
Macrophage Polarization Related to Crystal Phases of Calcium Phosphate Biomaterials

 and
and
Abstract
:1. Introduction
2. Results
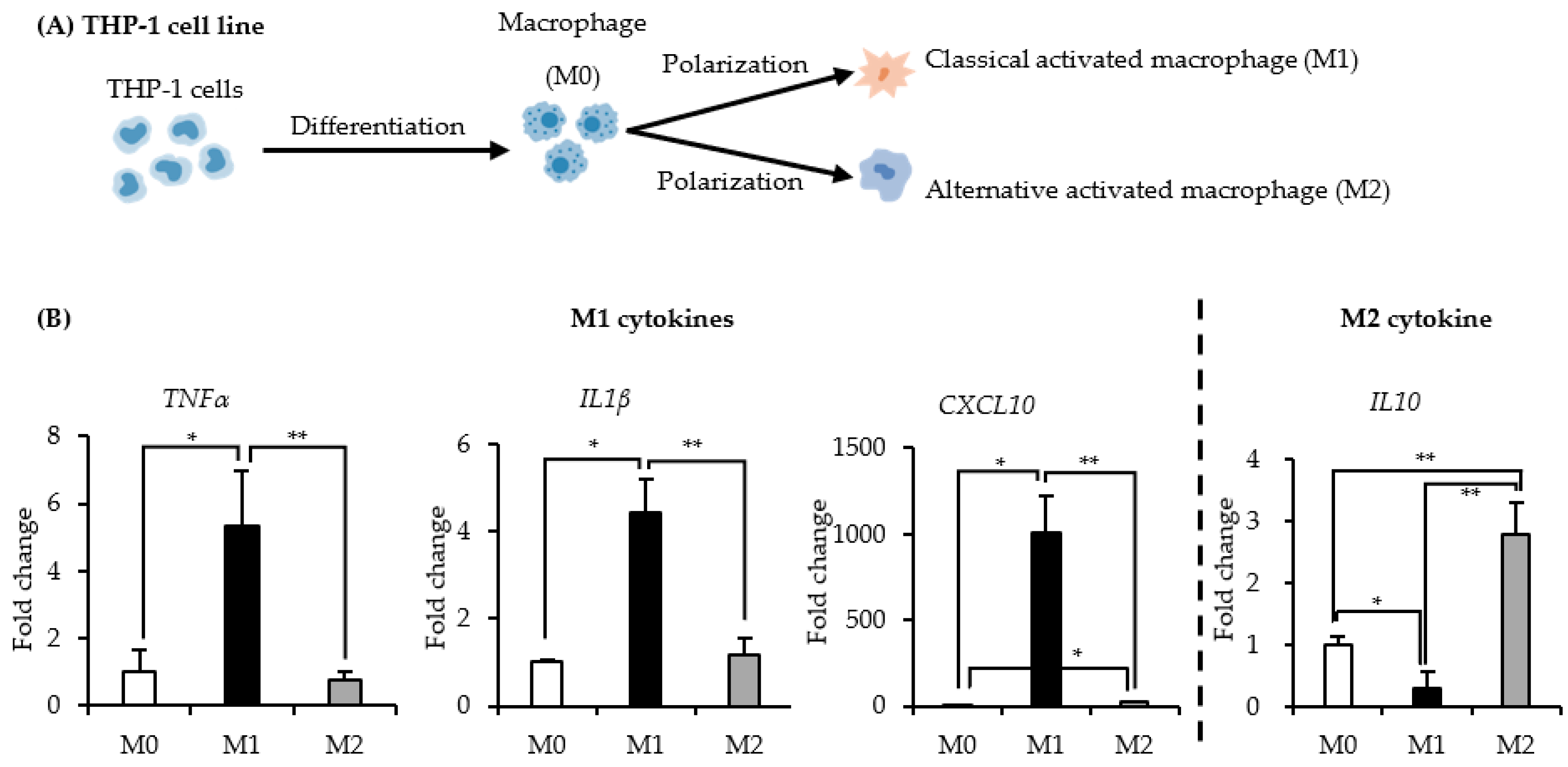
2.1. Identification of the THP-1 Derived M1 and M2 Macrophages
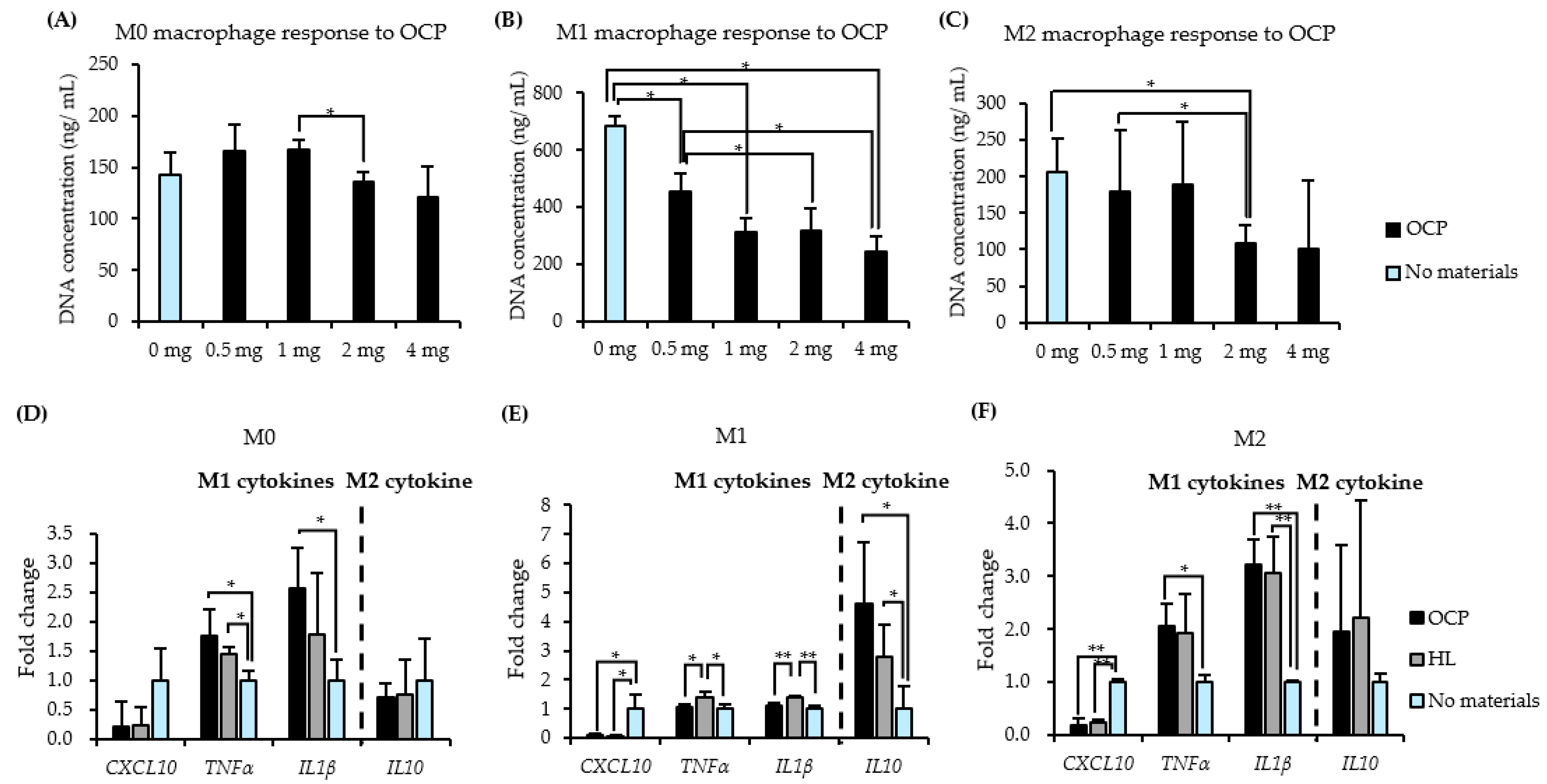
2.2. Responses of THP-1 Derived Macrophages to CaP Granules
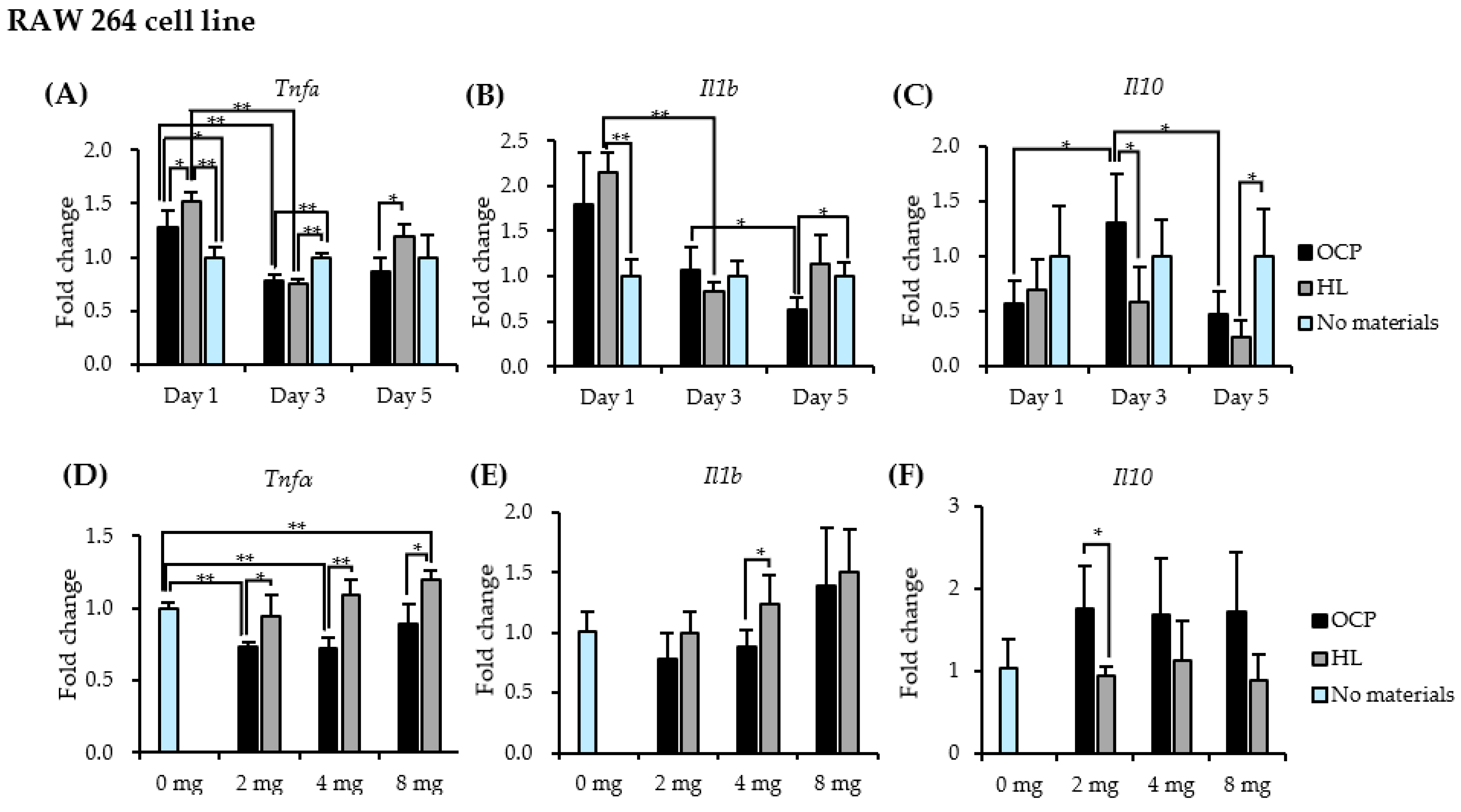
2.3. Responses of RAW 264 Cells to CaP granules
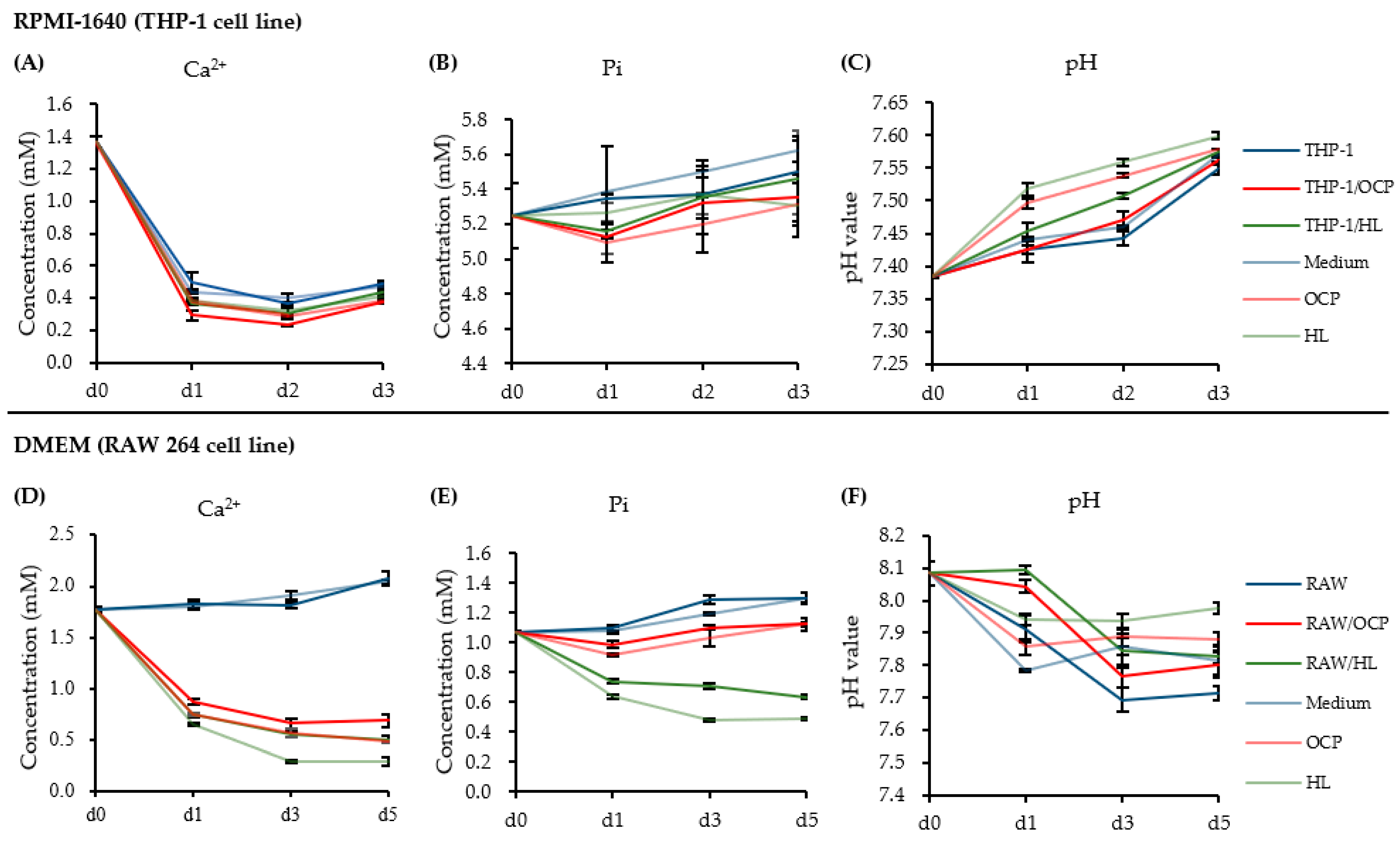
2.4. Changes in the Ionic Microenvironment in the Macrophage Medium in the Presence of CaP Granules
2.5. Dose-Response of CaP Granules in an Ionic Microenvironment
2.6. Analysis of the Degree of Supersaturation (DS) in DMEM
2.7. Characterization of the Spectroscopic Features of CaP Granules
3. Discussion
4. Materials and Methods
4.1. Synthesis of CaP Granules
4.2. Cell Differentiation, Polarization, and Treatment
4.3. Evaluation of the Expression of M1 and M2 Macrophage-Related Genes by qPCR
4.4. Measurement of Changes in the Ionic Microenvironment in the CaP Granule-Treated Macrophage Culture Medium
4.5. Determination of the DS in the DMEM Immersed with OCP or HL Granules
4.6. FTIR Analysis of OCP and HL after Immersion in the Macrophage Medium
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Julier, Z.; Park, A.J.; Briquez, P.S.; Martino, M.M. Promoting tissue regeneration by modulating the immune system. Acta Biomater. 2017, 53, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Ghiasi, M.S.; Chen, J.; Vaziri, A.; Rodriguez, E.K.; Nazarian, A. Bone fracture healing in mechanobiological modeling: A review of principles and methods. Bone Rep. 2017, 6, 87–100. [Google Scholar] [CrossRef]
- Tsukasaki, M.; Takayanagi, H. Osteoimmunology: Evolving concepts in bone-immune interactions in health and disease. Nat. Rev. Immunol. 2019, 19, 626–642. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Mao, X.; Tan, L.; Friis, T.; Wu, C.; Crawford, R.; Xiao, Y. Osteoimmunomodulatory properties of magnesium scaffolds coated with beta-tricalcium phosphate. Biomaterials 2014, 35, 8553–8565. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Klein, T.; Murray, R.Z.; Crawford, R.; Chang, J.; Wu, C.; Xiao, Y. Osteoimmunomodulation for the development of advanced bone biomaterials. Mater. Today 2016, 19, 304–321. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.M.; Rodriguez, A.; Chang, D.T. Foreign body reaction to biomaterials. Semin Immunol. 2008, 20, 86–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, W. Crystal growth of bone mineral. Clin. Orthop. Relat. Res. 1966, 44, 205–220. [Google Scholar] [CrossRef]
- Suzuki, O.; Nakamura, M.; Miyasaka, Y.; Kagayama, M.; Sakurai, M. Bone Formation on Synthetic Precursors of Hydroxyapatite. Tohoku J. Exp. Med. 1991, 164, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, O. Octacalcium phosphate: Osteoconductivity and crystal chemistry. Acta Biomater. 2010, 6, 3379–3387. [Google Scholar] [CrossRef]
- Suzuki, O.; Shiwaku, Y.; Hamai, R. Octacalcium phosphate bone substitute materials: Comparison between properties of biomaterials and other calcium phosphate materials. Dent. Mater. J. 2020, 39, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, O.; Kamakura, S.; Katagiri, T.; Nakamura, M.; Zhao, B.; Honda, Y.; Kamijo, R. Bone formation enhanced by implanted octacalcium phosphate involving conversion into Ca-deficient hydroxyapatite. Biomaterials 2006, 27, 2671–2681. [Google Scholar] [CrossRef] [PubMed]
- Anada, T.; Kumagai, T.; Honda, Y.; Masuda, T.; Kamijo, R.; Kamakura, S.; Yoshihara, N.; Kuriyagawa, T.; Shimauchi, H.; Suzuki, O. Dose-Dependent Osteogenic Effect of Octacalcium Phosphate on Mouse Bone Marrow Stromal Cells. Tissue Eng. Part A 2008, 14, 965–978. [Google Scholar] [CrossRef] [PubMed]
- Sai, Y.; Shiwaku, Y.; Anada, T.; Tsuchiya, K.; Takahashi, T.; Suzuki, O. Capacity of octacalcium phosphate to promote osteoblastic differentiation toward osteocytes in vitro. Acta Biomater. 2018, 69, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Hamai, R.; Shiwaku, Y.; Hasegawa, T.; Sakai, S.; Tsuchiya, K.; Sai, Y.; Iwama, R.; Amizuka, N.; Takahashi, T.; et al. Involvement of distant octacalcium phosphate scaffolds in enhancing early differentiation of osteocytes during bone regeneration. Acta Biomater. 2021, 129, 309–322. [Google Scholar] [CrossRef]
- Takami, M.; Mochizuki, A.; Yamada, A.; Tachi, K.; Zhao, B.; Miyamoto, Y.; Anada, T.; Honda, Y.; Inoue, T.; Nakamura, M.; et al. Osteoclast Differentiation Induced by Synthetic Octacalcium Phosphate Through Receptor Activator of NF-kB Ligand Expression in Osteoblasts. Tissue Eng. Part A 2009, 15, 3991–4000. [Google Scholar] [CrossRef]
- Honda, Y.; Anada, T.; Kamakura, S.; Nakamura, M.; Sugawara, S.; Suzuki, O. Elevated extracellular calcium stimulates secretion of bone morphogenetic protein 2 by a macrophage cell line. Biochem. Biophys. Res. Commun. 2006, 345, 1155–1160. [Google Scholar] [CrossRef]
- Shiwaku, Y.; Tsuchiya, K.; Xiao, L.; Suzuki, O. Effect of calcium phosphate phases affecting the crosstalk between osteoblasts and osteoclasts in vitro. J. Biomed. Mater. Res. A 2019, 107, 1001–1013. [Google Scholar] [CrossRef]
- Hirayama, B.; Anada, T.; Shiwaku, Y.; Miyatake, N.; Tsuchiya, K.; Nakamura, M.; Takahashi, T.; Suzuki, O. Immune cell response and subsequent bone formation induced by implantation of octacalcium phosphate in a rat tibia defect. RSC Adv. 2016, 6, 57475–57484. [Google Scholar] [CrossRef]
- Morimoto, S.; Anada, T.; Honda, Y.; Suzuki, O. Comparative study on in vitro biocompatibility of synthetic octacalcium phosphate and calcium phosphate ceramics used clinically. Biomed. Mater. 2012, 7, 045020. [Google Scholar] [CrossRef]
- Suzuki, O.; Kamakura, S.; Katagiri, T. Surface chemistry and biological responses to synthetic octacalcium phosphate. J. Biomed. Mater. Res. B Appl. Biomater. 2006, 77, 201–212. [Google Scholar] [CrossRef]
- Miyatake, N.; Kishimoto, K.N.; Anada, T.; Imaizumi, H.; Itoi, E.; Suzuki, O. Effect of partial hydrolysis of octacalcium phosphate on its osteoconductive characteristics. Biomaterials 2009, 30, 1005–1014. [Google Scholar] [CrossRef]
- Brown, B.N.; Badylak, S.F. Expanded applications, shifting paradigms and an improved understanding of host-biomaterial interactions. Acta Biomater. 2013, 9, 4948–4955. [Google Scholar] [CrossRef]
- Devlin, R.D.; Reddy, S.V.; Savino, R.; Ciliberto, G.; Roodman, G.D. IL-6 mediates the effects of IL-1 or TNF, but not PTHrP or 1,25(OH)2D3, on osteoclast-like cell formation in normal human bone marrow cultures. J. Bone Miner Res. 1998, 13, 393–399. [Google Scholar] [CrossRef]
- Walker, H.K.; Hall, W.D.; Hurst, J.W. (Eds.) Clinical Methods: The History, Physical, and Laboratory Examinations, 3rd ed.; Butterworths: Boston, WA, USA, 1990. [Google Scholar] [PubMed]
- Mokarram, N.; Bellamkonda, R.V. A perspective on immunomodulation and tissue repair. Ann. Biomed. Eng. 2014, 42, 338–351. [Google Scholar] [CrossRef]
- Freytes, D.O.; Kang, J.W.; Marcos-Campos, I.; Vunjak-Novakovic, G. Macrophages modulate the viability and growth of human mesenchymal stem cells. J. Cell Biochem. 2013, 114, 220–229. [Google Scholar] [CrossRef]
- Champagne, C.M.; Takebe, J.; Offenbacher, S.; Cooper, L.F. Macrophage cell lines produce osteoinductive signals that include bone morphogenetic protein-2. Bone 2002, 30, 26–31. [Google Scholar] [CrossRef]
- Chen, X.; Wang, M.; Chen, F.; Wang, J.; Li, X.; Liang, J.; Fan, Y.; Xiao, Y.; Zhang, X. Correlations between macrophage polarization and osteoinduction of porous calcium phosphate ceramics. Acta Biomater. 2020, 103, 318–332. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.S.; He, D.Q.; Luo, D.; Wang, Y.; Yu, M.; Guan, B.; Fu, Y.; Li, Z.X.; Zhang, T.; Zhou, Y.H.; et al. A Biomimetic Hierarchical Nanointerface Orchestrates Macrophage Polarization and Mesenchymal Stem Cell Recruitment to Promote Endogenous Bone Regeneration. ACS Nano 2019, 13, 6581–6595. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Guo, X.; Qi, W.; Wu, Z.; de Bruijn, J.D.; Xiao, Y.; Bao, C.; Yuan, H. Macrophage polarization plays roles in bone formation instructed by calcium phosphate ceramics. J. Mater. Chem. B 2020, 8, 1863–1877. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wu, C.; Gu, W.; Klein, T.; Crawford, R.; Xiao, Y. Osteogenic differentiation of bone marrow MSCs by beta-tricalcium phosphate stimulating macrophages via BMP2 signalling pathway. Biomaterials 2014, 35, 1507–1518. [Google Scholar] [CrossRef]
- Sadowska, J.M.; Wei, F.; Guo, J.; Guillem-Marti, J.; Lin, Z.; Ginebra, M.P.; Xiao, Y. The effect of biomimetic calcium deficient hydroxyapatite and sintered beta-tricalcium phosphate on osteoimmune reaction and osteogenesis. Acta Biomater. 2019, 96, 605–618. [Google Scholar] [CrossRef]
- Villa-Bellosta, R.; Hamczyk, M.R.; Andres, V. Novel phosphate-activated macrophages prevent ectopic calcification by increasing extracellular ATP and pyrophosphate. PLoS ONE 2017, 12, e0174998. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Yin, Y.; Hu, X.; Peng, C.; Liu, Y.; Li, Q.; Huang, W.; Huang, Q. Effects of Environmental pH on Macrophage Polarization and Osteoimmunomodulation. ACS Biomater. Sci. Eng. 2019, 5, 5548–5557. [Google Scholar] [CrossRef] [PubMed]
- Masuda, T.; Maruyama, H.; Arai, F.; Anada, T.; Tsuchiya, K.; Fukuda, T.; Suzuki, O. Application of an indicator-immobilized-gel-sheet for measuring the pH surrounding a calcium phosphate-based biomaterial. J. Biomater. Appl. 2017, 31, 1296–1304. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, K.R.; Zhang, Y.; Magri, A.M.P.; Renno, A.C.M.; van den Beucken, J. Biomaterial Property Effects on Platelets and Macrophages: An in Vitro Study. ACS Biomater. Sci. Eng. 2017, 3, 3318–3327. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Wang, Y.; Zheng, D.; Sun, Y.; Wang, Y.; Lee, V.W.S.; Zheng, G.; Tan, T.K.; Ince, J.; Alexander, S.I.; et al. IL-10/TGF-beta-modified macrophages induce regulatory T cells and protect against adriamycin nephrosis. J. Am. Soc. Nephrol. 2010, 21, 933–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoba, T.; Fukae, M.; Tanabe, T.; Shimizu, M.; Moreno, E.C. Selective Adsorption of Porcine-Amelogenins onto Hydroxyapatite and their Inhibitory Activity on Hydroxyapatite Growth in Supersaturated Solutions. Calcif. Tissue Int. 1987, 41, 281–289. [Google Scholar] [CrossRef]
- Moreno, E.C.; Aoba, T. Calcium bonding in enamel fluid and driving force for enamel mineralization in the secretory stage of amelogenesis. Adv. Dent. Res. 1987, 1, 245–251. [Google Scholar] [CrossRef]
- Moreno, E.C.; Aoba, T. Comparative solubility study of human dental enamel, dentin, and hydroxyapatite. Calcif. Tissue Int. 1991, 49, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Moreno, E.C.; Kresak, M.; Zahradnik, R.T. Fluoridated hydroxyapatite solubility and caries formation. Nature 1974, 247, 64–65. [Google Scholar] [CrossRef]
- Tung, M.S.; Eidelman, N.; Sieck, B.; Brown, W.E. Octacalcium Phosphate Solubility Product from 4 to 37 °C. J. Res. Natl. Bur. Stand. 1988, 93, 613–624. [Google Scholar] [CrossRef]
- Moreno, E.C.; Brown, W.E.; Osborn, G. Solubility of dicalcium phosphate dihydrate in aqueous systems. Soil. Sci. Soc. Am. J. 1960, 24, 94–98. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period (Days) | Ca2+ (mM) | Pi (mM) | pH | DS at pH (Each) and 37 °C | |||
|---|---|---|---|---|---|---|---|
| HA | OCP | DCPD | |||||
| DMEM | 0 | 1.78 | 1.07 | 8.08 | 3.28 × 1014 | 4.54 × 104 | 6.72 × 10−1 |
| RAW cells | 1 | 1.82 | 1.10 | 7.91 | 7.84 × 1013 | 2.31 × 104 | 6.90 × 10−1 |
| RAW/OCP | 1 | 0.88 | 0.99 | 8.04 | 6.13 × 1012 | 2.03 × 103 | 3.18 × 10−1 |
| RAW/HL | 1 | 0.75 | 0.74 | 8.10 | 2.03 × 1012 | 6.12 × 102 | 2.07 × 10−1 |
| OCP | 1 | 0.75 | 0.92 | 7.86 | 3.78 × 1011 | 3.46 × 102 | 2.48 × 10−1 |
| HL | 1 | 0.65 | 0.64 | 7.94 | 1.50 × 1011 | 1.06 × 102 | 1.54 × 10−1 |
| RAW cells | 3 | 1.82 | 1.28 | 7.69 | 1.44 × 1013 | 1.16 × 104 | 7.68 × 10−1 |
| RAW/OCP | 3 | 0.66 | 1.10 | 7.77 | 1.38 × 1011 | 2.19 × 102 | 2.56 × 10−1 |
| RAW/HL | 3 | 0.56 | 0.71 | 7.85 | 3.90 × 1010 | 4.96 × 101 | 1.45 × 10−1 |
| OCP | 3 | 0.57 | 1.03 | 7.89 | 1.82 × 1011 | 1.91 × 102 | 2.13 × 10−1 |
| HL | 3 | 0.30 | 0.48 | 7.94 | 1.37 × 1011 | 2.13 × 100 | 5.43 × 10−2 |
| RAW cells | 5 | 2.08 | 1.30 | 7.72 | 3.52 × 1013 | 2.26 × 104 | 8.87 × 10−1 |
| RAW/OCP | 5 | 0.69 | 1.12 | 7.80 | 2.56 × 1011 | 3.30 × 102 | 2.74 × 10−1 |
| RAW/HL | 5 | 0.51 | 0.64 | 7.83 | 1.44 × 1010 | 2.19 × 101 | 1.17 × 10−1 |
| OCP | 5 | 0.48 | 1.12 | 7.88 | 9.04 × 1010 | 1.18 × 102 | 1.95 × 10−1 |
| HL | 5 | 0.30 | 0.49 | 7.98 | 1.79 × 109 | 2.41 × 100 | 5.39 × 10−2 |
| Species | Gene | PCR Primers (5′-3′) | Universal Probe |
|---|---|---|---|
| Human | TNFα | Forward: agcccatgttgtagcaaacc | #79 |
| Reverse: tctcagctccacgccatt | |||
| IL1β | Forward: agccaggacagtcagctctc | #23 | |
| Reverse: agaggcctggctcaacaa | |||
| CXCL10 | Forward: aagcagttagcaaggaaaggtc | #34 | |
| Reverse: gacatatactccatgtagggaagtga | |||
| IL10 | Forward: ttgcctggtcctcctgact | #37 | |
| Reverse: gaagtgggtgcagctgttct | |||
| GAPDH | Forward: ccccggtttctataaattgagc | #63 | |
| Reverse: cttccccatggtgtctgag | |||
| Mouse | Tnfα | Forward: ctgtagcccacgtcgtagc | #25 |
| Reverse: tttgagatccatgccgttg | |||
| Il1β | Forward: agttgacggaccccaaaag | #38 | |
| Reverse: agctggatgctctcatcagg | |||
| Il10 | Forward: cagagccacatgctcctaga | #41 | |
| Reverse: tgtccagctggtcctttgtt | |||
| Gapdh | Forward: tgtccgtcgtggatctgac | #80 | |
| Reverse: cctgcttcaccaccttcttg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, L.; Shiwaku, Y.; Hamai, R.; Tsuchiya, K.; Sasaki, K.; Suzuki, O. Macrophage Polarization Related to Crystal Phases of Calcium Phosphate Biomaterials. Int. J. Mol. Sci. 2021, 22, 11252. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011252
Xiao L, Shiwaku Y, Hamai R, Tsuchiya K, Sasaki K, Suzuki O. Macrophage Polarization Related to Crystal Phases of Calcium Phosphate Biomaterials. International Journal of Molecular Sciences. 2021; 22(20):11252. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011252
Chicago/Turabian StyleXiao, Linghao, Yukari Shiwaku, Ryo Hamai, Kaori Tsuchiya, Keiichi Sasaki, and Osamu Suzuki. 2021. "Macrophage Polarization Related to Crystal Phases of Calcium Phosphate Biomaterials" International Journal of Molecular Sciences 22, no. 20: 11252. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011252