
AR-12 Has a Bactericidal Activity and a Synergistic Effect with Gentamicin against Group A Streptococcus

Abstract
:1. Introduction
2. Results
2.1. Inhibition of In Vitro GAS Growth by AR-12
2.2. AR-12 Reduced the Nucleic Acid and Protein Content of Group A Streptococcus
2.3. AR-12 Treatment Reduced the Production of Heat Shock Proteins and Exotoxins by GAS
2.4. The Analysis of Scanning Electron Microscopy
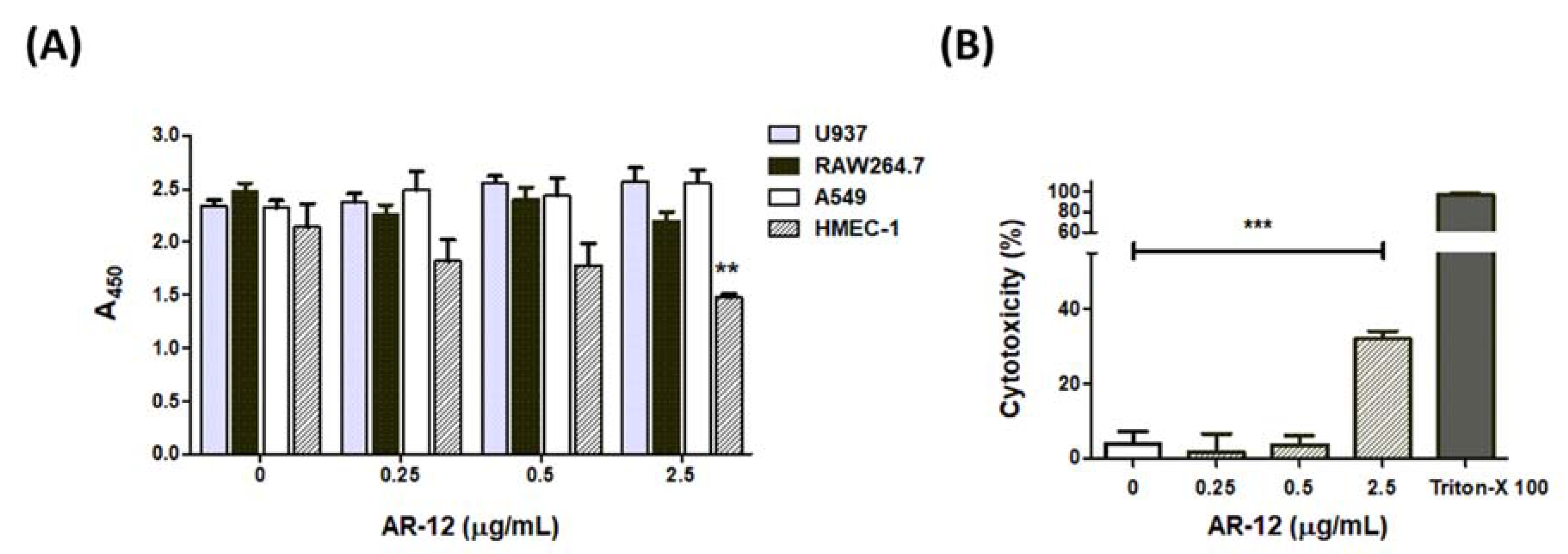
2.5. The Cell Effects of AR-12
2.6. AR-12 and Gentamicin Had a Synergistic Effect against Group A Streptococcus
2.7. The Survival Rate in the Mouse Model after Combination Treatment
3. Discussion
4. Materials and Methods
4.1. Bacteria
4.2. Bacterial Growth Curves Assay and Evaluation of Antibacterial Concentrations of AR-12 and Gentamicin
4.3. Cell Lines and Cell Culture
4.4. Flow Cytometry Assay
4.5. WST-1 Assay
4.6. LDH Assay
4.7. Western Blotting
4.8. Scanning Electron Microscopy Analysis
4.9. Evaluation of Drug Effects Using the Air Pouch GAS Infection Model
4.10. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lamagni, T.L.; Darenberg, J.; Luca-Harari, B.; Siljander, T.; Efstratiou, A.; Henriques-Normark, B.; Vuopio-Varkila, J.; Bouvet, A.; Creti, R.; Ekelund, K.; et al. Epidemiology of Severe Streptococcus pyogenes Disease in Europe. J. Clin. Microbiol. 2008, 46, 2359–2367. [Google Scholar] [CrossRef] [Green Version]
- Lappin, E.; Ferguson, A. Gram-positive toxic shock syndromes. Lancet Infect. Dis. 2009, 9, 281–290. [Google Scholar] [CrossRef]
- Schmitz, M.; Roux, X.; Huttner, B.; Pugin, J. Streptococcal toxic shock syndrome in the intensive care unit. Ann. Intensiv. Care 2018, 8, 88. [Google Scholar] [CrossRef] [PubMed]
- Steer, A.C.; Lamagni, T.; Curtis, N.; Carapetis, J. Invasive Group A Streptococcal Disease. Drugs 2012, 72, 1213–1227. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-N.; Chang, L.-L.; Lai, C.-H.; Lin, H.-H.; Chen, Y.-H. Emergence of Streptococcus pyogenes emm102 Causing Toxic Shock Syndrome in Southern Taiwan during 2005–2012. PLoS ONE 2013, 8, e81700. [Google Scholar] [CrossRef] [PubMed]
- Muhtarova, A.A.; Gergova, R.T.; Mitov, I.G. Distribution of macrolide resistance mechanisms in Bulgarian clinical isolates of Streptococcus pyogenes during the years of 2013–2016. J. Glob. Antimicrob. Resist. 2017, 10, 238–242. [Google Scholar] [CrossRef]
- Abraham, T.; Sistla, S. Trends in Antimicrobial Resistance Patterns of Group A Streptococci, Molecular Basis and Implications. Indian J. Med. Microbiol. 2018, 36, 186–191. [Google Scholar] [CrossRef]
- Mateo, J.; De Bono, J.S.; Ramanathan, R.K.; Lustberg, M.B.; Zivi, A.; Basset, D.; Ng, M.; Young, A.-M.; Garrett, M.; Decordova, S.; et al. A first-in-human phase I trial of AR-12, a PDK-1 inhibitor, in patients with advanced solid tumors. J. Clin. Oncol. 2013, 31, 2608. [Google Scholar] [CrossRef]
- Booth, L.; Roberts, J.L.; Cash, D.R.; Tavallai, S.; Jean, S.; Fidanza, A.; Cruz-Luna, T.; Siembiba, P.; Cycon, K.A.; Cornelissen, C.N.; et al. GRP78/BiP/HSPA5/Dna K is a universal therapeutic target for human disease. J. Cell. Physiol. 2015, 230, 1661–1676. [Google Scholar] [CrossRef] [Green Version]
- Porchia, L.M.; Guerra, M.; Wang, Y.-C.; Zhang, Y.; Espinosa, A.V.; Shinohara, M.; Kulp, S.K.; Kirschner, L.S.; Saji, M.; Chen, C.-S.; et al. 2-Amino-N-{4-[5-(2-phenanthrenyl)-3-(trifluoromethyl)-1H-pyrazol-1-yl]-phenyl} Acetamide (OSU-03012), a Celecoxib Derivative, Directly Targets p21-Activated Kinase. Mol. Pharmacol. 2007, 72, 1124–1131. [Google Scholar] [CrossRef]
- Zhang, S.; Suvannasankha, A.; Crean, C.D.; White, V.L.; Johnson, A.; Chen, C.-S.; Farag, S.S. OSU-03012, a Novel Celecoxib Derivative, Is Cytotoxic to Myeloma Cells and Acts through Multiple Mechanisms. Clin. Cancer Res. 2007, 13, 4750–4758. [Google Scholar] [CrossRef] [Green Version]
- Bai, L.-Y.; Weng, J.-R.; Tsai, C.-H.; Sargeant, A.; Lin, C.-W.; Chiu, C.-F. OSU-03012 sensitizes TIB-196 myeloma cells to imatinib mesylate via AMP-activated protein kinase and STAT3 pathways. Leuk. Res. 2010, 34, 816–820. [Google Scholar] [CrossRef]
- Ma, Y.; Mccarty, S.K.; Kapuriya, N.P.; Brendel, V.J.; Wang, C.; Zhang, X.; Jarjoura, D.; Saji, M.; Chen, C.-S.; Ringel, M.D. Development of p21 Activated Kinase-Targeted Multikinase Inhibitors That Inhibit Thyroid Cancer Cell Migration. J. Clin. Endocrinol. Metab. 2013, 98, E1314–E1322. [Google Scholar] [CrossRef] [Green Version]
- Booth, L.; Cazanave, S.C.; Hamed, H.A.; Yacoub, A.; Ogretmen, B.; Chen, C.-S.; Grant, S.; Dent, P. OSU-03012 suppresses GRP78/BiP expression that causes PERK-dependent increases in tumor cell killing. Cancer Biol. Ther. 2012, 13, 224–236. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.; Yeh, P.Y.; Lu, Y.-S.; Hsu, C.-H.; Chen, K.-F.; Lee, W.-C.; Feng, W.-C.; Chen, C.-S.; Kuo, M.-L.; Cheng, A.-L. OSU-03012, a Novel Celecoxib Derivative, Induces Reactive Oxygen Species–Related Autophagy in Hepatocellular Carcinoma. Cancer Res. 2008, 68, 9348–9357. [Google Scholar] [CrossRef] [Green Version]
- Booth, L.; Roberts, J.L.; Cruickshanks, N.; Grant, S.; Poklepovic, A.; Dent, P. Regulation of OSU-03012 Toxicity by ER Stress Proteins and ER Stress–Inducing Drugs. Mol. Cancer Ther. 2014, 13, 2384–2398. [Google Scholar] [CrossRef] [Green Version]
- Baxter, B.K.; DiDone, L.; Ogu, D.; Schor, S.; Krysan, D.J. Identification, in Vitro Activity and Mode of Action of Phosphoinositide-Dependent-1 Kinase Inhibitors as Antifungal Molecules. ACS Chem. Biol. 2011, 6, 502–510. [Google Scholar] [CrossRef] [Green Version]
- Koselny, K.; Green, J.; Favazzo, L.; Glazier, V.E.; DiDone, L.; Ransford, S.; Krysan, D.J. Antitumor/Antifungal Celecoxib Derivative AR-12 is a Non-Nucleoside Inhibitor of the ANL-Family Adenylating Enzyme Acetyl CoA Synthetase. ACS Infect. Dis. 2016, 2, 268–280. [Google Scholar] [CrossRef]
- Mohr, E.L.; McMullan, L.K.; Lo, M.K.; Spengler, J.R.; Bergeron, É.; Albariño, C.G.; Shrivastava-Ranjan, P.; Chiang, C.-F.; Nichol, S.T.; Spiropoulou, C.F.; et al. Inhibitors of cellular kinases with broad-spectrum antiviral activity for hemorrhagic fever viruses. Antivir. Res. 2015, 120, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-H.; Chen, C.-C.; Lin, Y.-S.; Chang, P.-C.; Lu, Z.-Y.; Lin, C.-F.; Chen, C.-L.; Chang, C.-P. AR-12 suppresses dengue virus replication by down-regulation of PI3K/AKT and GRP78. Antivir. Res. 2017, 142, 158–168. [Google Scholar] [CrossRef]
- Chan, J.F.-W.; Zhu, Z.; Chu, H.; Yuan, S.; Chik, K.K.-H.; Chan, C.C.-S.; Poon, V.K.-M.; Yip, C.C.-Y.; Zhang, X.; Tsang, J.O.-L.; et al. The celecoxib derivative kinase inhibitor AR-12 (OSU-03012) inhibits Zika virus via down-regulation of the PI3K/Akt pathway and protects Zika virus-infected A129 mice: A host-targeting treatment strategy. Antivir. Res. 2018, 160, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.-C.; Kulp, S.K.; Soni, S.; Wang, D.; Gunn, J.S.; Schlesinger, L.S.; Chen, C.-S. Eradication of Intracellular Salmonella enterica Serovar Typhimurium with a Small-Molecule, Host Cell-Directed Agent. Antimicrob. Agents Chemother. 2009, 53, 5236–5244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, H.-C.; Soni, S.; Kulp, S.K.; Curry, H.; Wang, D.; Gunn, J.S.; Schlesinger, L.S.; Chen, C.-S. Eradication of intracellular Francisella tularensis in THP-1 human macrophages with a novel autophagy inducing agent. J. Biomed. Sci. 2009, 16, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collier, M.; Peine, K.; Gautam, S.; Oghumu, S.; Varikuti, S.; Borteh, H.; Papenfuss, T.; Sataoskar, A.; Bachelder, E.; Ainslie, K. Host-mediated Leishmania donovani treatment using AR-12 encapsulated in acetalated dextran microparticles. Int. J. Pharm. 2016, 499, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zou, Y.; Guo, Q.; Chen, J.; Xu, L.; Wan, X.; Zhang, Z.; Li, B.; Chu, H. AR-12 Exhibits Direct and Host-Targeted Antibacterial Activity toward Mycobacterium abscessus. Antimicrob. Agents Chemother. 2020, 64, e00236-20. [Google Scholar] [CrossRef]
- Thangamani, S.; Younis, W.; Seleem, M. Repurposing celecoxib as a topical antimicrobial agent. Front. Microbiol. 2015, 6, 750. [Google Scholar] [CrossRef] [Green Version]
- Chiu, H.-C.; Lee, S.-L.; Kapuriya, N.; Wang, D.; Chen, Y.-R.; Yu, S.-L.; Kulp, S.K.; Teng, L.-J.; Chen, C.-S. Development of novel antibacterial agents against methicillin-resistant Staphylococcus aureus. Bioorg. Med. Chem. 2012, 20, 4653–4660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnamurthy, M.; Lemmon, M.M.; Falcinelli, E.M.; Sandy, R.A.; Dootz, J.N.; Mott, T.M.; Rajamani, S.; Schaecher, K.E.; Duplantier, A.J.; Panchal, R.G. Enhancing the antibacterial activity of polymyxins using a nonantibiotic drug. Infect. Drug Resist. 2019, 12, 1393–1405. [Google Scholar] [CrossRef] [Green Version]
- Kilic, S.; Dizbay, M.; Hizel, K.; Arman, D. In vitro synergistic activity of antibiotic combinations against Brucella melitensis using E-test methodology. Braz. J. Microbiol. 2008, 39, 233–237. [Google Scholar] [CrossRef] [Green Version]
- Meletiadis, J.; Pournaras, S.; Roilides, E.; Walsh, T.J. Defining Fractional Inhibitory Concentration Index Cutoffs for Additive Interactions Based on Self-Drug Additive Combinations, Monte Carlo Simulation Analysis, and In Vitro—In Vivo Correlation Data for Antifungal Drug Combinations against Aspergillus fumigatus. Antimicrob. Agents Chemother. 2010, 54, 602–609. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.-F.; Tsao, N.; Hsieh, I.-C.; Lin, Y.-S.; Wu, J.-J.; Hung, Y.-T. Immunization with a streptococcal multiple-epitope recombinant protein protects mice against invasive group A streptococcal infection. PLoS ONE 2017, 12, e0174464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booth, L.; Roberts, J.L.; Dent, P. HSPA5/Dna K May Be a Useful Target for Human Disease Therapies. DNA Cell Biol. 2015, 34, 153–158. [Google Scholar] [CrossRef]
- Lemos, J.; Luzardo, Y.; Burne, R.A. Physiologic Effects of Forced Down-Regulation of dnaK and groEL Expression in Streptococcus mutans. J. Bacteriol. 2007, 189, 1582–1588. [Google Scholar] [CrossRef] [Green Version]
- Uhlmann, J.; Rohde, M.; Siemens, N.; Kreikemeyer, B.; Bergman, P.; Johansson, L.; Norrby-Teglund, A. LL-37 Triggers Formation of Streptococcus pyogenes Extracellular Vesicle-Like Structures with Immune Stimulatory Properties. J. Innate Immun. 2015, 8, 243–257. [Google Scholar] [CrossRef] [Green Version]
- Goldfless, S.J.; Morag, A.S.; Belisle, K.A.; Sutera, V.A.; Lovett, S.T. DNA Repeat Rearrangements Mediated by DnaK-Dependent Replication Fork Repair. Mol. Cell 2006, 21, 595–604. [Google Scholar] [CrossRef]
- Kalle, A.M.; Rizvi, A. Inhibition of Bacterial Multidrug Resistance by Celecoxib, a Cyclooxygenase-2 Inhibitor. Antimicrob. Agents Chemother. 2010, 55, 439–442. [Google Scholar] [CrossRef] [Green Version]
- Zheng, P.X.; Chung, K.-T.; Chiang-Ni, C.; Wang, S.-Y.; Tsai, P.-J.; Chuang, W.-J.; Lin, Y.-S.; Liu, C.-C.; Wu, J.-J. Complete Genome Sequence of emm 1 Streptococcus pyogenes A20, a Strain with an Intact Two-Component System, CovRS, Isolated from a Patient with Necrotizing Fasciitis. Genome Announc. 2013, 1, e00149-12. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GAS Strain | NZ131 (M49) | A20 (M1) |
|---|---|---|
| MIC ranges of Gm (μg/mL) | 1–5 | 5–25 |
| MIC ranges of AR-12 (μg/mL) | 1–2.5 | 1–2.5 |
| MIC90 of Gm (μg/mL) | 5 | 25 |
| MIC90 of AR-12 (μg/mL) | 2.5 | 2.5 |
| FICGm | 0.25 | 0.2 |
| FICAR-12 | 0.05 | 0.1 |
| ΣFIC index | 0.3 | 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsao, N.; Chang, Y.-C.; Hsieh, S.-Y.; Li, T.-C.; Chiu, C.-C.; Yu, H.-H.; Hsu, T.-C.; Kuo, C.-F. AR-12 Has a Bactericidal Activity and a Synergistic Effect with Gentamicin against Group A Streptococcus. Int. J. Mol. Sci. 2021, 22, 11617. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111617
Tsao N, Chang Y-C, Hsieh S-Y, Li T-C, Chiu C-C, Yu H-H, Hsu T-C, Kuo C-F. AR-12 Has a Bactericidal Activity and a Synergistic Effect with Gentamicin against Group A Streptococcus. International Journal of Molecular Sciences. 2021; 22(21):11617. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111617
Chicago/Turabian StyleTsao, Nina, Ya-Chu Chang, Sung-Yuan Hsieh, Tang-Chi Li, Ching-Chen Chiu, Hai-Han Yu, Tzu-Ching Hsu, and Chih-Feng Kuo. 2021. "AR-12 Has a Bactericidal Activity and a Synergistic Effect with Gentamicin against Group A Streptococcus" International Journal of Molecular Sciences 22, no. 21: 11617. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111617