
Photoactivation of Cell-Free Expressed Archaerhodopsin-3 in a Model Cell Membrane


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
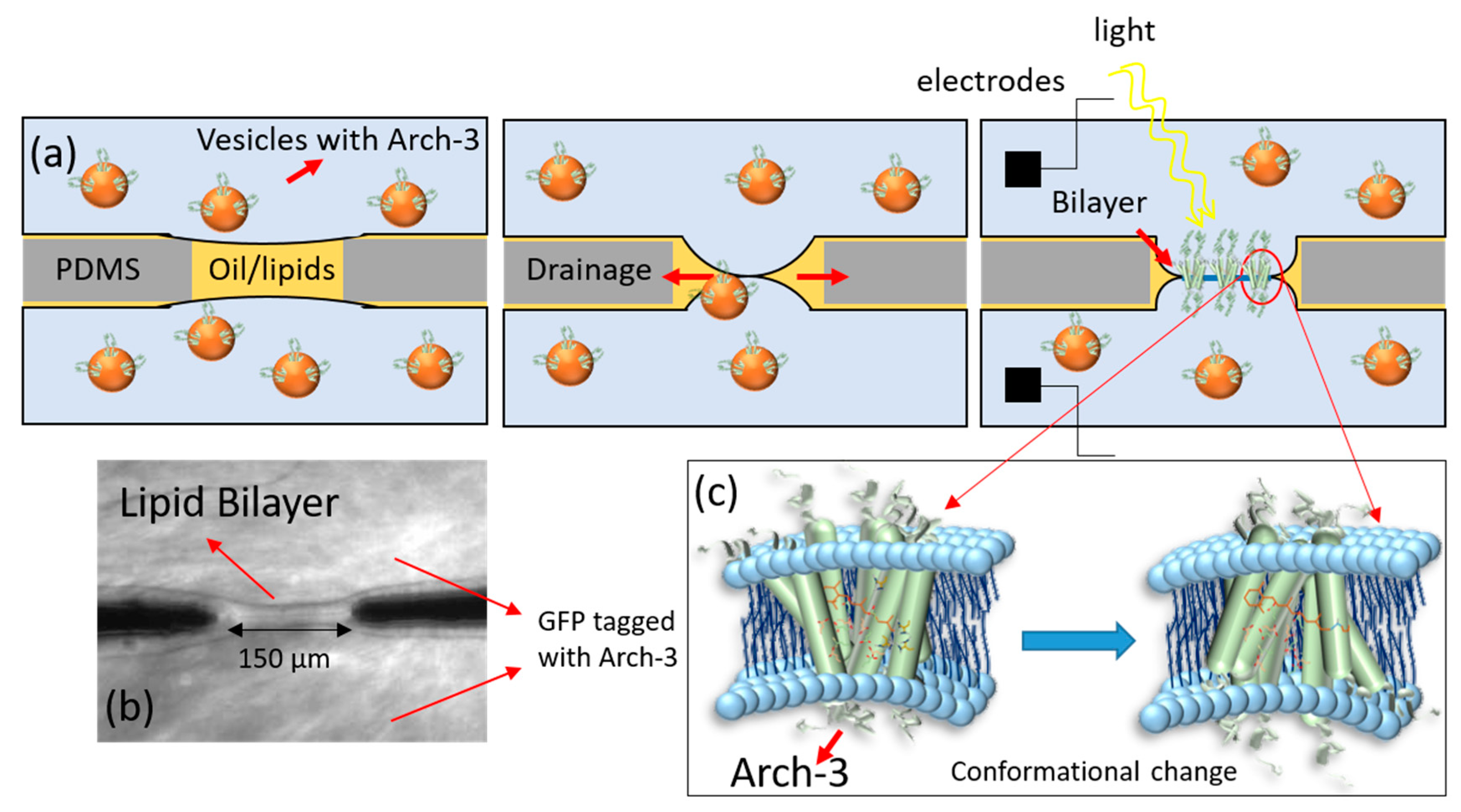
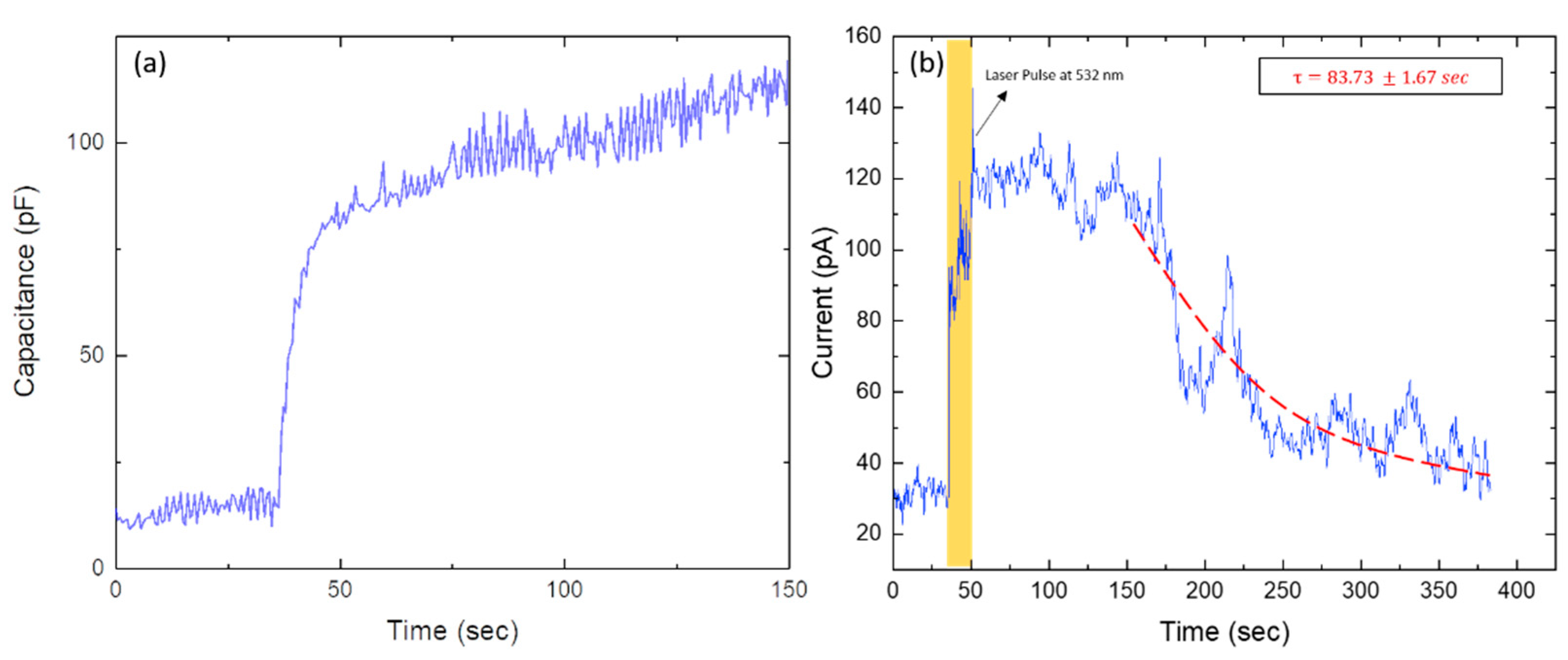
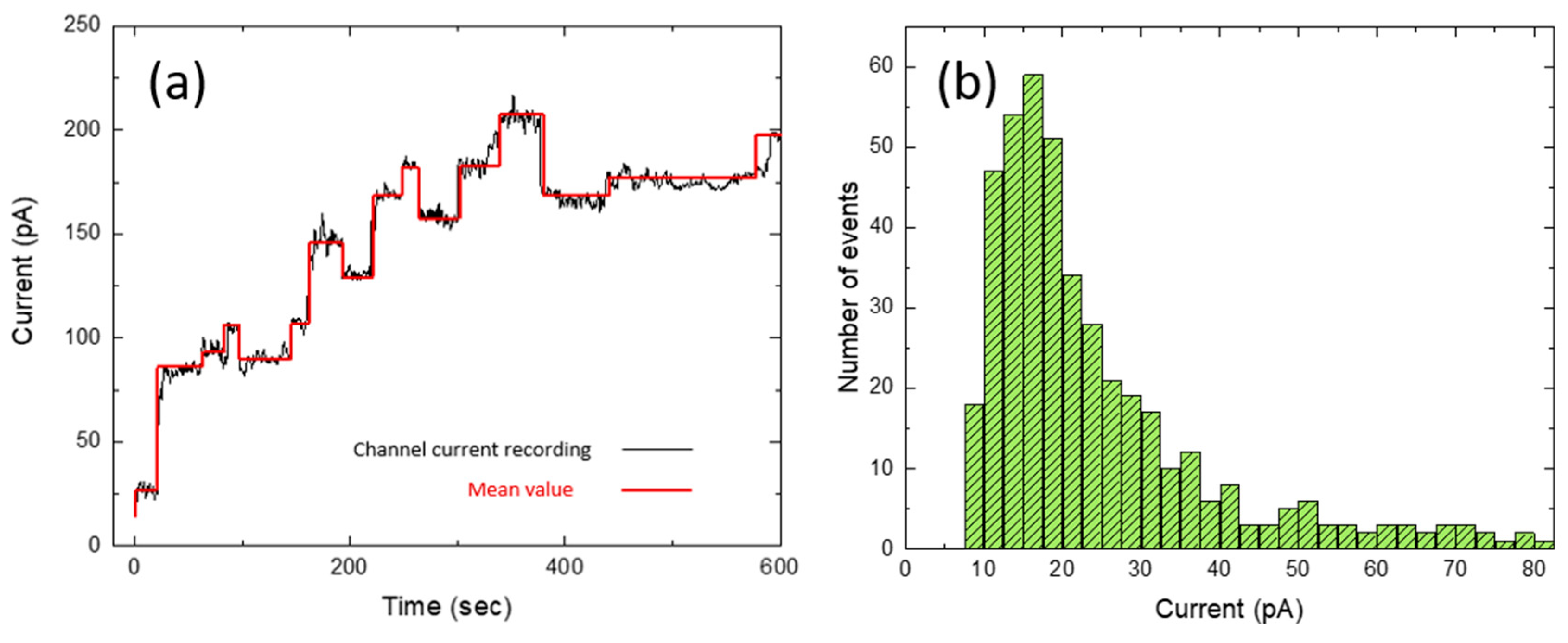
2. Results and Discussion
3. Conclusions
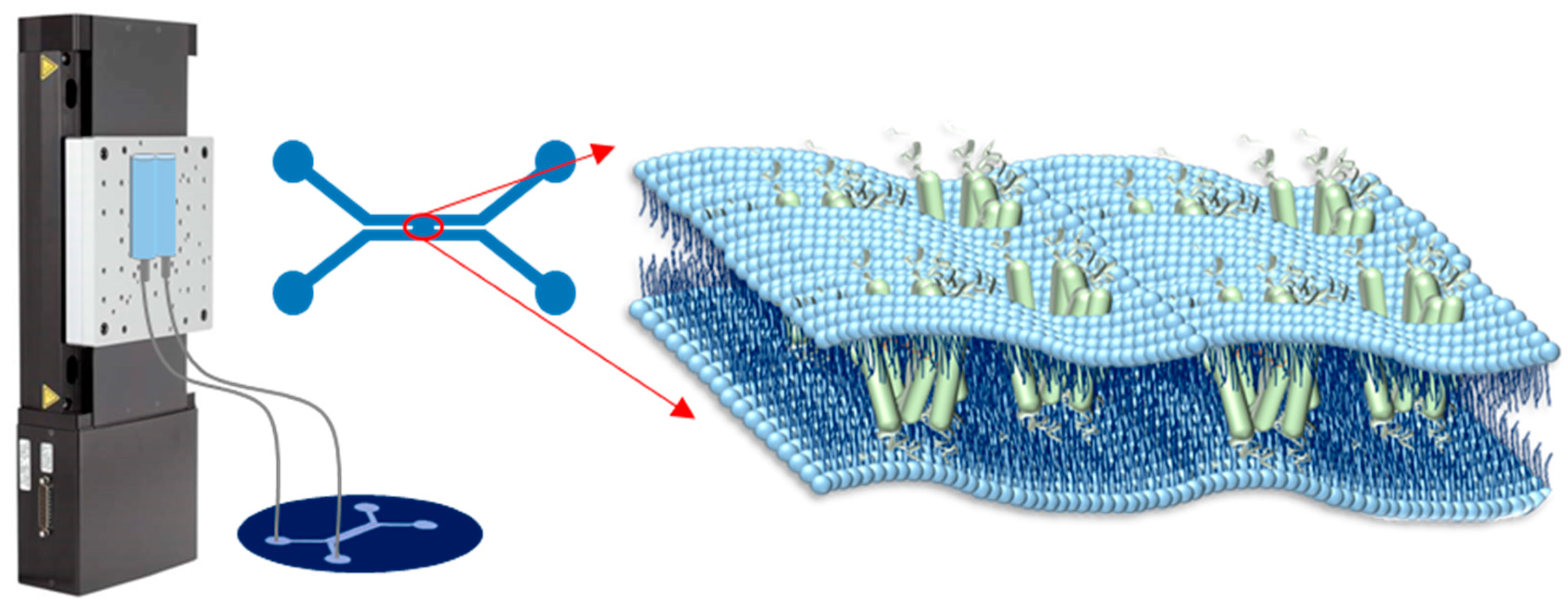
4. Materials and Methods
4.1. Gene Expression
4.2. Lipid Preparation, and Device and Bilayer Fabrication
4.3. Microscope Setup and Electrical Measurements
Author Contributions
Funding
Conflicts of Interest
References
- Lefkowitz, R.J. Seven transmembrane receptors: Something old, something new. Acta Physiol. 2007, 190, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Gudermann, T.; Nürnberg, B.; Schultz, G. Receptors and G proteins as primary components of transmembrane signal transduction-Part 1. G-protein-coupled receptors: Structure and function. J. Mol. Med. 1995, 73, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Orwick-Rydmark, M.; Lovett, J.E.; Graziadei, A.; Lindholm, L.; Hicks, M.R.; Watts, A. Detergent-free incorporation of a seven-transmembrane receptor protein into nanosized bilayer lipodisq particles for functional and biophysical studies. Nano Lett. 2012, 12, 4687–4692. [Google Scholar] [CrossRef]
- Zhulin, I.B.; Nikolskaya, A.N.; Galperin, M.Y. Common extracellular sensory domains in transmembrane receptors for diverse signal transduction pathways in Bacteria and Archaea. J. Bacteriol. 2013, 185, 285–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stock, J.B.; Stock, A.M.; Mottonen, J.M. Signal transduction in bacteria. Nature 1990, 344, 395–400. [Google Scholar] [CrossRef]
- Árnadóttir, J.; Chalfie, M. Eukaryotic mechanosensitive channels. Annu. Rev. Biophys. 2010, 39, 111–137. [Google Scholar] [CrossRef]
- Nagel, G.; Szellas, T.; Huhn, W.; Kateriya, S.; Adeishvili, N.; Berthold, P.; Ollig, D.; Hegemann, P.; Bamberg, E. Channelrhodopsin-2, a directly light-gated Cation-Selective Membrane Channel. Proc. Natl. Acad. Sci. USA 2003, 100, 13940–13945. [Google Scholar] [CrossRef] [Green Version]
- Sandoval, P.J.; Santiago, J. In Vitro Analytical Approaches to Study Plant Ligand-Receptor Interactions. Plant Physiol. 2020, 182, 1697–1712. [Google Scholar] [CrossRef] [Green Version]
- Stora, T.; Lakey, J.H.; Vogel, H. Ion-channel gating in transmembrane receptor proteins: Functional activity in tethered lipid membranes. Angew. Chem. Int. Ed. 1999, 38, 389–392. [Google Scholar] [CrossRef]
- Simon, M.I.; Strathmann, M.P.; Gautam, N. Diversity of G proteins in Signal Transduction. Science 1991, 252, 802–808. [Google Scholar] [CrossRef]
- Pierce, K.L.; Premont, R.T.; Lefkowitz, R.J. Seven-transmembrane receptors. Nat. Rev. Mol. Cell Biol. 2002, 3, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Bethani, I.; Skånland, S.S.; Dikic, I.; Acker-Palmer, A. Spatial organization of transmembrane receptor signalling. EMBO J. 2010, 9, 2677–2688. [Google Scholar] [CrossRef] [Green Version]
- Haswell, E.S.; Phillips, R.; Rees, D.C. Mechanosensitive channels: What can they do and how do they do it? Structure 2011, 19, 1356–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukohata, Y.; Sugiyama, Y.; Ihara, K.; Yoshida, M. An Australian Halobacterium contains a novel Proton Pump Retinal Protein: Archaerhodopsin. Biochem. Biophys. Res. Commun. 1988, 151, 1339–1345. [Google Scholar] [CrossRef]
- Ganapathy, S.; Kratz, S.; Chen, Q.; Hellingwerf, K.J.; de Groot, H.J.; Rothschild, K.J.; de Grip, W.J. Redshifted and Near-infrared Active Analog Pigments Based upon Archaerhodopsin-3. Photochem. Photobiol. 2019, 95, 959–968. [Google Scholar] [CrossRef]
- Maclaurin, D.; Venkatachalam, V.; Lee, H.; Cohen, A.E. Mechanism of voltage-sensitive fluorescence in a microbial rhodopsin. Proc. Natl. Acad. Sci. USA 2013, 110, 5939–5944. [Google Scholar] [CrossRef] [Green Version]
- Flytzanis, N.C.; Bedbrook, C.N.; Chiu, H.; Engqvist, M.K.; Xiao, C.; Chan, K.Y.; Sternberg, P.W.; Arnold, F.H.; Gradinaru, V. Archaerhodopsin variants with enhanced voltage-sensitive fluorescence in mammalian and Caenorhabditis elegans neurons. Nat. Commun. 2014, 5, 4894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juarez, J.F.B.; Judge, P.J.; Adam, S.; Axford, D.; Vinals, J.; Birch, J.; Kwan, T.O.; Hoi, K.K.; Yen, H.-Y.; Vial, A. Structures of the archaerhodopsin-3 transporter reveal that disordering of internal water networks underpins receptor sensitization. Nat. Commun. 2021, 12, 629. [Google Scholar] [CrossRef]
- El-Gaby, M.; Zhang, Y.; Wolf, K.; Schwiening, C.J.; Paulsen, O.; Shipton, O.A. Archaerhodopsin Selectively and Reversibly Silences Synaptic Transmission through Altered pH. Cell Rep. 2016, 16, 2259–2268. [Google Scholar] [CrossRef] [Green Version]
- Mukohata, Y.; Sugiyama, Y.; Ihara, K. Photophosphorylation Elements in Halobacteria: An A-type ATP Synthase and Bacterial Rhodopsins. J. Bioenerg. Biomembr. 1992, 24, 547–553. [Google Scholar] [CrossRef]
- Cao, Z.; Ding, X.; Peng, B.; Zhao, Y.; Ding, J.; Watts, A.; Zhao, X. Novel expression and characterization of a light driven proton pump archaerhodopsin 4 in a Halobacterium salinarum strain. Biochim. Biophys. Acta 2015, 1847, 390–398. [Google Scholar] [CrossRef] [Green Version]
- Deisseroth, K.; Hegemann, P. The form and function of channelrhodopsin. Science 2017, 357, eaan5544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Y.; Li, J.Z.; Schnitzer, M.J. Enhanced Archaerhodopsin Fluorescent Protein Voltage Indicators. PLoS ONE 2013, 8, e66959. [Google Scholar] [CrossRef] [Green Version]
- Khangholi, N.; Seemann, R.; Fleury, J.-B. Simultaneous measurement of surface and bilayer tension in a microfluidic chip. Biomicrofluidics 2020, 14, 024117. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Noireaux, V. An E. coli Cell-Free Expression Toolbox: Application to Synthetic Gene Circuits and Artificial Cells. ACS Synth. Biol. 2012, 1, 29–41. [Google Scholar] [CrossRef]
- Finkler, M.; Ott, A. Bead-based assay for spatiotemporal gene expression control in cell-free transcription–translation systems. Biotechniques 2019, 66, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Finkler, M.; Kurt, Ö.; Grimm, F.; Hartz, P.; Ott, A. A bead-based method for the removal of the amino acid lysine from cell-free transcription-translation systems. J. Biotechnol. 2020, 324S, 100024. [Google Scholar] [CrossRef]
- Finkler, M.; Ravanbodshirazi, S.; Grimm, F.; Hartz, P.; Ott, A. Full incorporation of the noncanonical amino acid hydroxylysine as a surrogate for lysine in green fluorescent protein. Bioorg. Med. Chem. 2021, 41, 116207. [Google Scholar] [CrossRef]
- Sunami, T.; Hosoda, K.; Suzuki, H.; Matsuura, T.; Yomo, T. Cellular compartment model for exploring the effect of the lipidic membrane on the kinetics of encapsulated biochemical reactions. Langmuir 2010, 26, 8544–8551. [Google Scholar] [CrossRef]
- Izri, Z.; Garenne, D.; Noireaux, V.; Maeda, Y.T. Gene Expression in on-Chip Membrane-Bound Artificial Cells. ACS Synth. Biol. 2019, 8, 1705–1712. [Google Scholar] [CrossRef]
- Nevenzal, H.; Noach-Hirsh, M.; Skornik-Bustan, O.; Brio, L.; Barbiro-Michaely, E.; Glick, Y.; Avrahami, D.; Lahmi, R.; Tzur, A.; Gerber, D. A high-throughput integrated microfluidics method enables tyrosine autophosphorylation discovery. Commun. Biol. 2019, 2, 42. [Google Scholar] [CrossRef]
- Silverman, A.D.; Karim, A.S.; Jewett, M.C. Cell-free gene expression: An expanded repertoire of applications. Nat. Rev. Genet. 2020, 21, 151–170. [Google Scholar] [CrossRef] [PubMed]
- Katzen, F.; Chang, G.; Kudlicki, W. The past, present and future of cell-free protein synthesis. Trends Biotechnol. 2005, 23, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. J. Biol. Eng. 2010, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- McSweeney, M.A.; Styczynski, M.P. Effective Use of Linear DNA in Cell-Free Expression Systems. Front. Bioeng. Biotechnol. 2021, 9, 715328. [Google Scholar] [CrossRef] [PubMed]
- Senoussi, A.; Lee Tin Wah, J.; Shimizu, Y.; Robert, J.; Jaramillo, A.; Findeiss, S.; Axmann, I.M.; Estevez-Torres, A. Quantitative Characterization of Translational Riboregulators Using an in Vitro Transcription-Translation System. ACS Synth. Biol. 2018, 7, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Damiati, S.; Mhanna, R.; Kodzius, R.; Ehmoser, E.-K. Cell-Free Approaches in Synthetic Biology Utilizing Microfluidics. Genes 2018, 9, 144. [Google Scholar] [CrossRef] [Green Version]
- Koch, M.; Dencher, N.; Oesterhelt, D.; Plöhn, H.; Rapp, G.; Büldt, G. Time-resolved X-ray diffraction study of structural changes associated with the photocycle of bacteriorhodopsin. EMBO J. 1991, 10, 521–526. [Google Scholar] [CrossRef] [Green Version]
- Nakasako, M.; Kataoka, M.; Amemiya, Y.; Tokunaga, F. Crystallographic characterization by X-ray diffraction of the M-intermediate from the photo-cycle of bacteriorhodopsin at room temperature. FEBS Lett. 1991, 292, 73–75. [Google Scholar]
- Dencher, N.A.; Dresselhaus, D.; Zaccai, G.; Büldt, G. Structural changes in bacteriorhodopsin during proton translocation revealed by neutron diffraction. Proc. Natl. Acad. Sci. USA 1989, 86, 7876–7879. [Google Scholar] [CrossRef] [Green Version]
- Montal, M.; Darszon, A.; Trissl, H.W. Transmembrane channel formation in rhodopsin-containing bilayer membranes. Nature 1977, 267, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Radzwill, N.; Gerwert, K.; Steinhoff, H.J. Time-resolved detection of transient movement of helices F and G in doubly spin-labeled bacteriorhodopsin. Biophys. J. 2001, 80, 2856–2866. [Google Scholar] [CrossRef] [Green Version]
- Heo, P.; Ramakrishnan, S.; Coleman, J.; Rothman, J.E.; Fleury, J.B.; Pincet, F. Highly Reproducible Physiological Asymmetric Membrane with Freely Diffusing Embedded Proteins in a 3D-Printed Microfluidic Setup. Small 2019, 15, 1900725. [Google Scholar] [CrossRef] [PubMed]
- Worst, E.G.; Exner, M.P.; De Simone, A.; Schenkelberger, M.; Noireaux, V.; Budisa, N.; Ott, A. Cell-free expression with the toxic amino acid canavanine. Bioorg. Med. Chem. Lett. 2015, 25, 3658–3660. [Google Scholar] [CrossRef] [PubMed]
- Worst, E.G.; Exner, M.P.; De Simone, A.; Schenkelberger, M.; Noireaux, V.; Budisa, N.; Ott, A. Residue-specific Incorporation of Noncanonical Amino Acids into Model Proteins Using an Escherichia coli Cell-free Transcription-translation System. J. Vis. Exp. 2016, 114, 54273. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khangholi, N.; Finkler, M.; Seemann, R.; Ott, A.; Fleury, J.-B. Photoactivation of Cell-Free Expressed Archaerhodopsin-3 in a Model Cell Membrane. Int. J. Mol. Sci. 2021, 22, 11981. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111981
Khangholi N, Finkler M, Seemann R, Ott A, Fleury J-B. Photoactivation of Cell-Free Expressed Archaerhodopsin-3 in a Model Cell Membrane. International Journal of Molecular Sciences. 2021; 22(21):11981. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111981
Chicago/Turabian StyleKhangholi, Navid, Marc Finkler, Ralf Seemann, Albrecht Ott, and Jean-Baptiste Fleury. 2021. "Photoactivation of Cell-Free Expressed Archaerhodopsin-3 in a Model Cell Membrane" International Journal of Molecular Sciences 22, no. 21: 11981. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111981