Effect of Polyhexamethylene Biguanide in Combination with Undecylenamidopropyl Betaine or PslG on Biofilm Clearance


Abstract
:1. Introduction
2. Results
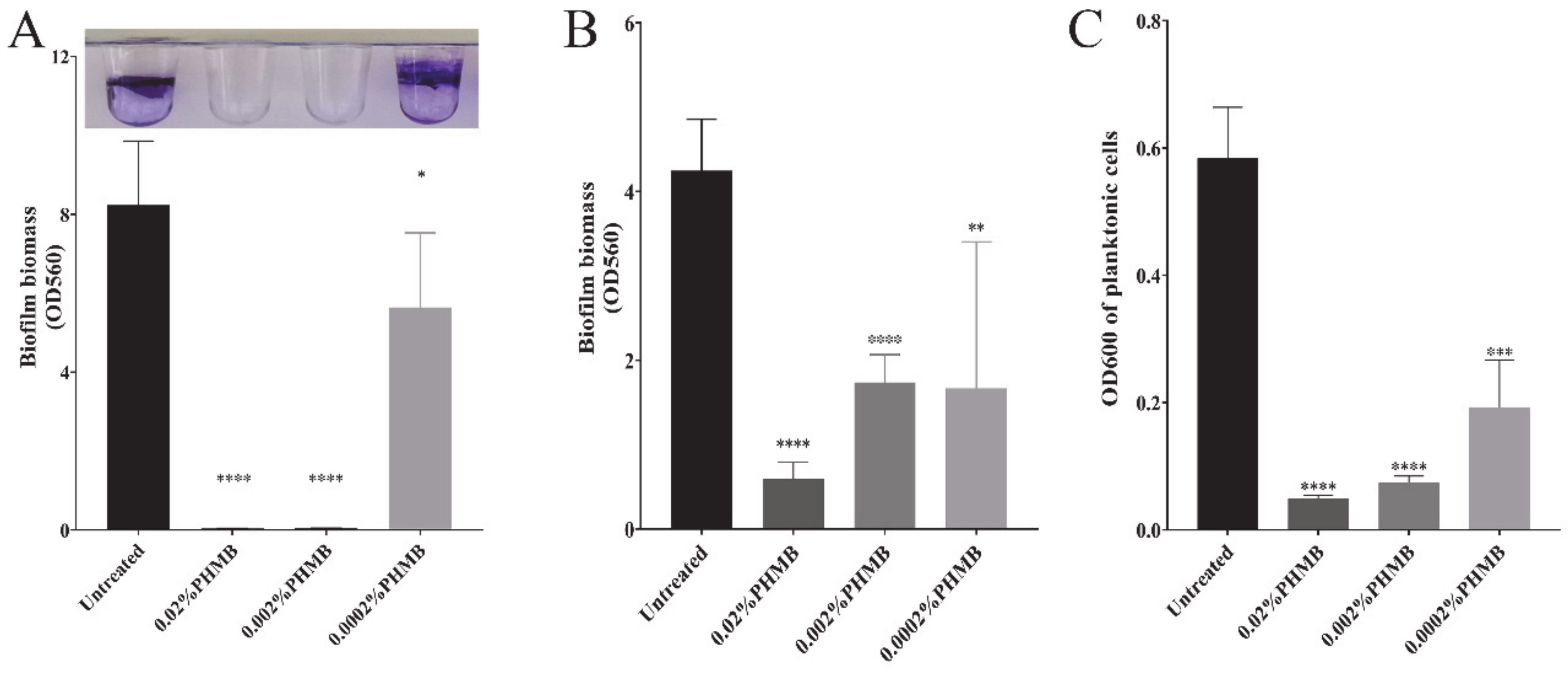
2.1. The Concentration of PHMB that Can Prevent Biofilm Formation and Disrupt the Preformed Biofilm of P. aeruginosa
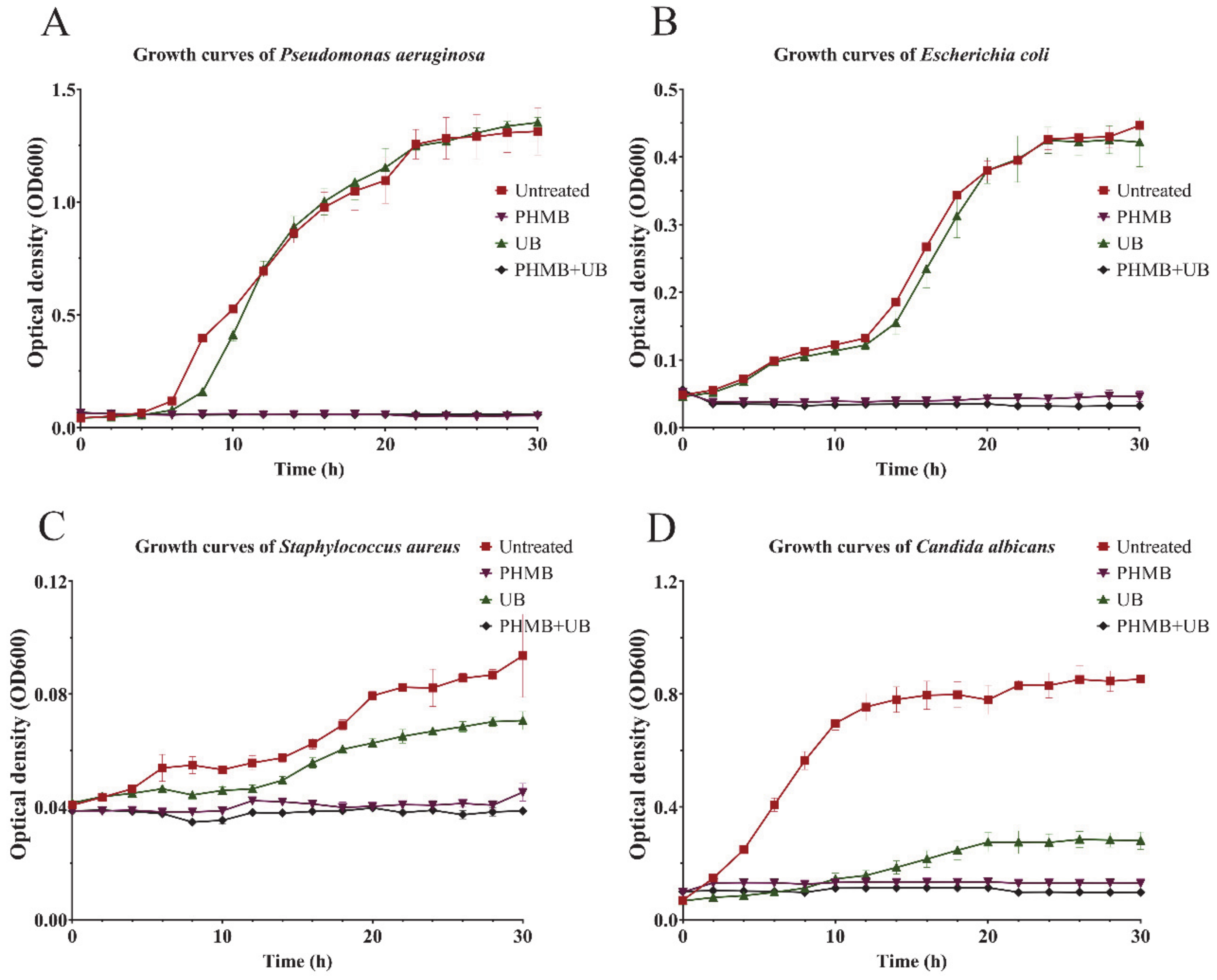
2.2. Effect of PHMB, UB, and PHMB + UB on Different Stages of P. aeruginosa Biofilm Development
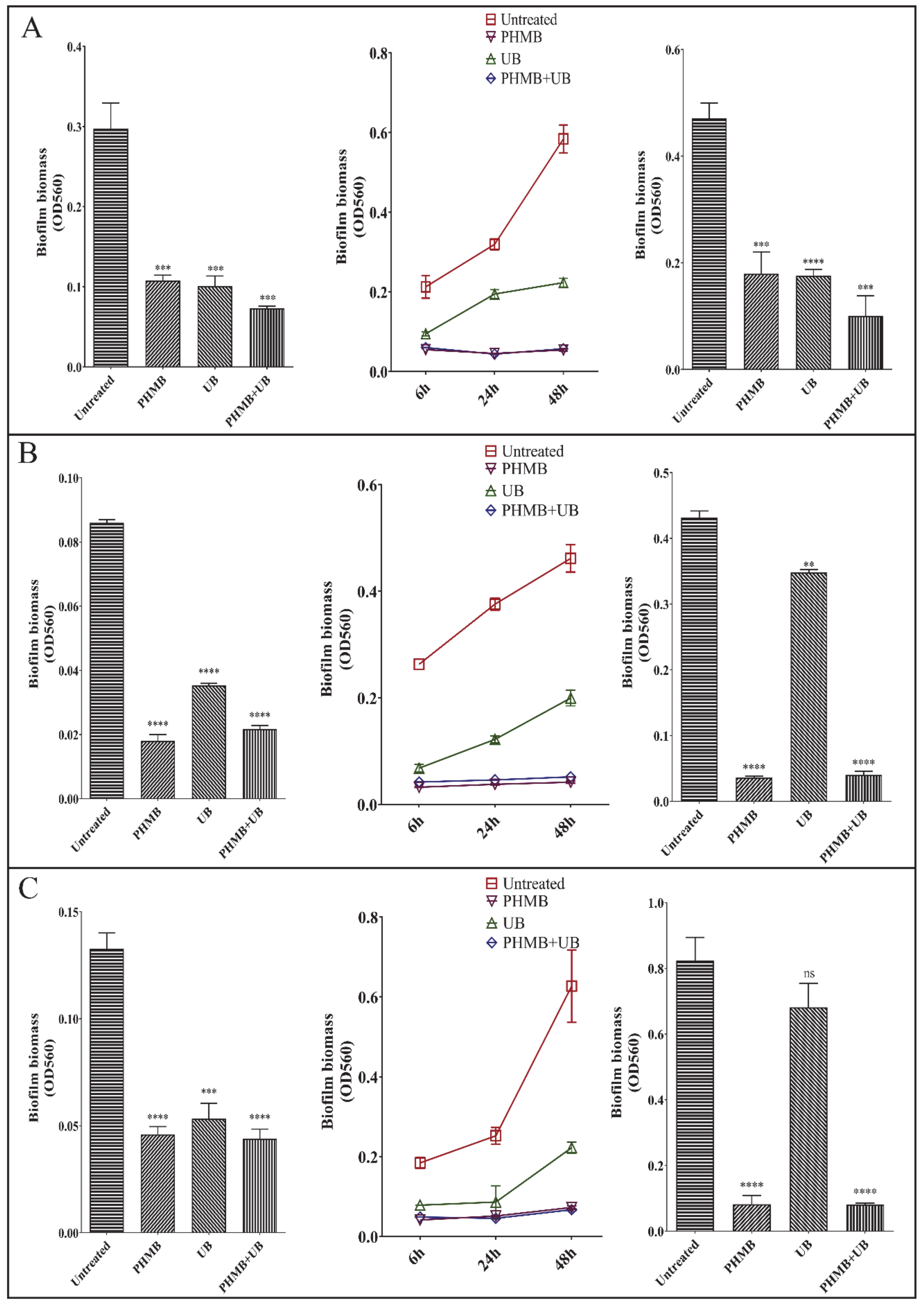
2.3. PHMB and UB against Biofilms Formed by Different Types of Pathogens
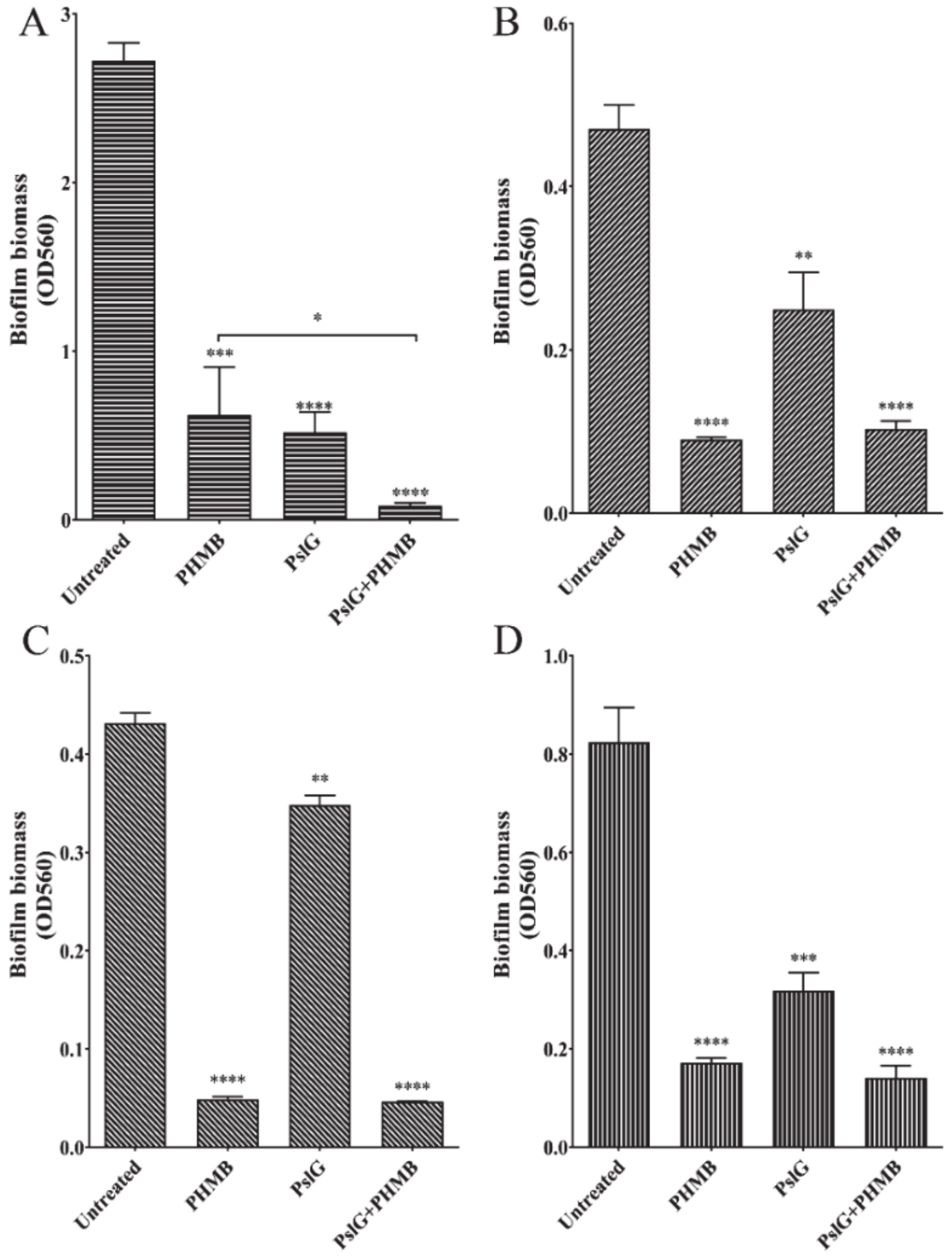
2.4. Combination of PHMB and PslG Enhanced Biofilms Clearance
3. Discussion
4. Materials and Methods
4.1. Strains, Media, and Growth Conditions
4.2. Growth Curves
4.3. Initial Attachment Assay
4.4. Biofilm Formation Assay
4.5. Disruption of Preformed Biofilms
4.6. Image Acquisition and Analysis of Air-Liquid Interface Biofilms (Pellicles)
4.7. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, F.; Li, H.; Gu, L.; Liu, M.; Xue, C.X.; Cao, B.; Wang, C.; The National Influenza A(H1N1)pdm09 Clinical Investigation Group of China. Risk factors for nosocomial infection among hospitalised severe influenza A(H1N1)pdm09 patients. Respir. Med. 2018, 134, 86–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donlan, R.M. Biofilms and device-associated infections. Emerg. Infect. Dis. 2001, 7, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Parsek, M.R.; Singh, P.K. Bacterial biofilms: An emerging link to disease pathogenesis. Annu. Rev. Microbiol. 2003, 57, 677–701. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Tolker-Nielsen, T. Biofilm Development. Microbiol. Spectr. 2015, 3, MB-0001-2014. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Conover, M.; Lu, H.; Parsek, M.R.; Bayles, K.; Wozniak, D.J. Assembly and development of the Pseudomonas aeruginosa biofilm matrix. PLoS Pathog. 2009, 5, e1000354. [Google Scholar] [CrossRef] [Green Version]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as complex differentiated communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Whitchurch, C.B.; Tolker-Nielsen, T.; Ragas, P.C.; Mattick, J.S. Extracellular DNA required for bacterial biofilm formation. Science 2002, 295, 1487. [Google Scholar] [CrossRef]
- Parks, Q.M.; Young, R.L.; Poch, K.R.; Malcolm, K.C.; Vasil, M.L.; Nick, J.A. Neutrophil enhancement of Pseudomonas aeruginosa biofilm development: Human F-actin and DNA as targets for therapy. J. Med. Microbiol. 2009, 58, 492–502. [Google Scholar] [CrossRef]
- Shukla, S.K.; Rao, T.S. Staphylococcus aureus biofilm removal by targeting biofilm-associated extracellular proteins. Indian J. Med. Res. 2017, 146, S1–S8. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Su, T.; Wu, H.; Liu, S.; Wang, D.; Zhao, T.; Jin, Z.; Du, W.; Zhu, M.-J.; Chua, S.L.; et al. PslG, a self-produced glycosyl hydrolase, triggers biofilm disassembly by disrupting exopolysaccharide matrix. Cell Res. 2015, 25, 1352–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Wang, D.; Tang, M.; Ma, L.Z. The advance of assembly of exopolysaccharide Psl biosynthesis machinery in Pseudomonas aeruginosa. Microbiologyopen 2019, 8, e857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, G.; Kramer, A. Biocompatibility index of antiseptic agents by parallel assessment of antimicrobial activity and cellular cytotoxicity. J. Antimicrob. Chemother. 2008, 61, 1281–1287. [Google Scholar] [CrossRef] [Green Version]
- Oule, M.K.; Azinwi, R.; Bernier, A.-M.; Kablan, T.; Maupertuis, A.-M.; Mauler, S.; Nevry, R.K.; Dembele, K.; Forbes, L.; Diop, L. Polyhexamethylene guanidine hydrochloride-based disinfectant: A novel tool to fight meticillin-resistant Staphylococcus aureus and nosocomial infections. J. Med. Microbiol. 2008, 57, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, P.; Moore, L.E. Cationic antiseptics: Diversity of action under a common epithet. J. Appl. Microbiol. 2005, 99, 703–715. [Google Scholar] [CrossRef]
- Albert, M.; Feiertag, P.; Hayn, G.; Saf, R.; Honig, H. Structure—Activity relationships of oligoguanidines-influence of counterion, diamine, and average molecular weight on blocidal activities. Biomacromolecules 2003, 4, 1811–1817. [Google Scholar] [CrossRef]
- Wiegand, C.; Eberlein, T.; Andriessen, A. Antibacterial activity of polihexanide formulations in a co-culture of HaCaT keratinocytes and Staphylococcus aureus and at different pH levels. Wound Repair Regen. 2017, 25, 423–431. [Google Scholar] [CrossRef]
- Medvedec Mikic, I.; Cigic, L.; Kero, D.; Kalibovic Govorko, D.; Prpic Mehicic, G.; Tambic Andrasevic, A.; Simeon, P. Antimicrobial effectiveness of polyhexamethylene biguanide on Enterococcus faecalis, Staphylococcus epidermidis and Candida albicans. Med. Glas. 2018, 15, 132–138. [Google Scholar] [CrossRef]
- Hubner, N.O.; Kramer, A. Review on the Efficacy, Safety and Clinical Applications of Polihexanide, a Modern Wound Antiseptic. Ski. Pharmacol. Physiol. 2010, 23, 17–27. [Google Scholar] [CrossRef]
- Ikeda, T.; Tazuke, S.; Watanabe, M. Interaction of Biologically-Active Molecules with Phospholipid-Membranes.1. Fluorescence Depolarization Studies on the Effect of Polymeric Biocide Bearing Biguanide Groups in the Main Chain. Biochim. Biophys. Acta 1983, 735, 380–386. [Google Scholar] [CrossRef]
- López-Rojas, R.; Fernández-Cuenca, F.; Serrano-Rocha, L.; Pascual, Á. In vitro activity of a polyhexanide–betaine solution against high-risk clones of multidrug-resistant nosocomial pathogens. Enferm. Infect. Microbiol. Clin. 2017, 35, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Koburger, T.; Hubner, N.O.; Braun, M.; Siebert, J.; Kramer, A. Standardized comparison of antiseptic efficacy of triclosan, PVP-iodine, octenidine dihydrochloride, polyhexanide and chlorhexidine digluconate. J. Antimicrob. Chemother. 2010, 65, 1712–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romanelli, M.; Dini, V.; Barbanera, S.; Bertone, M.S. Evaluation of the efficacy and tolerability of a solution containing propyl betaine and polihexanide for wound irrigation. Ski. Pharm. Physiol. 2010, 23, 41–44. [Google Scholar] [CrossRef]
- Minnich, K.E.; Stolarick, R.; Wilkins, R.G.; Chilson, G.; Pritt, S.L.; Unverdorben, M. The effect of a wound care solution containing polyhexanide and betaine on bacterial counts: Results of an in vitro study. Ostomy Wound Manag. 2012, 58, 32–36. [Google Scholar]
- Machuca, J.; Lopez-Rojas, R.; Fernandez-Cuenca, F.; Pascual, A. Comparative activity of a polyhexanide-betaine solution against biofilms produced by multidrug-resistant bacteria belonging to high-risk clones. J. Hosp. Infect. 2019, 103, e92–e96. [Google Scholar] [CrossRef]
- Cazzaniga, A.; Serralta, V.; Davis, S.; Orr, R.; Eaglstein, W.; Mertz, P.M. The effect of an antimicrobial gauze dressing impregnated with 0.2-percent polyhexamethylene biguanide as a barrier to prevent Pseudomonas aeruginosa wound invasion. Wounds Compend. Clin. Res. Pract. 2002, 14, 169–176. [Google Scholar]
- Welk, A.; Splieth, C.H.; Schmidt-Martens, G.; Schwahn, C.; Kocher, T.; Kramer, A.; Rosin, M. The effect of a polyhexamethylene biguanide mouthrinse compared with a triclosan rinse and a chlorhexidine rinse on bacterial counts and 4-day plaque re-growth. J. Clin. Periodontol. 2005, 32, 499–505. [Google Scholar] [CrossRef]
- Davis, S.C.; Harding, A.; Gil, J.; Parajon, F.; Valdes, J.; Solis, M.; Higa, A. Effectiveness of a polyhexanide irrigation solution on methicillin-resistant Staphylococcus aureus biofilms in a porcine wound model. Int. Wound J. 2017, 14, 937–944. [Google Scholar] [CrossRef]
- Woodford, N.; Turton, J.F.; Livermore, D.M. Multiresistant Gram-negative bacteria: The role of high-risk clones in the dissemination of antibiotic resistance. FEMS Microbiol. Rev. 2011, 35, 736–755. [Google Scholar] [CrossRef] [Green Version]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the ESKAPE pathogens. Expert Rev. Anti-Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odds, F.C.; Brown, A.J.; Gow, N.A. Candida albicans genome sequence: A platform for genomics in the absence of genetics. Genome Biol. 2004, 5, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horrocks, A. Prontosan wound irrigation and gel: Management of chronic wounds. Br. J. Nurs. 2006, 15, 1224–1228. [Google Scholar] [CrossRef] [PubMed]
- Mulder, G.D.; Cavorsi, J.P.; Lee, D.K. Polyhexamethylene biguanide (PHMB): An addendum to current topical antimicrobials. Wounds Compend. Clin. Res. Pract. 2007, 19, 173–182. [Google Scholar]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.L.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef]
- Ma, L.; Jackson, K.D.; Landry, R.M.; Parsek, M.R.; Wozniak, D.J. Analysis of Pseudomonas aeruginosa conditional psl variants reveals roles for the psl polysaccharide in adhesion and maintaining biofilm structure postattachment. J. Bacteriol. 2006, 188, 8213–8221. [Google Scholar] [CrossRef] [Green Version]
- Jensen, S.E.; Fecycz, I.T.; Campbell, J.N. Nutritional factors controlling exocellular protease production by Pseudomonas-aeruginosa. J. Bacteriol. 1980, 144, 844–847. [Google Scholar] [CrossRef] [Green Version]
- Yanisch-Perron, C.; Vieira, J.; Messing, J. Improved M13 phage cloning vectors and host strains: Nucleotide sequences of the M13mp18 and pUC19 vectors. Gene 1985, 33, 103–119. [Google Scholar] [CrossRef]
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Schembri, M.A.; Klemm, P. Biofilm formation in a hydrodynamic environment by novel FimH variants and ramifications for virulence. Infect. Immun. 2001, 69, 1322–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, G.D.; Simpson, W.A.; Younger, J.J.; Baddour, L.M.; Barrett, F.F.; Melton, D.M.; Beachey, E.H. Adherence of coagulase-negative staphylococci to plastic tissue culture plates: A quantitative model for the adherence of staphylococci to medical devices. J. Clin. Microbiol. 1985, 22, 996–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonzi, W.A.; Irwin, M.Y. Isogenic strain construction and gene mapping in Candida albicans. Genetics 1993, 134, 717–728. [Google Scholar] [PubMed]
- Alvarez-Peral, F.J.; Arguelles, J.C. Changes in external trehalase activity during human serum-induced dimorphic transition in Candida albicans. Res. Microbiol. 2000, 151, 837–843. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Parsek, M.R.; Wozniak, D.J.; Ma, L.Z. A spider web strategy of type IV pili-mediated migration to build a fibre-like Psl polysaccharide matrix in Pseudomonas aeruginosa biofilms. Environ. Microbiol. 2013, 15, 2238–2253. [Google Scholar] [CrossRef] [Green Version]
- Heydorn, A.; Nielsen, A.T.; Hentzer, M.; Sternberg, C.; Givskov, M.; Ersboll, B.K.; Molin, S. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology 2000, 146, 2395–2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Rich Medium | Minimal Medium * |
|---|---|---|
| Pseudomonas aeruginosa PAO1 [37] | LBNS medium [38] | Jensen’s medium [39] |
| Escherichia coli JM109 [40] | LB medium [41] | M9 medium [42] |
| Staphylococcus aureus ATCC6538 | TSB medium [43] | Jensen’s medium |
| Candida albicans SC5314 [44] | YPD medium [45] | RPMI 1640 medium (Gibco, Life technologies, New York, NY, USA) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Wang, D.; Ma, L.Z. Effect of Polyhexamethylene Biguanide in Combination with Undecylenamidopropyl Betaine or PslG on Biofilm Clearance. Int. J. Mol. Sci. 2021, 22, 768. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020768
Zheng Y, Wang D, Ma LZ. Effect of Polyhexamethylene Biguanide in Combination with Undecylenamidopropyl Betaine or PslG on Biofilm Clearance. International Journal of Molecular Sciences. 2021; 22(2):768. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020768
Chicago/Turabian StyleZheng, Yaqian, Di Wang, and Luyan Z. Ma. 2021. "Effect of Polyhexamethylene Biguanide in Combination with Undecylenamidopropyl Betaine or PslG on Biofilm Clearance" International Journal of Molecular Sciences 22, no. 2: 768. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020768