
OCT1 Is a Poor Prognostic Factor for Breast Cancer Patients and Promotes Cell Proliferation via Inducing NCAPH

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
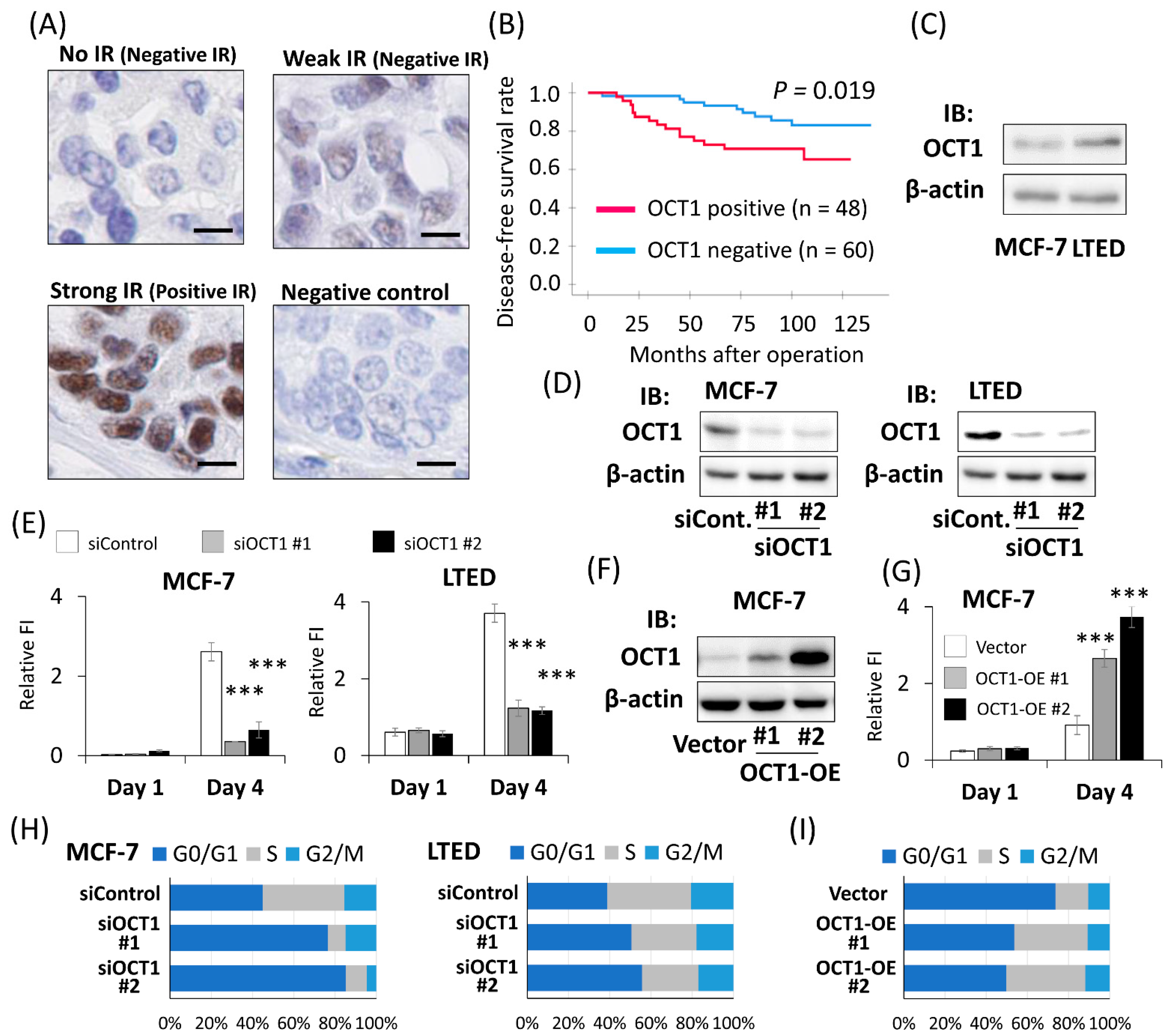
2.1. Positive OCT1 Immunoreactivity Was Associated with the Poor Prognosis of Breast Cancer Patients
2.2. OCT1 Promoted the Cell Proliferation of Breast Cancer Cells
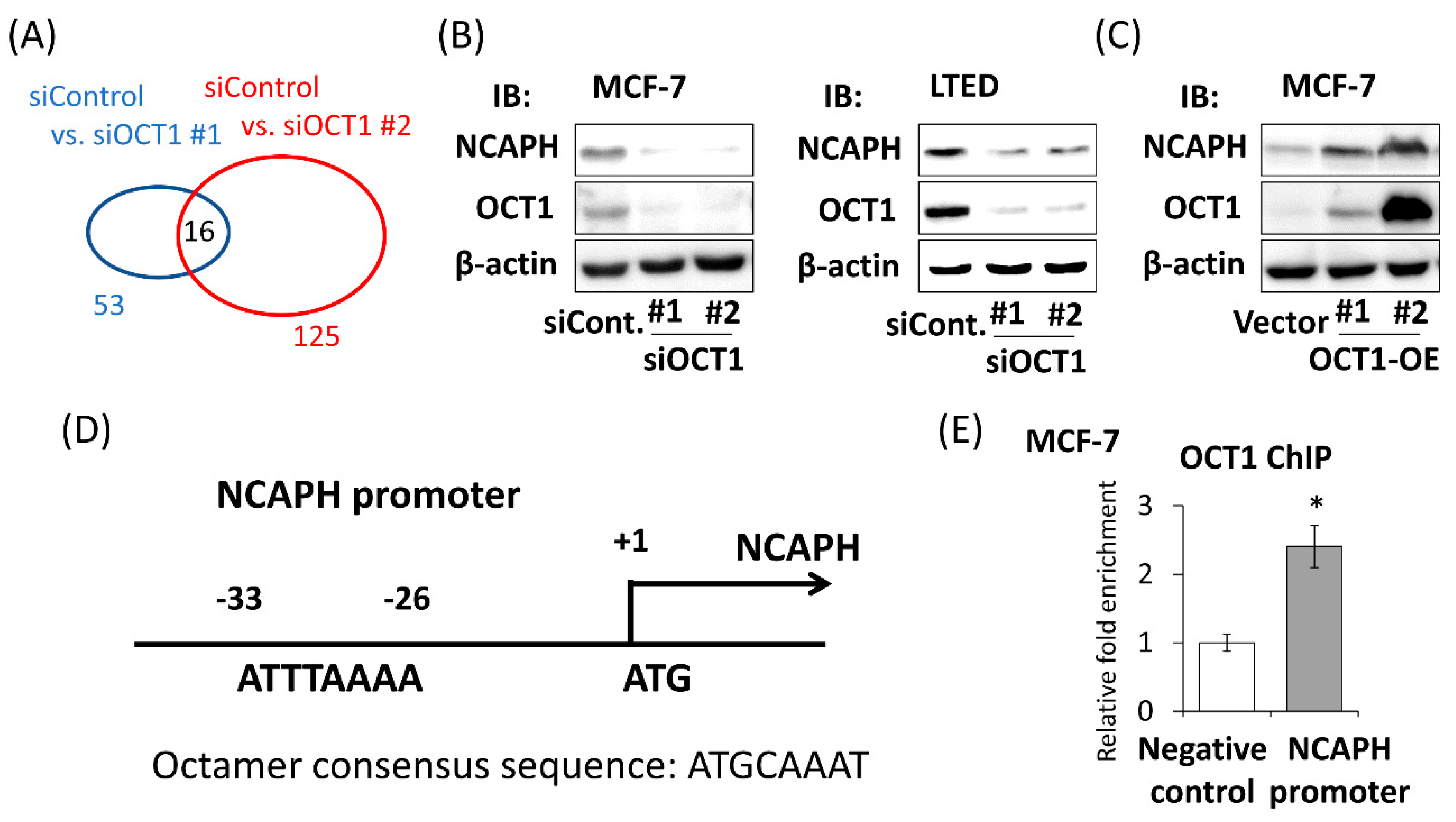
2.3. Identification of OCT1-Induced Genes
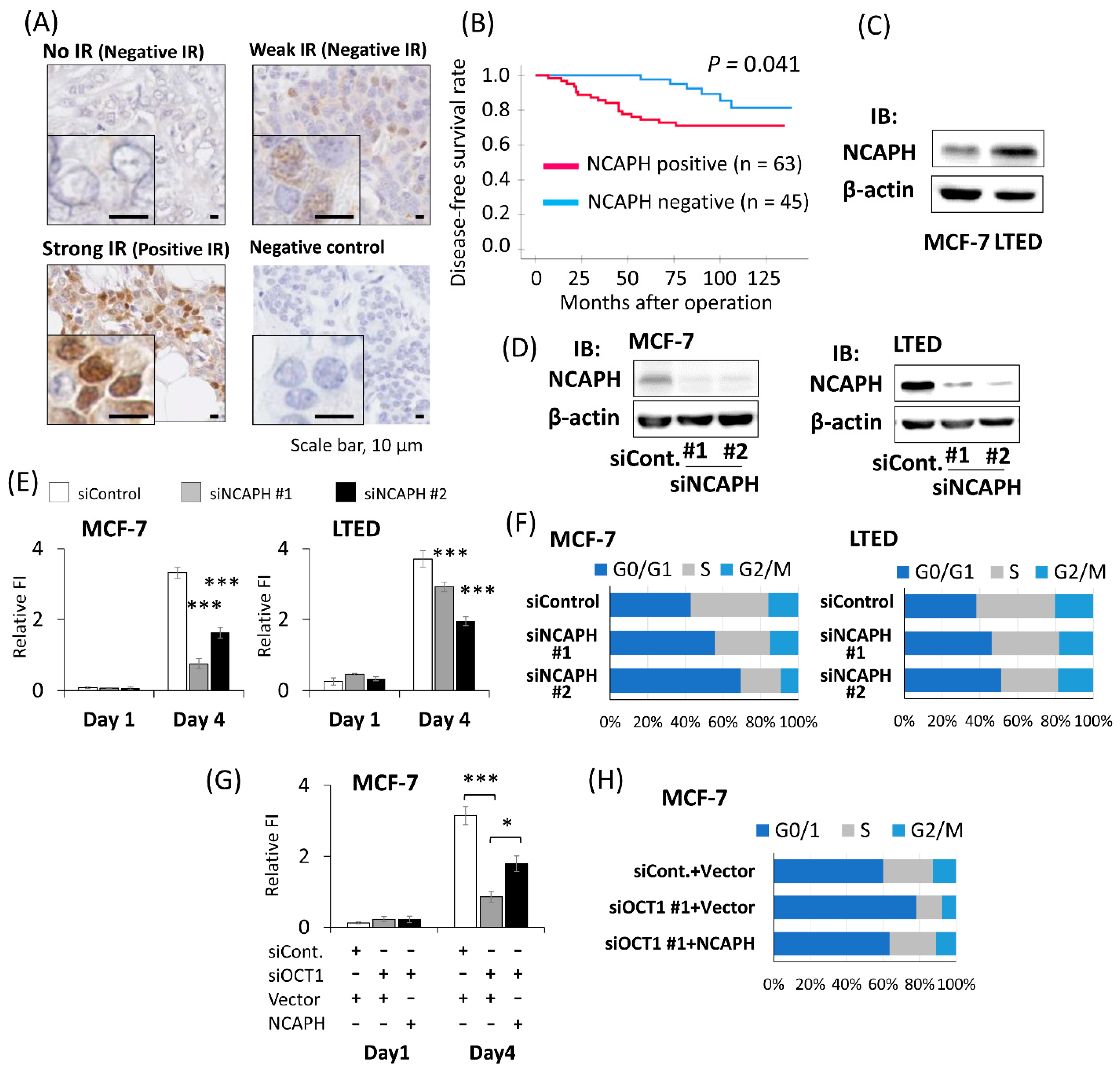
2.4. Positive NCAPH Immunoreactivity Was Associated with the Poor Prognosis of Breast Cancer Patients
2.5. Knockdown of NCAPH Suppressed the Proliferation of Breast Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Collection of Human Tissue Samples and Clinical Data
4.2. Antibodies
4.3. Immunohistochemical Analysis
4.4. Cell Culture
4.5. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) Analysis
4.6. Western Blot Analysis
4.7. Small Interfering RNA Transfection
4.8. Plasmid Construction and Transfection
4.9. Cell Proliferation Assay
4.10. Flow Cytometric Analysis
4.11. Microarray Analysis
4.12. ChIP Assay
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Heer, E.; Harper, A.; Escandor, N.; Sung, H.; McCormack, V.; Fidler-Benaoudia, M.M. Global burden and trends in premenopausal and postmenopausal breast cancer: A population-based study. Lancet Glob. Health 2020, 8, e1027–e1037. [Google Scholar] [CrossRef]
- Acheampong, T.; Kehm, R.D.; Terry, M.B.; Argov, E.L.; Tehranifar, P. Incidence Trends of Breast Cancer Molecular Subtypes by Age and Race/Ethnicity in the US From 2010 to 2016. JAMA Netw. Open 2020, 3, e2013226. [Google Scholar] [CrossRef] [PubMed]
- Emens, L.A. Breast Cancer Immunotherapy: Facts and Hopes. Clin. Cancer Res. 2017, 24, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Saji, S.; Kimura-Tsuchiya, R. Combination of molecular-targeted drugs with endocrine therapy for hormone-resistant breast cancer. Int. J. Clin. Oncol. 2015, 20, 268–272. [Google Scholar] [CrossRef]
- Clarke, R.; Tyson, J.J.; Dixon, J.M. Endocrine resistance in breast cancer—An overview and update. Mol. Cell. Endocrinol. 2015, 418, 220–234. [Google Scholar] [CrossRef] [Green Version]
- Sturm, R.A.; Das, G.; Herr, W. The ubiquitous octamer-binding protein Oct-1 contains a POU domain with a homeo box subdomain. Genes Dev. 1988, 2, 1582–1599. [Google Scholar] [CrossRef] [PubMed]
- Obinata, D.; Takayama, K.; Fujiwara, K.; Suzuki, T.; Tsutsumi, S.; Fukuda, N.; Nagase, H.; Fujimura, T.; Urano, T.; Homma, Y.; et al. Targeting Oct1 genomic function inhibits androgen receptor signaling and castration-resistant prostate cancer growth. Oncogene 2016, 35, 6350–6358. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Takayama, K.; Obinata, D.; Fujiwara, K.; Ashikari, D.; Takahashi, S.; Inoue, S. Identification of new octamer transcription factor 1-target genes upregulated in castration-resistant prostate cancer. Cancer Sci. 2019, 110, 3476–3485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayama, K.-I.; Suzuki, Y.; Yamamoto, S.; Obinata, D.; Takahashi, S.; Inoue, S. Integrative Genomic Analysis of OCT1 Reveals Coordinated Regulation of Androgen Receptor in Advanced Prostate Cancer. Endocrinology 2019, 160, 463–472. [Google Scholar] [CrossRef]
- Boulon, S.; Dantonel, J.-C.; Binet, V.; Vié, A.; Blanchard, J.-M.; Hipskind, R.A.; Philips, A. Oct-1 Potentiates CREB-Driven Cyclin D1 Promoter Activation via a Phospho-CREB- and CREB Binding Protein-Independent Mechanism. Mol. Cell Biol. 2002, 22, 7769–7779. [Google Scholar] [CrossRef] [Green Version]
- Dalvai, M.; Schubart, K.; Besson, A.; Matthias, P. Oct1 is required for mTOR-induced G1cell cycle arrest via the control of p27Kip1expression. Cell Cycle 2010, 9, 3933–3944. [Google Scholar] [CrossRef] [Green Version]
- Fan, W.; Jin, S.; Tong, T.; Zhao, H.; Fan, F.; Antinore, M.J.; Rajasekaran, B.; Wu, M.; Zhan, Q. BRCA1 Regulates GADD45 through Its Interactions with the OCT-1 and CAAT Motifs. J. Biol. Chem. 2002, 277, 8061–8067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.-B.; Fan, J.; Lin, R.; Elf, S.; Ji, Q.; Zhao, L.; Jin, L.; Seo, J.H.; Shan, C.; Arbiser, J.L.; et al. Metabolic Rewiring by Oncogenic BRAF V600E Links Ketogenesis Pathway to BRAF-MEK1 Signaling. Mol. Cell 2015, 59, 345–358. [Google Scholar] [CrossRef] [Green Version]
- Maddox, J.; Shakya, A.; South, S.; Shelton, D.; Andersen, J.N.; Chidester, S.; Kang, J.; Gligorich, K.M.; Jones, D.A.; Spangrude, G.J.; et al. Transcription Factor Oct1 Is a Somatic and Cancer Stem Cell Determinant. PLoS Genet. 2012, 8, e1003048. [Google Scholar] [CrossRef]
- Sharpe, D.J.; Orr, K.S.; Moran, M.; White, S.J.; McQuaid, S.; Lappin, T.R.; Thompson, A.; James, J.A. POU2F1 activity regulates HOXD10 and HOXD11 promoting a proliferative and invasive phenotype in Head and Neck cancer. Oncotarget 2014, 5, 8803–8815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang-Verslues, W.W.; Chang, P.-H.; Jeng, Y.-M.; Kuo, W.-H.; Chiang, P.-H.; Chang, Y.-C.; Hsieh, T.-H.; Su, F.-Y.; Lin, L.-C.; Abbondante, S.; et al. Loss of corepressor PER2 under hypoxia up-regulates OCT1-mediated EMT gene expression and enhances tumor malignancy. Proc. Natl. Acad. Sci. USA 2013, 110, 12331–12336. [Google Scholar] [CrossRef] [Green Version]
- Obinata, D.; Takayama, K.-I.; Urano, T.; Murata, T.; Kumagai, J.; Fujimura, T.; Ikeda, K.; Horie-Inoue, K.; Homma, Y.; Ouchi, Y.; et al. Oct1 regulates cell growth of LNCaP cells and is a prognostic factor for prostate cancer. Int. J. Cancer 2011, 130, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Kong, X.; Deng, N.; Tan, P.; Chen, H.; Wang, J.; Li, Z.; Hu, Y.; Zou, W.; Xu, J.; et al. OCT1 is a determinant of synbindin-related ERK signalling with independent prognostic significance in gastric cancer. Gut 2014, 64, 37–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.-P.; Song, G.-H.; Chen, J.; Xiao, C.; Li, C.; Zhong, L.; Sun, X.; Wang, Z.-W.; Deng, G.-L.; Yu, F.-D.; et al. Elevated OCT1 participates in colon tumorigenesis and independently predicts poor prognoses of colorectal cancer patients. Tumor Biol. 2015, 37, 3247–3255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Y.; Huang, H.; Chen, M.; Huang, J.; Wu, Q.; Yan, G.-R.; Chen, D. POU2F1 over-expression correlates with poor prognoses and promotes cell growth and epithelial-to-mesenchymal transition in hepatocellular carcinoma. Oncotarget 2017, 8, 44082–44095. [Google Scholar] [CrossRef]
- Györffy, B.; Lanczky, A.; Eklund, A.C.; Denkert, C.; Budczies, J.; Li, Q.; Szallasi, Z. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1809 patients. Breast Cancer Res. Treat. 2009, 123, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, D.R.; Yu, J.; Shanker, K.; Deshpande, N.; Varambally, R.; Ghosh, D.; Barrette, T.; Pander, A.; Chinnaiyan, A.M. ONCOMINE: A Cancer Microarray Database and Integrated Data-Mining Platform. Neoplasia 2004, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T. Condensins: Universal organizers of chromosomes with diverse functions. Genes Dev. 2012, 26, 1659–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schleiffer, A.; Kaitna, S.; Maurer-Stroh, S.; Glotzer, M.; Nasmyth, K.; Eisenhaber, F. Kleisins: A Superfamily of Bacterial and Eukaryotic SMC Protein Partners. Mol. Cell 2003, 11, 571–575. [Google Scholar] [CrossRef]
- Hosey, A.M.; Gorski, J.J.; Murray, M.M.; Quinn, J.E.; Chung, W.Y.; Stewart, G.E.; James, C.R.; Farragher, S.M.; Mulligan, J.M.; Scott, A.N.; et al. Molecular Basis for Estrogen Receptor Deficiency in BRCA1-Linked Breast Cancer. J. Natl. Cancer Inst. 2007, 99, 1683–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harte, M.T.; O’Brien, G.J.; Ryan, N.M.; Gorski, J.J.; Savage, K.; Crawford, N.T.; Mullan, P.B.; Harkin, D.P. BRD7, a Subunit of SWI/SNF Complexes, Binds Directly to BRCA1 and Regulates BRCA1-Dependent Transcription. Cancer Res. 2010, 70, 2538–2547. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Jin, T.; Dai, X.; Xu, J. Lentivirus-mediated knockdown of CEP55 suppresses cell proliferation of breast cancer cells. Biosci. Trends 2016, 10, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Shah, K.; Wilson, E.A.; Malla, R.; Elford, H.L.; Faridi, J.S. Targeting Ribonucleotide Reductase M2 and NF-κB Activation with Didox to Circumvent Tamoxifen Resistance in Breast Cancer. Mol. Cancer Ther. 2015, 14, 2411–2421. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.; Takano, A.; Thang, P.M.; Tsevegjav, B.; Zhu, M.; Yokose, T.; Yamashita, T.; Miyagi, Y.; Daigo, Y. Characterization of KIF20A as a prognostic biomarker and therapeutic target for different subtypes of breast cancer. Int. J. Oncol. 2020, 57, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Phan, N.N.; Wang, C.-Y.; Li, K.-L.; Chen, C.-F.; Chiao, C.-C.; Yu, H.-G.; Huang, P.-L.; Lin, Y.-C. Distinct expression of CDCA3, CDCA5, and CDCA8 leads to shorter relapse free survival in breast cancer patient. Oncotarget 2018, 9, 6977–6992. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Huang, S.; Wang, H.; Xie, R.; Zhang, L.; Zhou, Q.; He, X.; Ju, W. Non-SMC condensin I complex subunit H enhances proliferation, migration, and invasion of hepatocellular carcinoma. Mol. Carcinog. 2019, 58, 2266–2275. [Google Scholar] [CrossRef]
- Yin, L.; Jiang, L.-P.; Shen, Q.-S.; Xiong, Q.-X.; Zhuo, X.; Zhang, L.-L.; Yu, H.-J.; Guo, X.; Luo, Y.; Dong, J.; et al. NCAPH plays important roles in human colon cancer. Cell Death Dis. 2017, 8, e2680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Qiao, X.; Cooper, T.; Pan, W.; Liu, L.; Hayball, J.; Lin, J.; Cui, X.; Zhou, Y.; Zhang, S.; et al. HPV E7-mediated NCAPH ectopic expression regulates the carcinogenesis of cervical carcinoma via PI3K/AKT/SGK pathway. Cell Death Dis. 2020, 11, 1049. [Google Scholar] [CrossRef]
- Kim, B.; Kim, S.W.; Lim, J.-Y.; Park, S.-J. NCAPH Is Required for Proliferation, Migration and Invasion of Non-small-cell Lung Cancer Cells. Anticancer. Res. 2020, 40, 3239–3246. [Google Scholar] [CrossRef]
- Shimomura, H.; Sasahira, T.; Nakashima, C.; Kurihara-Shimomura, M.; Kirita, T. Non-SMC Condensin I Complex Subunit H (NCAPH) Is Associated with Lymphangiogenesis and Drug Resistance in Oral Squamous Cell Carcinoma. J. Clin. Med. 2019, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Paulson, J.R.; Bakhrebah, M.; Kim, J.H.; Nowell, C.; Kalitsis, P.; Hudson, D.F. Condensin I and II behaviour in interphase nuclei and cells undergoing premature chromosome condensation. Chromosom. Res. 2016, 24, 243–269. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, X.; Li, G. Explore a novel function of human condensins in cellular senescence. Cell Biosci. 2020, 10, 147. [Google Scholar] [CrossRef]
- Sun, D.-P.; Lin, C.-C.; Hung, S.-T.; Kuang, Y.-Y.; Hseu, Y.-C.; Fang, C.-L.; Lin, K.-Y. Aberrant Expression of NCAPG is Associated with Prognosis and Progression of Gastric Cancer. Cancer Manag. Res. 2020, 12, 7837–7846. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, H.; Han, Y.; Zhu, M.; Song, Z.; Zhan, D.; Jia, J. NCAPG Induces Cell Proliferation in Cardia Adenocarcinoma via PI3K/AKT Signaling Pathway. OncoTargets Ther. 2020, 13, 11315–11326. [Google Scholar] [CrossRef] [PubMed]
- NCCN Clinical Practice Guidelines in Oncology Breast Cancer, Version 8. 2021. Available online: https://www.nccn.org/professionals/physician_gls/PDF/breast.pdf (accessed on 22 September 2021).
- Brierley, J.D.; Gospodarowicz, M.K.; Wittekind, C. (Eds.) TNM Classification of Malignant Tumours, 8th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2017. [Google Scholar]
- Kawabata, H.; Azuma, K.; Ikeda, K.; Sugitani, I.; Kinowaki, K.; Fujii, T.; Osaki, A.; Saeki, T.; Horie-Inoue, K.; Inoue, S. TRIM44 Is a Poor Prognostic Factor for Breast Cancer Patients as a Modulator of NF-κB Signaling. Int. J. Mol. Sci. 2017, 18, 1931. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, H.; Ando, Y.; Nishio, M.; Zhang, Z.; Hamaguchi, M.; Mita, K.; Kobayashi, S.; Fuji, Y.; Iwase, H. Immunohistochemical evaluation of hormone receptor status for predicting response to endocrine therapy in metastatic breast cancer. Breast Cancer 2006, 13, 74–83. [Google Scholar] [CrossRef]
- Hammond, M.E.H.; Hayes, D.F.; Dowsett, M.; Allred, D.C.; Hagerty, K.L.; Badve, S.; Fitzgibbons, P.L.; Francis, G.; Goldstein, N.S.; Hayes, M.; et al. American Society of Clinical Oncology/College of American Pathologists Guideline Recommendations for Immunohistochemical Testing of Estrogen and Progesterone Receptors in Breast Cancer. J. Clin. Oncol. 2010, 28, 2784–2795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolff, A.; Hammond, M.E.H.; Hicks, D.G.; Dowsett, M.; McShane, L.M.; Allison, K.H.; Allred, D.C.; Bartlett, J.M.; Bilous, M.; Fitzgibbons, P.; et al. Recommendations for Human Epidermal Growth Factor Receptor 2 Testing in Breast Cancer: American Society of Clinical Oncology/College of American Pathologists Clinical Practice Guideline Update. J. Clin. Oncol. 2013, 31, 3997–4013. [Google Scholar] [CrossRef]
- Ikeda, K.; Horie-Inoue, K.; Ueno, T.; Suzuki, T.; Sato, W.; Shigekawa, T.; Osaki, A.; Saeki, T.; Berezikov, E.; Mano, H.; et al. miR-378a-3p modulates tamoxifen sensitivity in breast cancer MCF-7 cells through targeting GOLT1A. Sci. Rep. 2015, 5, 13170. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Gene Expression Omnibus. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/geo/ (accessed on 25 October 2021).




{kind=link}
{kind=link}
{kind=link}
| OCT1 Status | p-Value | |||
|---|---|---|---|---|
| Positive IR (n = 48) | Negative IR (n = 60) | |||
| Age | ≤50 years old | 26 | 27 | 0.344 |
| >50 years old | 22 | 33 | ||
| Stage | I | 17 | 25 | 0.508 |
| II, III | 31 | 35 | ||
| Invasive tumor size | ≤20 mm | 26 | 34 | 0.795 |
| >20 mm | 22 | 26 | ||
| Lymph node status | Positive | 21 | 22 | 0.455 |
| Negative | 27 | 38 | ||
| Nuclear grade | 1 | 24 | 36 | 0.299 |
| 2, 3 | 24 | 24 | ||
| PgR status | Positive | 40 | 52 | 0.628 |
| Negative | 8 | 8 | ||
| HER2 status | Positive | 6 | 8 | 0.898 |
| Negative | 42 | 52 | ||
| Variables | Univariate | Multivariate | ||
|---|---|---|---|---|
| p-Value | Hazard Ratio | 95% CI | p-Value | |
| Age (≤50 vs. >50 years old) | 0.186 | |||
| Invasivetumor size (≤20 vs. >20 mm) | 0.012 | 2.39 | 0.99–5.75 | 0.052 |
| Lymph node status (+ vs. −) | 0.014 | 2.09 | 0.89–4.91 | 0.091 |
| Nuclear grade (1 vs. 2, 3) | 0.007 | 2.21 | 0.87–5.62 | 0.095 |
| PgR status (+ vs. −) | 0.628 | |||
| HER2 status (+ vs. −) | 0.656 | |||
| OCT1 IR (+ vs. −) | 0.024 | 2.38 | 1.03–5.52 | 0.043 |
| NCAPH Status | p-Value | |||
|---|---|---|---|---|
| Positive IR (n = 63) | Negative IR (n = 45) | |||
| Age | ≤50 years old | 32 | 21 | 0.344 |
| >50 years old | 31 | 24 | ||
| Stage | I | 21 | 21 | 0.161 |
| II, III | 42 | 24 | ||
| Invasive tumor size | ≤20 mm | 35 | 25 | 1.000 |
| >20 mm | 28 | 20 | ||
| Lymph node status | Positive | 28 | 15 | 0.245 |
| Negative | 35 | 30 | ||
| Nuclear grade | 1 | 31 | 29 | 0.116 |
| 2, 3 | 32 | 16 | ||
| PgR status | Positive | 51 | 41 | 0.143 |
| Negative | 12 | 4 | ||
| HER2 status | Positive | 9 | 5 | 0.628 |
| Negative | 54 | 40 | ||
| OCT1 status | Positive IR | 42 | 6 | <0.001 |
| Negative IR | 21 | 39 | ||
| Variables | Univariate | Multivariate | ||
|---|---|---|---|---|
| p-Value | Hazard Ratio | 95% CI | p-Value | |
| Age (≤50 vs. >50 years old) | 0.186 | |||
| Invasive tumor size (≤20 vs. >20 mm) | 0.012 | 2.76 | 1.15–6.67 | 0.024 |
| Lymph node status (+ vs. −) | 0.014 | 2.22 | 0.95–5.19 | 0.066 |
| Nuclear grade (1 vs. 2, 3) | 0.007 | 2.15 | 0.85–5.44 | 0.105 |
| PgR status (+ vs. −) | 0.628 | |||
| HER2 status (+ vs. −) | 0.656 | |||
| NCAPH IR (+ vs. −) | 0.049 | 2.61 | 1.01–6.78 | 0.048 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogura, T.; Azuma, K.; Sato, J.; Kinowaki, K.; Takayama, K.-I.; Takeiwa, T.; Kawabata, H.; Inoue, S. OCT1 Is a Poor Prognostic Factor for Breast Cancer Patients and Promotes Cell Proliferation via Inducing NCAPH. Int. J. Mol. Sci. 2021, 22, 11505. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111505
Ogura T, Azuma K, Sato J, Kinowaki K, Takayama K-I, Takeiwa T, Kawabata H, Inoue S. OCT1 Is a Poor Prognostic Factor for Breast Cancer Patients and Promotes Cell Proliferation via Inducing NCAPH. International Journal of Molecular Sciences. 2021; 22(21):11505. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111505
Chicago/Turabian StyleOgura, Takuya, Kotaro Azuma, Junichiro Sato, Keiichi Kinowaki, Ken-Ichi Takayama, Toshihiko Takeiwa, Hidetaka Kawabata, and Satoshi Inoue. 2021. "OCT1 Is a Poor Prognostic Factor for Breast Cancer Patients and Promotes Cell Proliferation via Inducing NCAPH" International Journal of Molecular Sciences 22, no. 21: 11505. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111505