
Seed Dormancy and Pre-Harvest Sprouting in Rice—An Updated Overview

 ,
,  ,
,  ,
,  ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
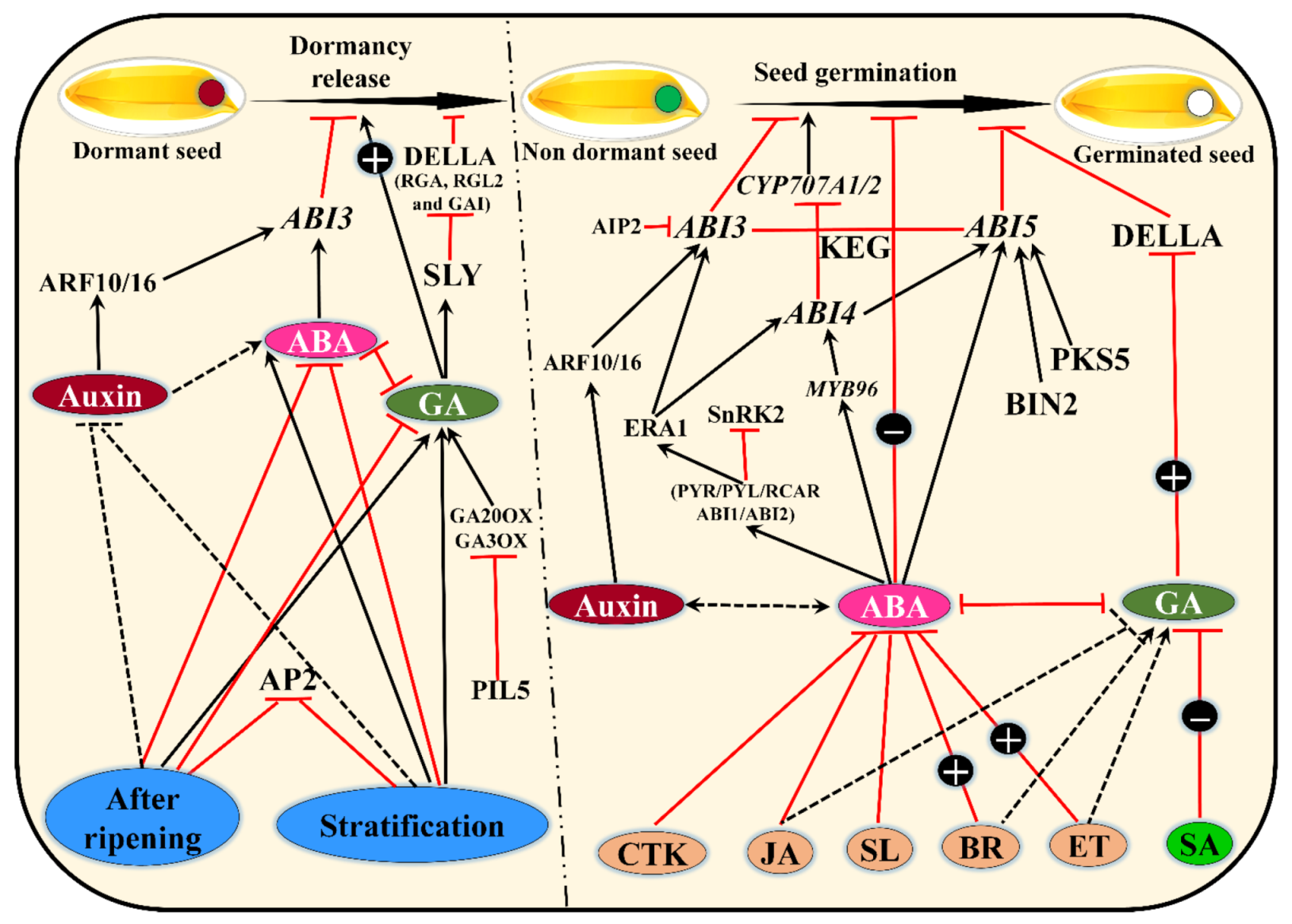
2. Seed Dormancy and Germination—A Game of Hormones
3. Pre-Harvest Sprouting
4. Factors Affecting Seed Dormancy and Pre-Harvest Sprouting
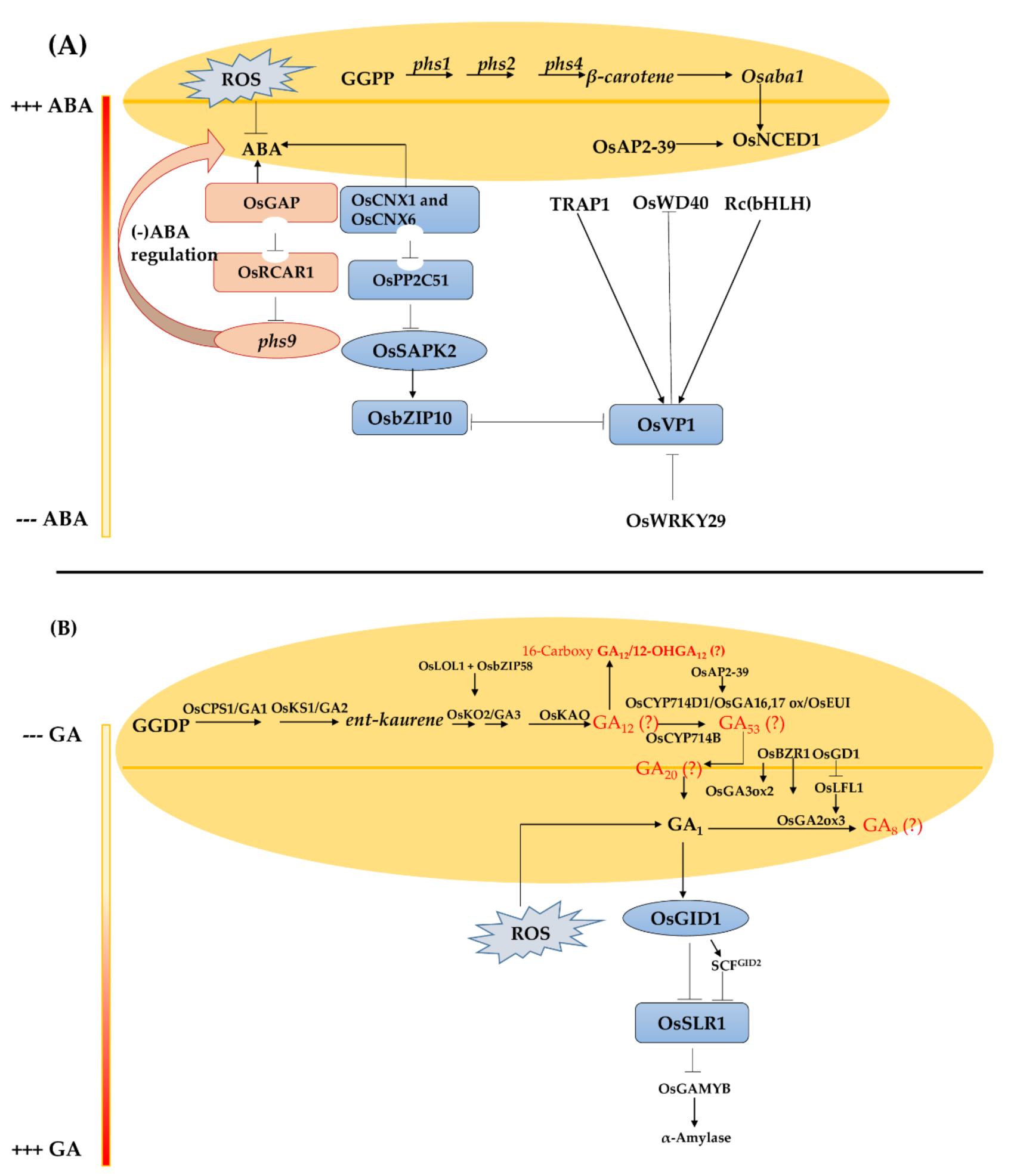
5. Pre-Harvest Sprouting in Rice
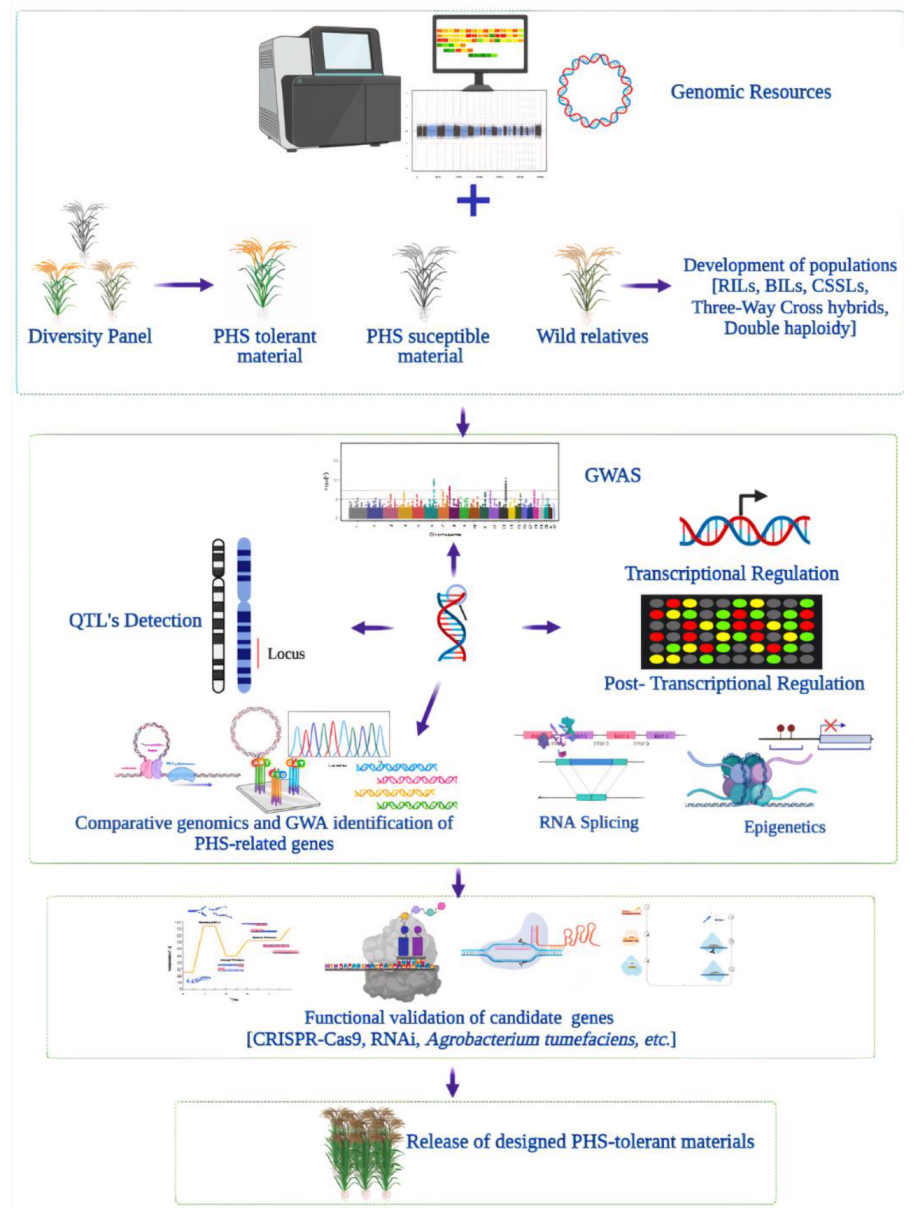
6. Omics Approaches for Pre-Harvest Sprouting in Rice
7. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular Aspects of Seed Dormancy. Annu. Rev. Plant Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Wang, W.; Lyu, Y.; Wu, Y.; Huang, P.; Hu, S.; Wei, X.; Jiao, G.; Sheng, Z.; Tang, S.; et al. OsVP1 activates Sdr4 expression to control rice seed dormancy via the ABA signaling pathway. Crop J. 2020, 9, 68–78. [Google Scholar] [CrossRef]
- He, D.; Yang, P. Proteomics of rice seed germination. Front. Plant Sci. 2013, 4, 246. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, M.V.; Barrero, J.M.; Corbineau, F.; Gubler, F.; Benech-Arnold, R.L. Dormancy in cereals (not too much, not so little): About the mechanisms behind this trait. Seed Sci. Res. 2015, 25, 99–119. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Sehgal, S.K.; Li, J.; Lin, M.; Trick, H.N.; Yu, J.; Gill, B.S.; Bai, G. Cloning and characterization of a critical regulator for preharvest sprouting in wheat. Genetics 2013, 195, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Shu, K.; Meng, Y.J.; Shuai, H.W.; Liu, W.G.; Du, J.B.; Liu, J.; Yang, W.Y. Dormancy and germination: How does the crop seed decide? Plant Biol. 2015, 17, 1104–1112. [Google Scholar] [CrossRef]
- Li, C.; Ni, P.; Francki, M.; Hunter, A.; Zhang, Y.; Schibeci, D.; Li, H.; Tarr, A.; Wang, J.; Cakir, M.; et al. Genes controlling seed dormancy and pre-harvest sprouting in a rice-wheat-barley comparison. Funct. Integr. Genom. 2004, 4, 84–93. [Google Scholar] [CrossRef]
- Wang, J.W.; Korkmaz, U.; Guo, M.; Pipatpongpinyo, W.; Gu, X.Y. Pyramiding seed dormancy genes to improve resistance of semi-dwarf varieties to pre-harvest sprouting in rice. Mol. Breed. 2020, 40, 93. [Google Scholar] [CrossRef]
- Liao, Y.; Bai, Q.; Xu, P.; Wu, T.; Guo, D.; Peng, Y.; Zhang, H.; Deng, X.; Chen, X.; Luo, M.; et al. Mutation in Rice Abscisic Acid2 Results in Cell Death, Enhanced Disease-Resistance, Altered Seed Dormancy and Development. Front. Plant Sci. 2018, 9, 405. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.; Abe, F.; Kawahigashi, H.; Nakazono, K.; Tagiri, A.; Matsumoto, T.; Utsugi, S.; Ogawa, T.; Handa, H.; Ishida, H.; et al. A Wheat Homolog of MOTHER OF FT AND TFL1 Acts in the Regulation of Germination. Plant Cell 2011, 23, 3215–3229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentsink, L.; Jowett, J.; Hanhart, C.J.; Koornneef, M. Cloning of DOG1, a quantitative trait locus controlling seed dormancy in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 17042–17047. [Google Scholar] [CrossRef] [Green Version]
- Hattori, T.; Terada, T.; Hamasuna, S.T. Sequence and functional analyses of the rice gene homologous to the maize Vp1. Plant Mol. Biol. 1994, 24, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.-A.; Jeon, Y.-A.; Lee, H.-S.; Hyun, D.Y.; Lee, J.-R.; Lee, M.-C.; Lee, S.-Y.; Ma, K.-H.; Koh, H.-J. New Genetic Loci Associated with Preharvest Sprouting and Its Evaluation Based on the Model Equation in Rice. Front. Plant Sci. 2017, 8, 1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, S. Gibberellin Metabolism and its Regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef]
- Pearce, S.; Huttly, A.K.; Prosser, I.M.; Li, Y.D.; Vaughan, S.P.; Gallova, B.; Patil, A.; Coghill, J.A.; Dubcovsky, J.; Hedden, P.; et al. Heterologous expression and transcript analysis of gibberellin biosynthetic genes of grasses reveals novel functionality in the GA3ox family. BMC Plant Biol. 2015, 15, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falade, K.; Semon, O.; Fadairo, M.; Oladunjoye, O.S.; Orou, K.K. Functional and physico-chemical properties of flours and starches of African rice cultivars. Food Hydrocoll. 2014, 39, 41–50. [Google Scholar] [CrossRef]
- Lee, H.S.; Choi, M.G.; Hwang, W.H.; Jeong, J.H.; Yang, S.Y.; Lee, C.G. Occurrence of rice preharvest sprouting varies greatly depending on past weather conditions during grain filling. Field Crop. Res. 2021, 264, 108087. [Google Scholar] [CrossRef]
- Muthuramalingam, P.; Jeyasri, R.; Kalaiyarasi, D.; Pandian, S.; Krishnan, S.R.; Satish, L.; Ramesh, M. Emerging advances in computational omics tools for systems analysis of gramineae family grass species and their abiotic stress responsive functions. OMICS Based Approach Plant Biotechnol. 2019, 185, 185. [Google Scholar]
- Yang, Y.; Saand, M.A.; Huang, L.; Abdelaal, W.B.; Zhang, J.; Wu, Y.; Li, J.; Sirohi, M.H.; Wang, F. Applications of Multi-Omics Technologies for Crop Improvement. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Omoarelojie, L.O.; Kulkarni, M.G.; Finnie, J.F.; van Staden, J. Smoke-derived cues in the regulation of seed germina-tion: Are Ca2+-dependent signals involved? Plant Growth Regul. 2021, 28, 1–3. [Google Scholar]
- Hilhorst, H.W.M.; Karssen, C.M. Seed dormancy and germination: The role of abscisic acid and gibberellins and the importance of hormone mutants. Plant Growth Regul. 1992, 11, 225–238. [Google Scholar] [CrossRef]
- Sano, N.; Marion-Poll, A. ABA Metabolism and Homeostasis in Seed Dormancy and Germination. Int. J. Mol. Sci. 2021, 22, 5069. [Google Scholar] [CrossRef]
- He, H.; de Souza Vidigal, D.; Snoek, L.B.; Schnabel, S.; Nijveen, H.; Hilhorst, H.; Bentsink, L. Interaction between parental environment and genotype affects plant and seed performance in Arabidopsis. J. Exp. Bot. 2014, 65, 6603–6615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Hou, X. Antagonistic Regulation of ABA and GA in Metabolism and Signaling Pathways. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Lin, C.; Guan, Y.; Sheteiwy, M.S.; Hu, W.; Hu, J. Inhibitory effect of eugenol on seed ger-mination and pre-harvest sprouting of hybrid rice (Oryza sativa L.). Sci. Rep. 2017, 7, 5295. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, H.S.; Benech-Arnold, R.L.; Sanchez, R.A. Hormonal regulation of dormancy in developing sorghum seeds. Plant Physiol. 1997, 113, 149–154. [Google Scholar] [CrossRef] [Green Version]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic Acid: Emergence of a Core Signaling Network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef] [Green Version]
- Carrillo-Barral, N.; Rodríguez-Gacio, M.D.C.; Matilla, A.J. Delay of Germination-1 (DOG1): A Key to Understanding Seed Dormancy. Plants 2020, 9, 480. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Ye, N.; Zhang, J. Glucose-Induced Delay of Seed Germination in Rice is Mediated by the Suppression of ABA Catabolism Rather Than an Enhancement of ABA Biosynthesis. Plant Cell Physiol. 2009, 50, 644–651. [Google Scholar] [CrossRef]
- Suriyasak, C.; Oyama, Y.; Ishida, T.; Mashiguchi, K.; Yamaguchi, S.; Hamaoka, N.; Iwaya-Inoue, M.; Ishibashi, Y. Mechanism of delayed seed germination caused by high temperature during grain filling in rice (Oryza sativa L.). Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Tanamachi, K.; Miyazaki, M.; Matsuo, K.; Suriyasak, C.; Tamada, A.; Matsuyama, K.; Iwaya-Inoue, M.; Ishibashi, Y. Differential responses to high temperature during maturation in heat-stress-tolerant cultivars of Japonica rice. Plant Prod. Sci. 2016, 19, 300–308. [Google Scholar] [CrossRef] [Green Version]
- Perreau, F.; Frey, A.; Effroy-Cuzzi, D.; Savane, P.; Berger, A.; Gissot, L.; Marion-Poll, A. ABSCISIC AC-ID-DEFICIENT4 has an essential function in both cis-violaxanthin and cis-neoxanthin synthesis. Plant Physiol. 2020, 184, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- McGinty, E.M.; Murphy, K.M.; Hauvermale, A.L. Seed Dormancy and Preharvest Sprouting in Quinoa (Chenopodium quinoa Willd). Plants 2021, 10, 458. [Google Scholar] [CrossRef] [PubMed]
- Hauvermale, A.L.; Tuttle, K.M.; Takebayashi, Y.; Seo, M.; Steber, C.M. Loss of Arabidopsis thaliana seed dormancy is associated with increased accumulation of the GID1 GA hormone receptors. Plant Cell Physiol. 2015, 56, 1773–1785. [Google Scholar] [CrossRef] [Green Version]
- Tuttle, K.M.; Martinez, S.A.; Schramm, E.C.; Takebayashi, Y.; Seo, M.; Steber, C.M. Grain dor-mancy loss is associated with changes in ABA and GA sensitivity and hormone accumulation in bread wheat, Triticum aestivum L. Seed Sci. Res. 2015, 25, 179–193. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, M.; Inukai, Y.; Ueguchi-Tanaka, M.; Itoh, H.; Izawa, T.; Kobayashi, Y.; Hattori, T.; Miyao, A.; Hirochika, H.; Ashikari, M.; et al. Loss-of-Function Mutations of the Rice GAMYB Gene Impair α-Amylase Expression in Aleurone and Flower Development. Plant Cell 2003, 16, 33–44. [Google Scholar] [CrossRef] [Green Version]
- Patwa, N.; Penning, B.W. Environmental Impact on Cereal Crop Grain Damage from Preharvest Sprouting and Late Maturity Alpha-Amylase. In Sustainable Agriculture in the Era of Climate Change; Springer: Cham, Switzerland, 2020; pp. 23–41. [Google Scholar] [CrossRef]
- Shu, K.; Liu, X.-D.; Xie, Q.; He, Z.-H. Two Faces of One Seed: Hormonal Regulation of Dormancy and Germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finch-Savage, W.E.; Footitt, S. Seed dormancy cycling and the regulation of dormancy mechanisms to time germination in variable field environments. J. Exp. Bot. 2017, 68, 843–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuan, P.A.; Kumar, R.; Rehal, P.K.; Toora, P.K.; Ayele, B.T. Molecular Mechanisms Underlying Abscisic Acid/Gibberellin Balance in the Control of Seed Dormancy and Germination in Cereals. Front. Plant Sci. 2018, 9, 668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Agriculture Organization of United Nations (FAO). FAO Cereal Supply and Demand Brief. Available online: http://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 13 November 2020).
- Fang, J.; Chai, C.; Qian, Q.; Li, C.; Tang, J.; Sun, L.; Huang, Z.; Guo, X.; Sun, C.; Liu, M.; et al. Mutations of genes in synthesis of the carotenoid precursors of ABA lead to pre-harvest sprouting and photo-oxidation in rice. Plant J. 2008, 54, 177–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olaerts, H.; Courtin, C.M. Impact of preharvest sprouting on endogenous hydrolases and techno-logical quality of wheat and bread: A review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 698–713. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Cao, J.; Jiang, H.; Chang, C.; Zhang, H.-P.; Sheikh, S.W.; Shah, L.; Ma, C. Unraveling Molecular and Genetic Studies of Wheat (Triticum aestivum L.) Resistance against Factors Causing Pre-Harvest Sprouting. Agronomy 2019, 9, 117. [Google Scholar] [CrossRef] [Green Version]
- Tai, L.; Wang, H.-J.; Xu, X.-J.; Sun, W.-H.; Ju, L.; Liu, W.-T.; Li, W.-Q.; Sun, J.; Chen, K.-M. Pre-harvest sprouting in cereals: Genetic and biochemical mechanisms. J. Exp. Bot. 2021, 72, 2857–2876. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Wu, J.; Luo, Q.; Li, J.; Zhuang, W.; Xiao, G.; Deng, Q.; Lei, D.; Bai, B. Influence of high natural field tem-perature during grain filling stage on the morphological structure and physico-chemical properties of rice (Oryza sativa L.) starch. Food Chem. 2020, 310, 125718. [Google Scholar] [CrossRef]
- Lee, G.A.; Jeon, Y.A.; Lee, H.S.; Hyun, D.Y.; Lee, J.R.; Lee, M.C.; Lee, S.Y.; Ma, K.H.; Koh, H.J. Variation in pre-harvest sprouting resistance, seed germination and changes in abscisic acid levels during grain development in di-verse rice genetic resources. Plant Genet Resour. 2018, 16, 18–27. [Google Scholar] [CrossRef]
- King, R.; Von Wettstein-Knowles, P. Epicuticular waxes and regulation of ear wetting and pre-harvest sprouting in barley and wheat. Euphytica 2000, 112, 157–166. [Google Scholar] [CrossRef]
- Vetch, J.M.; Stougaard, R.N.; Martin, J.M.; Giroux, M.J. Review: Revealing the genetic mechanisms of pre-harvest sprouting in hexaploid wheat (Triticum aestivum L.). Plant Sci. 2019, 281, 180–185. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy; Springer: New York, NY, USA, 2013. [Google Scholar]
- Farnsworth, E. The Ecology and Physiology of Viviparous and Recalcitrant Seeds. Annu. Rev. Ecol. Syst. 2000, 31, 107–138. [Google Scholar] [CrossRef]
- Hoang, H.H.; Sechet, J.; Bailly, C.; Leymarie, J.; Corbineau, F. Inhibition of germination of dormant barley (Hordeum vulgare L.) grains by blue light as related to oxygen and hormonal regulation. Plant Cell Environ. 2013, 37, 1393–1403. [Google Scholar] [CrossRef] [PubMed]
- Barrero, J.M.; Downie, A.B.; Xu, Q.; Gubler, F. A role for barley CRYPTOCHROME1 in light reg-ulation of grain dormancy and germination. Plant Cell 2014, 26, 1094–1104. [Google Scholar] [CrossRef] [Green Version]
- Dashab, S.; Omidi, H. Effects of hydro-and bio-priming on some physiological and biochemical characteristics of quinoa (Chenopodium quinoa) seedlings under drought stress. Iran. J. Plant Physiol. 2021, 11, 3659–3682. [Google Scholar]
- Nonogaki, H. Seed germination and dormancy: The classic story, new puzzles, and evolution. J. Integr. Plant Biol. 2018, 61, 541–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphey, M.; Kovach, K.; Elnacash, T.; He, H.; Bentsink, L.; Donohue, K. DOG1-imposed dormancy mediates germination responses to temperature cues. Environ. Exp. Bot. 2015, 112, 33–43. [Google Scholar] [CrossRef]
- Née, G.; Xiang, Y.; Soppe, W.J. The release of dormancy, a wake-up call for seeds to germinate. Curr. Opin. Plant Biol. 2017, 35, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, K.; Takeuchi, Y.; Ebana, K.; Miyao, A.; Hirochika, H.; Hara, N.; Ishiyama, K.; Kobayashi, M.; Ban, Y.; Hattori, T.; et al. Molecular cloning of Sdr4, a regulator involved in seed dormancy and domes-tication of rice. Proc. Natl. Acad. Sci. USA 2010, 107, 5792–5797. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Han, Y.; Li, J.; Ding, M.; Li, Y.; Li, X.; Chen, F.; Soppe, W.J.; Liu, Y. Arabidopsis thaliana SEED DORMAN-CY 4-LIKE regulates dormancy and germination by mediating the gibberellin pathway. J. Exp. Bot. 2020, 71, 919–933. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, Y.; Hu, G.; Wang, X.; Chen, H.; Shi, Q.; Xiang, J.; Zhang, Y.; Zhu, D.; Zhang, Y. Reduced bioactive gibberellin content in rice seeds under low temperature leads to decreased sugar consumption and low seed germination rates. Plant Physiol. Biochem. 2018, 133, 1–10. [Google Scholar] [CrossRef]
- Xu, F.; Tang, J.; Gao, S.; Cheng, X.; Du, L.; Chu, C. Control of rice pre-harvest sprouting by glutaredoxin-mediated abscisic acid signaling. Plant J. 2019, 100, 1036–1051. [Google Scholar] [CrossRef] [PubMed]
- Topham, A.T.; Taylor, R.E.; Yan, D.; Nambara, E.; Johnston, I.G.; Bassel, G.W. Temperature vari-ability is inte-grated by a spatially embedded decision-making center to break dormancy in Arabidopsis seeds. Proc. Natl. Acad. Sci. USA 2017, 114, 6629–6634. [Google Scholar] [CrossRef] [Green Version]
- Malalgoda, M.; Ohm, J.-B.; Ransom, J.K.; Howatt, K.; Simsek, S. Effects of Pre-Harvest Glyphosate Application on Spring Wheat Quality Characteristics. Agriculture 2020, 10, 111. [Google Scholar] [CrossRef] [Green Version]
- Cabral, A.L.; Jordan, M.C.; McCartney, C.A.; You, F.M.; Humphreys, D.G.; MacLachlan, R.; Pozniak, C.J. Identification of candidate genes, regions and markers for pre-harvest sprouting resistance in wheat (Triticum aestivum L.). BMC Plant Biol. 2014, 14, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S. Grain dormancy genes responsible for preventing pre-harvest sprouting in barley and wheat. Breed. Sci. 2018, 68, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Henry, R.J.; Battershell, V.G.; Brennan, P.S.; Oono, K. Control of wheat α-amylase using inhibitors from cereals. J. Sci. Food Agric. 1992, 58, 281–284. [Google Scholar] [CrossRef]
- Park, K.B.; Park, R.K. Studies on the viviparous germination of indica X japonica type cultivars in paddy rice. Korean J. Crop Sci. 1984, 29, 15–18. [Google Scholar]
- Zhang, Z.; Chen, J.; Lin, S.; Li, Z.; Cheng, R.; Fang, C.; Chen, H.; Lin, W. Proteomic and phospho-proteomic de-termination of ABA’s effects on grain-filling of Oryza sativa L. Inferior spikelets. Plant Sci. 2012, 185–186, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-S.; Chebotarov, D.; McNally, K.L.; Pede, V.; Setiyono, T.D.; Raquid, R.; Hyun, W.-J.; Jeung, J.-U.; Kohli, A.; Mo, Y. Novel Sources of Pre-Harvest Sprouting Resistance for Japonica Rice Improvement. Plants 2021, 10, 1709. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.Y.; Liu, T.; Feng, J.; Suttle, J.C.; Gibbons, J. The qSD12 underlying gene promotes abscisic acid accumulation in early developing seeds to induce primary dormancy in rice. Plant Mol. Biol. 2010, 73, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fang, J.; Xu, F.; Chu, J.; Yan, C.; Schläppi, M.R.; Wang, Y.; Chu, C. Expression Patterns of ABA and GA Metabolism Genes and Hormone Levels during Rice Seed Development and Imbibition: A Comparison of Dormant and Non-Dormant Rice Cultivars. J. Genet. Genom. 2014, 41, 327–338. [Google Scholar] [CrossRef]
- Debeaujon, I.; Leon-Kloosterziel, K.M.; Koornneef, M. Influence of the testa on seed dormancy, germination, and longevity in Arabidopsis. Plant Physiol. 2000, 122, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.S.; Ann, D.J.; Choi, C.D.; Lee, S.P.; Choi, B.S. Phase of overhead flooding damage during maturation of rice. Korean J. Agric. Meteorol. 2000, 2, 148–155. [Google Scholar]
- Nonogaki, M.; Nonogaki, H. Prevention of preharvest sprouting through hormone engineering and ger-mination recovery by chemical biology. Front. Plant Sci. 2017, 8, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, P.D.T.; Van Vu, B. Improving the pre-harvest sprouting resistance of rice cultivar IR36 using wild rice (Oryza rufipogon) W630. Cereal Res. Commun. 2021, 1–7. [Google Scholar] [CrossRef]
- Li, C.; Zhou, A.; Sang, T. Genetic analysis of rice domestication syndrome with the wild annual species, Oryza nivara. New Phytol. 2006, 170, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.Y.; Foley, M.E.; Horvath, D.P.; Anderson, J.V.; Feng, J.; Zhang, L.; Mowry, C.R.; Ye, H.; Suttle, J.C.; Kadowaki, K.-I. Association between seed dormancy and pericarp color is controlled by a pleiotropic gene that regulates ab-scisic acid and flavonoid synthesis in weedy red rice. Genetics 2011, 189, 1515–1524. [Google Scholar] [CrossRef] [Green Version]
- Viruel, J.; Kantar, M.B.; Gargiulo, R.; Hesketh-Prichard, P.; Leong, N.; Cockel, C.; Forest, F.; Gravendeel, B.; Pérez-Barrales, R.; Leitch, I.J.; et al. Crop wild phylorelatives (CWPs): Phylogenetic distance, cytogenetic compatibility and breeding system data enable estimation of crop wild relative gene pool classification. Bot. J. Linn. Soc. 2020, 195, 1–33. [Google Scholar] [CrossRef]
- Desta, Z.A.; Ortiz, R. Genomic selection: Genome-wide prediction in plant improvement. Trends Plant Sci. 2014, 19, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Hickey, J.M.; Participants, I.G.S.I.C.B.P.W.; Chiurugwi, T.; Mackay, I.; Powell, W. Genomic prediction unifies animal and plant breeding programs to form platforms for biological discovery. Nat. Genet. 2017, 49, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.H.; Choudhary, M.; Kumar, P.; Akram, N.A.; Surekha, C.; Ahmad, P.; Gosal, S.S. Marker-assisted breeding for abiotic stress tolerance in crop plants. In Biotechnologies of Crop. Improvement; Gosal, S.S., Wani, S.H., Eds.; Springer: Berlin, Germany, 2018; Volume 3, pp. 1–23. [Google Scholar]
- Dong, Y.; Tsuzuki, E.; Kamiunten, H.; Terao, H.; Lin, D.; Matsuo, M.; Zheng, Y. Identification of quantitative trait loci associated with pre-harvest sprouting resistance in rice (Oryza sativa L.). Field Crop. Res. 2003, 81, 133–139. [Google Scholar] [CrossRef]
- Jang, S.G.; Lar, S.M.; Zhang, H.; Lee, A.; Lee, J.; Kim, N.; Park, S.; Lee, J.; Ham, T.; Kwon, S. Detection of Whole-Genome Resequencing-Based QTLs Associat-ed with Pre-Harvest Sprouting in Rice (Oryza sativa L.). Plant. Breed. Biotech. 2020, 8, 396–404. [Google Scholar] [CrossRef]
- Mizuno, Y.; Yamanouchi, U.; Hoshino, T.; Nonoue, Y.; Nagata, K.; Fukuoka, S.; Ando, T.; Yano, M.; Sugimoto, K. Genetic dissection of pre-harvest sprouting resistance in an upland rice cultivar. Breed. Sci. 2018, 68, 200–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheon, K.-S.; Won, Y.J.; Jeong, Y.-M.; Lee, Y.-Y.; Kang, D.-Y.; Oh, J.; Oh, H.; Kim, S.L.; Kim, N.; Lee, E.; et al. QTL mapping for pre-harvest sprouting resistance in japonica rice varieties utilizing genome re-sequencing. Mol. Genet. Genom. 2020, 295, 1129–1140. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Sugimoto, K.; Nonoue, Y.; Ono, N.; Matsubara, K.; Yamanouchi, U.; Abe, A.; Takeuchi, Y.; Yano, M. Detection of quantitative trait loci controlling pre-harvest sprouting resistance by using backcrossed populations of japonica rice cultivars. Theor. Appl. Genet. 2010, 120, 1547–1557. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.Y.; Sasaki, T.; Yano, M. Mapping quantitative trait loci controlling seed dormancy and heading date in rice, Oryza sativa L., using backcross inbred lines. Theor. Appl. Genet. 1998, 96, 997–1003. [Google Scholar] [CrossRef]
- Miura, K.; Lin, S.Y.; Yano, M.; Nagamine, T. Mapping Quantitative Trait Loci Controlling Low Temperature Germinability in Rice (Oryza sativa L.). Breed. Sci. 2001, 51, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Zhu, L.; Xu, Y.; Zeng, D.; Wu, P.; Qian, Q. QTL analysis of seed dormancy in rice (Oryza sativa L.). Euphytica 2004, 140, 155–162. [Google Scholar] [CrossRef]
- Wan, J.M.; Cao, Y.J.; Wang, C.M.; Ikehashi, H. Quantitative Trait Loci Associated with Seed Dormancy in Rice. Crop Sci. 2005, 45, 712–716. [Google Scholar] [CrossRef]
- Wan, J.M.; Jiang, L.; Tang, J.Y.; Wang, C.M.; Hou, M.Y.; Jing, W.; Zhang, L.X. Genetic dissection of the seed dormancy trait in cultivated rice (Oryza sativa L.). Plant. Sci. 2006, 170, 786–792. [Google Scholar] [CrossRef]
- Gao, F.Y.; Ren, G.J.; Lu, X.J.; Sun, S.X.; Li, H.J.; Gao, Y.M.; Luo, H.; Yan, W.G.; Zhang, Y.Z. QTL analysis for resistance to preharvest sprouting in rice (Oryza sativa). Plant. Breed. 2008, 127, 268–273. [Google Scholar] [CrossRef]
- Cai, H.-W.; Morishima, H. Genomic regions affecting seed shattering and seed dormancy in rice. Theor. Appl. Genet. 2000, 100, 840–846. [Google Scholar] [CrossRef]
- Chen, L.; Lou, Q.J.; Sun, Z.X.; Xing, Y.Z.; Yu, X.Q.; Luo, L.J. QTL mapping of low temperature on germination rate of rice. Rice Sci. 2006, 13, 93–98. [Google Scholar]
- Jiang, L.; Liu, S.; Hou, M.; Tang, J.; Chen, L.; Zhai, H.; Wan, J. Analysis of QTLs for seed low temperature germinability and anoxia germinability in rice (Oryza sativa L.). Field Crop. Res. 2006, 98, 68–75. [Google Scholar] [CrossRef]
- Ji, S.L.; Jiang, L.; Wang, Y.H.; Zhang, W.W.; Liu, X.; Liu, S.J.; Chen, L.M.; Zhai, H.Q.; Wan, J.M. Quantitative trait loci mapping and stability for low temperature germination ability of rice. Plant. Breed. 2009, 128, 387–392. [Google Scholar] [CrossRef]
- Xie, K.; Jiang, L.; Lu, B.; Yang, C.; Li, L.; Liu, X.; Zhang, L.; Zhao, Z.; Wan, J. Identification of QTLs for seed dormancy in rice (Oryza sativa) L. Plant. Breed. 2010, 130, 328–332. [Google Scholar] [CrossRef]
- Marzougui, S.; Sugimoto, K.; Yamanouchi, U.; Shimono, M.; Hoshino, T.; Hori, K.; Kobayashi, M.; Ishiyama, K.; Yano, M. Mapping and characterization of seed dormancy QTLs using chromosome segment substitution lines in rice. Theor. Appl. Genet. 2011, 124, 893–902. [Google Scholar] [CrossRef]
- Li, L.; Liu, X.; Xie, K.; Wang, Y.; Liu, F.; Lin, Q.; Wang, W.; Yang, C.; Lu, B.; Liu, S.; et al. qLTG-9, a stable quantitative trait locus for low-temperature germination in rice (Oryza sativa L.). Theor. Appl. Genet. 2013, 126, 2313–2322. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Kazama, Y.; Chae, Y.; Sato, T. Confirmation of Novel Quantitative Trait Loci for Seed Dormancy at Different Ripening Stages in Rice. Rice Sci. 2013, 20, 207–212. [Google Scholar] [CrossRef]
- Wang, L.; Cheng, J.; Lai, Y.; Du, W.; Huang, X.; Wang, Z.; Zhang, H. Identification of QTLs with additive, epistatic and QTL × development interaction effects for seed dormancy in rice. Planta 2013, 239, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kwon, S.W. Analysis of quantitative trait loci associated with seed germination and coleop-tile length under low temperature condition. J. Crop Sci. Biotechnol. 2015, 18, 273–278. [Google Scholar] [CrossRef]
- Li, W.; Xu, L.; Bai, X.; Xing, Y. Quantitative trait loci for seed dormancy in rice. Euphytica 2010, 178, 427–435. [Google Scholar] [CrossRef]
- Fujino, K.; Sekiguchi, H.; Sato, T.; Kiuchi, H.; Nonoue, Y.; Takeuchi, Y.; Ando, T.; Lin, S.Y.; Yano, M. Mapping of quantitative trait loci controlling low-temperature germinability in rice (Oryza sativa L.). Theor. Appl. Genet. 2003, 108, 794–799. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.-Y.; Kianian, S.F.; Foley, M.E. Multiple Loci and Epistases Control Genetic Variation for Seed Dormancy in Weedy Rice (Oryza sativa). Genetics 2004, 166, 1503–1516. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.-Y.; Kianian, S.F.; Hareland, G.A.; Hoffer, B.L.; Foley, M.E. Genetic analysis of adaptive syndromes interrelated with seed dormancy in weedy rice (Oryza sativa). Theor. Appl. Genet. 2005, 110, 1108–1118. [Google Scholar] [CrossRef]
- Ye, H.; Foley, M.E.; Gu, X. New seed dormancy loci detected from weedy rice-derived ad-vanced populations with major QTL alleles removed from the background. Plant. Sci. 2010, 179, 612–619. [Google Scholar] [CrossRef]
- Subudhi, P.K.; Parco, A.; Singh, P.K.; DeLeon, T.; Karan, R.; Biradar, H.; Cohn, M.A.; Brar, D.S.; Sasaki, T. Genetic Architecture of Seed Dormancy in U.S. Weedy Rice in Different Genetic Backgrounds. Crop Sci. 2012, 52, 2564–2575. [Google Scholar] [CrossRef]
- Ye, H.; Feng, J.; Zhang, L.; Zhang, J.; Mispan, M.S.; Cao, Z.; Beighley, D.H.; Yang, J.; Gu, X. Map-Based Cloning of Seed Dormancy1-2 Identified a Gibberellin Synthesis Gene Regulating the Development of Endosperm-Imposed Dormancy in Rice. Plant. Physiol. 2015, 169, 2152–2165. [Google Scholar]
- Fujino, K.; Sekiguchi, H.; Matsuda, Y.; Sugimoto, K.; Ono, K.; Yano, M. Molecular identification of a major quantitative trait locus, qLTG3-1, controlling low-temperature germinability in rice. Proc. Natl. Acad. Sci. USA 2008, 105, 12623–12628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lü, Q.; Niu, X.; Zhang, M.; Wang, C.; Xu, Q.; Feng, Y.; Yang, Y.; Wang, S.; Yuan, X.; Yu, H.; et al. Genome-Wide Association Study of Seed Dormancy and the Genomic Consequences of Improvement Footprints in Rice (Oryza sativa L.). Front. Plant. Sci. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magwa, R.A.; Zhao, H.; Xing, Y. Genome-wide association mapping revealed a diverse genetic basis of seed dormancy across subpopulations in rice (Oryza sativa L.). BMC Genet. 2016, 17, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Huh, S.M.; Hwang, Y.-S.; Shin, Y.S.; Nam, M.H.; Kim, O.Y.; Yoon, I.S. Comparative transcriptome profiling of developing caryopses from two rice cultivars with differential dormancy. J. Plant. Physiol. 2013, 170, 1090–1100. [Google Scholar] [CrossRef]
- Mangrauthia, S.K.; Agarwal, S.; Sailaja, B.; Sarla, N.; Voleti, S.R. Transcriptome Analysis of Oryza sativa (Rice) Seed Germination at High Temperature Shows Dynamics of Genome Expression Associated with Hormones Signalling and Abiotic Stress Pathways. Trop. Plant. Biol. 2016, 9, 215–228. [Google Scholar] [CrossRef]
- Xie, K.; Bai, J.; Yang, Y.; Duan, N.; Ma, Y.; Guo, T.; Yao, F.; Ding, H. The RNA-seq transcriptome analysis identified genes related to rice seed dormancy. Biol. Plant. 2019, 63, 308–313. [Google Scholar] [CrossRef]
- Park, M.; Choi, W.; Shin, S.-Y.; Moon, H.; Lee, D.; Gho, Y.-S.; Jung, K.-H.; Jeon, J.-S.; Shin, C. Identification of Genes and MicroRNAs Affecting Pre-harvest Sprouting in Rice (Oryza sativa L.) by Transcriptome and Small RNAome Analyses. Front. Plant. Sci. 2021, 12. [Google Scholar] [CrossRef]
- Reyes, J.L.; Chua, N.-H. ABA induction of miR159 controls transcript levels of two MYB factors during Arabidopsis seed germination. Plant. J. 2007, 49, 592–606. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.-G.; Lee, H.-S.; Hwang, W.-H.; Yang, S.-Y.; Lee, Y.-H.; Lee, C.; Jeong, J.-H. Analysis of differences in germination between rice varieties using RNA-Sequencing. J. Korean Crop Soc. 2020, 65, 274–283. [Google Scholar] [CrossRef]
- Qin, H.; Wu, F.; Xie, K.; Cheng, Z.; Guo, X.; Zhang, X.; Wang, J.; Lei, C.; Wang, J.; Mao, L.; et al. Transcriptomics Analysis Identified Candidate Genes Colocalized with Seed Dormancy QTLs in Rice (Oryza sativa L.). J. Plant. Biol. 2010, 53, 330–337. [Google Scholar] [CrossRef]
- Zhu, C.-C.; Wang, C.-X.; Lu, C.-Y.; Wang, J.-D.; Zhou, Y.; Xiong, M.; Zhang, C.-Q.; Liu, Q.-Q.; Li, Q.-F. Genome-Wide Identification and Expression Analysis of OsbZIP09 Target Genes in Rice Reveal Its Mechanism of Controlling Seed Germination. Int. J. Mol. Sci. 2021, 22, 1661. [Google Scholar] [CrossRef]
- Shi, J.; Shi, J.; Liang, W.; Zhang, D. Integrating GWAS and transcriptomics to identify genes involved in seed dormancy in rice. Theor. Appl. Genet. 2021, 134, 3553–3562. [Google Scholar] [CrossRef]
- Ogawa, M.; Hanada, A.; Yamauchi, Y.; Kuwahara, A.; Kamiya, Y.; Yamaguchi, S. Gibberellin biosynthesis and response during Arabidopsis seed germination. Plant Cell 2003, 15, 1591–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobo, T.; Kowyama, Y.; Hattori, T. A bZIP factor, TRAB1, interacts with VP1 and mediates abscisic acid-induced transcription. Proc. Natl. Acad. Sci. USA 1999, 96, 15348–15353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Xu, S.; Fei, Y.; Cao, Y.; Wu, H. The rice (Oryza sativa) Rc gene improves resistance to preharvest sprouting and retains seed and milled rice quality. Plant. Breed. 2020, 140, 53–64. [Google Scholar] [CrossRef]
- Yaish, M.W.; El-Kereamy, A.; Zhu, T.; Beatty, P.; Good, A.; Bi, Y.-M.; Rothstein, S.J. The APETALA-2-Like Transcription Factor OsAP2-39 Controls Key Interactions between Abscisic Acid and Gibberellin in Rice. PLoS Genet. 2010, 6, e1001098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Singh, S.; Randhawa, H.; Singh, J. Polymorphic Homoeolog of Key Gene of RdDM Pathway, ARGONAUTE4_9 class Is Associated with Pre-Harvest Sprouting in Wheat (Triticum aestivum L.). PLoS ONE 2013, 8, e77009. [Google Scholar] [CrossRef]
- Luján-Soto, E.; Dinkova, T. Time to Wake Up: Epigenetic and Small-RNA-Mediated Regulation during Seed Germination. Plants 2021, 10, 236. [Google Scholar] [CrossRef]
- Karlson, C.K.S.; Mohd-Noor, S.N.; Nolte, N.; Tan, B.C. CRISPR/dCas9-Based Systems: Mechanisms and Applications in Plant Sciences. Plants 2021, 10, 2055. [Google Scholar] [CrossRef]
- Rani, D.P.; Sherif, M.S. Application of Exogenous dsRNAs-induced RNAi in Agriculture: Challenges and Triumphs. Front. Plant. Sci. 2020, 11, 946. [Google Scholar] [CrossRef]
- Nonaka, S.; Someya, T.; Kadota, Y.; Nakamura, K.; Ezura, H. Super-Agrobacterium ver. 4: Im-proving the Transformation Frequencies and Genetic Engineering Possibilities for Crop Plants. Front. Plant. Sci. 2019, 10, 1204. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Specialization of Study | Traits | QTL/Genes | Growth Condition | Markers Type | Markers No. | Mapping Population | Parents | References |
|---|---|---|---|---|---|---|---|---|
| QTL | PHS | qPHS-1-1, qPHS-1-2, qPHS-4, qPHS-5, qPHS-7, qPHS-8 | NG, Yellow and White paper bag | RFLPs | 6 | 71 F6 RILs | IR64 (Indica) × Asominori (Japonica) | [83] |
| PHS | qPHS1-1FC, qPHS1-2FC, qPHS1-1GC, qPHS1-2GC | FC, GH | SNPs | 8 | 88 F2:3 RILs | Jinsang (Japonica) × Gopum (Japonica) | [84] | |
| SD | qSD1-1, qSD1-2 | |||||||
| LTG | qLTG1-1, qLTG1-2 | |||||||
| SD | qSDR9.1 and qSDR9.2 | FC | SNPs | 2 | 44 BC4F5 CSSLs | Owarihatamochi (Japonica) × Koshihikari (Japonica) | [85] | |
| PHS | qPHS-3FD, qPHS-11FD, qPHS-3GH, qPHS-4GH, qPHS-11GH | FC, GH | KASP, CAPS, SNPs | 5 | F9 160 RILs | Odae (Japonica) × Unbong40 (Japonica) | [86] | |
| PHS | RM4108-RM5849, RM3455-RM6905 | FC | SSRs, SNPs | 2 | 79 N-BILs, 48 BC3F1 N-CSSLs, 41 BC4F1 K-CSSLs | Nipponbare × Koshihikari (Japonica) | [87] | |
| SD | C1488, R830, R1440, R1245, C390 | FC | RFLPs | 5 | 98 BC1F5 | Nipponbare (Japonica) × Kasalath (indica) | [88] | |
| LTG | qLTG-2, qLTG-4-1, qLTG-4-2, qLTG-5, qLTG-11 | FC | RFLPs | 5 | 98 BILs | Nipponbare (Japonica) × Kasalath (indica) | [89] | |
| SD | qSD-3, qSD-5, qSD-6 qSD-11 | FC | RFLPs, SSRs | 4 | 127 Double haploid (DH) lines | ZYQ8 (indica) × JX17 (Japonica) | [90] | |
| SD | qSD-1, qSD-3, qSD-7 | FC | SSRs | 3 | 166 F1, 12 F2, | IR50 (indica) × Tatsumimochi (Japonica) × Miyukimochi (Japonica) 3-way Breeding | [91] | |
| SD | qSdn-1, qSdnj-3, qSdn-5, qSdn-7, qSdn-11 | FC | SSRs | 5 | 168 BC1, 82 BC1, 148 F2 individuals | Nanjing35 (Japonica) × N22 (indica) and USSR (Japonica) × N22 (Indica) | [92] | |
| PHS | qPSR-2, qPSR-5, qPSR-8 | FC | SSRs | 3 | 164 F2 | K81 × G46B | [93] | |
| SD | qDOR-2, qDOR-3-1, qDOR-3-2, qDOR-3-3, qDOR-5-1, qDOR-5-2, qDOR-6-1, qDOR-6-2, qDOR-8, qDOR-9-1, qDOR-9-2, qDOR-11-1, qDOR-11-2, qDOR-11-3, qDOR-11-4, qDOR-11-5, qDOR-11-6 | FC | RFLPs | 17 | 125 F7 RILs | Pei-kuh × O. rufipogon | [94] | |
| LTG | qLTG-3, qLTG-10 | GC | SSRs | 2 | 198 Double haploid F1 | Zhenshan 97B (indica) × AAV002863 (Japonica) | [95] | |
| LTG | qLTG-3, qLTG-4, qLTG-5-1, qLTG-5-2, qLTG-5-3, qLTG-5-4, qLTG-7, qLTG-9, qLTG-10, qLTG-11-1, qLTG-11-2 | GC | SSRs | 11 | 148 F2 | USSR5 (Japonica) × N22 (Indica) | [96] | |
| GR | qGR-2, qGR-3, qGR-11, qGR-12, qGR-2, qGR-10, qGR-11, qGR-7 | GC | RFLPs, SSRs | 13 | 71 F6 RILs | IR64 (Indica) × Asominori (Japonica) | [97] | |
| GI | qGI-2, qGI-7, qGI-10, qGI-11 | |||||||
| MGT | qMGT-2 | |||||||
| SD | qSd-1, qSd-2, qSd-1-1, qSd-1-2 | GC | SSR | 4 | 122 BILS, 536 BC6F2 | N22 (Indica) × Nanjing35 (Japonica) | [98] | |
| SD | Sdr6, Sdr9, Sdr10 | CC (Short Day) | SSRs | 3 | 44 CSSLs, 87 F2 RILs | Koshihikari × Nona Bokra, F2 population of SL506 × Koshihikari | [99] | |
| LTG | qLTG-7, qLTG-9, qLTG-12, qLTG-7, qLTG-9 (Os09g0395600,Os09g0396300,Os09g0396900, Os09g0395700, and Os09g0395800) | FC | SSRs | 5 | F7 RILs, 181 individuals | USSR5 (Japonica) × N22 (Indica) | [100] | |
| SD | qSD-3.1, qSD-6.1, qSD-7.1, qSD-10.1 | FC | RFLPs | 4 | 98 BILs, 4 CSSLs | Nipponbare × Kasalath | [101] | |
| SD | qSD1.1, qSD2.2, qSD4.1, qSD4.2, qSD5.1, qSD2.1, qSD3.1, qSD7.1 | FC | SSRs | 8 | 150 RILs (F2:9) | Jiucaiqing (Japonica) × IR26 (Indica) | [102] | |
| LTG | qLTG3, qLTG7-1, qLTG7-2, qLTG12, qLTG8 | FC | SSRs, STS | 5 | 160 RILs | Tong88-7 × Millyang23 | [103] | |
| SD | qDGE1, qDGE5a, qDGE5b, qDGE7 | FC | RFLPs, SSRs | 4 | 240 RILs | ZS97 × MH63 (Hybrid Shanyou 63) | [104] | |
| LTG | qLTG-3-1, qLTG-3-2, qLTG-4 | FC | SSRs | 3 | F1 BC1F1, 122 BILs BC1F5 | Hayamasari (Japonica) × Italica Livorno (Japonica) | [105] | |
| SD | qSDS-4, qSDS-6, qSDS-7, qSDS-8, qSDS-12 | GH | SSRs | 5 | BC1 | EM93-1 × EM93-1 (indica-type Breeding line) × SS18-2 (indica wild-type weedy rice) | [106] | |
| SD | qSD4, qSD7-1, qSD7-2, qSD8, qSD12 | GH | SSRs | 5 | F1, 156 BC1 | SS18-2 (weedy Rice) × EM93-1 (Cultivated rice) | [107] | |
| SD | qSD1-2, qSD3, qSD6, qSD7-2, qSD10 | GH | SSRs | 5 | BC1F1 | SS18-2 × EM93-1 | [108] | |
| SD | qSD1, qSD3, qSD4, qSD7-1, qSD7-2, qSD7-3, qSD10, qSD11, qSD12 | FC | SSRs | 9 | BR RIL 198 indivuduls, CR RIL 174 individuals F8:9 generation | s Bengal × PSRR-1; Cypresss × PSRR-1 | [109] | |
| SD, PHS (R) | qSDR9.1, qSDR9.2 | FC | SSRs | 2 | 44 BC4F5, CSSL | Owarihatamochi × Koshihikari | [85] | |
| SD | qDOR-2, qDOR-3-1, qDOR-3-2, qDOR-3-3, qDOR-5-1, qDOR-5-2, qDOR-6-1, qDOR-6-2, qDOR-8, qDOR-9-1, qDOR-9-2, qDOR-11-1, qDOR-11-2, qDOR-11-3, qDOR-11-4, qDOR-11-5, qDOR-11-6 | FC | RFLPs | 17 | 189 F2, 158 F2 progency | Pei-kuh × O. rufipogon | [94] | |
| SD | qSD1-2 (* Map-based cloning) | CC | SSRs | 1 | BC5F3 F2 RIL | EM93-1 (Indica) × SS18-2 (weedy rice) | [110] | |
| LTG | qLTG3–1 (* Map-based cloning) | CC | SSRs, SNP, Indels | 1 | BILs 116 | Hayamasari × Italica Livorno | [111] | |
| SD | Sdr4 (* Map-based cloning) | CC | FNPs, SNPs, Indels | 1 | 28 BC4F2 | Nipponbare (Japonica) × Kasalath (Indica) | [59] | |
| PHS | Sdr6, qSD-1, qSD1, qDEG1, qSdn-1, Sdr1, qDT-SGC3.1, qSD-3, qSdn-5, qMT-SGC5.1, Sdr9, qDOR6-2, qSD6, SDR4, qMT-SGC7.2, qSD-7-2, qPHS-7, qSD12, qLTG3-1 | FC | GBS | 6 | 21 | Japonica (14) Indica (7) | [14] | |
| GWAS | PHS | Os01g03740, Os01g03730, Os01g03820, Os01g03840, Os01g03890, Os01g03914, Os01g03950, Os04g08460, Os04g08470, Os04g08570 | FC | SNPs | 10 | Indica, Japonica | 277 accessions | [70] |
| SD | RM6902, RM525, RM231, RM5963, qSD7-1, FHS7.0, RM234, FH8.1, qSD-11 | FC | SNPs | Indica | 453 accessions | [112] | ||
| SD | GA2ox3, GH3–2, EUI1, Sdr4, GA2ox3, OsEF3, OsbohE, OsISA, OsHPL2, EXP4, OsMADS13, AP59, OsAsr1, OsABI5, OSH43, Pid3, OSH43, OsCLC-1, OsLHY, OsBOR1 | FC | SNPs | 20 | Indica, Japonica, Aus | 350 accessions | [113] |
| Category | Material | Plant Organ/Developmental Stage | Study Objective | Methodology | Reference |
|---|---|---|---|---|---|
| PHS | Gopum (PHS-susceptible) and Jowoon (PHS-resistant) | 4 embryo and endosperm | miRNA PHS | RNASeq and small RNASeq (Illumina HiSeq 2500) | [117] |
| SD | Nona Bokra | Seeds (dormant and dormant broken) | SD | RNASeq (Illumina Hiseq 2000) | [116] |
| PHS | Gopum and Samgwang | Caryopses | PHS, SD | Microarray (Agilent DNA Microarray Scanner) | [114] |
| SD | Cultivar N22 and Q4646 | Seeds | SD | GeneChip arrays (Affymetrix Fluidics Station 450 and GeneChip Scanner 3000) | [120] |
| Germination | Cultivar N22 | Seedlings | GHT | RNASeq (Ion Proton sequencer) | [115] |
| PHS | Joun and Jopyeong | Seeds | PHS | RNASeq | [119] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sohn, S.-I.; Pandian, S.; Kumar, T.S.; Zoclanclounon, Y.A.B.; Muthuramalingam, P.; Shilpha, J.; Satish, L.; Ramesh, M. Seed Dormancy and Pre-Harvest Sprouting in Rice—An Updated Overview. Int. J. Mol. Sci. 2021, 22, 11804. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111804
Sohn S-I, Pandian S, Kumar TS, Zoclanclounon YAB, Muthuramalingam P, Shilpha J, Satish L, Ramesh M. Seed Dormancy and Pre-Harvest Sprouting in Rice—An Updated Overview. International Journal of Molecular Sciences. 2021; 22(21):11804. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111804
Chicago/Turabian StyleSohn, Soo-In, Subramani Pandian, Thamilarasan Senthil Kumar, Yedomon Ange Bovys Zoclanclounon, Pandiyan Muthuramalingam, Jayabalan Shilpha, Lakkakula Satish, and Manikandan Ramesh. 2021. "Seed Dormancy and Pre-Harvest Sprouting in Rice—An Updated Overview" International Journal of Molecular Sciences 22, no. 21: 11804. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111804