
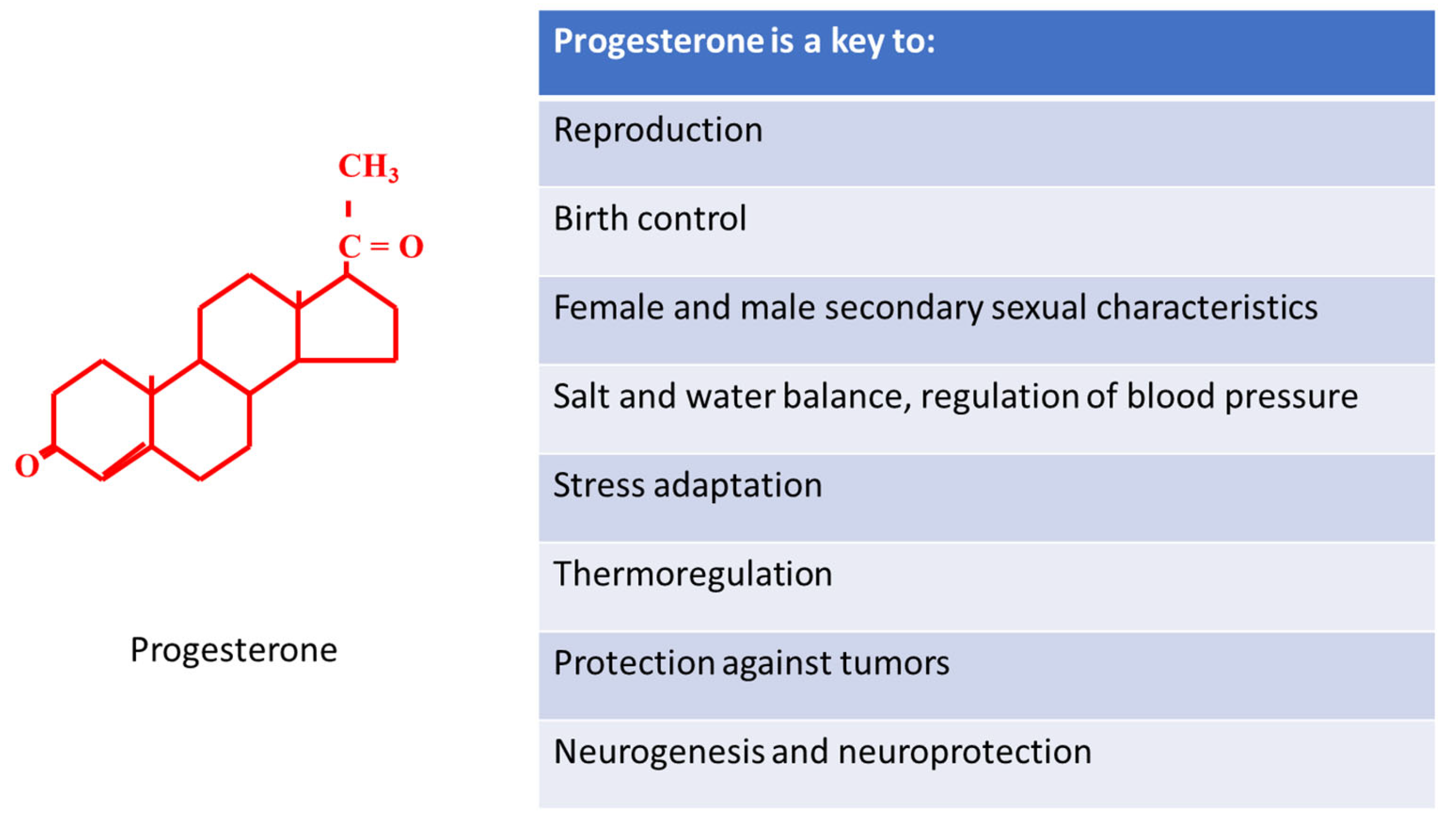
Key to Life: Physiological Role and Clinical Implications of Progesterone

 ,
, 
Abstract
:1. Introduction
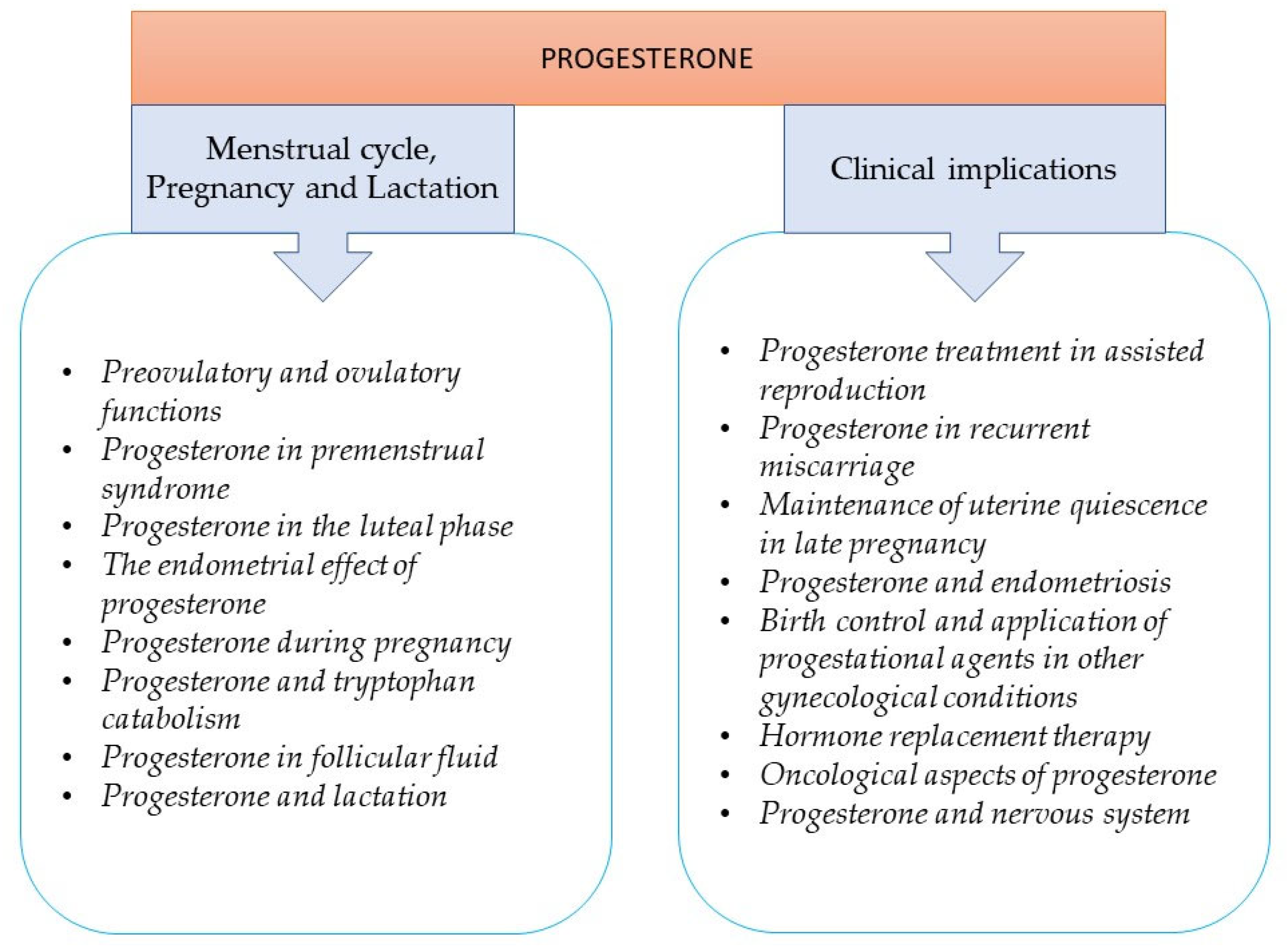
2. The Role of Progesterone in the Menstrual Cycle, Pregnancy, and Lactation
2.1. Preovulatory and Ovulatory Functions
2.2. Progesterone in Premenstrual Syndrome
2.3. Progesterone in the Luteal Phase
2.4. The Endometrial Effect of Progesterone
2.5. Progesterone during Pregnancy
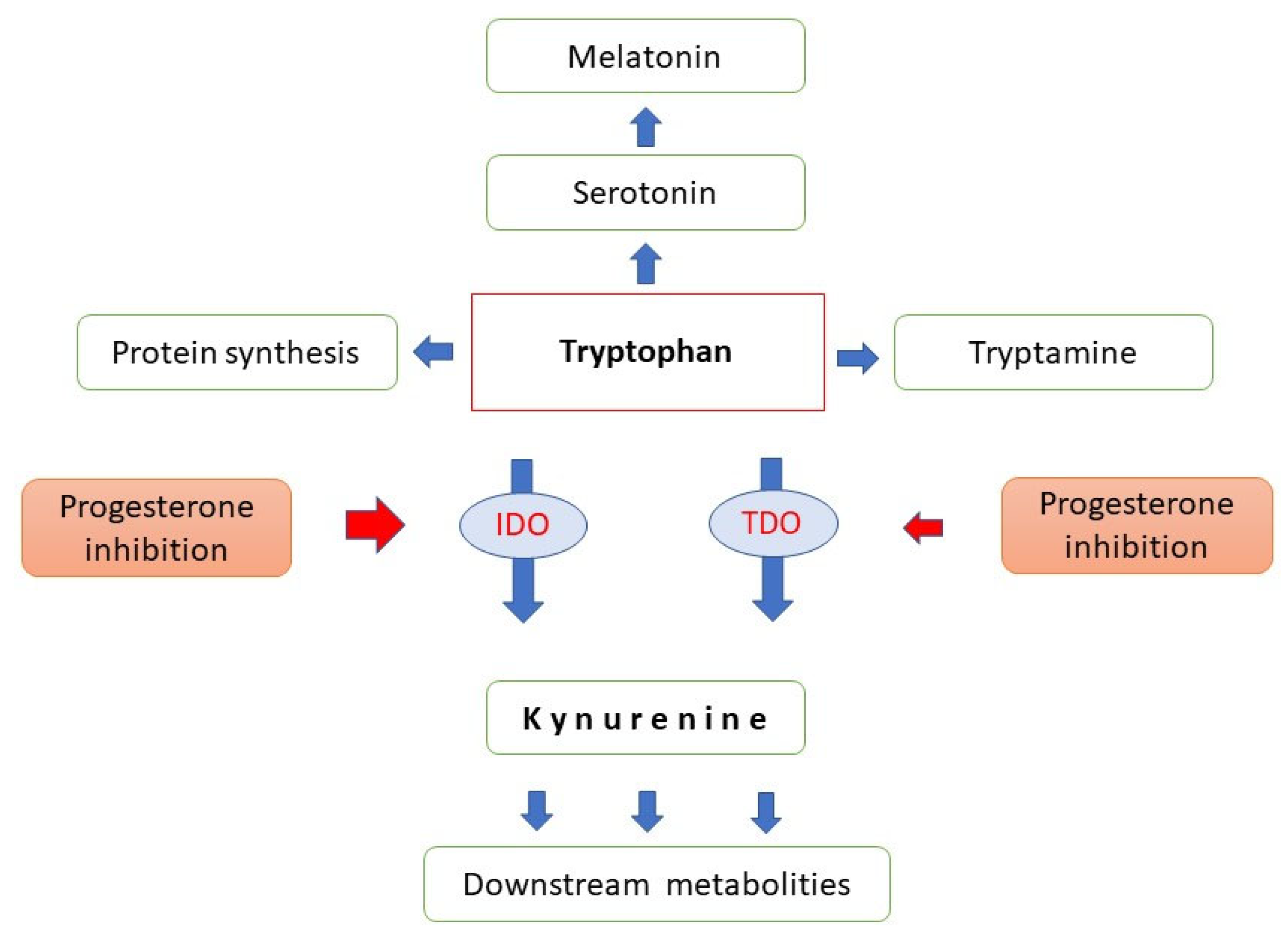
2.6. Progesterone and Tryptophan Catabolism
2.7. Progesterone in Follicular Fluid
2.8. Progesterone and Lactation
3. Clinical Implications of Progesterone
3.1. Applications of Progesterone in Reproductive Medicine
3.1.1. Progesterone Treatment in Assisted Reproduction
3.1.2. Progesterone in Recurrent Miscarriage
3.1.3. Maintenance of Uterine Quiescence in Late Pregnancy
3.1.4. Progesterone and Endometriosis
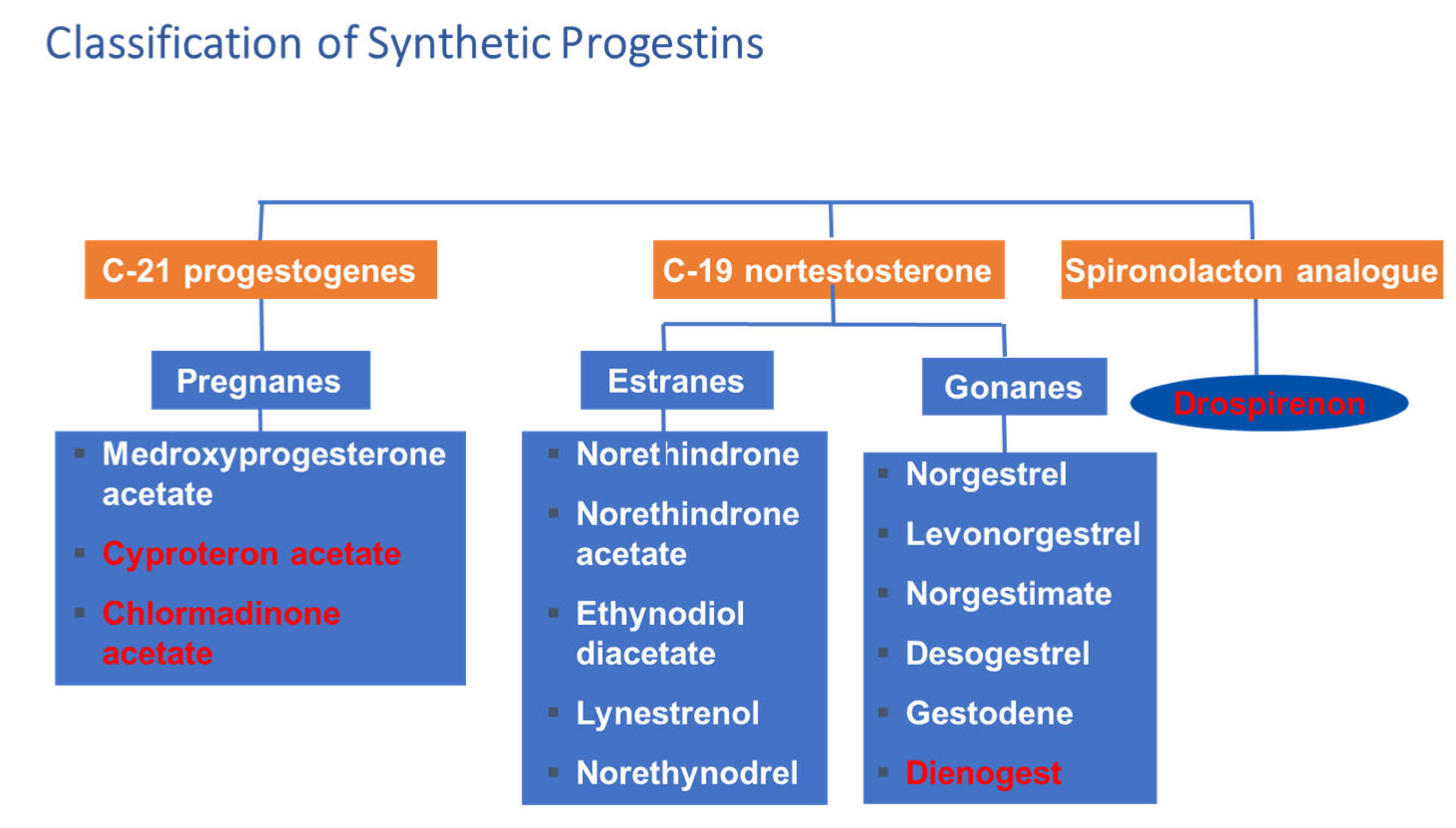
3.2. Birth Control
Application of Progestational Agents in Other Gynecological Conditions
3.3. Hormone Replacement Therapy
3.4. Oncological Aspects of Progesterone
3.5. Progesterone and Nervous System
4. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allen, W.M.; Corner, G.W. Physiology of the corpus luteum: III. Normal Growth and Implantation of Embryos After Very Early Ablation of the Ovaries, Under the Influence of Extracts of the Corpus Luteum. Am. J. Physiol.-Leg. Content 1929, 88, 340–346. [Google Scholar] [CrossRef]
- Allen, E.; Doisy, E.A. An ovarian hormone: Preliminary report on its localization, extraction and partial purification, and action in test animals. J. Am. Med. Assoc. 1923, 81, 819–821. [Google Scholar] [CrossRef]
- Taraborrelli, S. Physiology, production and action of progesterone. Acta Obstet. Gynecol. Scand. 2015, 94, 8–16. [Google Scholar] [CrossRef]
- Bodis, J.; Torok, A.; Tinneberg, H.R.; Hanf, V.; Hamori, M.; Cledon, P. Influence of serotonin on progesterone and estradiol secretion of cultured human granulosa cells. Fertil. Steril. 1992, 57, 1008–1011. [Google Scholar] [CrossRef]
- Bodis, J.; Koppan, M.; Kornya, L.; Tinneberg, H.R.; Torok, A. The effect of catecholamines, acetylcholine and histamine on progesterone release by human granulosa cells in a granulosa cell superfusion system. Gynecol. Endocrinol. 2002, 16, 259–264. [Google Scholar] [CrossRef]
- Bodis, J.; Tinneberg, H.R.; Torok, A.; Cledon, P.; Hanf, V.; Papenfuss, F. Effect of noradrenaline and dopamine on progesterone and estradiol secretion of human granulosa cells. Acta Endocrinol. 1993, 129, 165–168. [Google Scholar] [CrossRef]
- Bodis, J.; Tinneberg, H.R.; Papenfuss, F.; Török, A.; Cledon, P.H.; Hanf, V.; Schwarz, H. Cholinergic stimulation of progesterone and estradiol secretion by human granulosa cells cultured in serum-free medium. Gynecol. Endocrinol. 1993, 7, 83–87. [Google Scholar] [CrossRef]
- Bodis, J.; Torok, A.; Tinneberg, H.R.; Hanf, V.; Papenfuss, F.; Schwarz, H. Serotonin induces progesterone release from human granulosa cells in a superfused granulosa cell system. Arch. Gynecol. Obstet. 1993, 253, 59–64. [Google Scholar] [CrossRef]
- Hamori, M.; Torok, A.; Zwirner, M.; Batteux, C.; Schinkmann, W.; Bodis, J. In-vitro progesterone production of human granulosa—Luteal cells: The impact of different stimulation protocols, poor ovarian response and polycystic ovarian syndrome. Hum. Reprod. 1992, 7, 592–596. [Google Scholar] [CrossRef]
- Bodis, J.; Hanf, V.; Torok, A.; Tinneberg, H.R.; Borsay, P.; Szabo, I. Influence of nicotine on progesterone and estradiol production of cultured human granulosa cells. Early Pregnancy 1997, 3, 34–37. [Google Scholar]
- Peluso, J.J. Progesterone receptor membrane component 1 and its role in ovarian follicle growth. Front. Neurosci. 2013, 7, 99. [Google Scholar] [CrossRef] [Green Version]
- Mesen, T.B.; Young, S.L. Progesterone and the luteal phase: A requisite to reproduction. Obstet. Gynecol. Clin. N. Am. 2015, 42, 135–151. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, H.; Ueda, M.; Hattori, N.; Mori, T.; Maeda, M. A differentiation antigen of human large luteal cells in corpora lutea of the menstrual cycle and early pregnancy. Biol. Reprod. 1996, 54, 1173–1183. [Google Scholar] [CrossRef]
- Walsh, S.; Ismaili, E.; Naheed, B.; O’Brien, S. Diagnosis, pathophysiology and management of premenstrual syndrome. Obstet. Gynecol. 2015, 17, 99–104. [Google Scholar] [CrossRef]
- Ford, O.; Lethaby, A.; Roberts, H.; Mol, B.W. Progesterone for premenstrual syndrome. Cochrane Database Syst. Rev. 2012, CD003415. [Google Scholar] [CrossRef]
- De Bie, J.; Lim, C.K.; Guillemin, G.J. Progesterone Alters Kynurenine Pathway Activation in IFN-gamma-Activated Macrophages—Relevance for Neuroinflammatory Diseases. Int. J. Tryptophan. Res. 2016, 9, 89–93. [Google Scholar] [CrossRef] [Green Version]
- Songtachalert, T.; Roomruangwong, C.; Carvalho, A.F.; Bourin, M.; Maes, M. Anxiety Disorders: Sex Differences in Serotonin and Tryptophan Metabolism. Curr. Top. Med. Chem. 2018, 18, 1704–1715. [Google Scholar] [CrossRef]
- Carroll, B.J.; Steiner, M. The psychobiology of premenstrual dysphoria: The role of prolactin. Psychoneuroendocrinology 1978, 3, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.; Archibong, A.E. Ansah, T.A.; Nayyar, T. Correlation between beta-arrestin 1 and prolactin levels in women with depression during premenstrual dysphoric disorders. FASEB J. 2018, 31, 1000.7. [Google Scholar]
- Filicori, M.; Santoro, N.; Merriam, G.R.; Crowley, W.F., Jr. Characterization of the physiological pattern of episodic gonadotropin secretion throughout the human menstrual cycle. J. Clin. Endocrinol. Metab. 1986, 62, 1136–1144. [Google Scholar] [CrossRef]
- Roomruangwong, C.; Carvalho, A.F.; Comhaire, F.; Maes, M. Lowered Plasma Steady-State Levels of Progesterone Combined With Declining Progesterone Levels During the Luteal Phase Predict Peri-Menstrual Syndrome and Its Major Subdomains. Front. Psychol. 2019, 10, 2446. [Google Scholar] [CrossRef] [PubMed]
- Khan-Dawood, F.S.; Goldsmith, L.T.; Weiss, G.; Dawood, M.Y. Human corpus luteum secretion of relaxin, oxytocin, and progesterone. J. Clin. Endocrinol. Metab. 1989, 68, 627–631. [Google Scholar] [CrossRef] [PubMed]
- Lessey, B.A.; Killam, A.P.; Metzger, D.A.; Haney, A.F.; Greene, G.L.; McCarty, K.S., Jr. Immunohistochemical analysis of human uterine estrogen and progesterone receptors throughout the menstrual cycle. J. Clin. Endocrinol. Metab. 1988, 67, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Labarta, E.; Sebastian-Leon, P.; Devesa-Peiro, A.; Celada, P.; Vidal, C.; Giles, J.; Rodriguez-Varela, C.; Bosch, E.; Diaz-Gimeno, P. Analysis of serum and endometrial progesterone in determining endometrial receptivity. Hum. Reprod. 2021, deab184. [Google Scholar] [CrossRef]
- Nara, B.S.; Darmadja, D.; First, N.L. Effect of removal of follicles, corpora lutea or ovaries on maintenance of pregnancy in swine. J. Anim. Sci. 1981, 52, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Maria, B.; Stampf, F.; Goepp, A.; Ulmann, A. Termination of early pregnancy by a single dose of mifepristone (RU 486), a progesterone antagonist. Eur. J. Obstet. Gynecol. Reprod. Biol. 1988, 28, 249–255. [Google Scholar] [CrossRef]
- Halasz, M.; Szekeres-Bartho, J. The role of progesterone in implantation and trophoblast invasion. J. Reprod. Immunol. 2013, 97, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arck, P.; Hansen, P.J.; Mulac Jericevic, B.; Piccinni, M.P.; Szekeres-Bartho, J. Progesterone during pregnancy: Endocrine-immune cross talk in mammalian species and the role of stress. Am. J. Reprod. Immunol. 2007, 58, 268–279. [Google Scholar] [CrossRef]
- Szekeres-Bartho, J. Immonusupression by Progesterone in Pregnancy; CRC Press Inc.: Boca Raton, FL, USA, 1992; ISBN 0-8493-5396-3. [Google Scholar]
- Polgar, B.; Kispal, G.; Lachmann, M.; Paar, G.; Nagy, E.; Csere, P.; Miko, E.; Szereday, L.; Varga, P.; Szekeres-Bartho, J. Molecular cloning and immunologic characterization of a novel cDNA coding for progesterone-induced blocking factor. J. Immunol. 2003, 171, 5956–5963. [Google Scholar] [CrossRef] [Green Version]
- Munn, D.H.; Zhou, M.; Attwood, J.T.; Bondarev, I.; Conway, S.J.; Marshall, B.; Brown, C.; Mellor, A.L. Prevention of allogeneic fetal rejection by tryptophan catabolism. Science 1998, 281, 1191–1193. [Google Scholar] [CrossRef]
- Badawy, A.A. Tryptophan metabolism, disposition and utilization in pregnancy. Biosci. Rep. 2015, 35, e00261. [Google Scholar] [CrossRef] [PubMed]
- Bódis, J.; Sulyok, E.; Koppán, M.; Prémusz, V.; Gödöny, K.; Rascher, W.; Rauh, M. Tryptophan catabolism to serotonin and kynurenine in women undergoing in-vitro fertilization. Physiol. Res. 2020, 69, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Bomsel-Helmreich, O.; Gougeon, A.; Thebault, A.; Saltarelli, D.; Milgrom, E.; Frydman, R.; Papiernik, E. Healthy and atretic human follicles in the preovulatory phase: Differences in evolution of follicular morphology and steroid content of follicular fluid. J. Clin. Endocrinol. Metab. 1979, 48, 686–694. [Google Scholar] [CrossRef]
- Edwards, R.G.; Steptoe, P.C.; Fowler, R.E.; Baillie, J. Observations on preovulatory human ovarian follicles and their aspirates. Br. J. Obstet. Gynaecol. 1980, 87, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Oda, T.; Yoshimura, Y.; Takehara, Y.; Natori, M.; Nozawa, S. Androstenedione and progesterone concentrations in preovulatory follicular fluid correlate with successful fertilization and cleavage of human oocytes in vitro. Fertil. Steril. 1991, 56, 301–305. [Google Scholar] [CrossRef]
- Nagy, B.; Poto, L.; Farkas, N.; Koppan, M.; Varnagy, A.; Kovacs, K.; Papp, S.; Bohonyi, N.; Bodis, J. Follicular fluid progesterone concentration is associated with fertilization outcome after IVF: A systematic review and meta-analysis. Reprod. Biomed. Online 2019, 38, 871–882. [Google Scholar] [CrossRef]
- Salehnia, M.; Zavareh, S. The effects of progesterone on oocyte maturation and embryo development. Int. J. Fertil. Steril. 2013, 7, 74–81. [Google Scholar]
- Wen, X.; Li, D.; Tozer, A.J.; Docherty, S.M.; Iles, R.K. Estradiol, progesterone, testosterone profiles in human follicular fluid and cultured granulosa cells from luteinized pre-ovulatory follicles. Reprod. Biol. Endocrinol. 2010, 8, 117. [Google Scholar] [CrossRef] [Green Version]
- Tsafriri, A.; Motola, S. Are steroids dispensable for meiotic resumption in mammals? Trends Endocrinol. Metab. 2007, 18, 321–327. [Google Scholar] [CrossRef]
- Barrera, D.; Avila, E.; Diaz, L. Immunological role of progesterone in the maintenance of pregnancy. Rev. Investig. Clin. 2007, 59, 139–145. [Google Scholar]
- Loutradis, D.; Beretsos, P.; Arabatzi, E.; Anagnostou, E.; Drakakis, P. The role of steroid hormones in ART. J. Steroid Biochem. Mol. Biol. 2008, 112, 1–4. [Google Scholar] [CrossRef]
- Sengupta, J.; Ghosh, D. Role of progesterone on peri-implantation stage endometrium-embryo interaction in the primate. Steroids 2000, 65, 753–762. [Google Scholar] [CrossRef]
- Dumesic, D.A.; Meldrum, D.R.; Katz-Jaffe, M.G.; Krisher, R.L.; Schoolcraft, W.B. Oocyte environment: Follicular fluid and cumulus cells are critical for oocyte health. Fertil. Steril. 2015, 103, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Basuino, L.; Silveira, C.F., Jr. Human follicular fluid and effects on reproduction. JBRA Assist. Reprod. 2016, 20, 38–40. [Google Scholar] [CrossRef] [PubMed]
- Revelli, A.; Delle Piane, L.; Casano, S.; Molinari, E.; Massobrio, M.; Rinaudo, P. Follicular fluid content and oocyte quality: From single biochemical markers to metabolomics. Reprod. Biol. Endocrinol. 2009, 7, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conley, A.J.; Bird, I.M. The role of cytochrome P450 17 alpha-hydroxylase and 3 beta-hydroxysteroid dehydrogenase in the integration of gonadal and adrenal steroidogenesis via the delta 5 and delta 4 pathways of steroidogenesis in mammals. Biol. Reprod. 1997, 56, 789–799. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, J.; Yanaihara, A.; Iwasaki, S.; Otsuka, Y.; Negishi, M.; Akahane, T.; Okai, T. Reduction of progesterone receptor expression in human cumulus cells at the time of oocyte collection during IVF is associated with good embryo quality. Hum. Reprod. 2005, 20, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.H.; Nah, H.Y.; Lee, Y.J.; Lee, J.W.; Park, J.H.; Kim, S.J.; Lee, J.B.; Yoon, H.S.; Kim, C.H. Expression of estrogen receptor-alpha and -beta, glucocorticoid receptor, and progesterone receptor genes in human embryonic stem cells and embryoid bodies. Mol. Cells 2004, 18, 320–325. [Google Scholar]
- Hou, Q.; Gorski, J. Estrogen receptor and progesterone receptor genes are expressed differentially in mouse embryos during preimplantation development. Proc. Natl. Acad. Sci. USA 1993, 90, 9460–9464. [Google Scholar] [CrossRef] [Green Version]
- Rambags, B.P.; van Tol, H.T.; van den Eng, M.M.; Colenbrander, B.; Stout, T.A. Expression of progesterone and oestrogen receptors by early intrauterine equine conceptuses. Theriogenology 2008, 69, 366–375. [Google Scholar] [CrossRef]
- Ying, C.; Yang, Y.C.; Hong, W.F.; Cheng, W.T.; Hsu, W.L. Progesterone receptor gene expression in preimplantation pig embryos. Eur. J. Endocrinol. 2000, 143, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, J.R.; Dumesic, D.A.; Abbott, D.H.; Strauss, J.F., 3rd. Molecular abnormalities in oocytes from women with polycystic ovary syndrome revealed by microarray analysis. J. Clin. Endocrinol. Metab. 2007, 92, 705–713. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, L.; Gagliardi, A.; Landi, C.; Focarelli, R.; De Leo, V.; Luddi, A.; Bini, L.; Piomboni, P. Protein pathways working in human follicular fluid: The future for tailored IVF? Expert Rev. Mol. Med. 2016, 18, e9. [Google Scholar] [CrossRef]
- Moor, R.M.; Polge, C.; Willadsen, S.M. Effect of follicular steroids on the maturation and fertilization of mammalian oocytes. J. Embryol. Exp. Morphol. 1980, 56, 319–335. [Google Scholar]
- Thibault, C.; Gerard, M.; Menezo, Y. Preovulatory and ovulatory mechanisms in oocyte maturation. J. Reprod. Fertil. 1975, 45, 605–610. [Google Scholar] [CrossRef] [Green Version]
- Lonergan, P. Influence of progesterone on oocyte quality and embryo development in cows. Theriogenology 2011, 76, 1594–1601. [Google Scholar] [CrossRef]
- Reinthaller, A.; Deutinger, J.; Riss, P.; Müller-Tyl, E.; Fischl, F.; Bieglmayer, C.; Janisch, H. Relationship between the steroid and prolactin concentration in follicular fluid and the maturation and fertilization of human oocytes. J. Vitr. Fertil. Embryo Transf. 1987, 4, 228–231. [Google Scholar] [CrossRef]
- Urrego, R.; Herrera-Puerta, E.; Chavarria, N.A.; Camargo, O.; Wrenzycki, C.; Rodriguez-Osorio, N. Follicular progesterone concentrations and messenger RNA expression of MATER and OCT-4 in immature bovine oocytes as predictors of developmental competence. Theriogenology 2015, 83, 1179–1187. [Google Scholar] [CrossRef]
- Carpintero, N.L.; Suarez, O.A.; Mangas, C.C.; Varea, C.G.; Rioja, R.G. Follicular steroid hormones as markers of oocyte quality and oocyte development potential. J. Hum. Reprod. Sci. 2014, 7, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Lamb, J.D.; Zamah, A.M.; Shen, S.; McCulloch, C.; Cedars, M.I.; Rosen, M.P. Follicular fluid steroid hormone levels are associated with fertilization outcome after intracytoplasmic sperm injection. Fertil. Steril. 2010, 94, 952–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asimakopoulos, B.; Abu-Hassan, D.; Metzen, E.; Al-Hasani, S.; Diedrich, K.; Nikolettos, N. The levels of steroid hormones and cytokines in individual follicles are not associated with the fertilization outcome after intracytoplasmic sperm injection. Fertil. Steril. 2008, 90, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Artini, P.G.; Battaglia, C.; Ambrogio, G.; Barreca, A.; Droghini, F.; Volpe, A.; Genazzani, A.R. Relationship between human oocyte maturity, fertilization and follicular fluid growth factors. Hum. Reprod. 1994, 9, 902–906. [Google Scholar] [CrossRef] [PubMed]
- Ruan, W.; Monaco, M.E.; Kleinberg, D.L. Progesterone stimulates mammary gland ductal morphogenesis by synergizing with and enhancing insulin-like growth factor-I action. Endocrinology 2005, 146, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- Knight, C.H. An endocrine hypothesis to explain obesity-related lactation insufficiency in breastfeeding mothers. J. Dairy Res. 2020, 87, 78–81. [Google Scholar] [CrossRef]
- Kelsey, J.J. Hormonal contraception and lactation. J. Hum. Lact. 1996, 12, 315–318. [Google Scholar] [CrossRef]
- Goletiani, N.V.; Keith, D.R.; Gorsky, S.J. Progesterone: Review of safety for clinical studies. Exp. Clin. Psychopharmacol. 2007, 15, 427–444. [Google Scholar] [CrossRef]
- Carroll, J.S.; Hickey, T.E.; Tarulli, G.A.; Williams, M.; Tilley, W.D. Deciphering the divergent roles of progestogens in breast cancer. Nat. Rev. Cancer 2017, 17, 54–64. [Google Scholar] [CrossRef]
- Ali, S.; Balachandran, K.; O’Malley, B. 90 Years of progesterone: Ninety years of progesterone: The ‘other’ ovarian hormone. J. Mol. Endocrinol. 2020, 65, E1–E4. [Google Scholar] [CrossRef]
- Warren, M.P.; Shantha, S. Uses of progesterone in clinical practice. Int. J. Fertil. Womens Med. 1999, 44, 96–103. [Google Scholar] [PubMed]
- Apgar, B.S.; Greenberg, G. Using progestins in clinical practice. Am. Fam. Physician 2000, 62, 1839–1846. [Google Scholar]
- Doblinger, J.; Cometti, B.; Trevisan, S.; Griesinger, G. Subcutaneous Progesterone Is Effective and Safe for Luteal Phase Support in IVF: An Individual Patient Data Meta-Analysis of the Phase III Trials. PLoS ONE 2016, 11, e0151388. [Google Scholar] [CrossRef] [PubMed]
- Van der Linden, M.; Buckingham, K.; Farquhar, C.; Kremer, J.A.; Metwally, M. Luteal phase support for assisted reproduction cycles. Cochrane Database Syst. Rev. 2015, 2015, CD009154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miralpeix, E.; Gonzalez-Comadran, M.; Sola, I.; Manau, D.; Carreras, R.; Checa, M.A. Efficacy of luteal phase support with vaginal progesterone in intrauterine insemination: A systematic review and meta-analysis. J. Assist. Reprod. Genet. 2014, 31, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Filicori, M. Clinical roles and applications of progesterone in reproductive medicine: An overview. Acta Obstet. Gynecol. Scand. 2015, 94, 3–7. [Google Scholar] [CrossRef]
- Connell, M.T.; Szatkowski, J.M.; Terry, N.; DeCherney, A.H.; Propst, A.M.; Hill, M.J. Timing luteal support in assisted reproductive technology: A systematic review. Fertil. Steril. 2015, 103, 939–946.e3. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.R.; Mu, H.Q.; Shi, Q.; Xiao, X.Q.; Qi, H.B. The optimal duration of progesterone supplementation in pregnant women after IVF/ICSI: A meta-analysis. Reprod. Biol. Endocrinol. 2012, 10, 107. [Google Scholar] [CrossRef] [Green Version]
- Coomarasamy, A.; Devall, A.J.; Brosens, J.J.; Quenby, S.; Stephenson, M.D.; Sierra, S.; Christiansen, O.B.; Small, R.; Brewin, J.; Roberts, T.E.; et al. Micronized vaginal progesterone to prevent miscarriage: A critical evaluation of randomized evidence. Am. J. Obstet. Gynecol. 2020, 223, 167–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, R.; Conde-Agudelo, A.; Da Fonseca, E.; O’Brien, J.M.; Cetingoz, E.; Creasy, G.W.; Hassan, S.S.; Nicolaides, K.H. Vaginal progesterone for preventing preterm birth and adverse perinatal outcomes in singleton gestations with a short cervix: A meta-analysis of individual patient data. Am. J. Obstet. Gynecol. 2018, 218, 161–180. [Google Scholar] [CrossRef] [Green Version]
- Young, S.L. Oestrogen and progesterone action on endometrium: A translational approach to understanding endometrial receptivity. Reprod. Biomed. Online 2013, 27, 497–505. [Google Scholar] [CrossRef] [Green Version]
- Bulun, S.E.; Cheng, Y.H.; Pavone, M.E.; Xue, Q.; Attar, E.; Trukhacheva, E.; Tokunaga, H.; Utsunomiya, H.; Yin, P.; Luo, X.; et al. Estrogen receptor-beta, estrogen receptor-alpha, and progesterone resistance in endometriosis. Semin. Reprod. Med. 2010, 28, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Tanbo Tom, F.P. Endometriosis-associated infertility: Aspects ofpathophysiological mechanisms and treatment options. Acta Obstet. Gynecol. Scand. 2017, 96, 659–667. [Google Scholar] [CrossRef]
- Speroff, L.; Kase, N.G. Oral contraception. In Clinical Gynecologic Endocrinology and Infertility, 6th ed.; Lippincott Williams & Wilkins: Baltimore, MD, USA, 2005; Volume 1999, pp. 867–945. [Google Scholar]
- Krattenmacher, R. Drospirenone: Pharmacology and pharmacokinetics of a unique progestogen. Contraception 2000, 62, 29–38. [Google Scholar] [CrossRef]
- Apter, D.; Zimmerman, Y.; Beekman, L.; Mawet, M.; Maillard, C.; Foidart, J.M.; Coelingh Bennink, H.J. Estetrol combined with drospirenone: An oral contraceptive with high acceptability, user satisfaction, well-being and favourable body weight control. Eur. J. Contracept. Reprod. Health Care 2017, 22, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Bharucha, M.R. Progestogens in gynaecological practice. Indian Med. Trib. 1995, 3, 6–7. [Google Scholar]
- Abdi, S.; Salehnia, M.; Hosseinkhani, S. Steroid Production and Follicular Development of Neonatal Mouse Ovary during in vitro Culture. Int. J. Fertil. Steril. 2013, 7, 181–186. [Google Scholar]
- Harper-Harrison, G.; Shanahan, M.M. Hormone Replacement Therapy; StatPearls: Treasure Island, FL, USA, 2021. [Google Scholar]
- Edwards, M.; Can, A.S. Progestin; StatPearls: Treasure Island, FL, USA, 2021. [Google Scholar]
- Cobin, R.H.; Goodman, N.F.; Committee ARES. American Association of Clinical Endocrinologists and American College of Endocrinology Position Statement on Menopause-2017 Update. Endocr. Pract. 2017, 23, 869–880. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, D.E.; Ruan, Y.; Forbes, N.; Heitman, S.J.; Hilsden, R.J.; Pader, J.; Brenner, D.R. Long-term Use of Hormone Replacement Therapy is Associated With a Lower Risk of Developing High-risk Serrated Polyps in Women. J. Clin. Gastroenterol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, A.; Curtis, L. In Defense of Progesterone: A Review of the Literature. Altern. Ther. Health Med. 2017, 23, 24–32. [Google Scholar]
- Lange, C.A.; Yee, D. Progesterone and breast cancer. Womens Health 2008, 4, 151–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Orozco, J.C.; Camacho-Arroyo, I. Progesterone Actions During Central Nervous System Development. Front. Neurosci. 2019, 13, 503. [Google Scholar] [CrossRef] [Green Version]
- Espinoza, T.R.; Wright, D.W. The role of progesterone in traumatic brain injury. J. Head Trauma Rehabil. 2011, 26, 497–499. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1897 | J Beard | Absence of ovulation during pregnancy |
| 1898 | L A Prenant | Corpus luteum is necessary for implantation |
| 1916 | E Herrmann and M Stein | Extract from the corpus luteum blocks the ovulation |
| 1923 | W M Allen | Effect of the corpus luteum extract on castrated rabbit’s endometria |
| 1931 | L Haberlandt | Birth control through temporary hormonal sterilization |
| 1933–1934 | K H Slotta, P Wintersteiner, A Butenandt, U Westphal, W M Allen, M Hartmann, A Wettstein | Isolation of progesterone |
| 1951 | C Djerassi | First synthesis of an oral contraceptive norethindrone |
| 1970 | B W O’Malley | Progesterone receptors and isoforms |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagy, B.; Szekeres-Barthó, J.; Kovács, G.L.; Sulyok, E.; Farkas, B.; Várnagy, Á.; Vértes, V.; Kovács, K.; Bódis, J. Key to Life: Physiological Role and Clinical Implications of Progesterone. Int. J. Mol. Sci. 2021, 22, 11039. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011039
Nagy B, Szekeres-Barthó J, Kovács GL, Sulyok E, Farkas B, Várnagy Á, Vértes V, Kovács K, Bódis J. Key to Life: Physiological Role and Clinical Implications of Progesterone. International Journal of Molecular Sciences. 2021; 22(20):11039. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011039
Chicago/Turabian StyleNagy, Bernadett, Júlia Szekeres-Barthó, Gábor L. Kovács, Endre Sulyok, Bálint Farkas, Ákos Várnagy, Viola Vértes, Kálmán Kovács, and József Bódis. 2021. "Key to Life: Physiological Role and Clinical Implications of Progesterone" International Journal of Molecular Sciences 22, no. 20: 11039. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011039