1.522 | MF | voltage-gated potassium channel activity | 8 | 1.00 × 10−2 | 3.22 |
| BP | potassium ion transmembrane transport | 12 | 2.00 × 10−2 | 2.294 | ||
| MF | delayed rectifier potassium channel activity | 6 | 2.00 × 10−2 | 3.891 | ||
| CC | voltage-gated potassium channel complex | 9 | 3.00 × 10−2 | 2.424 | ||
| BP | regulation of ion transmembrane transport | 10 | 5.00 × 10−2 | 2.084 | ||
| BP | potassium ion transport | 7 | 1.00 × 10−1 | 1.974 | ||
| 3 | 1.413 | BP | membrane repolarization during cardiac muscle cell action potential | 4 | 8.00 × 10−3 | 9.252 |
| BP | regulation of membrane repolarization | 4 | 1.00 × 10−2 | 7.71 | ||
| MF | voltage-gated potassium channel activity involved in cardiac muscle cell action potential repolarization | 3 | 3.00 × 10−2 | 10.007 | ||
| BP | positive regulation of potassium ion transmembrane transport | 3 | 1.00 × 10−1 | 4.626 | ||
| BP | regulation of heart rate by cardiac conduction | 4 | 2.00 × 10−1 | 2.643 | ||
| 4 | 1.373 | CC | spectrin | 5 | 3.00 × 10−4 | 13.166 |
| BP | actin filament capping | 4 | 2.00 × 10−2 | 7.117 | ||
| CC | spectrin-associated cytoskeleton | 3 | 4.00 × 10−2 | 8.887 | ||
| MF | structural constituent of cytoskeleton | 9 | 1.00 × 10−1 | 1.91 | ||
| MF | phospholipid binding | 6 | 3.00 × 10−1 | 1.629 | ||
| BP | ER to Golgi vesicle-mediated transport | 6 | 8.00 × 10−1 | 0.867 | ||
| 5 | 1.333 | MF | glutathione peroxidase activity | 5 | 1.00 × 10−2 | 5.559 |
| BP | response to reactive oxygen species | 6 | 3.00 × 10−2 | 3.558 | ||
| BP | cellular oxidant detoxification | 5 | 4.00 × 10−1 | 1.652 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, M.Y.; Krishna Vasamsetti, B.M.; Kim, W.S.; Kang, H.J.; Kim, D.-Y.; Lim, B.; Cho, K.; Kim, J.S.; Chee, H.K.; Park, J.H.; et al. Comprehensive Analysis of Cardiac Xeno-Graft Unveils Rejection Mechanisms. Int. J. Mol. Sci. 2021, 22, 751. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020751
Park MY, Krishna Vasamsetti BM, Kim WS, Kang HJ, Kim D-Y, Lim B, Cho K, Kim JS, Chee HK, Park JH, et al. Comprehensive Analysis of Cardiac Xeno-Graft Unveils Rejection Mechanisms. International Journal of Molecular Sciences. 2021; 22(2):751. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020751
Chicago/Turabian StylePark, Min Young, Bala Murali Krishna Vasamsetti, Wan Seop Kim, Hee Jung Kang, Do-Young Kim, Byeonghwi Lim, Kahee Cho, Jun Seok Kim, Hyun Keun Chee, Jung Hwan Park, and et al. 2021. "Comprehensive Analysis of Cardiac Xeno-Graft Unveils Rejection Mechanisms" International Journal of Molecular Sciences 22, no. 2: 751. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020751