
Probing the Structural Dynamics of the Activation Gate of KcsA Using Homo-FRET Measurements

 , , ,
, , , 
 and
and
Abstract
:1. Introduction
2. Results
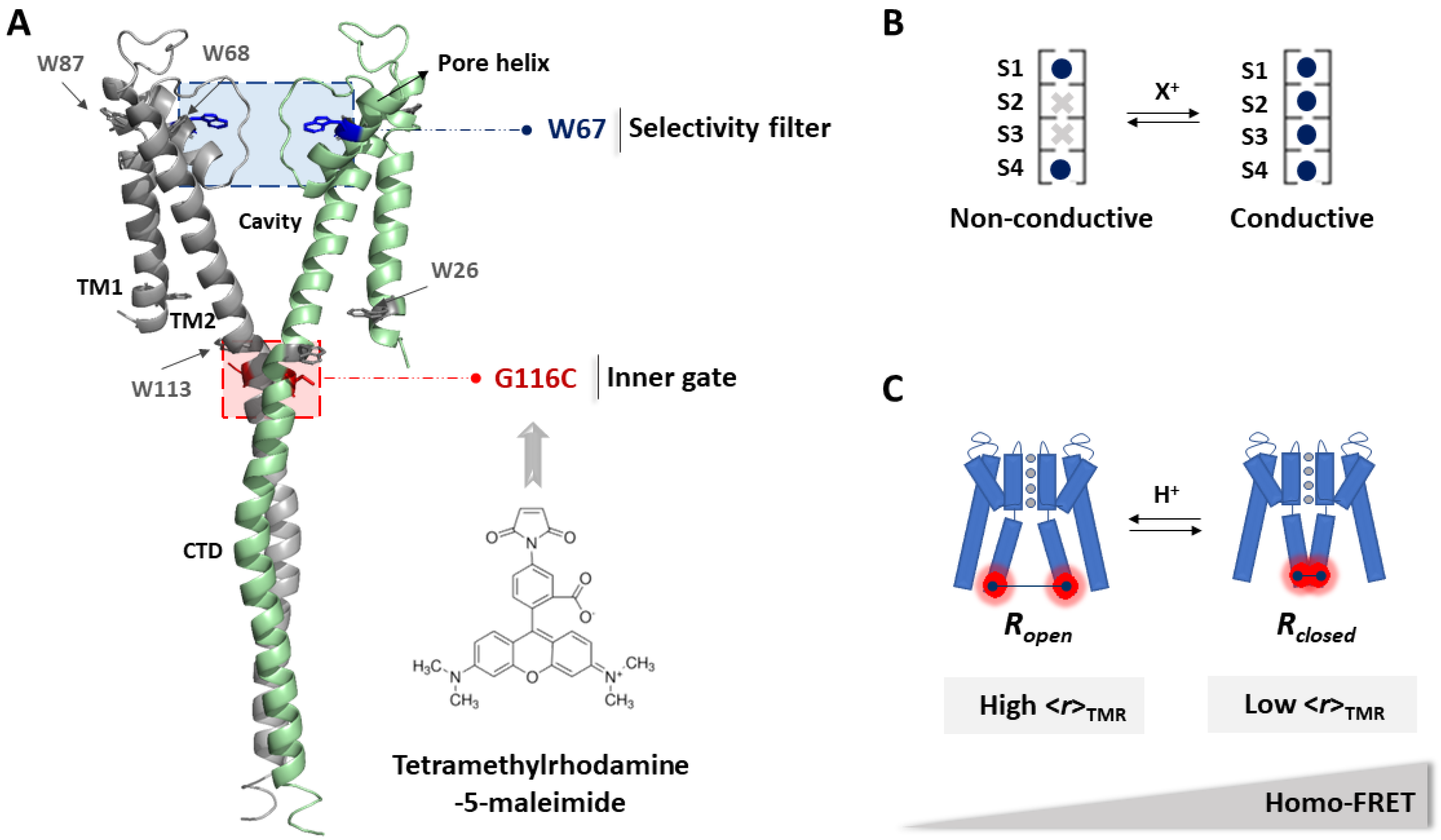
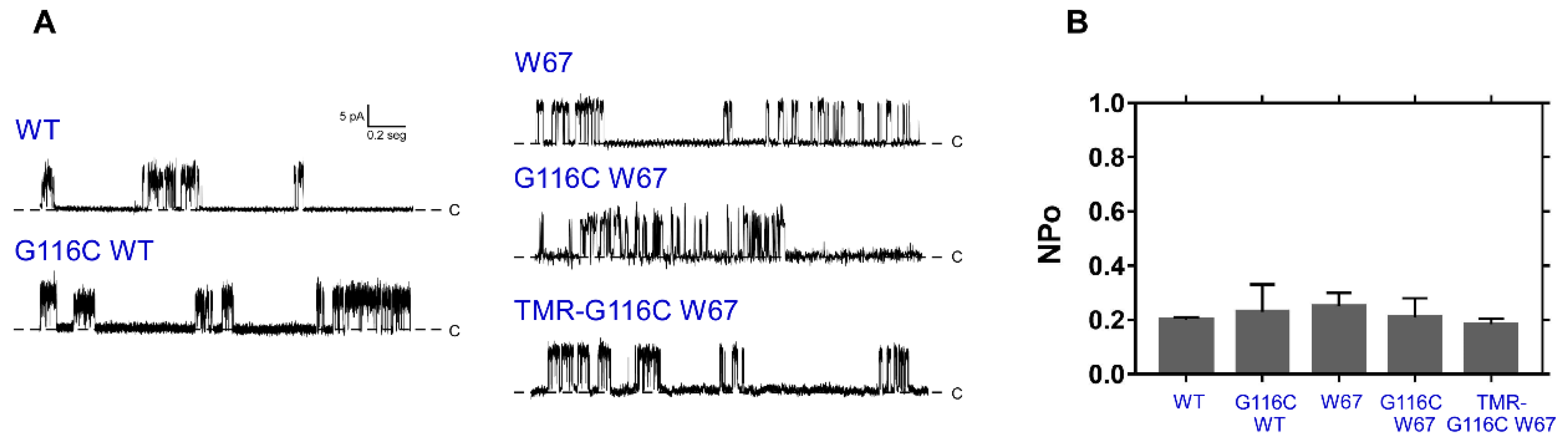
2.1. The G116C W67 KcsA Mutant Behaves as a Wild-Type Channel
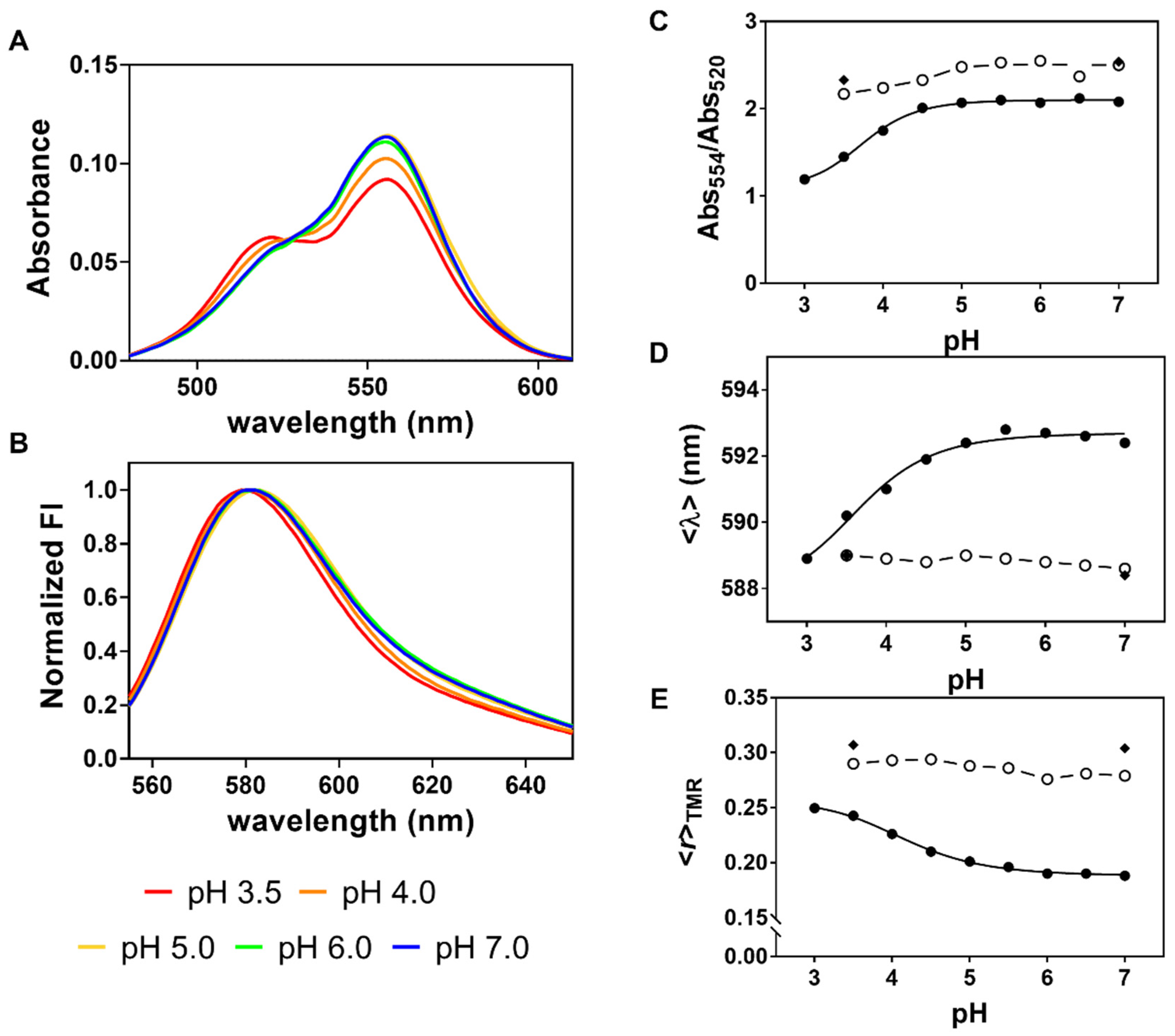
2.2. The Spectroscopic Properties of TMR Conjugated to G116C W67 KcsA Protein at a High Degree of Labeling Are Responsive to Channel Gating Induced by a pH Drop
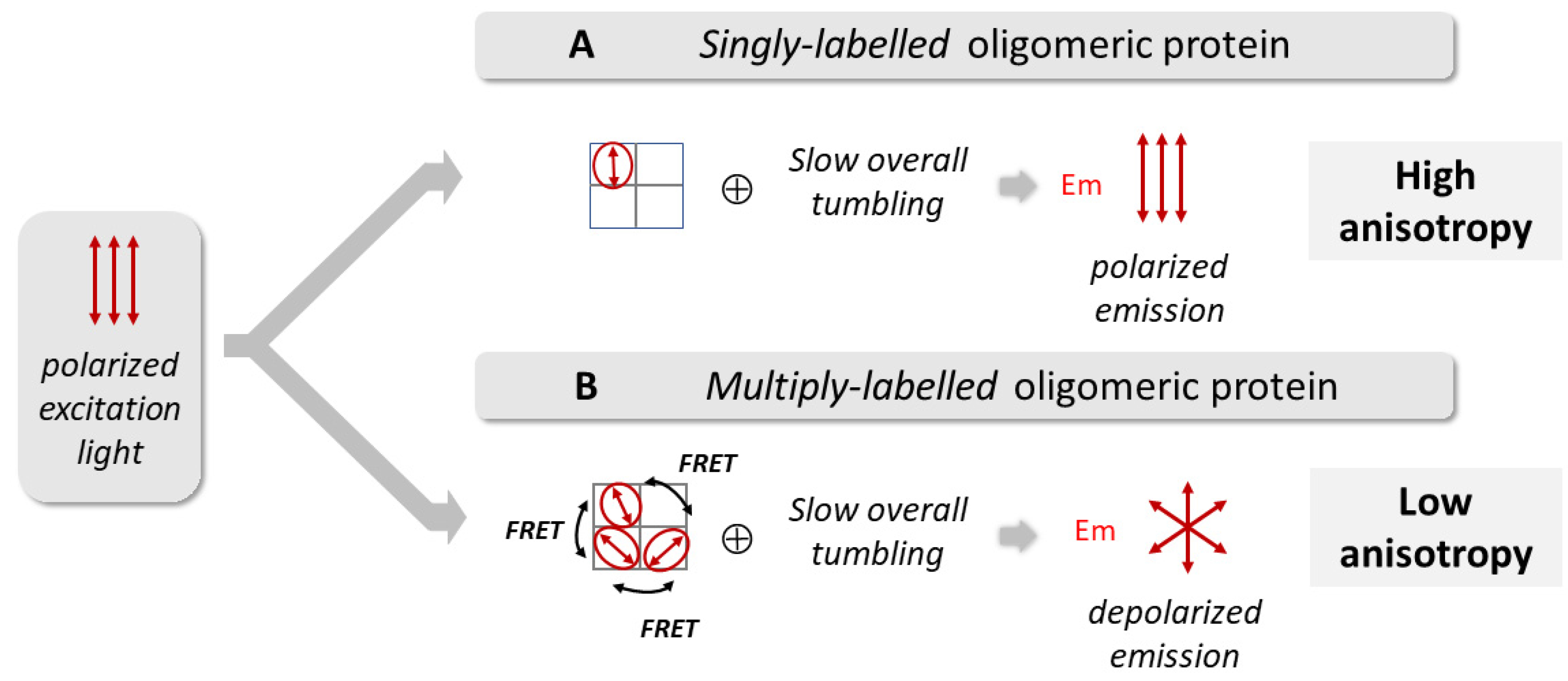
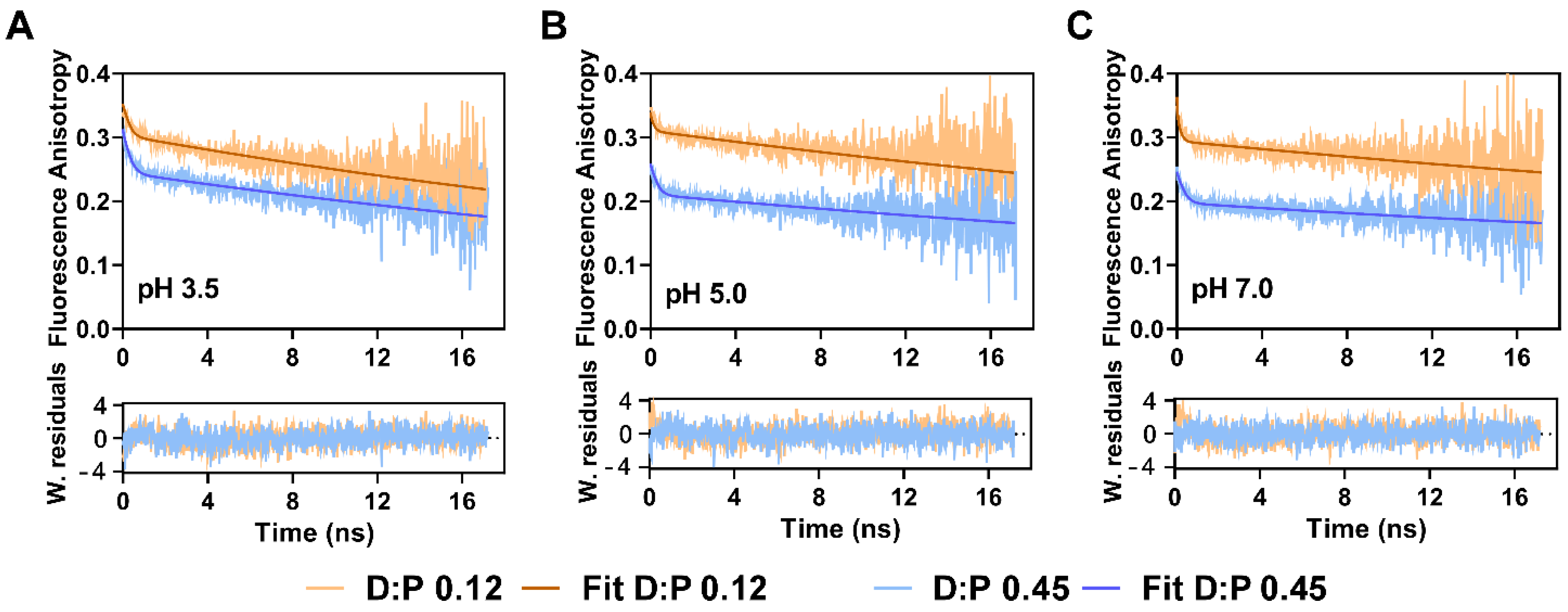
2.3. The Fluorescence Anisotropy of Multiply TMR-Labeled G116C W67 KcsA Is a Sensitive Reporter of pH Gating Due to an Efficient Intramolecular Homo-FRET Process
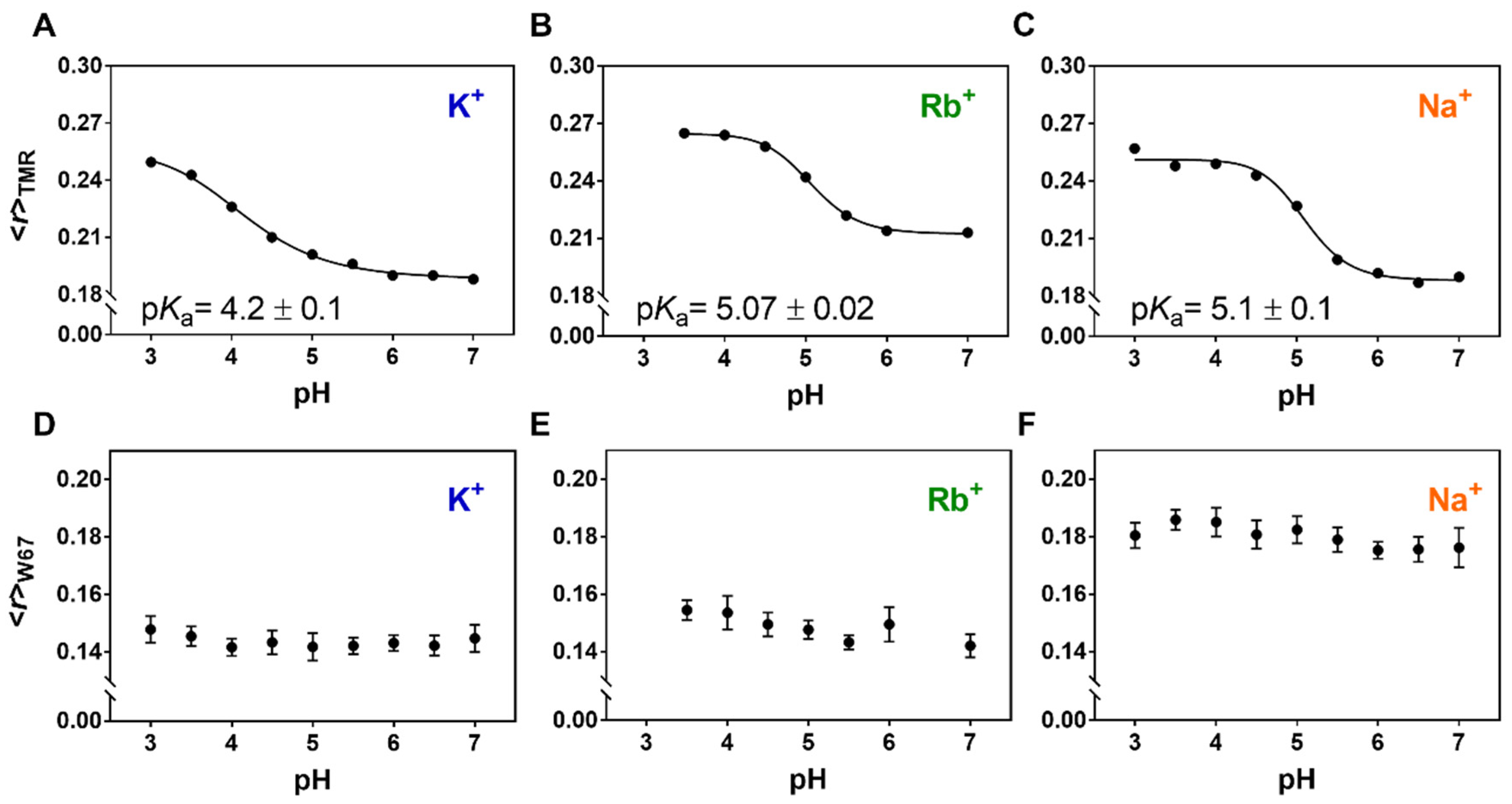
2.4. The Ion Occupancy of the SF Modulates the Allosteric Coupling of the Gates
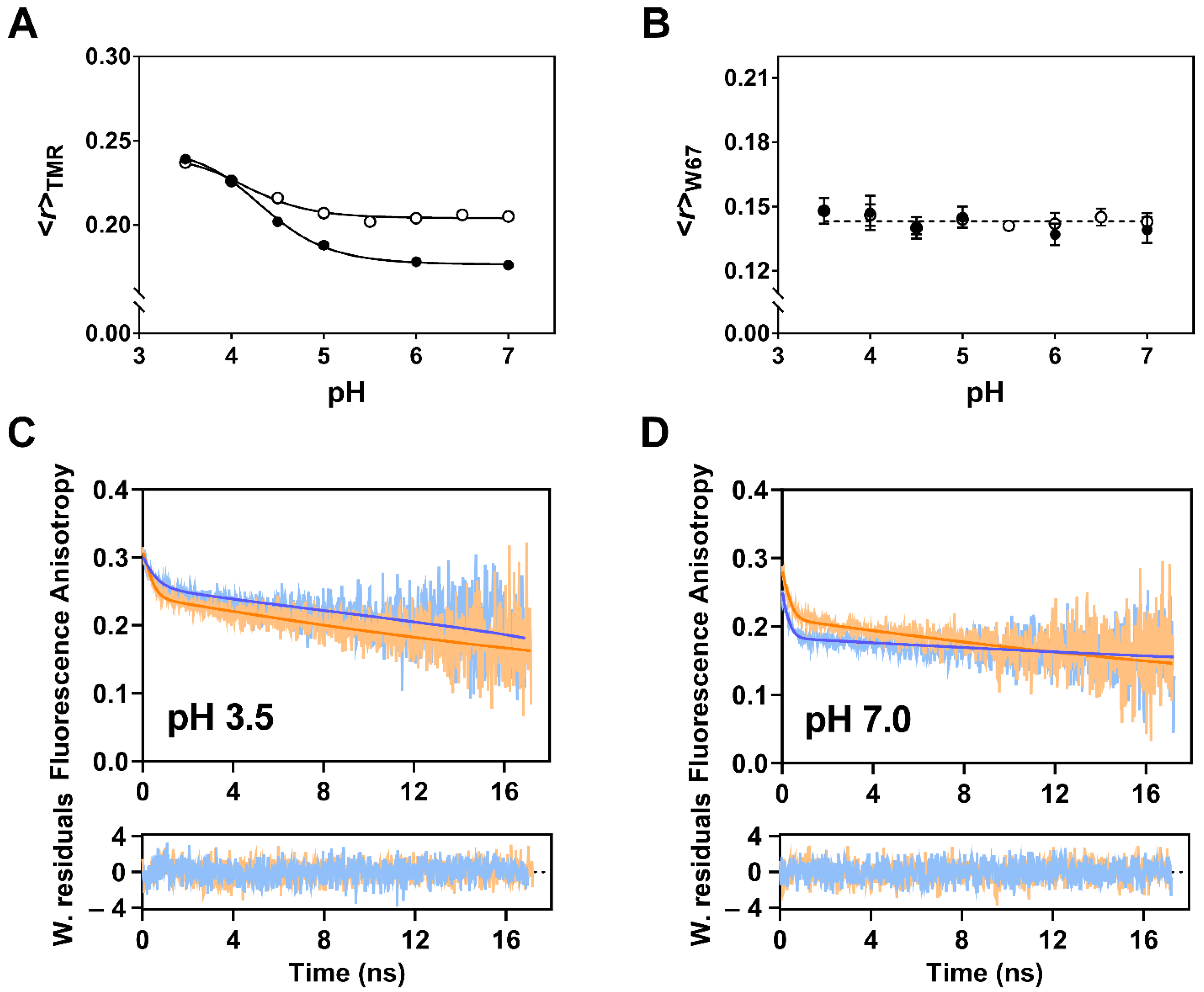
2.5. The Truncation of the C-Terminal Domain of KcsA Modestly Influences the Sensitivity of the Inner Gate to Protons
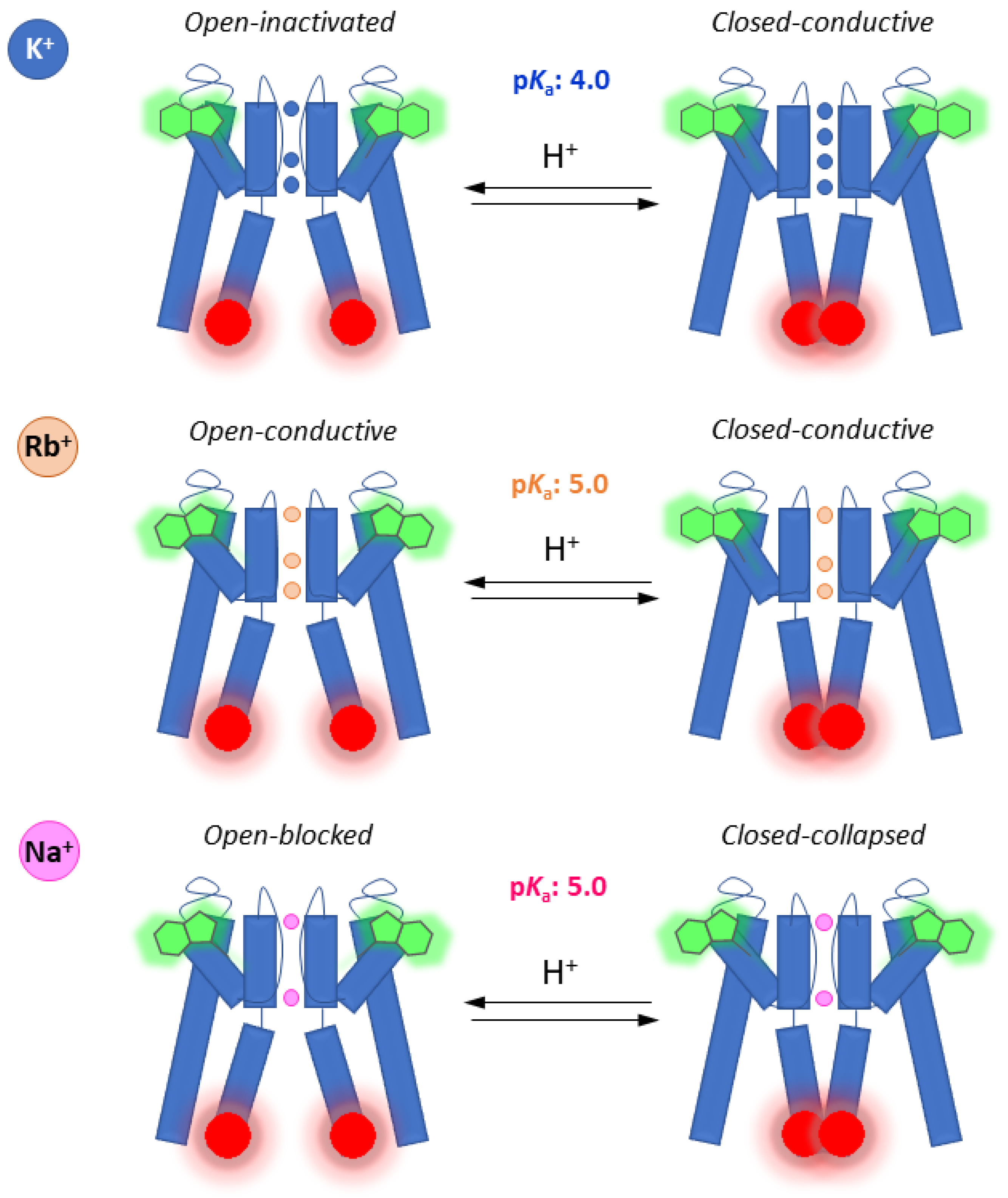
3. Discussion
4. Experimental Procedures
4.1. Heterologous Expression and Purification of KcsA
4.2. Functional and Structural Characterization of the Mutant Channels
4.3. TMR Labeling of G116C W67 KcsA Mutant Protein
4.4. Preparation of the Samples
4.5. Steady-State and Time-Resolved Spectroscopic Characterization
4.6. Quantum Yield and Förster Radius Determination of the TMR Labeled KcsA Channel
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hille, B. Ion Channels of Excitable Membranes, 3rd ed.; Mass Sinauer Assoc. Inc.: Sunderland, UK, 2001; ISBN 0-87893-321-2. [Google Scholar]
- Mironenko, A.; Zachariae, U.; de Groot, B.L.; Kopec, W. The Persistent Question of Potassium Channel Permeation Mechanisms. J. Mol. Biol. 2021, 433, 167002. [Google Scholar] [CrossRef] [PubMed]
- Kiss, L.; LoTurco, J.; Korn, S.J. Contribution of the selectivity filter to inactivation in potassium channels. Biophys. J. 1999, 76, 253–263. [Google Scholar] [CrossRef] [Green Version]
- González, C.; Baez-Nieto, D.; Valencia, I.; Oyarzún, I.; Rojas, P.; Naranjo, D.; Latorre, R. K+ Channels: Function-Structural Overview. Compr. Physiol. 2012, 2, 2087–2149. [Google Scholar] [PubMed]
- Xu, Y.; McDermott, A.E. Inactivation in the potassium channel KcsA. J. Struct. Biol. X 2019, 3, 100009. [Google Scholar] [CrossRef] [PubMed]
- Renart, M.L.; Giudici, A.M.; Díaz-García, C.; Molina, M.L.; Morales, A.; González-Ros, J.M.; Poveda, J.A. Modulation of Function, Structure and Clustering of K+ Channels by Lipids: Lessons Learnt from KcsA. Int. J. Mol. Sci. 2020, 21, 2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, D.A.; Morais, C.J.; Pfuetzner, R.A.; Kuo, A.; Gulbis, J.M.; Cohen, S.L.; Chait, B.T.; MacKinnon, R. The structure of the potassium channel: Molecular basis of K+ conduction and selectivity. Science 1998, 280, 69–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeMasurier, M.; Heginbotham, L.; Miller, C. KcsA: It’s a potassium channel. J. Gen. Physiol 2001, 118, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Uysal, S.; Vasquez, V.; Tereshko, V.; Esaki, K.; Fellouse, F.A.; Sidhu, S.S.; Koide, S.; Perozo, E.; Kossiakoff, A. Crystal structure of full-length KcsA in its closed conformation. Proc. Natl. Acad. Sci. USA 2009, 106, 16644–16649. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Morais-Cabral, J.H.; Kaufman, A.; Mackinnon, R. Chemistry of ion coordination and hydration revealed by a K+ channel-Fab complex at 2.0 Å resolution. Nature 2001, 414, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Morais-Cabral, J.H.; Zhou, Y.; MacKinnon, R. Energetic optimization of ion conduction rate by the K+ selectivity filter. Nature 2001, 414, 37–40. [Google Scholar] [CrossRef]
- Renart, M.L.; Giudici, A.M.; Poveda, J.A.; Fedorov, A.; Berberan-Santos, M.N.; Prieto, M.; Díaz-García, C.; González-Ros, J.M.; Coutinho, A. Conformational plasticity in the KcsA potassium channel pore helix revealed by homo-FRET studies. Sci. Rep. 2019, 9, 6215–6228. [Google Scholar] [CrossRef] [PubMed]
- Cordero-Morales, J.F.; Cuello, L.G.; Zhao, Y.; Jogini, V.; Cortes, D.M.; Roux, B.; Perozo, E. Molecular determinants of gating at the potassium-channel selectivity filter. Nat. Struct. Mol. Biol. 2006, 13, 311–318. [Google Scholar] [CrossRef]
- Poveda, J.A.; Marcela Giudici, A.; Lourdes Renart, M.; Morales, A.; González-Ros, J.M. Towards understanding the molecular basis of ion channel modulation by lipids: Mechanistic models and current paradigms. Biochim. Biophys. Acta-Biomembr. 2017, 1859, 1507–1516. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Mi, X.; Paajanen, V.; Wang, K.; Fan, Z. Activation-coupled inactivation in the bacterial potassium channel KcsA. Proc. Natl. Acad. Sci. USA 2005, 102, 17630–17635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuello, L.G.; Jogini, V.; Cortes, D.M.; Pan, A.C.; Gagnon, D.G.; Dalmas, O.; Cordero-Morales, J.F.; Chakrapani, S.; Roux, B.; Perozo, E. Structural basis for the coupling between activation and inactivation gates in K+channels. Nature 2010, 466, 272–275. [Google Scholar] [CrossRef]
- Ader, C.; Schneider, R.; Hornig, S.; Velisetty, P.; Vardanyan, V.; Giller, K.; Ohmert, I.; Becker, S.; Pongs, O.; Baldus, M. Coupling of activation and inactivation gate in a K-channel: Potassium and ligand sensitivity. EMBO J. 2009, 28, 2825–2834. [Google Scholar] [CrossRef] [Green Version]
- Wylie, B.J.; Bhate, M.P.; McDermott, A.E. Transmembrane allosteric coupling of the gates in a potassium channel. Proc. Natl. Acad. Sci. USA 2014, 111, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Bhate, M.P.; McDermott, A.E. Transmembrane allosteric energetics characterization for strong coupling between proton and potassium ion binding in the KcsA channel. Proc. Natl. Acad. Sci. USA 2017, 114, 8788–8793. [Google Scholar] [CrossRef] [Green Version]
- Labro, A.J.; Cortes, D.M.; Tilegenova, C.; Cuello, L.G. Inverted allosteric coupling between activation and inactivation gates in K+ channels. Proc. Natl. Acad. Sci. USA 2018, 115, 5426–5431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giudici, A.; Díaz-García, C.; Renart, M.; Coutinho, A.; Prieto, M.; González-Ros, J.; Poveda, J. Tetraoctylammonium, a Long Chain Quaternary Ammonium Blocker, Promotes a Noncollapsed, Resting-Like Inactivated State in KcsA. Int. J. Mol. Sci. 2021, 22, 490. [Google Scholar] [CrossRef] [PubMed]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy Principles of Fluorescence Spectroscopy; Springer: Boston, MA, USA, 2006; ISBN1 978-0-387-31278-1. ISBN2 978-0-387-46312-4. [Google Scholar]
- Stryer, L.; Haugland, R.P. Energy transfer: A spectroscopic ruler. Proc. Natl. Acad. Sci. USA 1967, 58, 719–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valeur, B.; Berberan-Santos, M.N. Molecular Fluorescence: Principles and Applications, 2nd ed.; Wiley-Vch: Weinheim, Germany, 2012. [Google Scholar]
- Bergström, F.; Hägglöf, P.; Karolin, J.; Ny, T.; Johansson, L. The use of site-directed fluorophore labeling and donor-donor energy migration to investigate solution structure and dynamics in proteins. Proc. Natl. Acad. Sci. USA 1999, 96, 12477–12481. [Google Scholar] [CrossRef] [Green Version]
- Bader, A.N.; Hoetzl, S.; Hofman, E.G.; Voortman, J.; van Bergen en Henegouwen, P.M.; van Meer, G.; Gerritsen, H.C. Homo-FRET imaging as a tool to quantify protein and lipid clustering. ChemPhysChem 2011, 12, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Tramier, M.; Coppey-Moisan, M. Fluorescence anisotropy imaging microscopy for homo-FRET in living cells. Methods Cell Biol. 2008, 85, 395–414. [Google Scholar]
- Ghosh, S.; Saha, S.; Goswami, D.; Bilgrami, S.; Mayor, S. Dynamic Imaging of Homo-FRET in Live Cells by Fluorescence Anisotropy Microscopy. Methods Enzymol. 2012, 505, 291–327. [Google Scholar]
- Thaler, C.; Koushik, S.V.; Puhl, H.L.; Blank, P.S.; Vogel, S.S. Structural rearrangement of CaMKIIα catalytic domains encodes activation. Proc. Natl. Acad. Sci. USA 2009, 106, 6369–6374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Runnels, L.; Scarlata, S. Theory and application of fluorescence homotransfer to melittin oligomerization. Biophys. J. 1995, 69, 1569–1583. [Google Scholar] [CrossRef] [Green Version]
- Warren, S.C.; Margineanu, A.; Katan, M.; Dunsby, C.; French, P.M.W. Homo-FRET Based Biosensors and Their Application to Multiplexed Imaging of Signalling Events in Live Cells. Int. J. Mol. Sci. 2015, 16, 14695–14716. [Google Scholar] [CrossRef] [Green Version]
- Cameron, W.D.; Bui, C.V.; Hutchinson, A.; Loppnau, P.; Gräslund, S.; Rocheleau, J. V Apollo-NADP+: A spectrally tunable family of genetically encoded sensors for NADP+. Nat. Methods 2016, 13, 352–358. [Google Scholar] [CrossRef]
- Ross, B.L.; Tenner, B.; Markwardt, M.L.; Zviman, A.; Shi, G.; Kerr, J.P.; Snell, N.E.; McFarland, J.J.; Mauban, J.R.; Ward, C.W.; et al. Single-color, ratiometric biosensors for detecting signaling activities in live cells. Elife 2018, 7, e35458. [Google Scholar] [CrossRef] [PubMed]
- Chan, F.; Kaminski, C.; Schierle, G.K. Homo-FRET fluorescence anisotropy imaging as a tool to study molecular self-assembly in live cells. ChemPhysChem 2011, 12, 500–509. [Google Scholar] [CrossRef]
- Perozo, E.; Cortes, D.M.; Cuello, L.G. Three-dimensional architecture and gating mechanism of a K+ channel studied by EPR spectroscopy. Nat. Struct. Biol. 1998, 5, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Perozo, E.; Cortes, D.; Cuello, L. Structural rearrangements underlying K+-channel activation gating. Science 1999, 285, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Tilegenova, C.; Cortes, D.M.; Cuello, L.G. Hysteresis of KcsA potassium channel’s activation– deactivation gating is caused by structural changes at the channel’s selectivity filter. Proc. Natl. Acad. Sci. USA 2017, 114, 3234–3239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, I.S.M.; Spence, M. The Molecular Probes Handbook. A Guide to Fluorescent Probes and Labelling Technologies, 11st ed.; Biochemistry: Moscow, Russia, 2011. [Google Scholar]
- Blunck, R.; Cordero-Morales, J.F.; Cuello, L.G.; Perozo, E.; Bezanilla, F. Detection of the opening of the bundle crossing in KcsA with fluorescence lifetime spectroscopy reveals the existence of two gates for ion conduction. J. Gen. Physiol. 2006, 128, 569–581. [Google Scholar] [CrossRef] [Green Version]
- Vaid, M.; Claydon, T.W.; Rezazadeh, S.; Fedida, D. Voltage clamp fluorimetry reveals a novel outer pore instability in a mammalian voltage-gated potassium channel. J. Gen. Physiol. 2008, 132, 209–222. [Google Scholar] [CrossRef] [Green Version]
- Cowgill, J.; Chanda, B. The contribution of voltage clamp fluorometry to the understanding of channel and transporter mechanisms. J. Gen. Physiol. 2019, 151, 1163–1172. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Lee, S.-J.; Heyman, S.; Enkvetchakul, D.; Nichols, C.G. Structural rearrangements underlying ligand-gating in Kir channels. Nat. Commun. 2012, 3, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Iwamoto, M.; Shimizu, H.; Inoue, F.; Konno, T.; Sasaki, Y.C.; Oiki, S. Surface Structure and Its Dynamic Rearrangements of the KcsA Potassium Channel upon Gating and Tetrabutylammonium Blocking. J. Biol. Chem. 2006, 281, 28379–28386. [Google Scholar] [CrossRef] [Green Version]
- Chakrapani, S.; Cordero-Morales, J.F.; Perozo, E. A quantitative description of KcsA gating II: Single-channel currents. J. Gen. Physiol. 2007, 130, 479–496. [Google Scholar] [CrossRef] [Green Version]
- Cuello, L.G.; Jogini, V.; Cortes, D.M.; Perozo, E. Structural mechanism of C-type inactivation in K+ channels. Nature 2010, 466, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Sadler, E.; Kapanidis, A.; Tucker, S. Solution-Based Single-Molecule FRET Studies of K+ Channel Gating in a Lipid Bilayer. Biophys. J. 2016, 110, 2663–2670. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, M.; Kosaka, N.; Choyke, P.L.; Kobayashi, H. H-Type Dimer Formation of Fluorophores: A Mechanism for Activatable, in Vivo Optical Molecular Imaging. ACS Chem. Biol. 2009, 4, 535–546. [Google Scholar] [CrossRef] [Green Version]
- Donaphon, B.; Bloom, L.; Levitus, M. Photophysical characterization of interchromophoric interactions between rhodamine dyes conjugated to proteins. Methods Appl. Fluoresc. 2018, 6, 045004. [Google Scholar] [CrossRef]
- Rotem, D.; Mason, A.; Bayley, H. Inactivation of the KcsA potassium channel explored with heterotetramers. J. Gen. Physiol. 2010, 135, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Setiawan, D.; Kazaryan, A.; Martoprawiro, M.; Filatov, M. A first principles study of fluorescence quenching in rhodamine B dimers: How can quenching occur in dimeric species? Phys. Chem. Chem. Phys. 2010, 12, 11238–11244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, T.; Nishikiori, H.; Tamura, T. Absorption spectra of rhodamine B dimers in dip-coated thin films prepared by the sol-gel method. Chem. Phys. Lett. 1995, 233, 424–429. [Google Scholar] [CrossRef]
- Del Monte, F.; Levy, D. Formation of fluorescent rhodamine B J-dimers in sol-gel glasses induced by the adsorption geometry on the silica surface. J. Phys. Chem. B 1998, 102, 8036–8041. [Google Scholar] [CrossRef]
- Terdale, S.; Tantray, A. Spectroscopic study of the dimerization of rhodamine 6G in water and different organic solvents. J. Mol. Liq. 2017, 225, 662–671. [Google Scholar] [CrossRef]
- Bergström, F.; Mikhalyov, I.; Hägglöf, P.; Wortmann, R.; Tor Ny, A.; Johansson, L.B.-Å. Dimers of Dipyrrometheneboron Difluoride (BODIPY) with Light Spectroscopic Applications in Chemistry and Biology. J. Am. Chem. Soc. 2001, 124, 196–204. [Google Scholar] [CrossRef]
- Mikhalyov, I.; Gretskaya, N.; Bergström, F.; Johansson, L.B.-Å. Electronic ground and excited state properties of dipyrrometheneboron difluoride (BODIPY): Dimers with application to biosciences. Phys. Chem. Chem. Phys. 2002, 4, 5663–5670. [Google Scholar] [CrossRef]
- Zimmer, J.; Doyle, D.A.; Grossmann, J.G. Structural Characterization and pH-Induced Conformational Transition of Full-Length KcsA. Biophys. J. 2006, 90, 1752–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qasim, A.; Sher, I.; Hirschhorn, O.; Shaked, H.; Qasem, Z.; Ruthstein, S.; Chill, J.H. Investigation of a KcsA Cytoplasmic pH Gate in Lipoprotein Nanodiscs. ChemBioChem 2019, 20, 813–821. [Google Scholar] [CrossRef]
- Thompson, A.N.; Posson, D.J.; Parsa, P.V.; Nimigean, C.M. Molecular mechanism of pH sensing in KcsA potassium channels. Proc. Natl. Acad. Sci. USA 2008, 105, 6900–6905. [Google Scholar] [CrossRef] [Green Version]
- Uysal, S.; Cuello, L.G.; Cortes, D.M.; Koide, S.; Kossiakoff, A.A.; Perozo, E. Mechanism of activation gating in the full-length KcsA K+ channel. Proc. Natl. Acad. Sci. USA 2011, 108, 11896–11899. [Google Scholar] [CrossRef] [Green Version]
- Pantazis, A.; Westerberg, K.; Althoff, T.; Abramson, J.; Olcese, R. Harnessing photoinduced electron transfer to optically determine protein sub-nanoscale atomic distances. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahl, P. Analysis of fluorescence anisotropy decays by a least square method. Biophys. Chem. 1979, 10, 91–104. [Google Scholar] [CrossRef]
- Jameson, D.M.; Ross, J.A. Fluorescence Polarization/Anisotropy in Diagnostics and Imaging. Chem. Rev. 2010, 110, 2685–2708. [Google Scholar] [CrossRef] [Green Version]
- Melo, A.M.; Fedorov, A.; Prieto, M.; Coutinho, A. Exploring homo-FRET to quantify the oligomer stoichiometry of membrane-bound proteins involved in a cooperative partition equilibrium. Phys. Chem. Chem. Phys. 2014, 16, 18105–18117. [Google Scholar] [CrossRef]
- Zhou, Y.; MacKinnon, R. The occupancy of ions in the K+ selectivity filter: Charge balance and coupling of ion binding to a protein conformational change underlie high conduction rates. J. Mol. Biol. 2003, 333, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Medintz, I.; Hildebrandt, N. FRET-Förster Resonance Energy Transfer: From Theory to Applications; Wiley-Vch: Weinheim, Germany, 2013; pp. 1–791. ISBN 978-3-527-32816. [Google Scholar]
- Chakrapani, S.; Cordero-Morales, J.F.; Perozo, E. A quantitative description of KcsA gating I: Macroscopic currents. J. Gen. Physiol. 2007, 130, 465–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulse, R.; Sachleben, J.; Wen, P.; Moradi, M.; Tajkhorshid, E.; Perozo, E. Conformational dynamics at the inner gate of KcsA during activation. Biochemistry 2014, 53, 2557–2559. [Google Scholar] [CrossRef]
- Liu, S.; Focke, P.J.; Matulef, K.; Bian, X.; Moënne-Loccoz, P.; Valiyaveetil, F.I.; Lockless, S.W. Ion-binding properties of a K+ channel selectivity filter in different conformations. Proc. Natl. Acad. Sci. USA 2015, 112, 15096–15100. [Google Scholar] [CrossRef] [Green Version]
- Giudici, A.M.; Renart, M.L.; Díaz-García, C.; Morales, A.; Poveda, J.A.; González-Ros, J.M. Accessibility of Cations to the Selectivity Filter of KcsA in the Inactivated State: An Equilibrium Binding Study. Int. J. Mol. Sci. 2019, 20, 689. [Google Scholar] [CrossRef] [Green Version]
- López Arbeloa, F.; Martínez Martínez, V.; Arbeloa, T.; López Arbeloa, I. Photoresponse and anisotropy of rhodamine dye intercalated in ordered clay layered films. J. Photochem. Photobiol. C Photochem. Rev. 2007, 8, 85–108. [Google Scholar] [CrossRef]
- Hirano, M.; Takeuchi, Y.; Aoki, T.; Yanagida, T.; Ide, T. Rearrangements in the KcsA cytoplasmic domain underlie its gating. J. Biol. Chem. 2010, 285, 3777–3783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwamoto, M.; Oiki, S. Amphipathic antenna of an inward rectifier K+ channel responds to changes in the inner membrane leaflet. Proc. Natl. Acad. Sci. USA 2013, 110, 749–754. [Google Scholar] [CrossRef] [Green Version]
- Barrera, F.N.; Renart, M.L.; Molina, M.L.; Poveda, J.A.; Encinar, J.A.; Fernández, A.M.; Neira, J.L.; González-Ros, J.M. Unfolding and refolding in vitro of a tetrameric, alpha-helical membrane protein: The prokaryotic potassium channel KcsA. Biochemistry 2005, 44, 14344–14352. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Humana Press Inc.: Totowa, NJ, USA, 2005. [Google Scholar]
- Riquelme, G.; Lopez, E.; Garcia-Segura, L.M.; Ferragut, J.A.; Gonzalez-Ros, J.M. Giant liposomes: A model system in which to obtain patch-clamp recordings of ionic channels. Biochemistry 1990, 29, 11215–11222. [Google Scholar] [CrossRef]
- Molina, M.L.; Encinar, J.A.; Barrera, F.N.; Fernandez-Ballester, G.; Riquelme, G.; Gonzalez-Ros, J.M. Influence of C-terminal protein domains and protein-lipid interactions on tetramerization and stability of the potassium channel KcsA. Biochemistry 2004, 43, 14924–14931. [Google Scholar] [CrossRef] [Green Version]
- Renart, M.L.; Triano, I.; Poveda, J.A.; Encinar, J.A.; Fernández, A.M.; Ferrer-Montiel, A.V.; Gómez, J.; González Ros, J.M. Ion binding to KcsA: Implications in ion selectivity and channel gating. Biochemistry 2010, 49, 9480–9487. [Google Scholar] [CrossRef] [PubMed]
- Renart, M.L.; Barrera, F.N.; Molina, M.L.; Encinar, J.A.; Poveda, J.A.; Fernández, A.M.; Gómez, J.; González-Ros, J.M. Effects of conducting and blocking ions on the structure and stability of the potassium channel KcsA. J. Biol. Chem. 2006, 281, 29905–29915. [Google Scholar] [CrossRef] [Green Version]
- Poveda, J.A.; Prieto, M.; Encinar, J.A.; González-Ros, J.M.; Mateo, C.R. Intrinsic tyrosine fluorescence as a tool to study the interaction of the Shaker B “ball” peptide with anionic membranes. Biochemistry 2003, 42, 7124–7132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Würth, C.; Grabolle, M.; Pauli, J.; Spieles, M.; Resch-Genger, U. Relative and absolute determination of fluorescence quantum yields of transparent samples. Nat. Protoc. 2013, 8, 1535–1550. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | D:P | β1 a | φ1 b (ns) | β2 a | φ2 c (ns) | r(0) d | |
|---|---|---|---|---|---|---|---|
| 3.5 | 0.12 | 0.08 | 0.19 | 0.30 | 51 [49–55] | 0.38 | 1.191 |
| 0.45 | 0.09 | 0.23 | 0.25 | 0.34 | |||
| 5.0 | 0.12 | 0.09 | 0.10 | 0.31 | 76 [71–84] | 0.40 | 1.278 |
| 0.45 | 0.06 | 0.22 | 0.21 | 0.27 | |||
| 7.0 | 0.12 | 0.08 | 0.03 | 0.30 | 92 [85–103] | 0.38 | 1.289 |
| 0.45 | 0.06 | 0.23 | 0.20 | 0.26 |
| Protein | pH | β1 a | φ1 b (ns) | β2 a | φ2 c (ns) | r(0) | |
|---|---|---|---|---|---|---|---|
| Δ125 | 3.5 | 0.09 | 0.25 | 0.24 | 42 [40–44] | 0.33 | 1.134 |
| 4.0 | 0.08 | 0.30 | 0.23 | 43 [40–45] | 0.31 | 1.134 | |
| 4.5 | 0.10 | 0.20 | 0.23 | 39 [37–42] | 0.33 | 1.117 | |
| 5.0 | 0.09 | 0.23 | 0.21 | 41 [39–44] | 0.30 | 1.153 | |
| 5.5 | 0.10 | 0.18 | 0.21 | 40 [38–42] | 0.31 | 1.088 | |
| 6.0 | 0.09 | 0.23 | 0.22 | 39 [37–41] | 0.31 | 1.110 | |
| 6.5 | 0.09 | 0.21 | 0.21 | 44 [41–46] | 0.30 | 1.135 | |
| 7.0 | 0.09 | 0.20 | 0.21 | 45 [42–48] | 0.30 | 1.184 | |
| FL | 3.5 | 0.07 | 0.52 | 0.26 | 57 [53–61] | 0.32 | 1.210 |
| 7.0 | 0.07 | 0.16 | 0.18 | 99 [87–114] | 0.26 | 1.033 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-García, C.; Renart, M.L.; Poveda, J.A.; Giudici, A.M.; González-Ros, J.M.; Prieto, M.; Coutinho, A. Probing the Structural Dynamics of the Activation Gate of KcsA Using Homo-FRET Measurements. Int. J. Mol. Sci. 2021, 22, 11954. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111954
Díaz-García C, Renart ML, Poveda JA, Giudici AM, González-Ros JM, Prieto M, Coutinho A. Probing the Structural Dynamics of the Activation Gate of KcsA Using Homo-FRET Measurements. International Journal of Molecular Sciences. 2021; 22(21):11954. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111954
Chicago/Turabian StyleDíaz-García, Clara, Maria Lourdes Renart, José Antonio Poveda, Ana Marcela Giudici, José M. González-Ros, Manuel Prieto, and Ana Coutinho. 2021. "Probing the Structural Dynamics of the Activation Gate of KcsA Using Homo-FRET Measurements" International Journal of Molecular Sciences 22, no. 21: 11954. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111954