
Primary Ciliogenesis by 2-Isopropylmalic Acid Prevents PM2.5-Induced Inflammatory Response and MMP-1 Activation in Human Dermal Fibroblasts and a 3-D-Skin Model

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Result
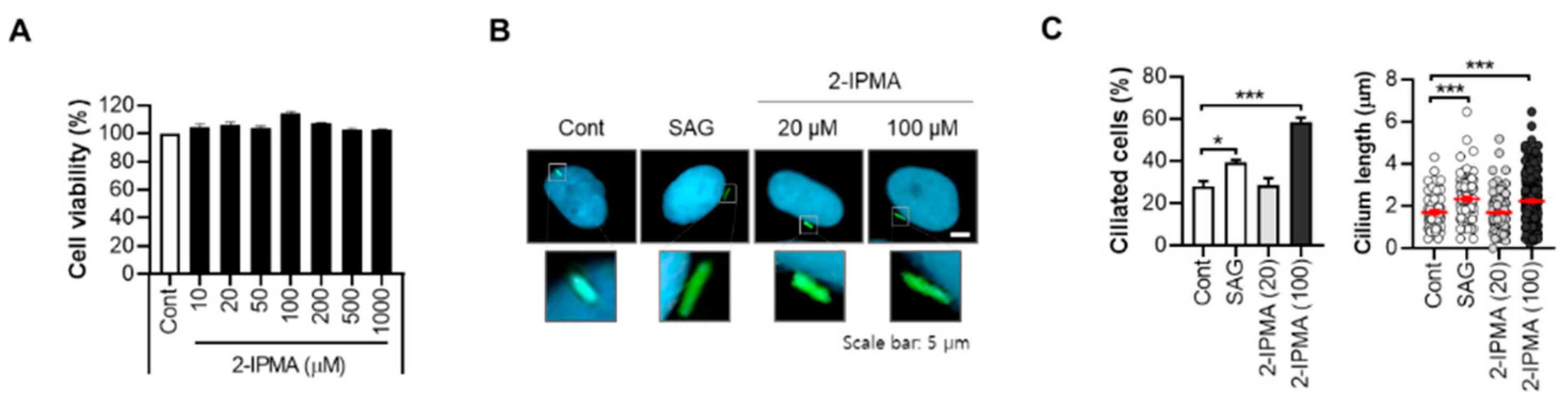
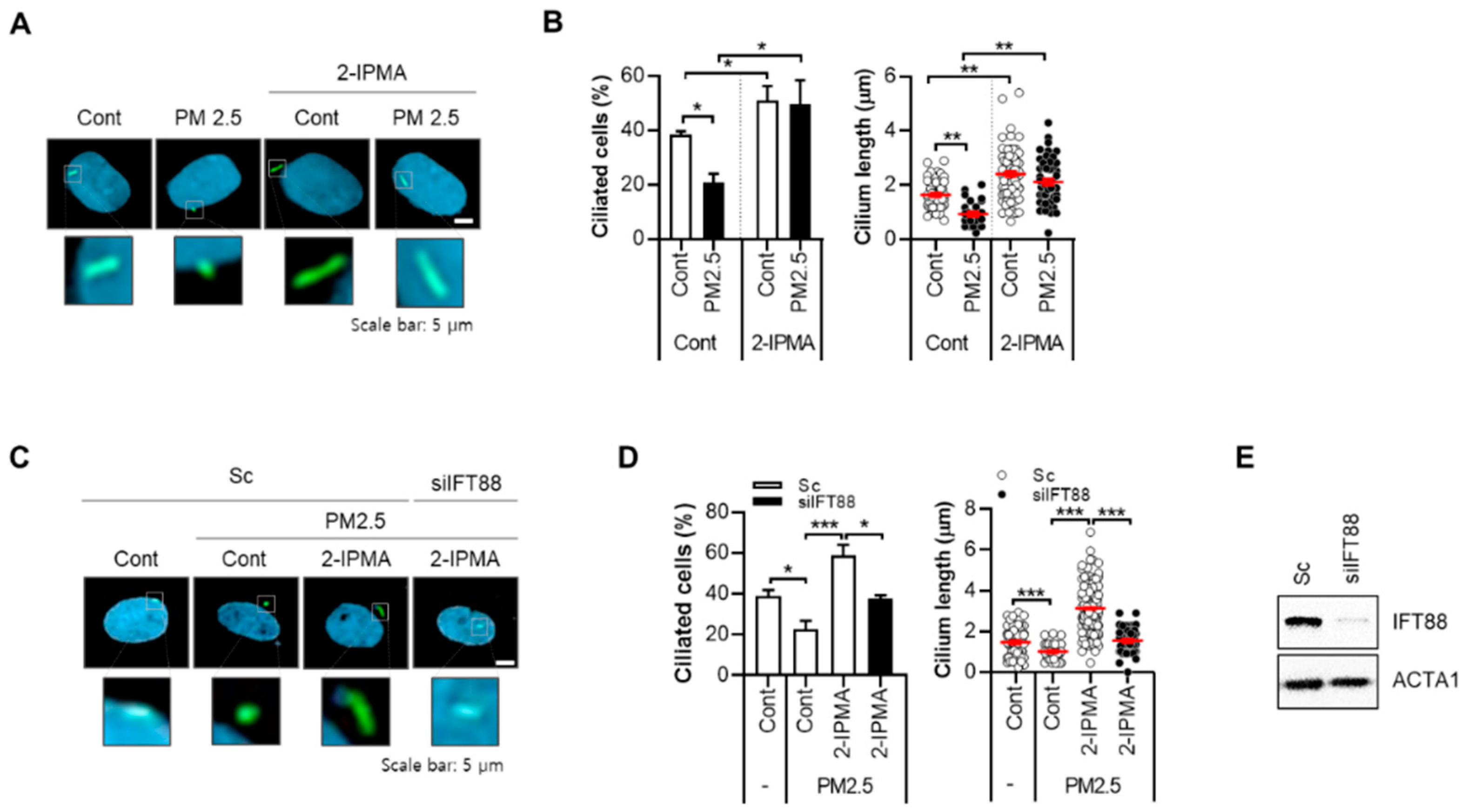
2.1. 2-IPMA Restores PM2.5-Mediated Ciliary Dysgenesis in Normal Dermal Fibroblasts
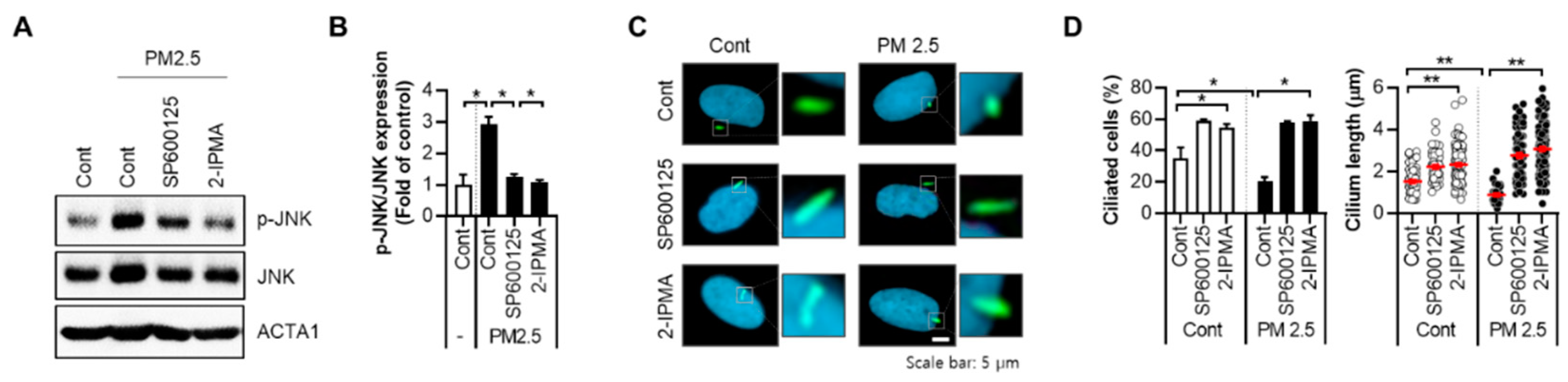
2.2. 2-IPMA Inhibits PM2.5-Induced JNK Activation and Inflammatory Response in Normal Dermal Fibroblasts
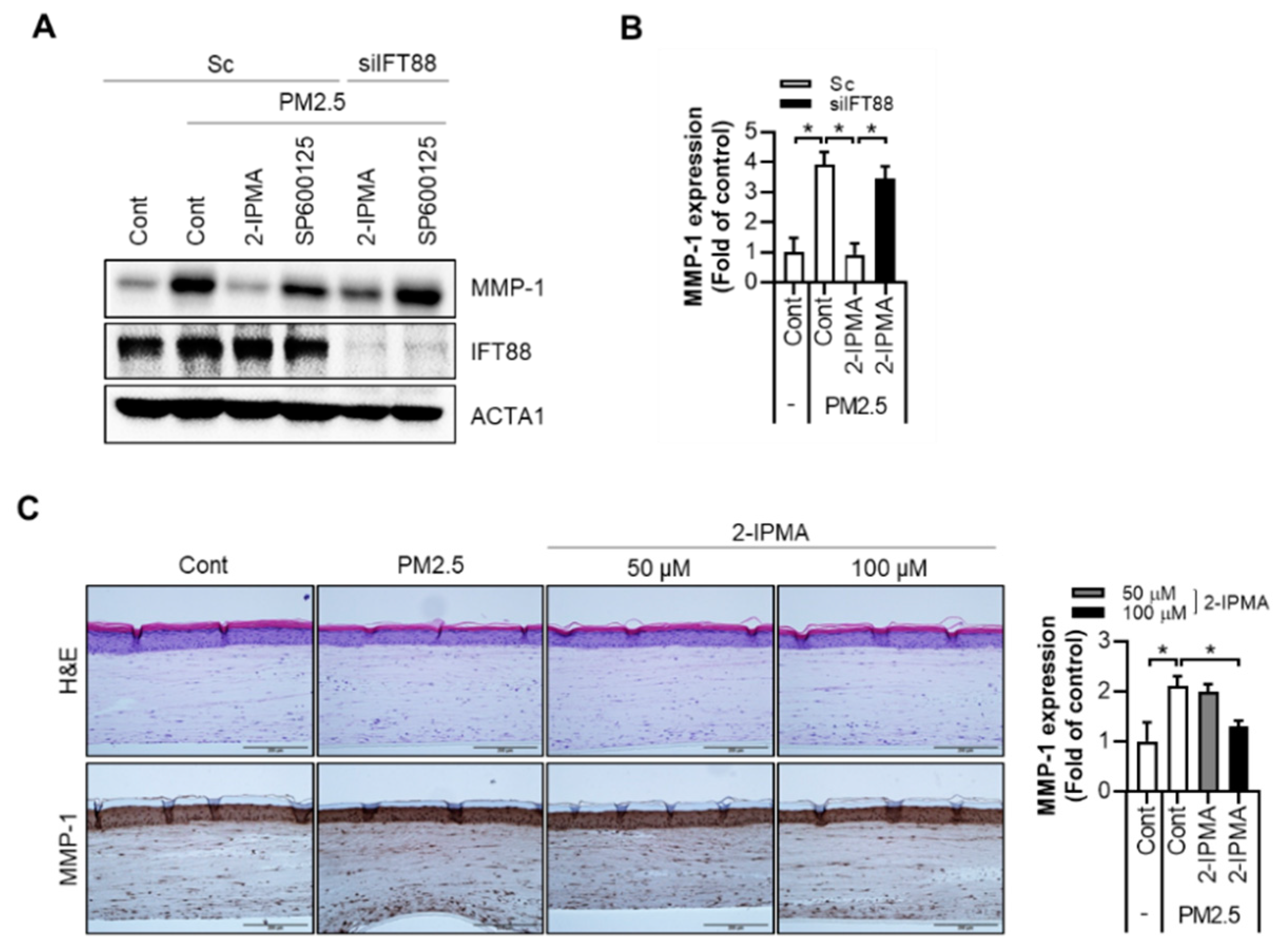
2.3. 2-IPMA Downregulates MMP-1 Expression by PM2.5 in Normal Dermal Fibroblasts and a 3-D-Skin Model
3. Discussion
4. Material and Method
4.1. Cell Culture
4.2. Reagents and Cell Viability
4.3. Western Blot Analysis and Densitometry Analysis
4.4. Primary Cilia Staining and Counting
4.5. ELISA Assay
4.6. Reconstruction of Full-Thickness Human 3-D-Skin Model
4.7. Histological Examination
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lippmann, M. Toxicological and epidemiological studies of cardiovascular effects of ambient air fine particulate matter (PM2.5) and its chemical components: Coherence and public health implications. Crit. Rev. Toxicol. 2014, 44, 299–347. [Google Scholar] [CrossRef]
- Pun, V.C.; Kazemiparkouhi, F.; Manjourides, J.; Suh, H.H. Long-Term PM2.5 exposure and respiratory, cancer, and cardiovascular mortality in older US adults. Am. J. Epidemiol. 2017, 186, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-N.; Lim, Y.-H.; Bae, H.J.; Kim, M.; Jung, K.; Hong, Y.-C. Long-term fine particulate matter exposure and major depressive disorder in a community-based urban cohort. Environ. Health Perspect. 2016, 124, 1547–1553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.-P.; Li, Z.; Choi, E.K.; Lee, S.; Kim, Y.K.; Seo, E.Y.; Chung, J.H.; Cho, S. Urban particulate matter in air pollution penetrates into the barrier-disrupted skin and produces ROS-dependent cutaneous inflammatory response in vivo. J. Dermatol. Sci. 2018, 91, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Satir, P.; Pedersen, L.; Christensen, S.T. The primary cilium at a glance. J. Cell Sci. 2010, 123, 499–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anvarian, Z.; Mykytyn, K.; Mukhopadhyay, S.; Pedersen, L.; Christensen, S.T. Cellular signalling by primary cilia in development, organ function and disease. Nat. Rev. Nephrol. 2019, 15, 199–219. [Google Scholar] [CrossRef] [PubMed]
- Toriyama, M.; Ishii, K.J. Primary cilia in the skin: Functions in immunity and therapeutic potential. Front. Cell Dev. Biol. 2021, 9, 621318. [Google Scholar] [CrossRef]
- Lehman, J.; Laag, E.; Michaud, E.J.; Yoder, B.K. An essential role for dermal primary cilia in hair follicle morphogenesis. J. Investig. Dermatol. 2009, 129, 438–448. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.; Shin, J.H.; Kim, E.S.; Park, S.J.; Bae, I.-H.; Jo, Y.K.; Jeong, I.Y.; Kim, H.-J.; Lee, Y.; Park, H.C.; et al. Primary cilia negatively regulate melanogenesis in melanocytes and pigmentation in a human skin model. PLoS ONE 2016, 11, e0168025. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.-E.; Choi, H.; Shin, D.W.; Na, H.-W.; Park, N.Y.; Kim, J.B.; Jo, D.S.; Cho, M.J.; Lyu, J.H.; Chang, J.H.; et al. Fine particulate matter (PM2.5) inhibits ciliogenesis by increasing SPRR3 expression via c-Jun activation in RPE cells and skin keratinocytes. Sci. Rep. 2019, 9, 3994. [Google Scholar] [CrossRef]
- Strassman, M.; Ceci, L.N. Enzymatic Formation of α-isopropylmalic acid, an intermediate in leucine biosynthesis. J. Biol. Chem. 1963, 238, 2445–2452. [Google Scholar] [CrossRef]
- Ricciutelli, M.; Moretti, S.; Galarini, R.; Sagratini, G.; Mari, M.; Lucarini, S.; Vittori, S.; Caprioli, G. Identification and quantification of new isomers of isopropyl-malic acid in wine by LC-IT and LC-Q-Orbitrap. Food Chem. 2019, 294, 390–396. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Tamura, S.; Nakanishi, H.; Tashiro, M.; Nishizawa, N.K.; Yoshimura, E. Reduction of aluminum toxicity by 2-isopropylmalic acid in the budding yeast saccharomyces cerevisiae. Biol. Trace Element Res. 2007, 120, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Kobayashi, T.; Nishiumi, S.; Hidaka, A.; Yamaji, T.; Sawada, N.; Hirata, Y.; Yamanaka, K.; Azuma, T.; Goto, A.; et al. Metabolome analysis for pancreatic cancer risk in nested case-control study: Japan Public Health Center-based prospective Study. Cancer Sci. 2018, 109, 1672–1681. [Google Scholar] [CrossRef] [Green Version]
- Skogerson, K.; Runnebaum, R.; Wohlgemuth, G.; de Ropp, J.; Heymann, H.; Fiehn, O. Comparison of gas chromatography-coupled time-of-flight mass spectrometry and 1H nuclear magnetic resonance spectroscopy metabolite identification in white wines from a sensory study investigating wine body. J. Agric. Food Chem. 2009, 57, 6899–6907. [Google Scholar] [CrossRef]
- Ricciutelli, M.; Bartolucci, G.; Campana, R.; Salucci, S.; Benedetti, S.; Caprioli, G.; Maggi, F.; Sagratini, G.; Vittori, S.; Lucarini, S. Quantification of 2- and 3-isopropylmalic acids in forty Italian wines by UHPLC-MS/MS triple quadrupole and evaluation of their antimicrobial, antioxidant activities and biocompatibility. Food Chem. 2020, 321, 126726. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wei, Q.; Zhang, Y.; Ling, K.; Hu, J. The small GTPases ARL-13 and ARL-3 coordinate intraflagellar transport and ciliogenesis. J. Cell Biol. 2010, 189, 1039–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyun, Y.J.; Piao, M.J.; Kang, K.A.; Zhen, A.X.; Madushan Fernando, P.D.S.; Kang, H.K.; Ahn, Y.S.; Hyun, J.W. Effect of fer-mented fish oil on fine particulate matter-induced skin aging. Mar. Drugs 2019, 17, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, W.J.; Hanson, P.S.; Critchley, A.; Griffiths, B.; Chavan, B.; Birch-Machin, M.A. Exposing human primary dermal fibroblasts to particulate matter induces changes associated with skin aging. FASEB J. 2020, 34, 14725–14735. [Google Scholar] [CrossRef] [PubMed]
- Boyle, C.J.; Plotczyk, M.; Villalta, S.F.; Patel, S.; Hettiaratchy, S.; Masouros, S.D.; Masen, M.A.; Higgins, C.A. Morphology and composition play distinct and complementary roles in the tolerance of plantar skin to mechanical load. Sci. Adv. 2019, 5, eaay0244. [Google Scholar] [CrossRef] [Green Version]
- Przekora, A. A Concise review on tissue engineered artificial skin grafts for chronic wound treatment: Can we reconstruct functional skin tissue in vitro? Cells 2020, 9, 1622. [Google Scholar] [CrossRef]
- Choi, J.Y.; Bae, J.-E.; Kim, J.B.; Jo, D.S.; Park, N.Y.; Kim, Y.H.; Lee, H.J.; Kim, S.H.; Kim, S.H.; Jeon, H.B.; et al. 2-IPMA Ameliorates PM2.5-induced inflammation by promoting primary ciliogenesis in RPE cells. Molecules 2021, 26, 5409. [Google Scholar] [CrossRef]
- Marobbio, C.M.T.; Giannuzzi, G.; Paradies, E.; Pierri, C.L.; Palmieri, F. α-Isopropylmalate, a leucine biosynthesis intermediate in yeast, is transported by the mitochondrial oxalacetate carrier. J. Biol. Chem. 2008, 283, 28445–28453. [Google Scholar] [CrossRef] [Green Version]
- Rembiesa, J.; Ruzgas, T.; Engblom, J.; Holefors, A. The Impact of pollution on skin and proper efficacy testing for anti-pollution claims. Cosmetics 2018, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Roberts, W. Air pollution and skin disorders. Int. J. Women’s Dermatol. 2021, 7, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.-S.; Kim, S.; Lee, S.-W.; Jung, J.; Lee, S.; Na, H.-W.; Kim, H.-J.; Hong, Y.; Park, W.; Lee, T.; et al. Molecule-resolved visualization of particulate matter on human skin using multimodal nonlinear optical imaging. Int. J. Mol. Sci. 2021, 22, 5199. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.-W.; Lee, H.-S.; Na, J.-I.; Huh, C.-H.; Park, K.-C.; Choi, H.-R. Resveratrol inhibits particulate matter-induced inflammatory responses in human keratinocytes. Int. J. Mol. Sci. 2020, 21, 3446. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Li, X.; Nan, A.; Zhang, N.; Chen, L.; Zhou, H.; Zhang, H.; Qiu, M.; Zhu, J.; Ling, Y.; et al. Circular RNA 406961 interacts with ILF2 to regulate PM2.5-induced inflammatory responses in human bronchial epithelial cells via activation of STAT3/JNK pathways. Environ. Int. 2020, 141, 105755. [Google Scholar] [CrossRef]
- Wang, J.; Huang, J.; Wang, L.; Chen, C.; Yang, D.; Jin, M.; Bai, C.; Song, Y. Urban particulate matter triggers lung inflammation via the ROS-MAPK-NF-κB signaling pathway. J. Thorac. Dis. 2017, 9, 4398–4412. [Google Scholar] [CrossRef] [Green Version]
- Zhen, A.X.; Piao, M.J.; Kang, K.A.; Fernando, P.D.S.M.; Kang, H.K.; Koh, Y.S.; Yi, J.M.; Hyun, J.W. Niacinamide protects skin cells from oxidative stress induced by particulate matter. Biomol. Ther. 2019, 27, 562–569. [Google Scholar] [CrossRef]
- Diao, P.; He, H.; Tang, J.; Xiong, L.; Li, L. Natural compounds protect the skin from airborne particulate matter by attenuating oxidative stress. Biomed. Pharmacother. 2021, 138, 111534. [Google Scholar] [CrossRef]
- Tan, P.; Thiyagarasaiyar, K.; Tan, C.-Y.; Jeon, Y.-J.; Nadzir, M.; Wu, Y.-J.; Low, L.-E.; Atanasov, A.; Ming, L.; Liew, K.; et al. Algae-derived anti-inflammatory compounds against particulate matters-induced respiratory diseases: A systematic review. Mar. Drugs 2021, 19, 317. [Google Scholar] [CrossRef] [PubMed]
- Roztocil, E.; Hammond, C.L.; Gonzalez, M.O.; Feldon, S.E.; Woeller, C.F. The aryl hydrocarbon receptor pathway controls matrix metalloproteinase-1 and collagen levels in human orbital fibroblasts. Sci. Rep. 2020, 10, 8477. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zeng, Z.; Liu, J.; Pi, Z.; Zou, P.; Deng, Q.; Ma, X.; Qiao, F.; Xiong, W.; Zhou, C.; et al. Particulate matter promotes hyperpigmentation via AhR/MAPK signaling activation and by increasing α-MSH paracrine levels in keratinocytes. Environ. Pollut. 2021, 278, 116850. [Google Scholar] [CrossRef]
- Villa, M.; Crotta, S.; Dingwell, K.S.; Hirst, E.M.A.; Gialitakis, M.; Ahlfors, H.; Smith, J.C.; Stockinger, B.; Wack, A. The aryl hydrocarbon receptor controls cyclin O to promote epithelial multiciliogenesis. Nat. Commun. 2016, 7, 12652. [Google Scholar] [CrossRef]
- Ryu, Y.S.; Kang, K.A.; Piao, M.J.; Ahn, M.J.; Yi, J.M.; Bossis, G.; Hyun, Y.-M.; Park, C.O.; Hyun, J.W. Particulate matter-induced senescence of skin keratinocytes involves oxidative stress-dependent epigenetic modifications. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Bae, I.-H.; Choi, H.; Choi, H.W.; Oh, S.; Marinho, P.A.; Min, D.J.; Kim, D.-Y.; Lee, T.R.; Lee, C.S.; et al. Ameliorating effect of dipotassium glycyrrhizinate on an IL-4- and IL-13-induced atopic dermatitis-like skin-equivalent model. Arch. Dermatol. Res. 2019, 311, 131–140. [Google Scholar] [CrossRef]
- Min, D.; Park, S.; Kim, H.-J.; Lee, S.H.; Ahn, Y.; Jung, W.; Cho, Y.W. Potential anti-ageing effect of chondroitin sulphate through skin regeneration. Int. J. Cosmet. Sci. 2020, 42, 520–527. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, J.-E.; Min, D.; Choi, J.Y.; Choi, H.; Kim, J.B.; Park, N.Y.; Jo, D.S.; Kim, Y.H.; Na, H.-W.; Kim, Y.J.; et al. Primary Ciliogenesis by 2-Isopropylmalic Acid Prevents PM2.5-Induced Inflammatory Response and MMP-1 Activation in Human Dermal Fibroblasts and a 3-D-Skin Model. Int. J. Mol. Sci. 2021, 22, 10941. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222010941
Bae J-E, Min D, Choi JY, Choi H, Kim JB, Park NY, Jo DS, Kim YH, Na H-W, Kim YJ, et al. Primary Ciliogenesis by 2-Isopropylmalic Acid Prevents PM2.5-Induced Inflammatory Response and MMP-1 Activation in Human Dermal Fibroblasts and a 3-D-Skin Model. International Journal of Molecular Sciences. 2021; 22(20):10941. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222010941
Chicago/Turabian StyleBae, Ji-Eun, Daejin Min, Ji Yeon Choi, Hyunjung Choi, Joon Bum Kim, Na Yeon Park, Doo Sin Jo, Yong Hwan Kim, Hye-Won Na, Yoon Jae Kim, and et al. 2021. "Primary Ciliogenesis by 2-Isopropylmalic Acid Prevents PM2.5-Induced Inflammatory Response and MMP-1 Activation in Human Dermal Fibroblasts and a 3-D-Skin Model" International Journal of Molecular Sciences 22, no. 20: 10941. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222010941