Elucidating the Functional Roles of Helper and Cytotoxic T Cells in the Cell-Mediated Immune Responses of Olive Flounder (Paralichthys olivaceus)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Husbandry
2.2. Synthesis of CD8α- and CD8β-Peptides
2.3. Production of mAbs (3H9-CD8α, 1G7-CD8β) Specific to CD8 Lymphocytes from Olive Flounder
2.4. Construction of Plasmids
2.5. Transfection
2.6. Western Blotting
2.7. Preparation of Leukocytes from Olive Flounder
2.8. Flow Cytometry
2.9. Immunofluorescence Staining
2.10. RT-PCR with Flow Cytometry Sorted Leukocytes
2.11. Assessing Populations of CD4-1-, CD4-2-, CD8α- and CD8β-Positive Lymphocytes in Olive Flounder
2.12. Quantitative Real-Time PCR (RT-qPCR) Analysis
2.13. Statistical Analysis
3. Results
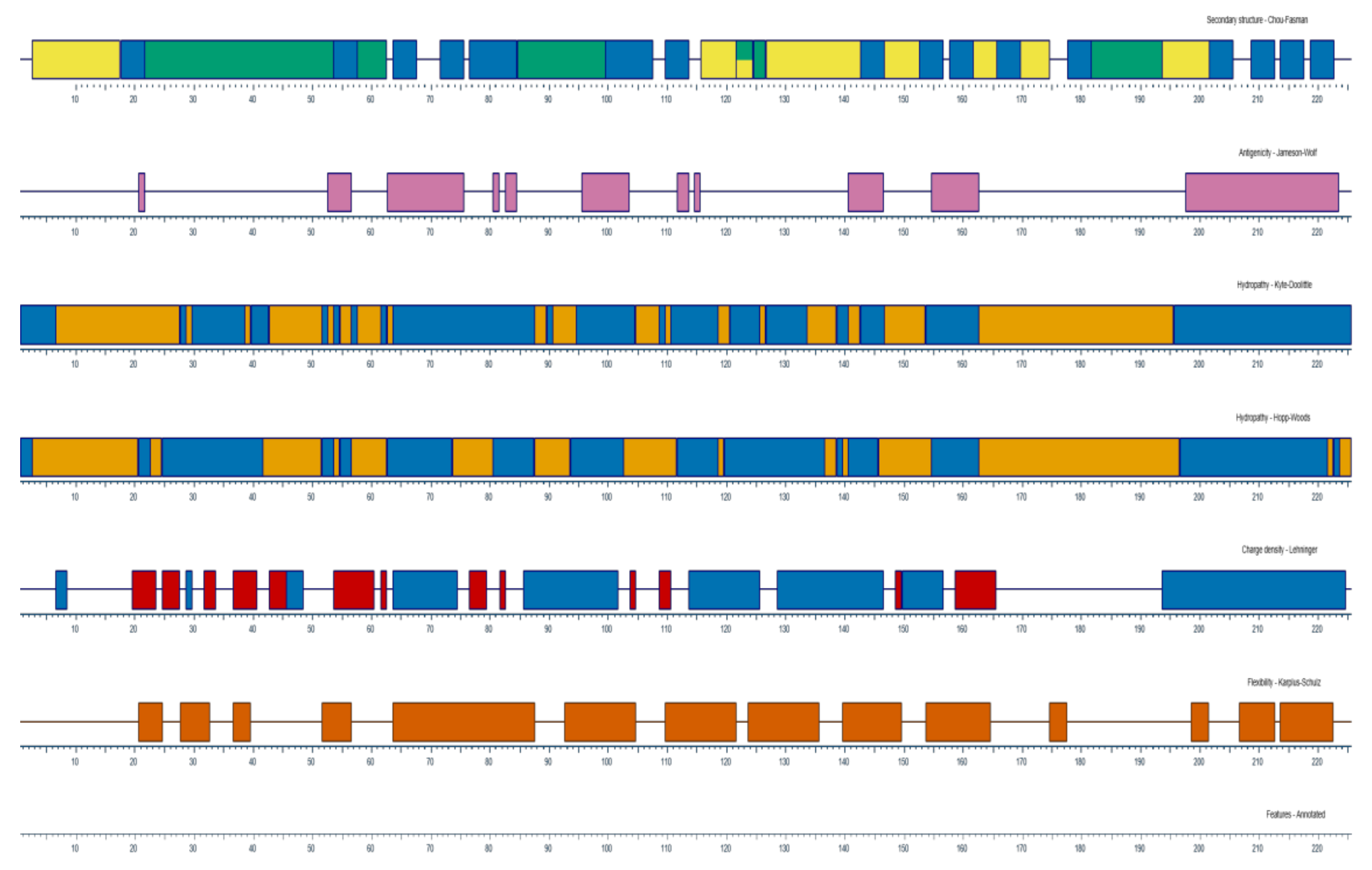
3.1. Synthesis and Selection of CD8α and CD8β Peptides
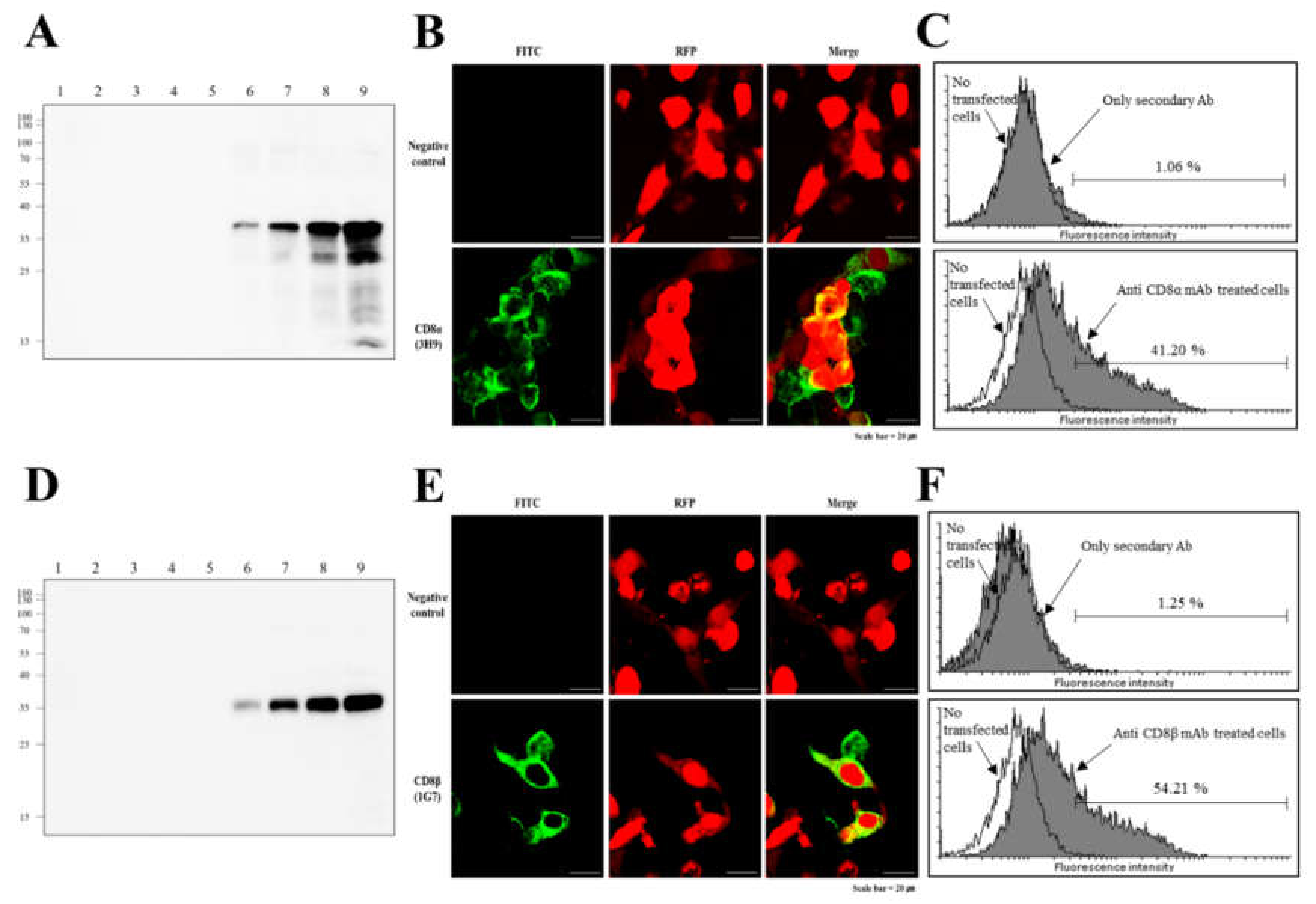
3.2. Reactivity of Anti-CD8α and Anti-CD8β mAbs
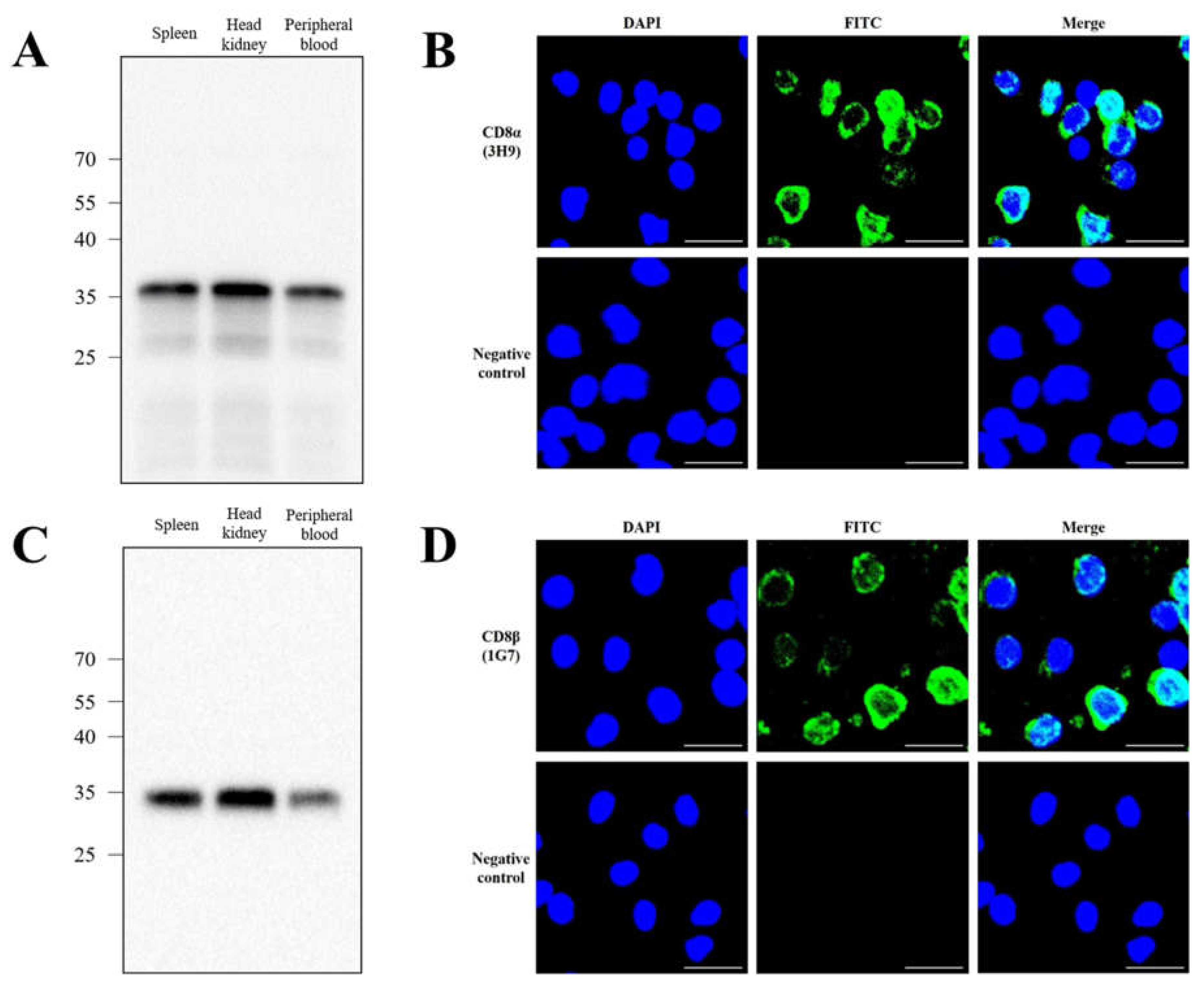
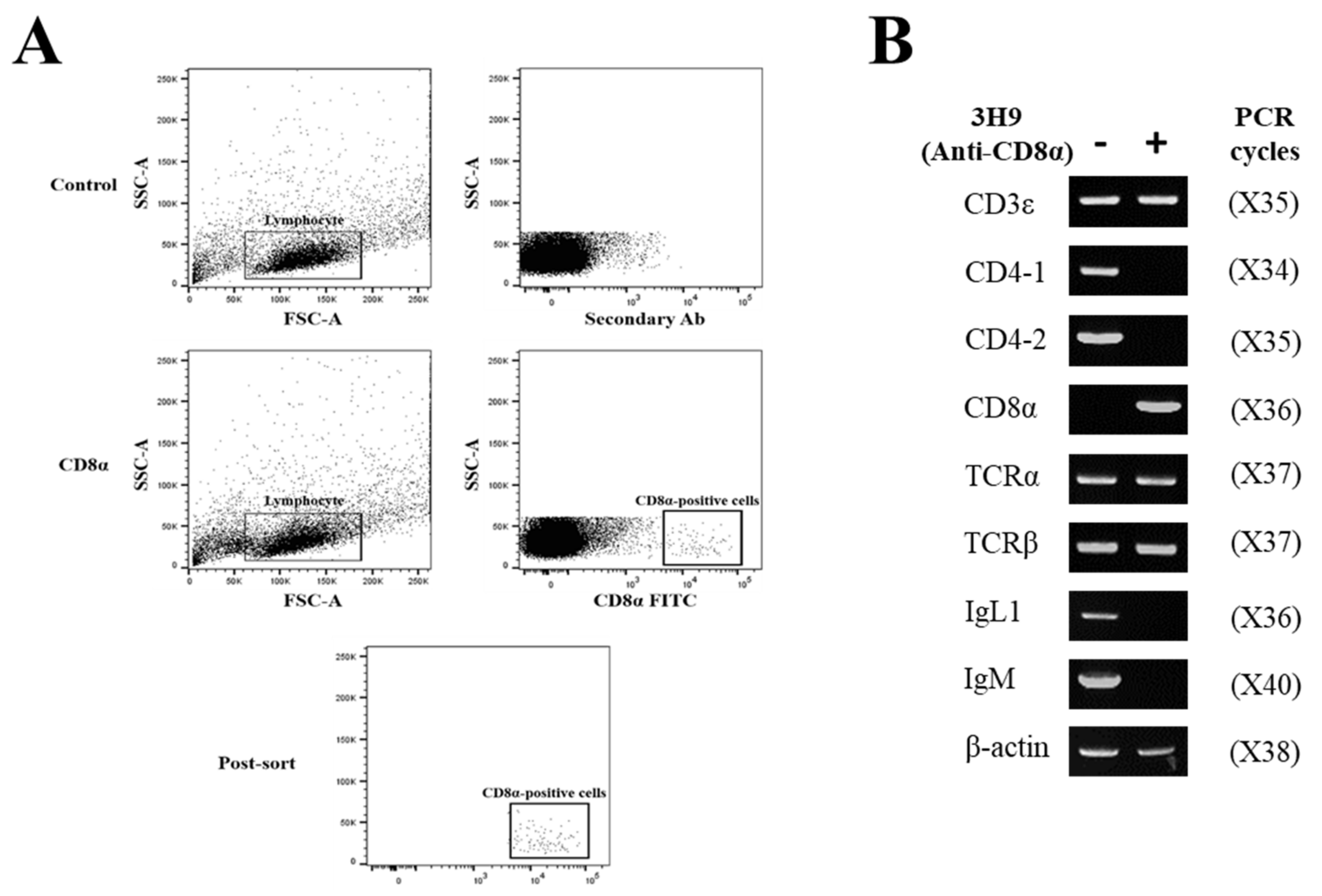
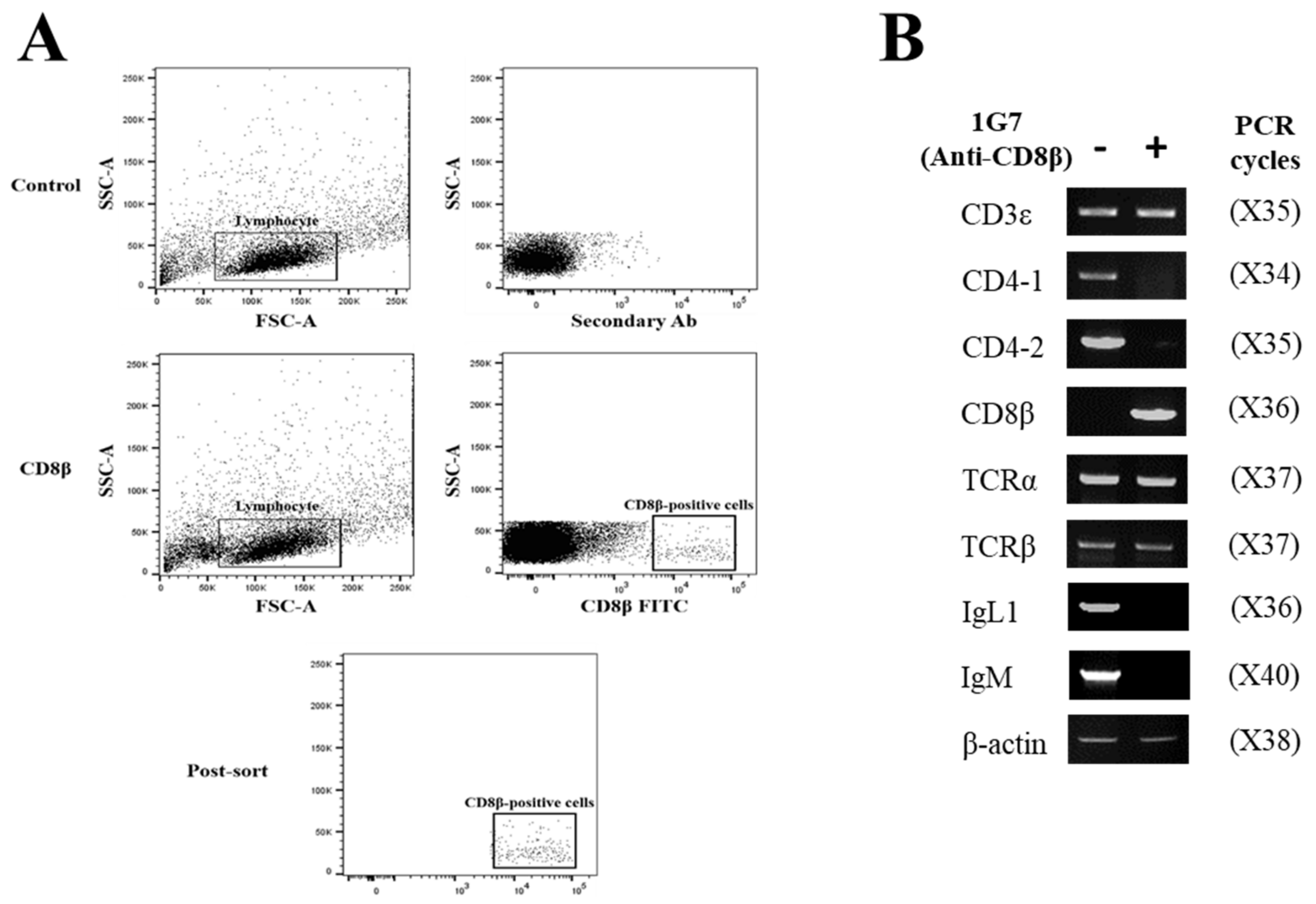
3.3. Specificity of Anti-CD8α and Anti-CD8β mAbs
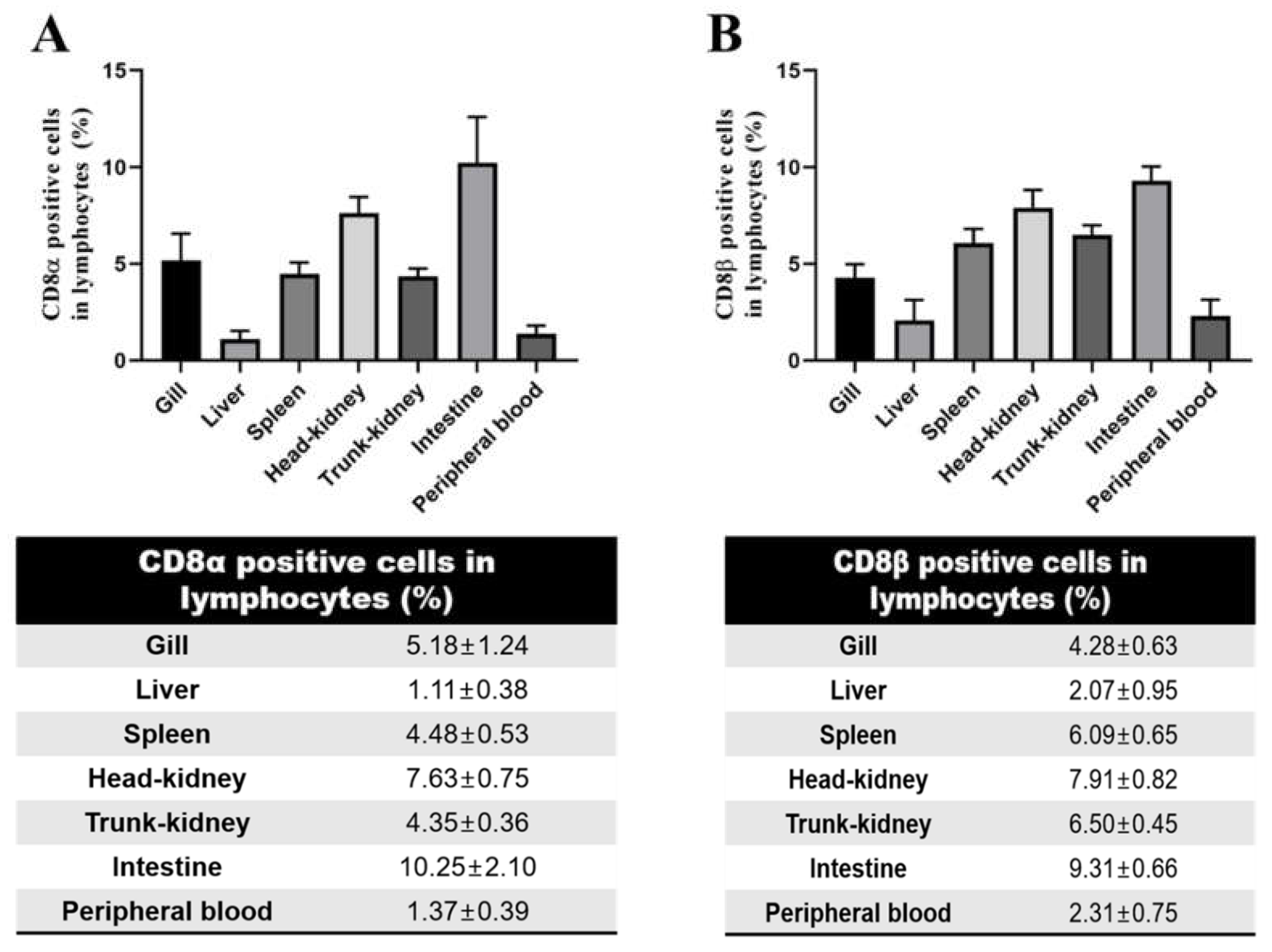
3.4. Distribution of CD8α- and CD8β-Positive Lymphocytes in Tissues by Flow Cytometry
3.5. Immune Gene Profiles of CD8α- and CD8β-Positive Lymphocytes
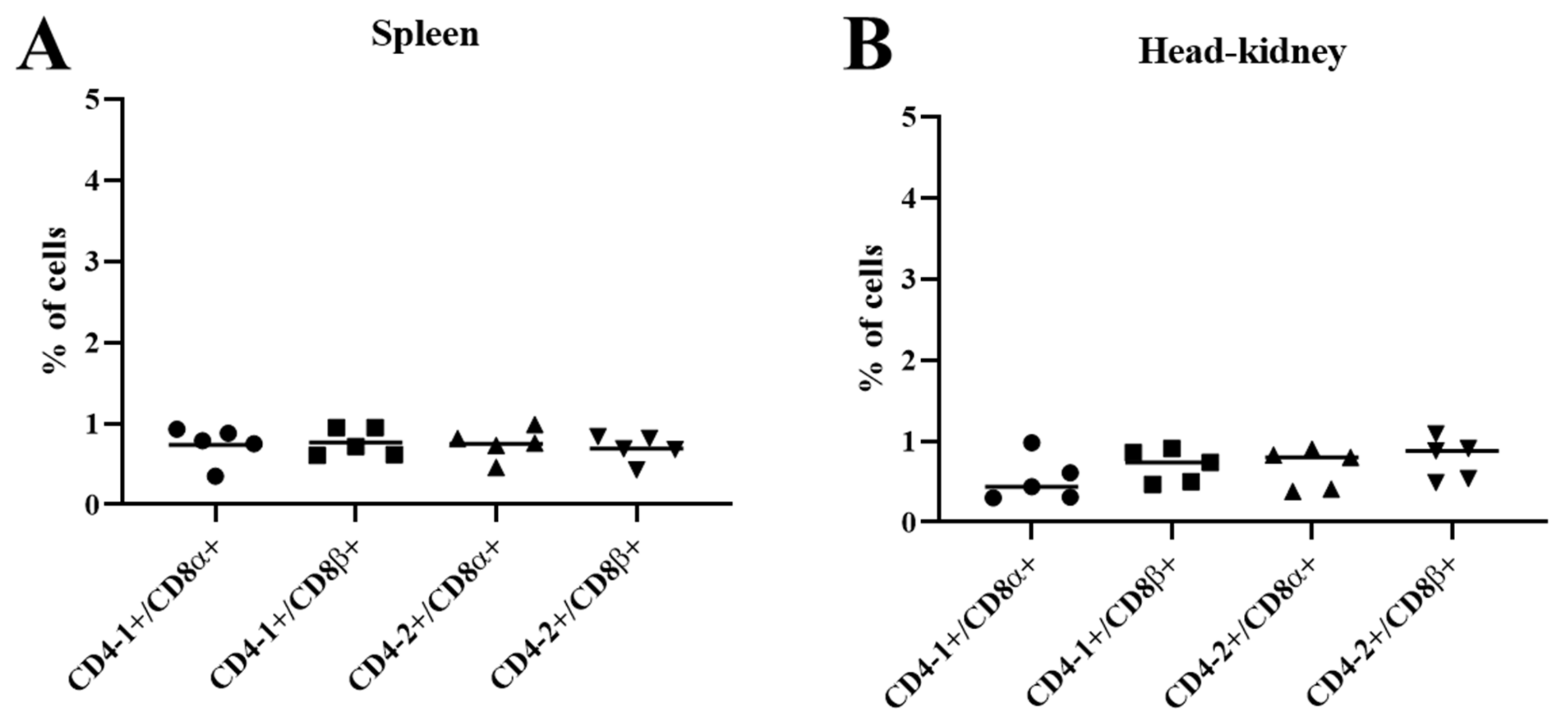
3.6. Two-Color Flow Cytometry
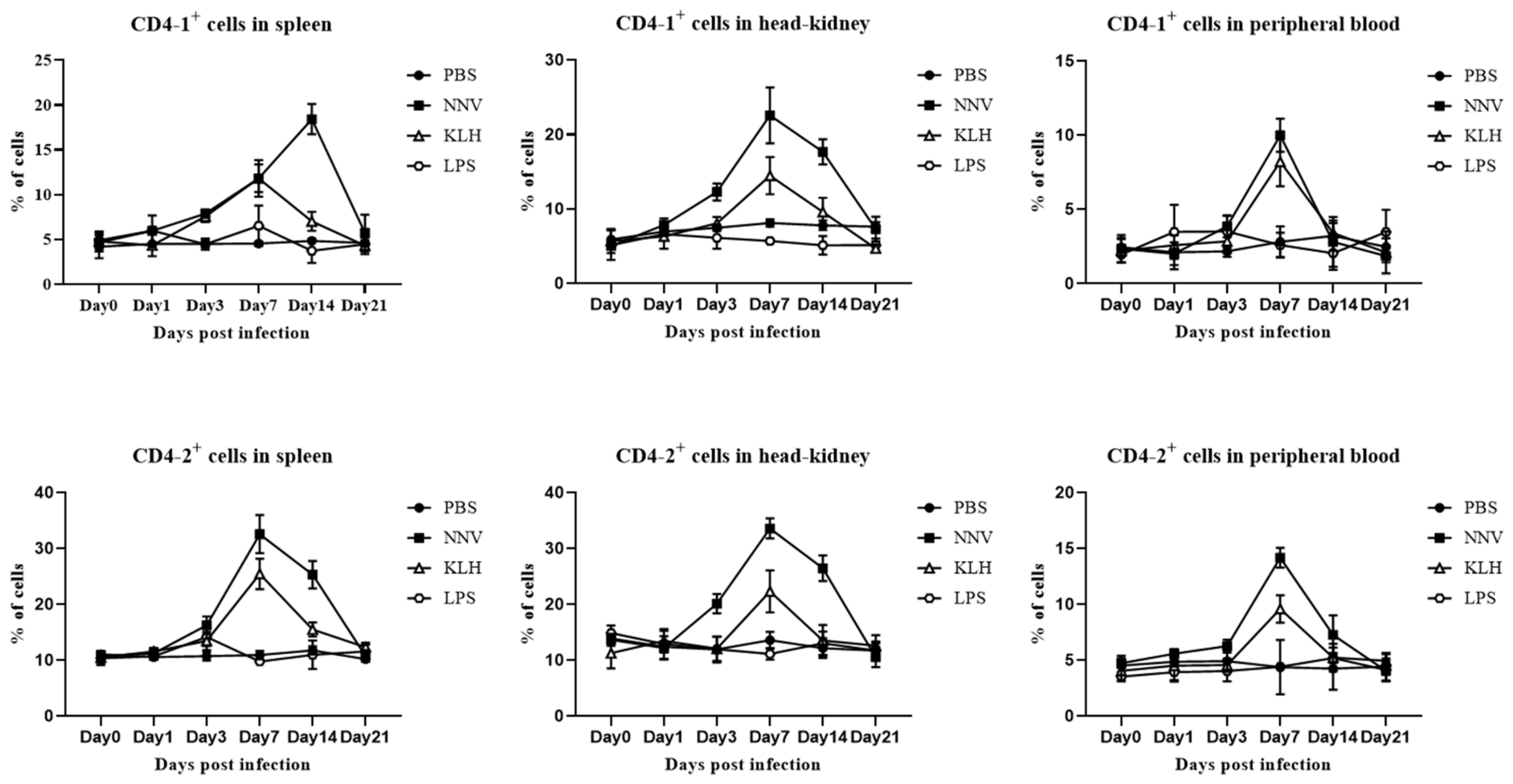
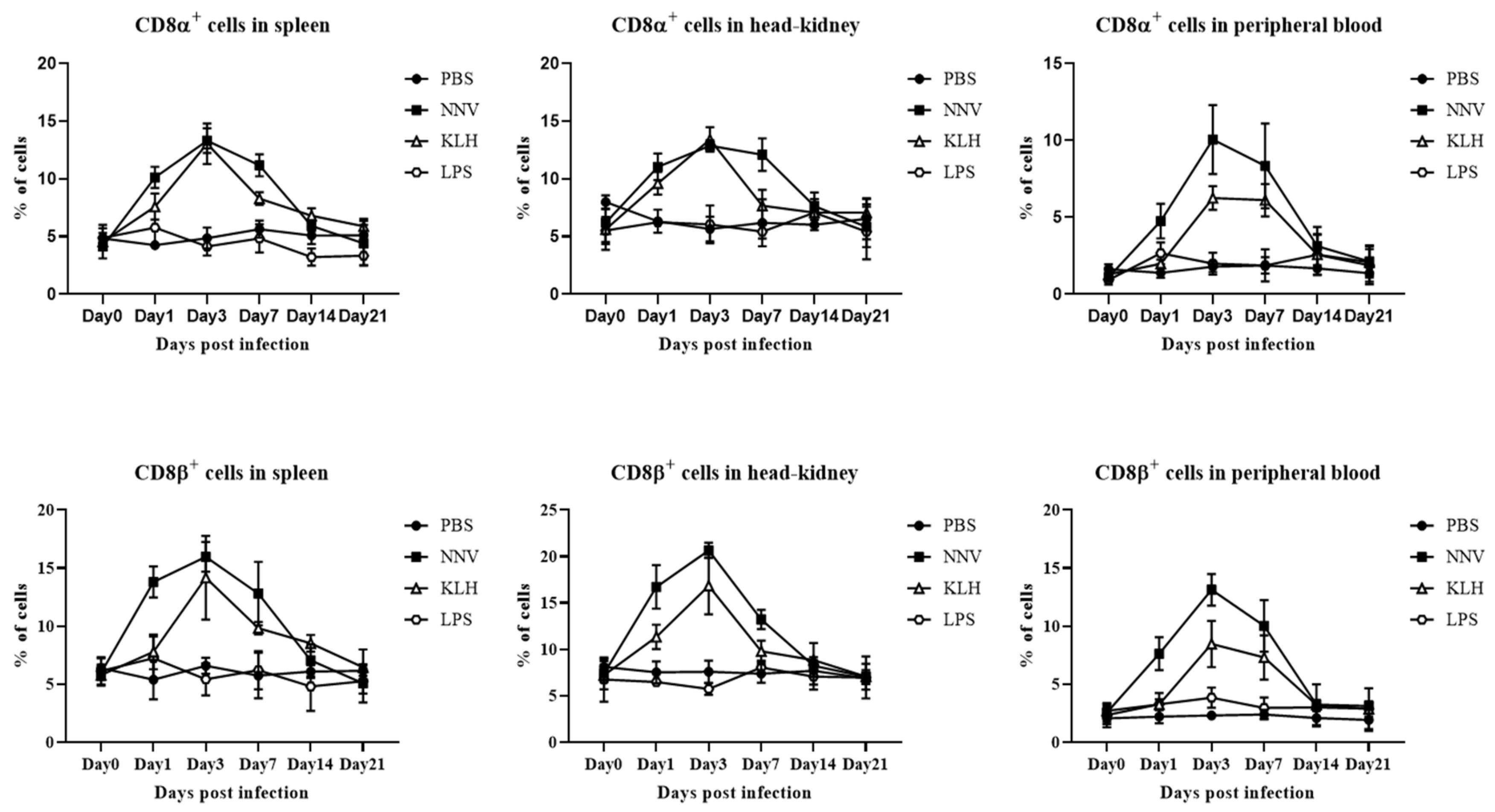
3.7. In Vivo Proliferation of CD4- and CD8-Positive Lymphocytes after Viral Infection with NNV or Administration of KLH or LPS
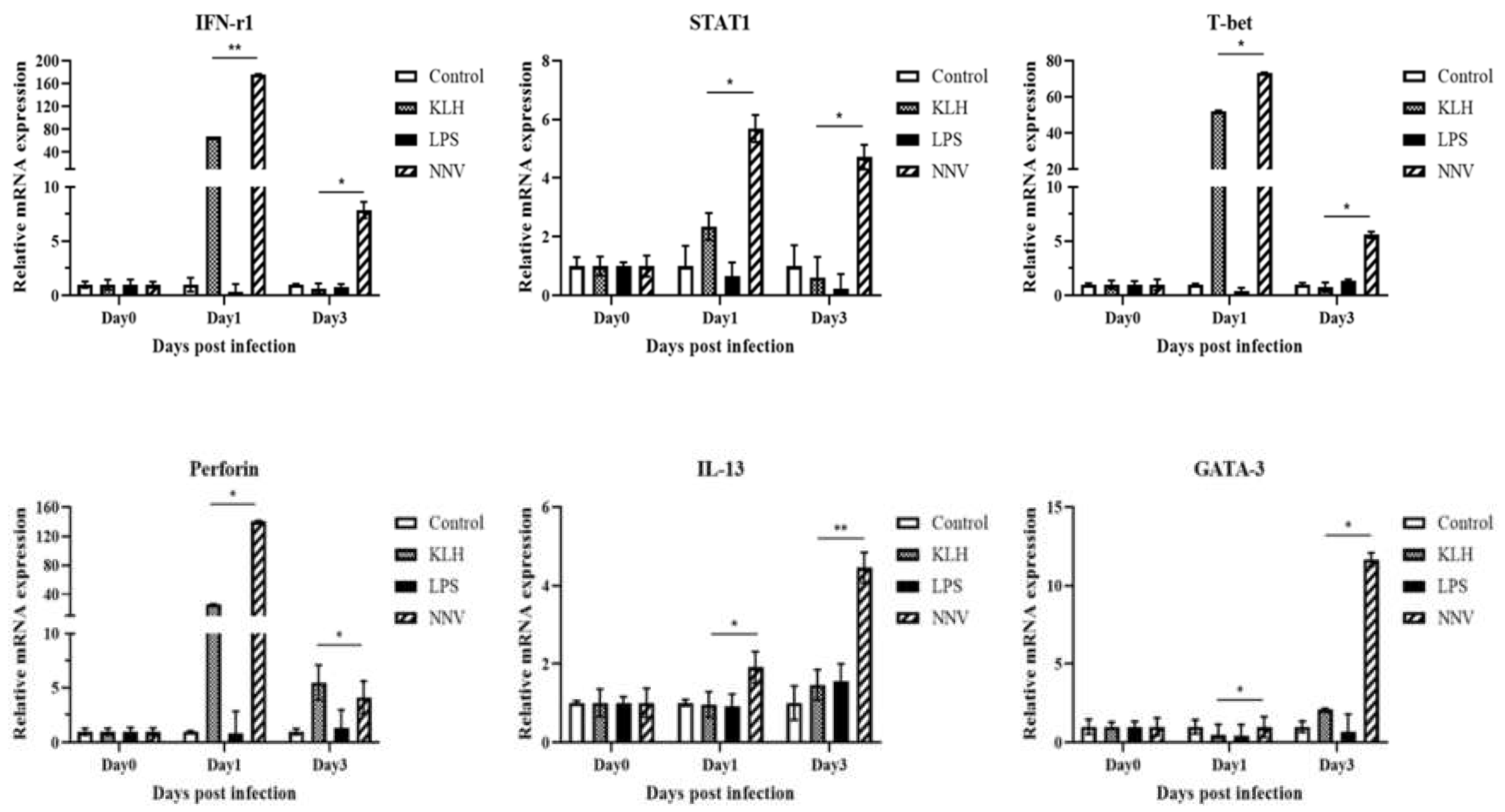
3.8. Expression of Transcription Factors and Cytokines by Lymphocytes after Virus Infection or Immuno-Stimulation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jung, J.W.; Chun, J.H.; Lee, J.S.; Kim, S.W.; Lee, A.R.; Kim, J.; Lazarte, J.M.S.; Kim, Y.R.; Kim, H.J.; Thompson, K.D.; et al. Characterization of CD4-Positive Lymphocytes in the Antiviral Response of Olive Flounder (Paralichthys oliveceus) to Nervous Necrosis Virus. Int. J. Mol. Sci. 2020, 21, 4180. [Google Scholar] [CrossRef] [PubMed]
- Teruyuki, N.; Shibasaki, Y.; Matsuura, Y. T cells in fish. Biology 2015, 4, 640–663. [Google Scholar]
- Kato, G.; Goto, K.; Akune, I.; Aoka, S.; Kondo, H.; Hirono, I. CD4 and CD8 homologues in Japanese flounder, Paralichthys olivaceus: Differences in the expressions and localizations of CD4-1, CD4-2, CD8α and CD8β. Dev. Comp. Immunol. 2013, 39, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.W.; Lee, J.S.; Kim, J.; Im, S.P.; Kim, S.W.; Lazarte, J.M.S.; Kim, Y.R.; Chun, J.H.; Ha, M.W.; Na Kim, N.; et al. Involvement of CD4-1 T cells in the cellular immune response of olive flounder (Paralichthys olivaceus) against viral hemorrhagic septicemia virus (VHSV) and nervous necrosis virus (NNV) infection. Dev. Comp. Immunol. 2019, 103, 103518. [Google Scholar] [CrossRef] [PubMed]
- Takizawa, F.; Dijkstra, J.M.; Kotterba, P.; Korytář, T.; Kock, H.; Köllner, B.; Jaureguiberry, B.; Nakanishi, T.; Fischer, U. The expression of CD8α discriminates distinct T cell subsets in teleost fish. Dev. Comp. Immunol. 2011, 35, 752–763. [Google Scholar] [CrossRef]
- Patel, S.; Øvergård, A.-C.; Nerland, A.H. CD8α and CD8β in Atlantic halibut, Hippoglossus hippoglossus: Cloning, characterization and gene expression during viral and bacterial infection. Fish Shellfish Immunol. 2008, 25, 570–580. [Google Scholar] [CrossRef]
- Maisey, K.; Montero, R.; Corripio-Miyar, Y.; Toro-Ascuy, D.; Valenzuela, B.; Reyes-Cerpa, S.; Sandino, A.M.; Zou, J.; Wang, T.; Secombes, C.J.; et al. Isolation and Characterization of Salmonid CD4+T Cells. J. Immunol. 2016, 196, 4150–4163. [Google Scholar] [CrossRef] [Green Version]
- Laing, K.J.; Zou, J.J.; Purcell, M.K.; Phillips, R.; Secombes, C.J.; Hansen, J.D. Evolution of the CD4 Family: Teleost Fish Possess Two Divergent Forms of CD4 in Addition to Lymphocyte Activation Gene-3. J. Immunol. 2006, 177, 3939–3951. [Google Scholar] [CrossRef] [Green Version]
- Moore, L.J.; Somamoto, T.; Lie, K.K.; Dijkstra, J.M.; Hordvik, I. Characterisation of salmon and trout CD8α and CD8β. Mol. Immunol. 2005, 42, 1225–1234. [Google Scholar] [CrossRef]
- Taniuchi, I. CD4 helper and CD8 cytotoxic T cell differentiation. Annu. Rev. Immunol. 2018, 36, 579–601. [Google Scholar] [CrossRef]
- Dijkstra, J.M.; Somamoto, T.; Moore, L.; Hordvik, I.; Ototake, M.; Fischer, U. Identification and characterization of a second CD4-like gene in teleost fish. Mol. Immunol. 2006, 43, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Edholm, E.-S.; Stafford, J.L.; Quiniou, S.M.; Waldbieser, G.; Miller, N.W.; Bengtén, E.; Wilson, M. Channel catfish, Ictalurus punctatus, CD4-like molecules. Dev. Comp. Immunol. 2007, 31, 172–187. [Google Scholar] [CrossRef] [PubMed]
- Suetake, H.; Araki, K.; Suzuki, Y. Cloning, expression, and characterization of fugu CD4, the first ectothermic animal CD4. Immunogenetics 2004, 56, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.-F.; Shang, N.; Hu, W.; Wang, Y.-P.; Guo, Q.-L. Molecular cloning and characterization of carp (Cyprinus carpio L.) CD8β and CD4-like genes. Fish Shellfish Immunol. 2007, 23, 1242–1255. [Google Scholar] [CrossRef]
- Buonocore, F.; Randelli, E.; Casani, D.; Guerra, L.; Picchietti, S.; Costantini, S.; Facchiano, A.M.; Zou, J.; Secombes, C.J.; Scapigliati, G. A CD4 homologue in sea bass (Dicentrarchus labrax): Molecular characterization and structural analysis. Mol. Immunol. 2008, 45, 3168–3177. [Google Scholar] [CrossRef] [Green Version]
- Ashfaq, H.; Soliman, H.; Saleh, M.; El-Matbouli, M. CD4: A vital player in the teleost fish immune system. Veter Res. 2019, 50, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lyons, G.E.; Moore, T.; Brasic, N.; Li, M.; Roszkowski, J.J.; Nishimura, M.I. Influence of Human CD8 on Antigen Recognition by T-Cell Receptor–Transduced Cells. Cancer Res. 2006, 66, 11455–11461. [Google Scholar] [CrossRef] [Green Version]
- Nagarajan, U.M.; O’Connell, C.; Rank, R.G. Molecular characterization of guinea-pig (Cavia porcellus) CD8α and CD8β cDNA. Tissue Antigens 2004, 63, 184–189. [Google Scholar] [CrossRef]
- Ureta-Vidal, A.; Garcia, Z.; Lemonnier, F.A.; Kazanji, M. Molecular characterization of cDNAs encoding squirrel monkey (Saïmiri sciureus) CD8 α and β chains. Immunogenetics 1999, 49, 718–721. [Google Scholar] [CrossRef]
- Buonocore, F.; Randelli, E.; Bird, S.; Secombes, C.J.; Costantini, S.; Facchiano, A.; Mazzini, M.; Scapigliati, G. The CD8α from sea bass (Dicentrarchus labrax L.): Cloning, expression and 3D modelling. Fish Shellfish Immunol. 2006, 20, 637–646. [Google Scholar] [CrossRef]
- Randelli, E.; Foglietta, A.; Mazzini, M.; Scapigliati, G.; Buonocore, F. Cloning and expression analysis of the co-receptor CD8α in sea bream (Sparus aurata L.). Aquaculture 2006, 256, 631–637. [Google Scholar] [CrossRef]
- Somamoto, T.; Yoshiura, Y.; Nakanishi, T.; Ototake, M. Molecular cloning and characterization of two types of CD8α from ginbuna crucian carp, Carassius auratus langsdorfii. Dev. Comp. Immunol. 2005, 29, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Suetake, H.; Araki, K.; Akatsu, K.; Somamoto, T.; Dijkstra, J.M.; Yoshiura, Y.; Kikuchi, K.; Suzuki, Y. Genomic organization and expression of CD8α and CD8β genes in fugu Takifugu rubripes. Fish Shellfish Immunol. 2007, 23, 1107–1118. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.D.; Strassburger, P. Description of an Ectothermic TCR Coreceptor, CD8α, in Rainbow Trout. J. Immunol. 2000, 164, 3132–3139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, R.; Bernard, D.; Lefranc, M.; Six, A.; Benmansour, A.; Boudinot, P. T cell diversity and TcR repertoires in teleost fish. Fish Shellfish Immunol. 2011, 31, 644–654. [Google Scholar] [CrossRef]
- Randelli, E.; Buonocore, F.; Scapigliati, G. Cell markers and determinants in fish immunology. Fish Shellfish Immunol. 2008, 25, 326–340. [Google Scholar] [CrossRef] [Green Version]
- Xing, J.; Ma, J.; Tang, X.; Sheng, X.; Zhan, W. Characterizations of CD4-1, CD4-2 and CD8β T cell subpopulations in peripheral blood leucocytes, spleen and head kidney of Japanese flounder (Paralichthys olivaceus). Mol. Immunol. 2017, 85, 155–165. [Google Scholar] [CrossRef]
- Jung, J.W.; Lee, J.S.; Kim, Y.R.; Im, S.P.; Kim, S.W.; Lazarte, J.M.S.; Kim, J.; Thompson, K.D.; Suh, J.P.; Jung, T.S. Development of a monoclonal antibody against the CD3ε of olive flounder (Paralichthys olivaceus) and its application in evaluating immune response related to CD3ε. Fish Shellfish Immunol. 2017, 65, 179–185. [Google Scholar] [CrossRef]
- Jung, J.W.; Lee, J.S.; Kim, J.; Im, S.P.; Kim, S.W.; Lazarte, J.M.S.; Kim, Y.R.; Chun, J.H.; Ha, M.W.; Kim, H.S.; et al. Characterization of Hagfish (Eptatretus burgeri) Variable Lymphocyte Receptor–Based Antibody and Its Potential Role in the Neutralization of Nervous Necrosis Virus. J. Immunol. 2019, 204, 718–725. [Google Scholar] [CrossRef] [Green Version]
- López-Muñoz, A.; Sepulcre, M.P.; García-Moreno, D.; Fuentes, I.; Béjar, J.; Manchado, M.; Álvarez, M.C.; Meseguer, J.; Mulero, V. Viral nervous necrosis virus persistently replicates in the central nervous system of asymptomatic gilthead seabream and promotes a transient inflammatory response followed by the infiltration of IgM+ B lymphocytes. Dev. Comp. Immunol. 2012, 37, 429–437. [Google Scholar] [CrossRef]
- Chen, C.-W.; Wu, M.-S.; Huang, Y.-J.; Cheng, C.-A.; Chang, C.-Y. Recognition of Linear B-Cell Epitope of Betanodavirus Coat Protein by RG-M18 Neutralizing mAB Inhibits Giant Grouper Nervous Necrosis Virus (GGNNV) Infection. PLoS ONE 2015, 10, e0126121. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Jimena, B.; Garcia-Rosado, E.; Thompson, K.D.; Adams, A.; Infante, C.; Borrego, J.J.; Alonso, M.D.C. Distribution of red-spotted grouper nervous necrosis virus (RGNNV) antigens in nervous and non-nervous organs of European seabass (Dicentrarchus labrax) during the course of an experimental challenge. J. Veter Sci. 2012, 13, 355–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, T.-Y.; Arancibia, S.; Born, R.; Tampe, R.; Villar, J.; Del Campo, M.; Manubens, A.; Becker, M.I. Hemocyanins Stimulate Innate Immunity by Inducing Different Temporal Patterns of Proinflammatory Cytokine Expression in Macrophages. J. Immunol. 2016, 196, 4650–4662. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.R.; Markl, J. Keyhole limpet hemocyanin (KLH): A biomedical review. Micron 1999, 30, 597–623. [Google Scholar] [CrossRef]
- Needham, B.D.; Carroll, S.M.; Giles, D.K.; Georgiou, G.; Whiteley, M.; Trent, M.S. Modulating the innate immune response by combinatorial engineering of endotoxin. Proc. Natl. Acad. Sci. USA 2013, 110, 1464–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swain, P.S.; Nayak, S.K.; Nanda, P.K.; Dash, S. Biological effects of bacterial lipopolysaccharide (endotoxin) in fish: A review. Fish Shellfish Immunol. 2008, 25, 191–201. [Google Scholar] [CrossRef]
- Xing, J.; Tian, H.-F.; Tang, X.-Q.; Sheng, X.-Z.; Zhan, W.-B. Kinetics of T lymphocyte subsets and B lymphocytes in response to immunostimulants in flounder (Paralichthys oliveceus): Implications for CD4+ T lymphocyte differentiation. Sci. Rep. 2020, 10, 13827. [Google Scholar] [CrossRef]
- Harlow, E.; Lane, D. A Laboratory Manual; Cold Spring Harbor Laboratory: New York, NY, USA, 1988; p. 579. [Google Scholar]
- Husband, A.J. Overview of the mammalian immune system. In Advances in Nutritional Research; Springer: Boston, MA, USA, 2001; pp. 3–14. [Google Scholar]
- Laing, K.J.; Hansen, J.D. Fish T cells: Recent advances through genomics. Dev. Comp. Immunol. 2011, 35, 1282–1295. [Google Scholar] [CrossRef]
- Toda, H.; Saito, Y.; Koike, T.; Takizawa, F.; Araki, K.; Yabu, T.; Somamoto, T.; Suetake, H.; Suzuki, Y.; Ototake, M. Conservation of characteristics and functions of CD4 positive lymphocytes in a teleost fish. Dev. Comp. Immunol. 2011, 35, 650–660. [Google Scholar] [CrossRef]
- Quiniou, S.M.A.; Sahoo, M.; Edholm, E.-S.; Bengtén, E.; Wilson, M. Channel catfish CD8α and CD8β co-receptors: Characterization, expression and polymorphism. Fish Shellfish Immunol. 2011, 30, 894–901. [Google Scholar] [CrossRef]
- Forlenza, M.; de CarvalhoDias, J.D.A.; Veselý, T.; Pokorová, D.; Savelkoul, H.F.J.; Wiegertjes, G.F. Transcription of signal-3 cytokines, IL-12 and IFNαβ, coincides with the timing of CD8αβ up-regulation during viral infection of common carp (Cyprinus carpio L.). Mol. Immunol. 2008, 45, 1531–1547. [Google Scholar] [CrossRef] [PubMed]
- Workenhe, S.T.; Rise, M.L.; Kibenge, M.J.; Kibenge, F.S. The fight between the teleost fish immune response and aquatic viruses. Mol. Immunol. 2010, 47, 2525–2536. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, S.-C.; Jiang, G.; Yang, N.; Lian, J.; Yang, Y. The development of the lymphoid organs of flounder, Paralichthys olivaceus, from hatching to 13 months. Fish Shellfish Immunol. 2004, 16, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Hetland, D.L.; Jørgensen, S.M.; Skjødt, K.; Dale, O.B.; Falk, K.; Xu, C.; Mikalsen, A.B.; Grimholt, U.; Gjøen, T.; Press, C.M. In situ localisation of major histocompatibility complex class I and class II and CD8 positive cells in infectious salmon anaemia virus (ISAV)-infected Atlantic salmon. Fish Shellfish Immunol. 2010, 28, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Haugarvoll, E.; Bjerkås, I.; Nowak, B.F.; Hordvik, I.; Koppang, E. Identification and characterization of a novel intraepithelial lymphoid tissue in the gills of Atlantic salmon. J. Anat. 2008, 213, 202–209. [Google Scholar] [CrossRef]
- Jiang, X.-B.; Wang, Z.-D.; Zhu, Y.; Zhang, X.-L.; Cui, X.-F.; Yao, K.; Yin, K.-S. Inhibition of CD8+ T lymphocytes attenuates respiratory syncytial virus-enhanced allergic inflammation. Respiration 2009, 77, 76–84. [Google Scholar] [CrossRef]
- Jabri, B.; Ebert, E. Human CD8+ intraepithelial lymphocytes: A unique model to study the regulation of effector cytotoxic T lymphocytes in tissue. Immunol. Rev. 2007, 215, 202–214. [Google Scholar] [CrossRef]
- Kunisawa, J.; Takahashi, I.; Kiyono, H. Intraepithelial lymphocytes: Their shared and divergent immunological behaviors in the small and large intestine. Immunol. Rev. 2007, 215, 136–153. [Google Scholar] [CrossRef]
- Zuckermann, F.A. Extrathymic CD4/CD8 double positive T cells. Veter Immunol. Immunopathol. 1999, 72, 55–66. [Google Scholar] [CrossRef]
- Laky, K.; Fleischacker, C.; Fowlkes, B. TCR and Notch signaling in CD4 and CD8 T-cell development. Immunol. Rev. 2006, 209, 274–283. [Google Scholar] [CrossRef]
- Ellmeier, W.; Sawada, S.; Littman, D.R. The regulation of CD4 and CD8 coreceptor gene expression during T cell development. Annu. Rev. Immunol. 1999, 17, 523–554. [Google Scholar] [CrossRef] [PubMed]
- Guidotti, L.G.; Chisari, F.V. Noncytolytic control of viral infections by the innate and adaptive immuneresponse. Annu. Rev. Immunol. 2001, 19, 65–91. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, A.; Lucas, R.M.; Dear, K.; McMichael, A.J. Keyhole limpet haemocyanin—A model antigen for human immunotoxicological studies. Br. J. Clin. Pharmacol. 2014, 78, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Kono, T.; Korenaga, H. Cytokine Gene Expression in CD4 Positive Cells of the Japanese Pufferfish, Takifugu rubripes. PLoS ONE 2013, 8, e66364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, T.; Katakura, F.; Someya, K.; Dijkstra, J.M.; Moritomo, T.; Nakanishi, T. Clonal growth of carp (Cyprinus carpio) T cells in vitro: Long-term proliferation of Th2-like cells. Fish Shellfish Immunol. 2013, 34, 433–442. [Google Scholar] [CrossRef]
- Biedermann, T.; Röcken, M.; Carballido, J.M. TH1 and TH2 Lymphocyte development and regulation of TH Cell–mediated immune responses of the skin. J. Investig. Dermatol. Symp. Proc. 2004, 9, 5–14. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Start | End | Peptide Sequence | Length |
|---|---|---|---|---|
| 1 | 37 | 46 | CKPAEMFNTV | 10 |
| 2 | 60 | 72 | IASFGRDGKMKSN | 13 |
| 3 | 107 | 114 | TIIQSNEM | 8 |
| No | Start | End | Peptide Sequence | Length |
|---|---|---|---|---|
| 1 | 44 | 53 | CNCNNSCDSV | 10 |
| 2 | 68 | 80 | LGKCNNAERVNYG | 13 |
| 3 | 116 | 123 | VLKAKSGT | 8 |
| Primer Name | Sequence (5′-3′) | |
|---|---|---|
| CD4-1 | Forward | TATAGGCCACCGGGGCCATGGAGAAGTTTGTCCTCATTC |
| Reverse | GGTGGGCCCCAGAGGCCTGTTCTGTAGAATCCTTTGGGC | |
| CD4-2 | Forward | TATAGGCCACCGGGGCCATGAACGTCATTGTGTTGTTTGGA |
| Reverse | GGTGGGCCCCAGAGGCCCTCCTTTAGCAGGGGCTTCAG | |
| CD8α | Forward | TATAGGCCACCGGGGCCATGGACCAAAAGTGGATTCAG |
| Reverse | GGTGGGCCCCAGAGGCCAACATGTGTGTTGTTCTTCATCTG | |
| CD8β | Forward | TATAGGCCACCGGGGCCATGAACCCGCTGCCGCTG |
| Reverse | GGTGGGCCCCAGAGGCCGGGCATCTGTCTCATCTTCTG |
| Primer Name | Sequence (5′-3′) | |
|---|---|---|
| CD3ε (AB081751.1) | Forward | ATGAAAATCAACACCATGGATGTC |
| Reverse | TCCCGTCCTGTTCACAATAGA | |
| CD4-1 (AB643634.1) | Forward | ATGAATCCCAGAGGAGAGATAATG |
| Reverse | CACGTAGTCTCCTCCGTCTTC | |
| CD4-2 (AB640684.1) | Forward | GTGATCCTAACAAAACCCAGGCAG |
| Reverse | AGCAGGTTCTTCAACTTTGATCTT | |
| CD8α (AB082957.1) | Forward | ATGGACCAAAAGTGGATTCAGATG |
| Reverse | AACATGTGTGTTGTTCTTCATCTG | |
| CD8β (AB643633.1) | Forward | ATGAACCCGCTGCCGCTG |
| Reverse | GGGCATCTGTCTCATCTTCTG | |
| TCRα (AB053227.1) | Forward | ATGCTCTCACTGCATCTTGGT |
| Reverse | GACTCTGTGACTGAGCCACAG | |
| TCRβ (AB053228.1) | Forward | ATGATTCCAAGCCTCAACACC |
| Reverse | GTGGTTCTGCTTCTCAGCTGA | |
| IgL1 (AB819734.1) | Forward | ATGAGCTTTACCTCCGTCCTC |
| Reverse | GGACTGGGAACACTGGTCTCT | |
| IgM (AB052744.1) | Forward | ATGTTTCCTGTAGCTGTGCTG |
| Reverse | CTGGGCCTTGCATGGTATGTT | |
| β-actin (HQ386788.1) | Forward | ATGGAAGATGAAATCGCCGCA |
| Reverse | GAAGCATTTGCGGTGGACGAT |
| NNV | KLH | LPS | |||||
|---|---|---|---|---|---|---|---|
| DPI (Days post infection) | Peak (Mean ± SD) | DPA (Days post administration) | Peak (Mean±SD) | DPA (Days post administration) | Peak (Mean±SD) | ||
| CD4-1 lymphocytes | Spleen | 14 | 18.44 ± 1.36 * | 7 | 11.82 ± 1.66 * | 7 | 6.53 ± 1.84 |
| Head-kidney | 7 | 22.59 ± 3.06 * | 7 | 14.50 ± 2.04 * | 1 | 6.63 ± 0.56 | |
| Peripheral blood | 7 | 10.00 ± 0.90 * | 7 | 8.21 ± 1.3 4 * | 3 | 3.52 ± 0.87 | |
| NNV | KLH | LPS | |||||
| DPI (Days post infection) | Peak (Mean±SD) | DPA (Days post administration) | Peak (Mean±SD) | DPA (Days post administration) | Peak (Mean±SD) | ||
| CD4-2 lymphocytes | Spleen | 7 | 32.59 ± 2.79 * | 7 | 25.48 ± 2.21 * | 14 | 10.99 ± 2.08 |
| Head-kidney | 7 | 33.62 ± 1.47 * | 7 | 22.36 ± 3.08 * | 0 | 14.89 ± 1.11 | |
| Peripheral blood | 7 | 14.19 ± 0.72 * | 7 | 9.60 ± 1.00 * | 14 | 5.20 ± 0.38 | |
| NNV | KLH | LPS | |||||
| DPI (Days post infection) | Peak (Mean ± SD) | DPA (Days post administration) | Peak (Mean ± SD) | DPA (Days post administration) | Peak (Mean ± SD) | ||
| CD8α lymphocytes | Spleen | 3 | 13.31 ± 0.88 * | 3 | 13.03 ± 1.44 * | 1 | 5.77 ± 1.25 |
| Head-kidney | 3 | 12.87 ± 0.41 * | 3 | 13.41 ± 0.87 * | 14 | 7.07 ± 0.96 | |
| Peripheral blood | 3 | 10.06 ± 1.83 * | 3 | 6.26 ± 0.63 * | 1 | 2.67 ± 0.57 | |
| NNV | KLH | LPS | |||||
| DPI (Days post infection) | Peak (Mean ± SD) | DPA (Days post administration) | Peak (Mean ± SD) | DPA (Days post administration) | Peak (Mean ± SD) | ||
| CD8β lymphocytes | Spleen | 3 | 15.98 ± 1.05 * | 3 | 14.17 ± 2.94 * | 1 | 7.24 ± 1.53 |
| Head-kidney | 3 | 20.66 ± 0.68 * | 3 | 16.83 ± 2.50 * | 7 | 8.05 ± 0.61 | |
| Peripheral blood | 3 | 13.13 ± 1.11 * | 3 | 8.47 ± 1.62 * | 1 | 3.85 ± 0.71 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, J.W.; Lee, A.R.; Kim, J.; Kim, Y.R.; Lazarte, J.M.S.; Lee, J.S.; Thompson, K.D.; Kim, H.; Jung, T.S. Elucidating the Functional Roles of Helper and Cytotoxic T Cells in the Cell-Mediated Immune Responses of Olive Flounder (Paralichthys olivaceus). Int. J. Mol. Sci. 2021, 22, 847. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020847
Jung JW, Lee AR, Kim J, Kim YR, Lazarte JMS, Lee JS, Thompson KD, Kim H, Jung TS. Elucidating the Functional Roles of Helper and Cytotoxic T Cells in the Cell-Mediated Immune Responses of Olive Flounder (Paralichthys olivaceus). International Journal of Molecular Sciences. 2021; 22(2):847. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020847
Chicago/Turabian StyleJung, Jae Wook, Ae Rin Lee, Jaesung Kim, Young Rim Kim, Jassy Mary S. Lazarte, Jung Suk Lee, Kim D. Thompson, Hyeongsu Kim, and Tae Sung Jung. 2021. "Elucidating the Functional Roles of Helper and Cytotoxic T Cells in the Cell-Mediated Immune Responses of Olive Flounder (Paralichthys olivaceus)" International Journal of Molecular Sciences 22, no. 2: 847. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020847