
Molecular Insights on the Possible Role of Annexin A2 in COVID-19 Pathogenesis and Post-Infection Complications

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
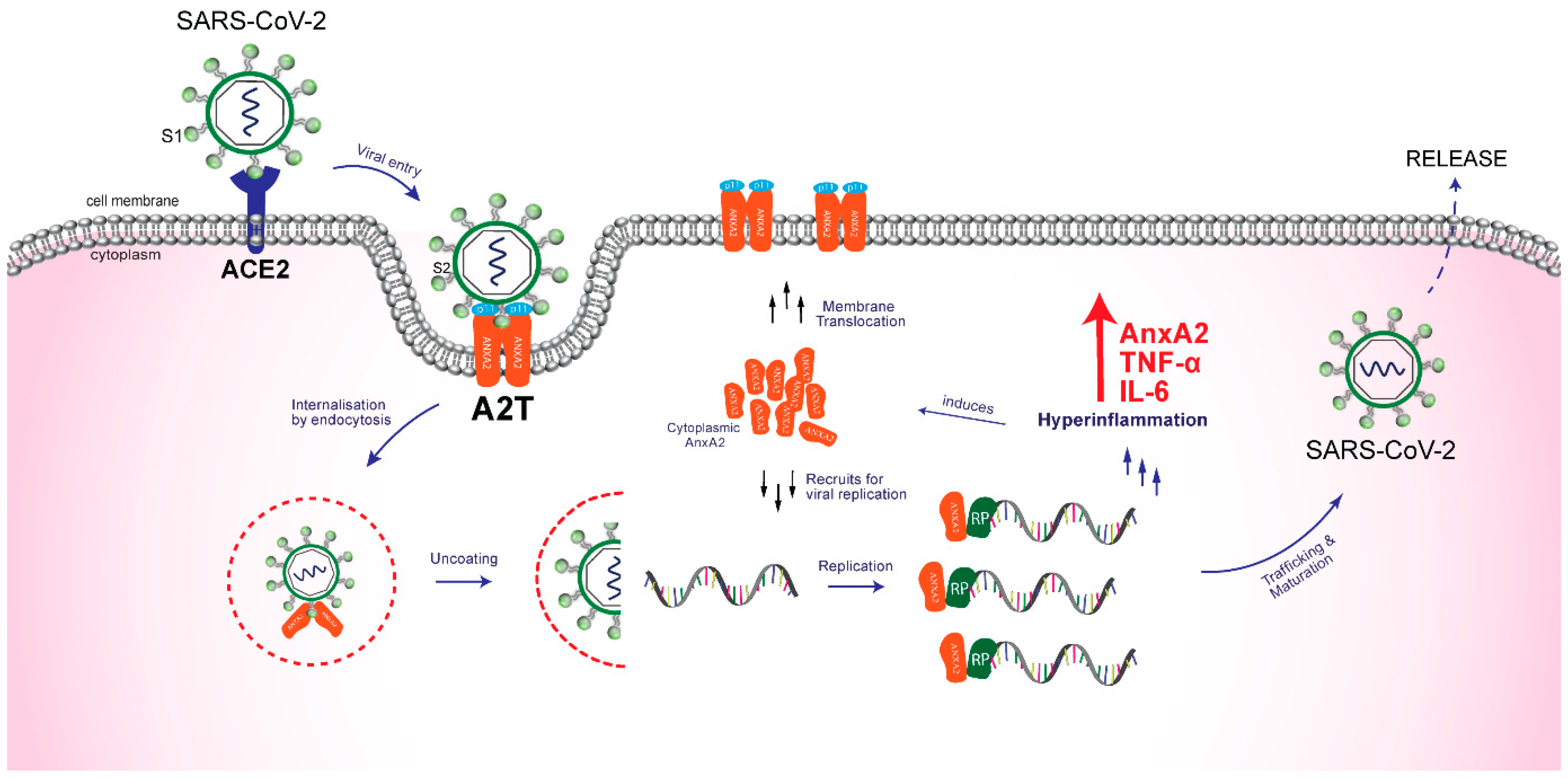
2. AnxA2 Role in SARS-CoV-2 Pathogenesis
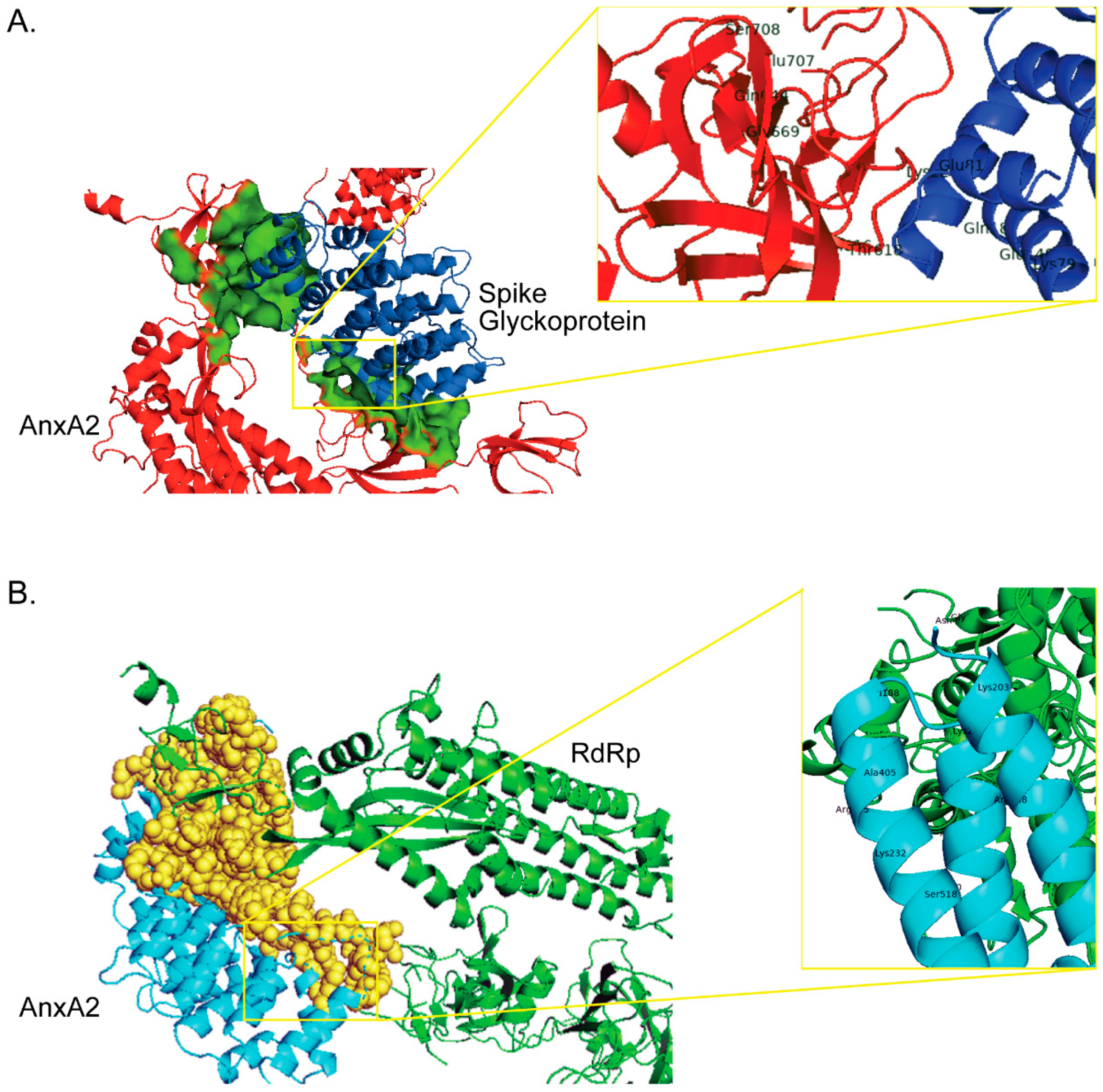
2.1. Role in Attachment and Internalization of the Virus
2.2. Role in Replication and Release of Virus
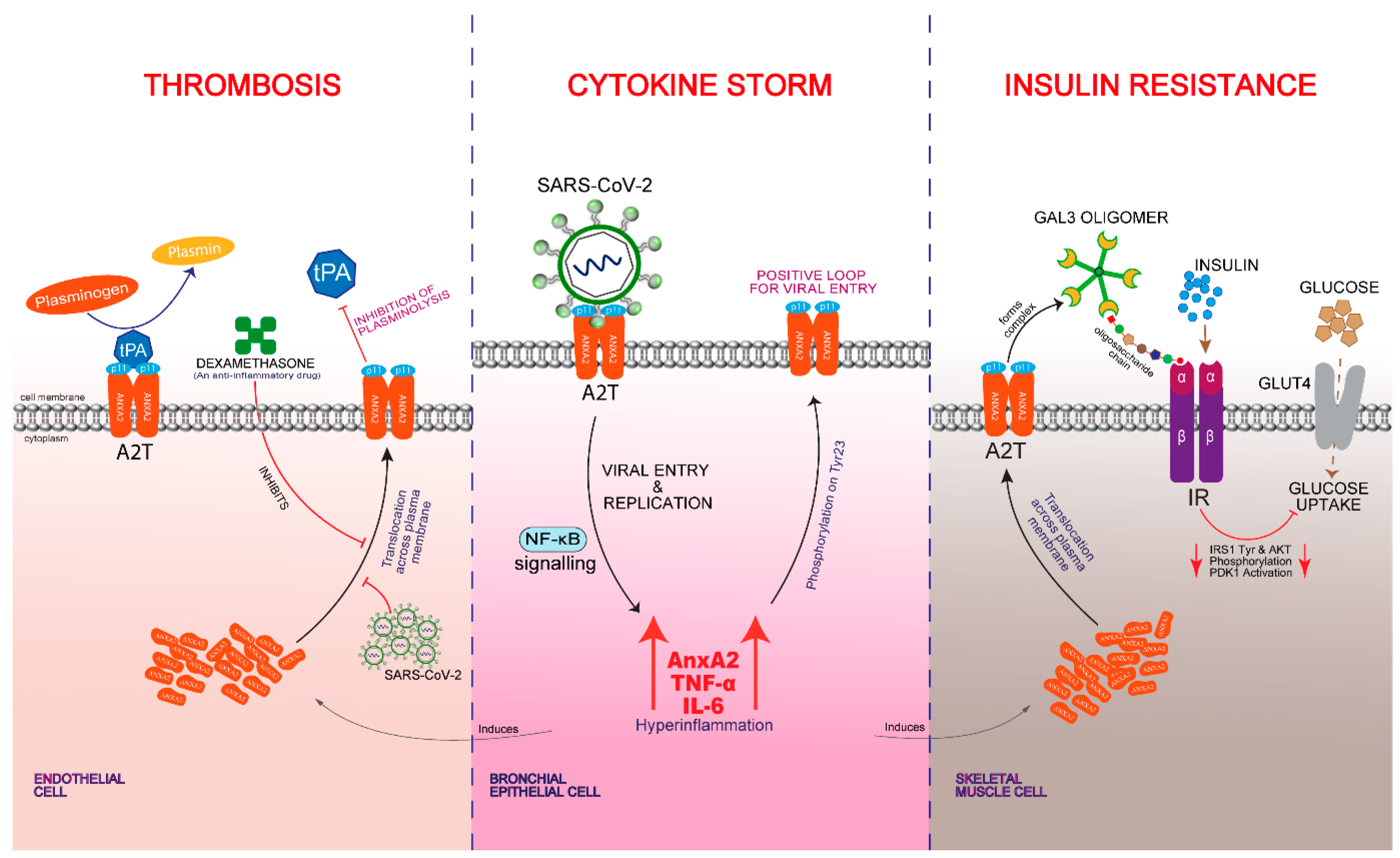
3. AnxA2 Role in Cytokine Storm
4. AnxA2 Role in Thrombosis
5. AnxA2 Role in Insulin Resistance
6. Summary and Future Perspectives
Supplementary Materials
Author Contributions
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- China CDC Weekly. The Epidemiological Characteristics of an Outbreak of 2019 Novel Coronavirus Diseases (COVID-19)—China, 2020. China CDC Weekly 2020, 2, 113–122. [Google Scholar] [CrossRef]
- Johns Hopkins Coronavirus Resource Center Home Page. Available online: https://0-coronavirus-jhu-edu.brum.beds.ac.uk/data (accessed on 5 October 2021).
- Wu, Y.; Ho, W.; Huang, Y.; Jin, D.-Y.; Li, S.; Liu, S.-L.; Liu, X.; Qiu, J.; Sang, Y.; Wang, Q.; et al. SARS-CoV-2 is an appropriate name for the new coronavirus. Lancet 2020, 395, 949–950. [Google Scholar] [CrossRef]
- Khan, S.; Siddique, R.; Shereen, M.A.; Ali, A.; Liu, J.; Bai, Q.; Bashir, N.; Xue, M. Emergence of a Novel Coronavirus, Severe Acute Respiratory Syndrome Coronavirus 2: Biology and Therapeutic Options. J. Clin. Microbiol. 2020, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, S.; Zarbock, A. Coronaviruses and SARS-CoV-2: A Brief Overview. Anesth. Analg. 2020, 131, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Yang, H.; Ji, W.; Wu, W.; Chen, S.; Zhang, W.; Duan, G. Virology, Epidemiology, Pathogenesis, and Control of COVID-19. Viruses 2020, 12, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umakanthan, S.; Chattu, V.K.; Ranade, A.V.; Das, D.; Basavarajegowda, A.; Bukelo, M. A rapid review of recent advances in diagnosis, treatment and vaccination for COVID-19. AIMS Public Health 2021, 8, 137–153. [Google Scholar] [CrossRef] [PubMed]
- Ahn, D.-G.; Shin, H.-J.; Kim, M.-H.; Lee, S.; Kim, H.-S.; Myoung, J.; Kim, B.-T.; Kim, S.-J. Current Status of Epidemiology, Diagnosis, Therapeutics, and Vaccines for Novel Coronavirus Disease 2019 (COVID-19). J. Microbiol. Biotechnol. 2020, 30, 313–324. [Google Scholar] [CrossRef]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; Peacock, S.J.; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM Structure of the 2019-NCoV Spike in the Prefusion Conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchis-Gomar, F.; Lavie, C.J.; Perez-Quilis, C.; Henry, B.M.; Lippi, G. Angiotensin-Converting Enzyme 2 and Antihyper-tensives (Angiotensin Receptor Blockers and Angiotensin-Converting Enzyme Inhibitors) in Coronavirus Disease 2019. Mayo Clin. Proc. 2020, 95, 1222–1230. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Chen, K.; Zou, J.; Han, P.; Hao, J.; Han, Z. Single-cell RNA-seq data analysis on the receptor ACE2 expression reveals the potential risk of different human organs vulnerable to 2019-nCoV infection. Front. Med. 2020, 14, 185–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyerstedt, S.; Casaro, E.B.; Rangel, É.B. COVID-19: Angiotensin-converting enzyme 2 (ACE2) expression and tissue suscep-tibility to SARS-CoV-2 infection. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 905–919. [Google Scholar] [CrossRef]
- Kragstrup, T.W.; Singh, H.S.; Grundberg, I.; Nielsen, A.L.-L.; Rivellese, F.; Mehta, A.; Goldberg, M.B.; Filbin, M.R.; Qvist, P.; Bibby, B.M. Plasma ACE2 predicts outcome of COVID-19 in hospitalized patients. PLoS ONE 2021, 16, e0252799. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Huang, Y.; Lau, S.K.P.; Yuen, K.-Y. Coronavirus Genomics and Bioinformatics Analysis. Viruses 2010, 2, 1804–1820. [Google Scholar] [CrossRef] [Green Version]
- Peiris, J.S.M.; Lai, S.T.; Poon, L.L.M.; Guan, Y.; Yam, L.Y.C.; Lim, W.; Nicholls, J.; Yee, W.K.S.; Yan, W.W.; Cheung, M.T.; et al. Coronavirus as a possible cause of severe acute respiratory syndrome. Lancet 2003, 361, 1319–1325. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, Y.; Ye, D.; Liu, Q. Review of the 2019 novel coronavirus (SARS-CoV-2) based on current evidence. Int. J. Antimicrob. Agents 2020, 55, 105948. [Google Scholar] [CrossRef]
- Zheng, J. SARS-CoV-2: An Emerging Coronavirus that Causes a Global Threat. Int. J. Biol. Sci. 2020, 16, 1678–1685. [Google Scholar] [CrossRef] [Green Version]
- Shereen, M.A.; Khan, S.; Kazmi, A.; Bashir, N.; Siddique, R. COVID-19 infection: Emergence, transmission, and characteristics of human coronaviruses. J. Adv. Res. 2020, 24, 91–98. [Google Scholar] [CrossRef]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef]
- Bojkova, D.; Klann, K.; Koch, B.; Widera, M.; Krause, D.; Ciesek, S.; Cinatl, J.; Münch, C. Proteomics of SARS-CoV-2-infected host cells reveals therapy targets. Nat. Cell Biol. 2020, 583, 469–472. [Google Scholar] [CrossRef]
- Wang, M.Y.; Zhao, R.; Gao, L.J.; Gao, X.F.; Wang, D.P.; Cao, J.M. SARS-CoV-2: Structure, Biology, and Structure-Based Ther-apeutics Development. Front. Cell. Infect. Microbiol. 2020, 10, 724. [Google Scholar] [CrossRef]
- Romano, M.; Ruggiero, A.; Squeglia, F.; Maga, G.; Berisio, R. A Structural View of SARS-CoV-2 RNA Replication Machinery: RNA Synthesis, Proofreading and Final Capping. Cells 2020, 9, 1267. [Google Scholar] [CrossRef]
- Haddad, C.; Davila-Calderon, J.; Tolbert, B.S. Integrated approaches to reveal mechanisms by which RNA viruses reprogram the cellular environment. Methods 2020, 183, 50–56. [Google Scholar] [CrossRef]
- Li, F. Structure, Function, and Evolution of Coronavirus Spike Proteins. Annu. Rev. Virol. 2016, 3, 237–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, X.; Sun, J.; Yan, Z.; Zhang, J.; Zhao, J.; Zhao, Z.; Gao, Q.; He, W.-T.; Veit, M.; Su, S. Comparison of Severe Acute Respiratory Syndrome Coronavirus 2 Spike Protein Binding to ACE2 Receptors from Human, Pets, Farm Animals, and Putative Intermediate Hosts. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, J.; Randeva, H.S.; Chatha, K.; Hall, M.; Spandidos, D.A.; Karteris, E.; Kyrou, I. Neuropilin-1 as a new potential SARS-CoV-2 infection mediator implicated in the neurologic features and central nervous system involvement of COVID-19. Mol. Med. Rep. 2020, 22, 4221–4226. [Google Scholar] [CrossRef]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; Van Der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 2020, eabd2985. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.L.; Simonetti, B.; Klein, K.; Chen, K.-E.; Williamson, M.K.; Antón-Plágaro, C.; Shoemark, D.K.; Simón-Gracia, L.; Bauer, M.; Hollandi, R.; et al. Neuropilin-1 is a host factor for SARS-CoV-2 infection. Science 2020, eabd3072. [Google Scholar] [CrossRef] [PubMed]
- Bayati, A.; Kumar, R.; Francis, V.; McPherson, P.S. SARS-CoV-2 infects cells after viral entry via clathrin-mediated endocytosis. J. Biol. Chem. 2021, 296, 100306. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Xiong, X.; Park, Y.-J.; Tortorici, M.A.; Snijder, J.; Quispe, J.; Cameroni, E.; Gopal, R.; Dai, M.; Lanzavecchia, A.; et al. Unexpected Receptor Functional Mimicry Elucidates Activation of Coronavirus Fusion. Cell 2019, 176, 1026–1039.e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, T.; Bidon, M.; Jaimes, J.A.; Whittaker, G.R.; Daniel, S. Coronavirus membrane fusion mechanism offers a potential target for antiviral development. Antivir. Res. 2020, 178, 104792. [Google Scholar] [CrossRef]
- Valapala, M.; Vishwanatha, J.K. Lipid Raft Endocytosis and Exosomal Transport Facilitate Extracellular Trafficking of Annexin A2. J. Biol. Chem. 2011, 286, 30911–30925. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Hajjar, K.A. The annexin A2 system and angiogenesis. Biol. Chem. 2016, 397, 1005–1016. [Google Scholar] [CrossRef]
- Taylor, J.R.; Skeate, J.; Kast, W.M. Annexin A2 in Virus Infection. Front. Microbiol. 2018, 9, 2954. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.-T.; Lin, C.-F.; Liao, P.-C.; Kuo, Y.-M.; Wang, S.; Yeh, T.-M.; Shieh, C.-C.K.; Su, I.-J.; Lei, H.-Y.; Lin, Y.-S. Annexin A2 on lung epithelial cell surface is recognized by severe acute respiratory syndrome-associated coronavirus spike domain 2 antibodies. Mol. Immunol. 2010, 47, 1000–1009. [Google Scholar] [CrossRef]
- Zuniga, M.; Gomes, C.; Carsons, S.E.; Bender, M.T.; Cotzia, P.; Miao, Q.R.; Lee, D.C.; Rodriguez, A. Autoimmunity to Annexin A2 predicts mortality among hospitalised COVID-19 patients. Eur. Respir. J. 2021, 2100918. [Google Scholar] [CrossRef]
- Filipenko, N.R.; MacLeod, T.J.; Yoon, C.-S.; Waisman, D. Annexin A2 Is a Novel RNA-binding Protein. J. Biol. Chem. 2004, 279, 8723–8731. [Google Scholar] [CrossRef] [Green Version]
- Bharadwaj, A.; Bydoun, M.; Holloway, R.; Waisman, D. Annexin A2 Heterotetramer: Structure and Function. Int. J. Mol. Sci. 2013, 14, 6259–6305. [Google Scholar] [CrossRef] [Green Version]
- Madureira, P.A.; Surette, A.P.; Phipps, K.D.; Taboski, M.A.S.; Miller, V.A.; Waisman, D.M. The role of the annexin A2 heterotetramer in vascular fibrinolysis. Blood 2011, 118, 4789–4797. [Google Scholar] [CrossRef] [Green Version]
- Rescher, U.; Gerke, V. Annexins—Unique membrane binding proteins with diverse functions. J. Cell Sci. 2004, 117, 2631–2639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajjar, K.A. The Biology of Annexin A2: From Vascular Fibrinolysis to Innate Immunity. Trans. Am. Clin. Clim. Assoc. 2015, 126, 144–155. [Google Scholar]
- Koyuncu, O.O.; Perlman, D.H.; Enquist, L.W. Efficient Retrograde Transport of Pseudorabies Virus within Neurons Requires Local Protein Synthesis in Axons. Cell Host Microbe 2013, 13, 54–66. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.-L.; Chou, Y.-T.; Wu, C.N.; Ho, M.-S. Annexin II Binds to Capsid Protein VP1 of Enterovirus 71 and Enhances Viral Infectivity. J. Virol. 2011, 85, 11809–11820. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Koike, S. Cellular receptors for enterovirus A71. J. Biomed. Sci. 2020, 27, 1–12. [Google Scholar] [CrossRef]
- Yen, T.-Y.; Shih, W.-L.; Huang, Y.-C.; Lee, J.-T.; Huang, L.-M.; Chang, L.-Y. Polymorphisms in enterovirus 71 receptors associated with susceptibility and clinical severity. PLoS ONE 2018, 13, e0206769. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.; Kurosky, A.; Wasi, S. An Endothelial Cell-Surface Form of Annexin II Binds Human Cytomegalovirus. Biochem. Biophys. Res. Commun. 1994, 198, 983–989. [Google Scholar] [CrossRef]
- Derry, M.C.; Sutherland, M.R.; Restall, C.M.; Waisman, D.; Pryzdial, E.L.G. Annexin 2-mediated enhancement of cytomegalovirus infection opposes inhibition by annexin 1 or annexin 5. J. Gen. Virol. 2007, 88, 19–27. [Google Scholar] [CrossRef]
- Raynor, C.M.; Wright, J.F.; Waisman, D.; Pryzdial, E.L.G. Annexin II Enhances Cytomegalovirus Binding and Fusion to Phospholipid Membranes. Biochemistry 1999, 38, 5089–5095. [Google Scholar] [CrossRef] [PubMed]
- Woodham, A.W.; Da Silva, D.M.; Skeate, J.; Raff, A.B.; Ambroso, M.R.; Brand, H.E.; Isas, J.M.; Langen, R.; Kast, W.M. The S100A10 Subunit of the Annexin A2 Heterotetramer Facilitates L2-Mediated Human Papillomavirus Infection. PLoS ONE 2012, 7, e43519. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.R.; Fernandez, D.J.; Thornton, S.M.; Skeate, J.; Lühen, K.P.; Da Silva, D.M.; Langen, R.; Kast, W.M. Heterotetrameric annexin A2/S100A10 (A2t) is essential for oncogenic human papillomavirus trafficking and capsid disassembly, and protects virions from lysosomal degradation. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Greenwell-Wild, T.; Lei, K.; Jin, W.; Swisher, J.; Hardegen, N.; Wild, C.T.; Wahl, S.M. Secretory Leukocyte Protease Inhibitor Binds to Annexin II, a Cofactor for Macrophage HIV-1 Infection. J. Exp. Med. 2004, 200, 1337–1346. [Google Scholar] [CrossRef] [Green Version]
- Rai, T.; Mosoian, A.; Resh, M.D. Annexin 2 Is Not Required for Human Immunodeficiency Virus Type 1 Particle Production but Plays a Cell Type-Dependent Role in Regulating Infectivity. J. Virol. 2010, 84, 9783–9792. [Google Scholar] [CrossRef] [Green Version]
- Aliyu, I.A.; Ling, K.-H.; Hashim, N.F.M.; Lam, J.-Y.; Chee, H.-Y. Annexin II as a Dengue Virus Serotype 2 Interacting Protein Mediating Virus Interaction on Vero Cells. Viruses 2019, 11, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhotra, R.; Ward, M.; Bright, H.; Priest, R.; Foster, M.R.; Hurle, M.; Blair, E.; Bird, M. Isolation and characterisation of potential respiratory syncytial virus receptor(s) on epithelial cells. Microbes Infect. 2003, 5, 123–133. [Google Scholar] [CrossRef]
- Janin, J.; Henrick, K.; Moult, J.; Eyck, L.T.; Sternberg, M.; Vajda, S.; Vakser, I.; Wodak, S. CAPRI: A Critical Assessment of PRedicted Interactions. Proteins: Struct. Funct. Bioinform. 2003, 52, 2–9. [Google Scholar] [CrossRef]
- Comeau, S.R.; Gatchell, D.W.; Vajda, S.; Camacho, C.J. ClusPro: A fully automated algorithm for protein-protein docking. Nucleic Acids Res. 2004, 32, W96–W99. [Google Scholar] [CrossRef] [Green Version]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S. The ClusPro web server for protein–protein docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Kozakov, D.; Beglov, D.; Bohnuud, T.; Mottarella, S.E.; Xia, B.; Hall, D.R.; Vajda, S. How good is automated protein docking? Proteins Struct. Funct. Bioinform. 2013, 81, 2159–2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrödinger, L.; De Lano, W. PyMOL; v2.4.0; Schrodinger: New York, NY, USA, 2021. [Google Scholar]
- Koga, R.; Kubota, M.; Hashiguchi, T.; Yanagi, Y.; Ohno, S. Annexin A2 Mediates the Localization of Measles Virus Matrix Protein at the Plasma Membrane. J. Virol. 2018, 92, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backes, P.; Quinkert, D.; Reiss, S.; Binder, M.; Zayas, M.; Rescher, U.; Gerke, V.; Bartenschlager, R.; Lohmann, V. Role of Annexin A2 in the Production of Infectious Hepatitis C Virus Particles. J. Virol. 2010, 84, 5775–5789. [Google Scholar] [CrossRef] [Green Version]
- Saxena, V.; Lai, C.-K.; Chao, T.-C.; Jeng, K.-S.; Lai, M.M.C. Annexin A2 Is Involved in the Formation of Hepatitis C Virus Replication Complex on the Lipid Raft. J. Virol. 2012, 86, 4139–4150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solbak, S.M.Ø.; Abdurakhmanov, E.; Vedeler, A.; Danielson, U.H. Characterization of interactions between hepatitis C virus NS5B polymerase, annexin A2 and RNA—Effects on NS5B catalysis and allosteric inhibition. Virol. J. 2017, 14, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeBouder, F.; Morello, E.; Rimmelzwaan, G.F.; Bosse, F.; Péchoux, C.; Delmas, B.; Riteau, B. Annexin II Incorporated into Influenza Virus Particles Supports Virus Replication by Converting Plasminogen into Plasmin. J. Virol. 2008, 82, 6820–6828. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Sun, J.; Gu, L.; Bao, H.; Zhao, Y.; Shi, L.; Yao, W.; Tian, G.; Wang, X.; Chen, H. Annexin A2 (ANXA2) interacts with nonstructural protein 1 and promotes the replication of highly pathogenic H5N1 avian influenza virus. BMC Microbiol. 2017, 17, 191. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Zhang, A.; Guo, J.; Yang, J.; Zhou, H.; Chen, H.; Jin, M. Identification of Human Host Proteins Contributing to H5N1 Influenza Virus Propagation by Membrane Proteomics. J. Proteome Res. 2012, 11, 5396–5405. [Google Scholar] [CrossRef]
- Ryzhova, E.V.; Vos, R.M.; Albright, A.V.; Harrist, A.V.; Harvey, T.; González-Scarano, F. Annexin 2: A Novel Human Immunodeficiency Virus Type 1 Gag Binding Protein Involved in Replication in Monocyte-Derived Macrophages. J. Virol. 2006, 80, 2694–2704. [Google Scholar] [CrossRef] [Green Version]
- Harrist, A.V.; Ryzhova, E.V.; Harvey, T.; González-Scarano, F. Anx2 Interacts with HIV-1 Gag at Phosphatidylinositol (4,5) Bisphosphate-Containing Lipid Rafts and Increases Viral Production in 293T Cells. PLoS ONE 2009, 4, e5020. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Chang, J.-S.; Song, M.-S.; Ahn, B.-Y.; Park, Y.I.; Lim, D.-S.; Han, Y.S. Association of hepatitis B virus polymerase with promyelocytic leukemia nuclear bodies mediated by the S100 family protein p11. Biochem. Biophys. Res. Commun. 2003, 305, 1049–1056. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, J.; Zhang, D.; Xu, Z.; Ji, J.; Wen, C. Cytokine Storm in COVID-19: The Current Evidence and Treatment Strategies. Front. Immunol. 2020, 11, 1708. [Google Scholar] [CrossRef]
- Ragab, D.; Eldin, S.H.; Taeimah, M.; Khattab, R.; Salem, R. The COVID-19 Cytokine Storm; What We Know So Far. Front. Immunol. 2020, 11, 1446. [Google Scholar] [CrossRef]
- Mokhtari, T.; Hassani, F.; Ghaffari, N.; Ebrahimi, B.; Yarahmadi, A.; Hassanzadeh, G. COVID-19 and multiorgan failure: A narrative review on potential mechanisms. J. Mol. Histol. 2020, 51, 613–628. [Google Scholar] [CrossRef]
- Yang, L.; Xie, X.; Tu, Z.; Fu, J.; Xu, D.; Zhou, Y. The signal pathways and treatment of cytokine storm in COVID-19. Signal Transduct. Target. Ther. 2021, 6, 1–20. [Google Scholar] [CrossRef]
- Darif, D.; Hammi, I.; Kihel, A.; El Idrissi Saik, I.; Guessous, F.; Akarid, K. The pro-inflammatory cytokines in COVID-19 pathogenesis: What goes wrong? Microb. Pathog. 2021, 153, 104799. [Google Scholar] [CrossRef]
- Swisher, J.F.A.; Khatri, U.; Feldman, G.M. Annexin A2 is a soluble mediator of macrophage activation. J. Leukoc. Biol. 2007, 82, 1174–1184. [Google Scholar] [CrossRef] [Green Version]
- Shetty, P.; Patil, V.S.; Mohan, R.; D’Souza, L.C.; Bargale, A.; Patil, B.R.; Dinesh, U.; Haridas, V.; Kulkarni, S.P. Annexin A2 and its downstream IL-6 and HB-EGF as secretory biomarkers in the differential diagnosis of Her-2 negative breast cancer. Ann. Clin. Biochem. Int. J. Lab. Med. 2016, 54, 463–471. [Google Scholar] [CrossRef]
- Jung, H.; Kim, J.M.; Kim, W.K.; Oh, K.-J.; Lee, H.J.; Han, B.S.; Kim, D.S.; Seo, Y.S.; Lee, S.C.; Park, S.G.; et al. Intracellular annexin A2 regulates NF-κB signaling by binding to the p50 subunit: Implications for gemcitabine resistance in pancreatic cancer. Cell Death Dis. 2015, 6, e1606. [Google Scholar] [CrossRef] [Green Version]
- Schuliga, M.; Langenbach, S.; Berhan, A.; Harris, T.; Keenan, C.R.; Royce, S.G.; Stewart, A.G. The Coagulant Factor Xa Induces Protease-Activated Receptor-1 and Annexin A2-Dependent Airway Smooth Muscle Cytokine Production and Cell Proliferation. Am. J. Respir. Cell Mol. Biol. 2016, 54, 200–209. [Google Scholar] [CrossRef]
- Schuliga, M.; Jaffar, J.; Berhan, A.; Langenbach, S.; Harris, T.; Waters, D.; Lee, P.V.S.; Grainge, C.; Westall, G.; Knight, D.; et al. Annexin A2 contributes to lung injury and fibrosis by augmenting factor Xa fibrogenic activity. Am. J. Physiol. Cell. Mol. Physiol. 2017, 312, L772–L782. [Google Scholar] [CrossRef] [Green Version]
- Schuliga, M.; Langenbach, S.; Xia, Y.C.; Qin, C.; Mok, J.S.; Harris, T.; Mackay, G.A.; Medcalf, R.L.; Stewart, A.G. Plasminogen-Stimulated Inflammatory Cytokine Production by Airway Smooth Muscle Cells Is Regulated by Annexin A2. Am. J. Respir. Cell Mol. Biol. 2013, 49, 751–758. [Google Scholar] [CrossRef]
- Renner, B.; Tong, H.H.; Laskowski, J.; Jonscher, K.; Goetz, L.; Woolaver, R.; Hannan, J.; Li, Y.X.; Hourcade, D.; Pickering, M.C.; et al. Annexin A2 Enhances Complement Activation by Inhibiting Factor H. J. Immunol. 2016, 196, 1355–1365. [Google Scholar] [CrossRef] [Green Version]
- Tsukamoto, H.; Tanida, S.; Ozeki, K.; Ebi, M.; Mizoshita, T.; Shimura, T.; Mori, Y.; Kataoka, H.; Kamiya, T.; Fukuda, S.; et al. Annexin A2 Regulates A Disintegrin and Metalloproteinase 17–mediated Ectodomain Shedding of Pro–Tumor Necrosis Factor-α in Monocytes and Colon Epithelial Cells. Inflamm. Bowel Dis. 2013, 19, 1365–1373. [Google Scholar] [CrossRef]
- Swisher, J.F.A.; Burton, N.; Bacot, S.M.; Vogel, S.N.; Feldman, G.M. Annexin A2 tetramer activates human and murine macrophages through TLR4. Blood 2010, 115, 549–558. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-J.; Jung, Y.H.; Song, E.J.; Jang, K.K.; Choi, S.H.; Han, H.J. Vibrio vulnificusVvpE Stimulates IL-1β Production by the Hypomethylation of the IL-1β Promoter and NF-κB Activation via Lipid Raft–Dependent ANXA2 Recruitment and Reactive Oxygen Species Signaling in Intestinal Epithelial Cells. J. Immunol. 2015, 195, 2282–2293. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-J.; Jung, Y.H.; Kim, J.S.; Lee, H.J.; Lee, S.H.; Lee, K.-H.; Jang, K.K.; Choi, S.H.; Han, H.J. A Vibrio vulnificus VvpM Induces IL-1β Production Coupled with Necrotic Macrophage Death via Distinct Spatial Targeting by ANXA2. Front. Cell. Infect. Microbiol. 2017, 7, 352. [Google Scholar] [CrossRef]
- Khamsi, R. Rogue antibodies could be driving severe COVID-19. Nat. Cell Biol. 2021, 590, 29–31. [Google Scholar] [CrossRef]
- Zaid, A.; Dawoodi, S.; Pirro, M.; Monti, M.; Mobaraki, P.D. key role of annexin a2 and plasmin in covid-19 pathophysiology, clinical presentation and outcomes. Ital. J. Prev. Diagnostic Ther. Med 2020, 3, 16–25. [Google Scholar]
- Cesarman-Maus, G.C.; Ríos-Luna, N.P.; Deora, A.B.; Huang, B.; Villa, R.; Cravioto, M.D.C.; Alarcón-Segovia, D.; Sánchez-Guerrero, J.; Hajjar, K.A. Autoantibodies against the fibrinolytic receptor, annexin 2, in antiphospholipid syndrome. Blood 2006, 107, 4375–4382. [Google Scholar] [CrossRef]
- Brichory, F.M.; Misek, D.E.; Yim, A.-M.; Krause, M.C.; Giordano, T.; Beer, D.G.; Hanash, S.M. An immune response manifested by the common occurrence of annexins I and II autoantibodies and high circulating levels of IL-6 in lung cancer. Proc. Natl. Acad. Sci. USA 2001, 98, 9824–9829. [Google Scholar] [CrossRef] [Green Version]
- Haridas, V.; Shetty, P.; Sarathkumar, E.; Bargale, A.; Vishwanatha, J.; Patil, V.; Dinesh, U.S. Reciprocal regulation of pro-inflammatory Annexin A2 and anti-inflammatory Annexin A1 in the pathogenesis of rheumatoid arthritis. Mol. Biol. Rep. 2019, 46, 83–95. [Google Scholar] [CrossRef]
- Tsai, W.-H.; Lai, S.-L.; Li, I.-T.; Chien, H.-Y.; Shih, C.-H.; Kou, Y.R.; Hsu, H.-C. Annexin A1 mediates the anti-adhesive effects of dexamethasone during the cell-cell interaction between the all-trans retinoic acid-treated acute promyelocytic leukemic cells and endothelial cells. J. Cell. Biochem. 2013, 114, 551–557. [Google Scholar] [CrossRef]
- McFadyen, J.D.; Stevens, H.; Peter, K. The Emerging Threat of (Micro)Thrombosis in COVID-19 and Its Therapeutic Implications. Circ. Res. 2020, 127, 571–587. [Google Scholar] [CrossRef]
- Klok, F.A.; Kruip, M.J.H.A.; van der Meer, N.J.M.; Arbous, M.S.; Gommers, D.; Kant, K.M.; Kaptein, F.H.J.; van Paassen, J.; Stals, M.A.M.; Huisman, M.V.; et al. Confirmation of the high cumulative incidence of thrombotic complications in critically ill ICU patients with COVID-19: An updated analysis. Thromb. Res. 2020, 191, 148–150. [Google Scholar] [CrossRef]
- Suh, Y.J.; Hong, H.; Ohana, M.; Bompard, F.; Revel, M.-P.; Valle, C.; Gervaise, A.; Poissy, J.; Susen, S.; Hékimian, G.; et al. Pulmonary Embolism and Deep Vein Thrombosis in COVID-19: A Systematic Review and Meta-Analysis. Radiology 2021, 298, E70–E80. [Google Scholar] [CrossRef]
- Middeldorp, S.; Coppens, M.; Van Haaps, T.F.; Foppen, M.; Vlaar, A.P.; Müller, M.C.A.; Bouman, C.C.S.; Beenen, L.F.M.; Kootte, R.S.; Heijmans, J.; et al. Incidence of venous thromboembolism in hospitalized patients with COVID-19. J. Thromb. Haemost. 2020, 18, 1995–2002. [Google Scholar] [CrossRef]
- Mohamed, M.F.H.; Al-Shokri, S.D.; Shunnar, K.M.; Mohamed, S.F.; Najim, M.S.; Ibrahim, S.I.; Elewa, H.; Abdalla, L.O.; El-Bardissy, A.; Elshafei, M.N.; et al. Prevalence of Venous Thromboembolism in Critically Ill COVID-19 Patients: Systematic Review and Meta-Analysis. Front. Cardiovasc. Med. 2021, 7, 598846. [Google Scholar] [CrossRef]
- Loo, J.; Spittle, D.A.; Newnham, M. COVID-19, immunothrombosis and venous thromboembolism: Biological mechanisms. Thorax 2021, 76, 412–420. [Google Scholar] [CrossRef]
- Fassel, H.; Chen, H.; Ruisi, M.; De Sancho, M.T.; Hajjar, K.A. Markedly Reduced Expression of Annexin A2 Is Associated with Hypofibrinolysis and Provoked and Unprovoked Venous Thromboembolism. Blood 2019, 134, 443. [Google Scholar] [CrossRef]
- Fassel, H.; Chen, H.; Ruisi, M.; Kumar, N.; DeSancho, M.T.; Hajjar, K.A. Reduced expression of annexin A2 is associated with impaired cell surface fibrinolysis and venous thromboembolism. Blood 2021, 137, 2221–2230. [Google Scholar] [CrossRef]
- Ma, K.; Simantov, R.; Zhang, J.-C.; Silverstein, R.; Hajjar, K.A.; McCrae, K.R. High Affinity Binding of β2-Glycoprotein I to Human Endothelial Cells Is Mediated by Annexin II. J. Biol. Chem. 2000, 275, 15541–15548. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; McCrae, K.R. Annexin A2 mediates endothelial cell activation by antiphospholipid/anti-β2 glycoprotein I antibodies. Blood 2005, 105, 1964–1969. [Google Scholar] [CrossRef] [Green Version]
- Cesarman-Maus, G.; Cantú-Brito, C.; Barinagarrementeria, F.; Villa, R.; Reyes, E.; Sanchez-Guerrero, J.; Hajjar, K.A.; Latorre, E.G. Autoantibodies Against the Fibrinolytic Receptor, Annexin A2, in Cerebral Venous Thrombosis. Stroke 2011, 42, 501–503. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, P.; Rengarajan, T.; Thangavel, J.; Nishigaki, Y.; Sakthisekaran, D.; Sethi, G.; Nishigaki, I. The Vascular Endothelium and Human Diseases. Int. J. Biol. Sci. 2013, 9, 1057–1069. [Google Scholar] [CrossRef] [Green Version]
- Romay-Penabad, Z.; Montiel-Manzano, M.G.; Shilagard, T.; Papalardo, E.; Vargas, G.; Deora, A.B.; Wang, M.; Jacovina, A.T.; Garcia-Latorre, E.; Reyes-Maldonado, E.; et al. Annexin A2 is involved in antiphospholipid antibody-mediated pathogenic effects in vitro and in vivo. Blood 2009, 114, 3074–3083. [Google Scholar] [CrossRef] [Green Version]
- Allen, K.L.; Fonseca, F.V.; Betapudi, V.; Willard, B.; Zhang, J.; McCrae, K.R. A novel pathway for human endothelial cell activation by antiphospholipid/anti-β2 glycoprotein I antibodies. Blood 2012, 119, 884–893. [Google Scholar] [CrossRef]
- Ishii, H.; Yoshida, M.; Hiraoka, M.; Hajjar, K.A.; Tanaka, A.; Yasukochi, Y.; Numano, F. Recombinant annexin II modulates impaired fibrinolytic activity in vitro and in rat carotid artery. Circ. Res. 2001, 89, 1240–1245. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Flood, E.C.; Almeida, D.; Yan, L.; Berlin, D.A.; Heerdt, P.M.; Hajjar, K.A. Annexin A2 supports pulmonary microvascular integrity by linking vascular endothelial cadherin and protein tyrosine phosphatases. J. Exp. Med. 2017, 214, 2535–2545. [Google Scholar] [CrossRef]
- Bachler, M.; Bösch, J.; Stürzel, D.P.; Hell, T.; Giebl, A.; Ströhle, M.; Klein, S.J.; Schäfer, V.; Lehner, G.F.; Joannidis, M.; et al. Impaired fibrinolysis in critically ill COVID-19 patients. Br. J. Anaesth. 2021, 126, 590–598. [Google Scholar] [CrossRef]
- Nougier, C.; Benoit, R.; Simon, M.; Desmurs-Clavel, H.; Marcotte, G.; Argaud, L.; David, J.S.; Bonnet, A.; Negrier, C.; Dargaud, Y. Hypofibrinolytic state and high thrombin generation may play a major role in SARS-COV2 associated thrombosis. J. Thromb. Haemost. 2020, 18, 2215–2219. [Google Scholar] [CrossRef] [PubMed]
- Arachchillage, D.J.; Stacey, A.; Akor, F.; Scotz, M.; Laffan, M. Thrombolysis restores perfusion in COVID-19 hypoxia. Br. J. Haematol. 2020, 190, e270–e274. [Google Scholar] [CrossRef]
- Wang, J.; Hajizadeh, N.; Moore, E.E.; McIntyre, R.C.; Moore, P.; Veress, L.A.; Yaffe, M.B.; Moore, H.B.; Barrett, C.D. Tissue plasminogen activator (tPA) treatment for COVID-19 associated acute respiratory distress syndrome (ARDS): A case series. J. Thromb. Haemost. 2020, 18, 1752–1755. [Google Scholar] [CrossRef]
- Goyal, A.; Saigal, S.; Niwariya, Y.; Sharma, J.; Singh, P. Successful use of tPA for thrombolysis in COVID related ARDS: A case series. J. Thromb. Thrombolysis 2021, 51, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Sarfraz, A.; Sarfraz, Z.; Razzack, A.A.; Patel, G.; Sarfraz, M. Venous Thromboembolism, Corticosteroids and COVID-19: A Systematic Review and Meta-Analysis. Clin. Appl. Thromb. 2021, 27, 27. [Google Scholar] [CrossRef]
- Li, X.; Xu, S.; Yu, M.; Wang, K.; Tao, Y.; Zhou, Y.; Shi, J.; Zhou, M.; Wu, B.; Yang, Z.; et al. Risk factors for severity and mortality in adult COVID-19 inpatients in Wuhan. J. Allergy Clin. Immunol. 2020, 146, 110–118. [Google Scholar] [CrossRef]
- Lim, S.; Bae, J.H.; Kwon, H.-S.; Nauck, M.A. COVID-19 and diabetes mellitus: From pathophysiology to clinical management. Nat. Rev. Endocrinol. 2021, 17, 11–30. [Google Scholar] [CrossRef]
- Carrasco-Sánchez, F.J.; López-Carmona, M.D.; Martínez-Marcos, F.J.; Pérez-Belmonte, L.M.; Hidalgo-Jiménez, A.; Buonaiuto, V.; Fernández, C.S.; Castro, S.J.F.; Luordo, D.; Fontan, P.M.P.; et al. Admission hyperglycaemia as a predictor of mortality in patients hospitalized with COVID-19 regardless of diabetes status: Data from the Spanish SEMI-COVID-19 Registry. Ann. Med. 2021, 53, 103–116. [Google Scholar] [CrossRef]
- Zhu, L.; She, Z.-G.; Cheng, X.; Qin, J.-J.; Zhang, X.-J.; Cai, J.; Lei, F.; Wang, H.; Xie, J.; Wang, W.; et al. Association of Blood Glucose Control and Outcomes in Patients with COVID-19 and Pre-existing Type 2 Diabetes. Cell Metab. 2020, 31, 1068–1077.e3. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kong, W.; Xia, P.; Xu, Y.; Li, L.; Li, Q.; Yang, L.; Wei, Q.; Wang, H.; Li, H.; et al. Impaired Fasting Glucose and Diabetes Are Related to Higher Risks of Complications and Mortality Among Patients With Coronavirus Disease 2019. Front. Endocrinol. 2020, 11, 525. [Google Scholar] [CrossRef]
- Zhang, W.; Li, C.; Xu, Y.; He, B.; Hu, M.; Cao, G.; Li, L.; Wu, S.; Wang, X.; Zhang, C.; et al. Hyperglycemia and Correlated High Levels of Inflammation Have a Positive Relationship with the Severity of Coronavirus Disease 2019. Mediat. Inflamm. 2021, 2021, 1–9. [Google Scholar] [CrossRef]
- Wu, C.-T.; Lidsky, P.V.; Xiao, Y.; Lee, I.T.; Cheng, R.; Nakayama, T.; Jiang, S.; Demeter, J.; Bevacqua, R.J.; Chang, C.A.; et al. SARS-CoV-2 infects human pancreatic β cells and elicits β cell impairment. Cell Metab. 2021, 33, 1565–1576.e5. [Google Scholar] [CrossRef]
- Ilias, I.; Diamantopoulos, A.; Pratikaki, M.; Botoula, E.; Jahaj, E.; Athanasiou, N.; Tsipilis, S.; Zacharis, A.; Vassiliou, A.; Vassiliadi, D.; et al. Glycemia, Beta-Cell Function and Sensitivity to Insulin in Mildly to Critically Ill Covid-19 Patients. Medicina 2021, 57, 68. [Google Scholar] [CrossRef]
- Steenblock, C.; Richter, S.; Berger, I.; Barovic, M.; Schmid, J.; Schubert, U.; Jarzebska, N.; von Mässenhausen, A.; Linkermann, A.; Schürmann, A.; et al. Viral infiltration of pancreatic islets in patients with COVID-19. Nat. Commun. 2021, 12, 1–12. [Google Scholar] [CrossRef]
- Dai, H.; Yu, Z.; Fan, X.; Liu, N.; Yan, M.; Chen, Z.; Lo, E.H.; Hajjar, K.A.; Dai, H.; Wang, X. Dysfunction of annexin A2 contributes to hyperglycaemia-induced loss of human endothelial cell surface fibrinolytic activity. Thromb. Haemost. 2013, 109, 1070–1078. [Google Scholar] [CrossRef] [Green Version]
- Ishii, H.; Yoshida, M.; Hajjar, K.A.; Tanaka, A.; Yasukochi, Y.; Shimokado, K.; Numano, F. Potential role of recombinant annexin II in diabetic vascular injury. Ann. N. Y. Acad. Sci. 2006, 947, 308–311. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, Y.-S.; Yin, X.-Q.; Yu, G.; Jia, B.-L. Anxa2 gene silencing attenuates obesity-induced insulin resistance by suppressing the NF-κB signaling pathway. Am. J. Physiol. Physiol. 2019, 316, C223–C234. [Google Scholar] [CrossRef]
- Li, P.; Liu, S.; Lu, M.; Bandyopadhyay, G.; Oh, D.; Imamura, T.; Johnson, A.M.F.; Sears, D.; Shen, Z.; Cui, B.; et al. Hematopoietic-Derived Galectin-3 Causes Cellular and Systemic Insulin Resistance. Cell 2016, 167, 973–984.e12. [Google Scholar] [CrossRef] [Green Version]
- Alves, M.T.; De Souza, I.D.P.; Ferreira, C.N.; Cândido, A.L.; Bizzi, M.F.; Oliveira, F.R.; Reis, F.M.; Gomes, K.B. Galectin-3 is a potential biomarker to insulin resistance and obesity in women with polycystic ovary syndrome. Gynecol. Endocrinol. 2020, 36, 760–763. [Google Scholar] [CrossRef]
- Kingwell, K. Turning down galectin 3 to combat insulin resistance. Nat. Rev. Drug Discov. 2017, 16, 18. [Google Scholar] [CrossRef]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Alblihed, M.; Guerreiro, S.G.; Cruz-Martins, N.; Batiha, G.E.-S. COVID-19 in Relation to Hyperglycemia and Diabetes Mellitus. Front. Cardiovasc. Med. 2021, 8, 644095. [Google Scholar] [CrossRef]
- Santos, A.; Magro, D.O.; Evangelista-Poderoso, R.; Saad, M.J.A. Diabetes, obesity, and insulin resistance in COVID-19: Mo-lecular interrelationship and therapeutic implications. Diabetol. Metab. Syndr. 2021, 13, 1–14. [Google Scholar] [CrossRef]
- Shetty, P.; Bargale, A.; Patil, B.R.; Mohan, R.; Dinesh, U.S.; Vishwanatha, J.; Gai, P.B.; Patil, V.S.; Amsavardani, T.S. Cell surface interaction of annexin A2 and galectin-3 modulates epidermal growth factor receptor signaling in Her-2 negative breast cancer cells. Mol. Cell. Biochem. 2015, 411, 221–233. [Google Scholar] [CrossRef]
- Storgaard, H.; Song, X.M.; Jensen, C.B.; Madsbad, S.; Björnholm, M.; Vaag, A.A.; Zierath, J. Insulin Signal Transduction in Skeletal Muscle From Glucose-Intolerant Relatives With Type 2 Diabetes. Diabetes 2001, 50, 2770–2778. [Google Scholar] [CrossRef] [Green Version]
- Shetty, P.K.; Thamake, S.I.; Biswas, S.; Johansson, S.L.; Vishwanatha, J.K. Reciprocal Regulation of Annexin A2 and EGFR with Her-2 in Her-2 Negative and Herceptin-Resistant Breast Cancer. PLoS ONE 2012, 7, e44299. [Google Scholar] [CrossRef]
- Yuan, S.; Chan, C.C.-Y.; Chik, K.K.-H.; Tsang, J.O.-L.; Liang, R.; Cao, J.; Tang, K.; Cai, J.-P.; Ye, Z.-W.; Yin, F.; et al. Broad-Spectrum Host-Based Antivirals Targeting the Interferon and Lipogenesis Pathways as Potential Treatment Options for the Pandemic Coronavirus Disease 2019 (COVID-19). Viruses 2020, 12, 628. [Google Scholar] [CrossRef]
- Waterer, G.W.; Rello, J. Steroids and COVID-19: We Need a Precision Approach, Not One Size Fits All. Infect. Dis. Ther. 2020, 9, 701–705. [Google Scholar] [CrossRef]
- Akhtar, S.; Das, J.K.; Ismail, T.; Wahid, M.; Saeed, W.; Bhutta, Z.A. Nutritional perspectives for the prevention and mitigation of COVID-19. Nutr. Rev. 2020, 79, 289–300. [Google Scholar] [CrossRef] [PubMed]
- National Institute for Health (NIH) Dietary Supplements in the Time of COVID-19: Fact Sheet for Health Professionals. Available online: https://ods.od.nih.gov/factsheets/COVID19-HealthProfessional/ (accessed on 23 September 2021).
- Nithin, K.K.; Patil, P.; Bhandary, S.K.; Haridas, V.; Suchetha, K.N.; Sarathkumar, E.; Shetty, P. Is butyrate a natural alternative to dexamethasone in the management of CoVID-19? F1000Research 2021, 10, 273. [Google Scholar] [CrossRef]
- Funk, C.D.; Laferrière, C.; Ardakani, A. A Snapshot of the Global Race for Vaccines Targeting SARS-CoV-2 and the COVID-19 Pandemic. Front. Pharmacol. 2020, 11, 937. [Google Scholar] [CrossRef] [PubMed]
- Peiffer-Smadja, N.; Rozencwajg, S.; Kherabi, Y.; Yazdanpanah, Y.; Montravers, P. COVID-19 vaccines: A race against time. Anaesth. Crit. Care Pain Med. 2021, 40, 100848. [Google Scholar] [CrossRef]
- Forni, G.; Mantovani, A.; Forni, G.; Mantovani, A.; Moretta, L.; Rappuoli, R.; Rezza, G.; Bagnasco, A.; Barsacchi, G.; Bussolati, G.; et al. COVID-19 vaccines: Where we stand and challenges ahead. Cell Death Differ. 2021, 28, 626–639. [Google Scholar] [CrossRef] [PubMed]
- Shah, B.; Modi, P.; Sagar, S.R. In silico studies on therapeutic agents for COVID-19: Drug repurposing approach. Life Sci. 2020, 252, 117652. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Jaffee, E.M. Annexin A2 is a new antigenic target for pancreatic cancer immunotherapy. OncoImmunology 2012, 1, 112–114. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Yu, L.; Hu, B.; Chen, L.; Jv, M.; Wang, L.; Zhou, C.; Wei, M.; Zhao, L. Advances in cancer treatment: A new therapeutic target, Annexin A2. J. Cancer 2021, 12, 3587–3596. [Google Scholar] [CrossRef] [PubMed]
- Raddum, A.M.; Hollås, H.; Shumilin, I.A.; Henklein, P.; Kretsinger, R.; Fossen, T.; Vedeler, A. The native structure of annexin A2 peptides in hydrophilic environment determines their anti-angiogenic effects. Biochem. Pharmacol. 2015, 95, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Virus | Family & Genus | Genome | AnxA2 Function | Ref. | ||
|---|---|---|---|---|---|---|
| Strand | Type | Size | ||||
| PRV Pseudorabies virus | Herpesviridae Varicellovirus | Double, Linear positive | DNA, envelope | 143 Kb | Involved in retrograde trafficking of PRV in axons | [48] |
| CMV Cytomegalovirus | Herpesviridae Cytomegalovirus | Double, Circular | DNA, envelope | 200 Kb | Bridge virus to host cell membrane and enhances infection | [52,53,54] |
| HPV Human Papillomavirus | Papillomaviridae Alphapapillomavirus | Double, Circular | DNA, non-envelope | 7.9 Kb | Interact with viral L2 capsid protein for internalization and infection | [55,56] |
| EV71 Enterovirus type 71 | Picornaviridae Enterovirus | Single, Linear positive | RNA, non-envelope | 7.5 Kb | Interact with viral VP1 for host cell entry | [49,50,51] |
| DENV Dengue virus | Flaviviridae Flavivirus | Single, Linear positive | RNA, envelope | 10.7 Kb | Requires active filopodia formation for a successful DENV infection | [59] |
| RSV Respiratory syncytial virus | Pneumoviridae Orthopneumovirus | Single, Linear negative | RNA, envelope | 15 Kb | Potential epithelial cell receptor for RSV infection | [60] |
| HIV Human Immunodeficiency Virus | Retroviridae Lentivirus | Single, Linear positive | RNA, envelope | 9.2 Kb | Regulates HIV1 infectivity in cell dependent manner | [57,58] |
| SARS-CoV Severe acute respiratory syndrome coronavirus | Coronaviridae Betacoronavirus | Single, Linear positive | RNA, envelope | 30 Kb | Colocalised with S2 protein of SARS-CoV, role in internalization | [41] |
| Virus | Family & Genus | Genome | AnxA2 Function | Ref | ||
|---|---|---|---|---|---|---|
| Strand | Type | Size | ||||
| MV Measles Virus | Paramyxoviridae Morbillivirus | Single, Linear negative | RNA, envelope | 16Kb | Interacts with M protein of the virus to mediates the cellular localization | [66] |
| HCV Hepatitis C virus | Flaviviridae Hepacivirus | Single, Linear positive | RNA, envelope | 9.6Kb | Interact with NS5A/B protein of virus involved in assembly and replication | [67,68,69] |
| IAV Influenza A viruses | Orthomyxoviridae Alphainfluenzavirus | Single, Linear negative | RNA, envelope | 13.6Kb | Activation of plasminogen helps the virus to replicate | [70,71,72] |
| HBV Hepatitis B Virus | Hepadnaviridae Hepadnavirus | Double, Linear negative | DNA, envelope | 3.2Kb | In modulating HBV Pol function for intracellular viral replication | [75] |
| HIV Human Immunodeficiency Virus | Retroviridae Lentivirus | Single, Linear positive | RNA, envelope | 9.2Kb | Interact with HIV-1 Gag binding protein for replication in monocyte-derived macrophages | [73,74] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patil, P.; Shetty, P.; Kuriakose, N.; Gollapalli, P.; Shetty, S.; Bhandary, R.; Vishwanatha, J.K.; Ghate, S.D. Molecular Insights on the Possible Role of Annexin A2 in COVID-19 Pathogenesis and Post-Infection Complications. Int. J. Mol. Sci. 2021, 22, 11028. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011028
Patil P, Shetty P, Kuriakose N, Gollapalli P, Shetty S, Bhandary R, Vishwanatha JK, Ghate SD. Molecular Insights on the Possible Role of Annexin A2 in COVID-19 Pathogenesis and Post-Infection Complications. International Journal of Molecular Sciences. 2021; 22(20):11028. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011028
Chicago/Turabian StylePatil, Prakash, Praveenkumar Shetty, Nithin Kuriakose, Pavan Gollapalli, Sukanya Shetty, Roopa Bhandary, Jamboor K. Vishwanatha, and Sudeep D. Ghate. 2021. "Molecular Insights on the Possible Role of Annexin A2 in COVID-19 Pathogenesis and Post-Infection Complications" International Journal of Molecular Sciences 22, no. 20: 11028. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011028